
Intra-Annual Xylem Growth of Larix principis-rupprechtii at Its Upper and Lower Distribution Limits on the Luyashan Mountain in North-Central China

Abstract
:1. Introduction
2. Materials and Methods
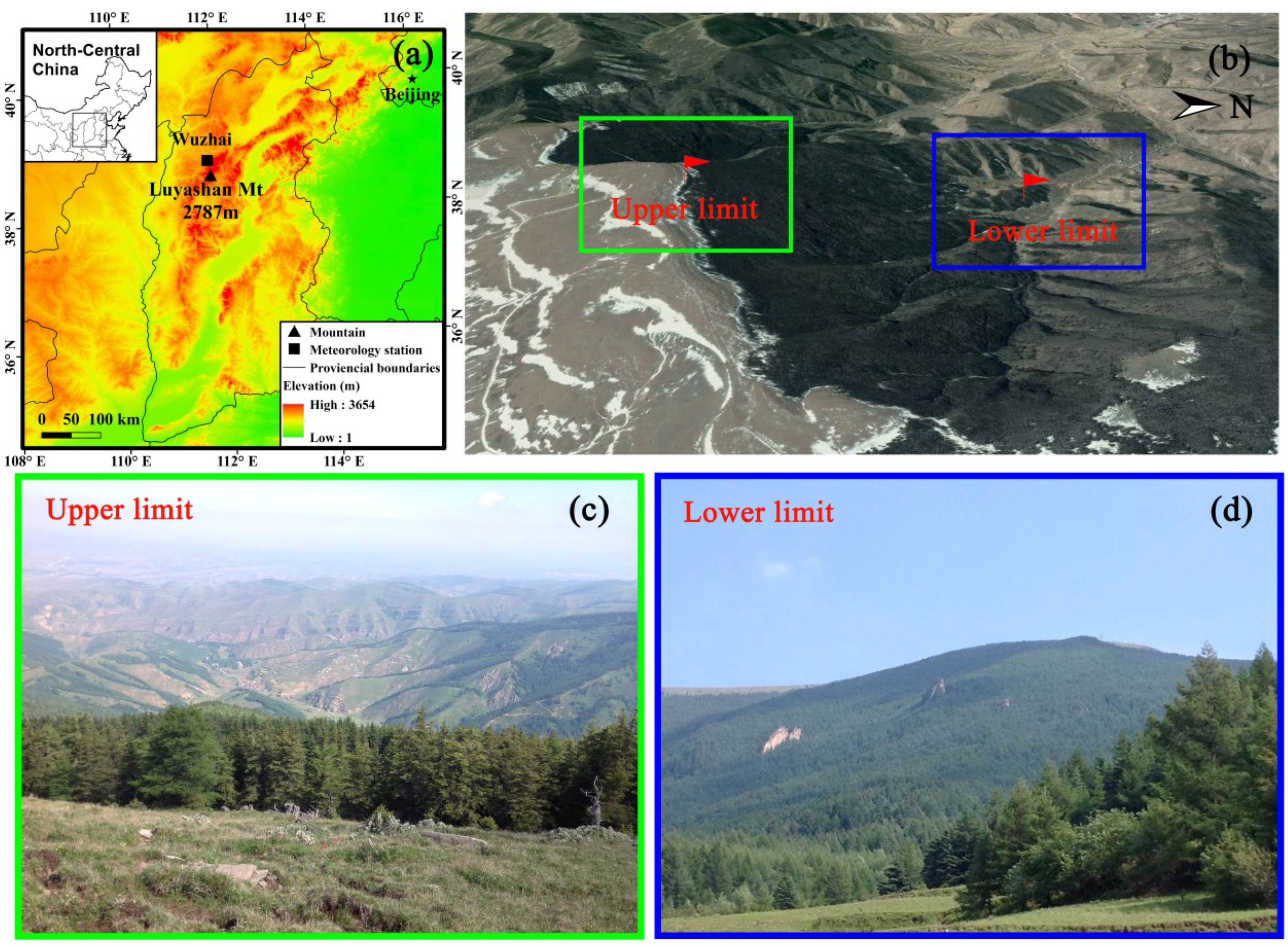
2.1. Study Sites


2.2. Tree Selection
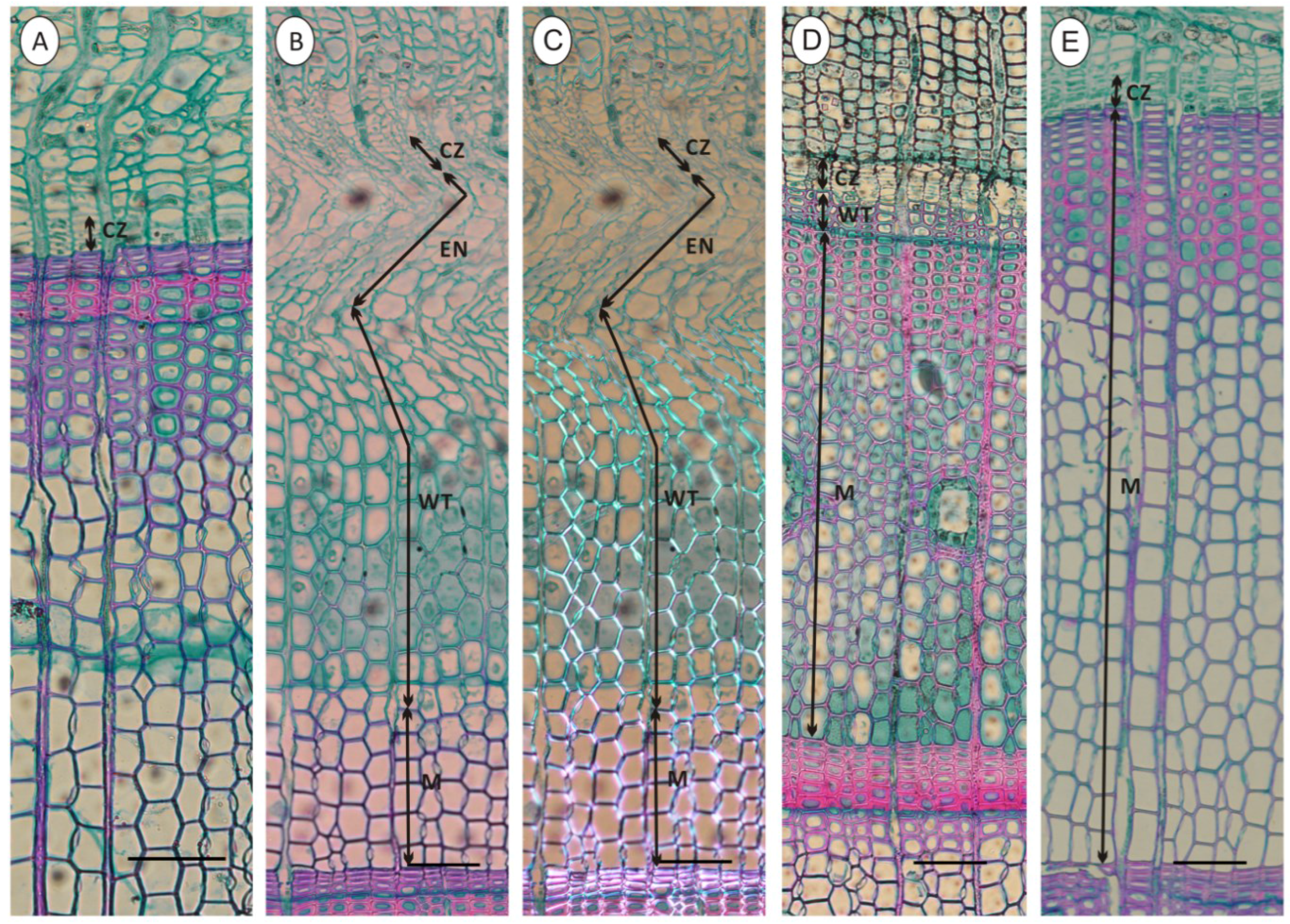
2.3. Cambial Activity and Xylem Differentiation

2.4. Environmental Factors and Climate-Growth Relationship
2.5. Threshold Temperature
3. Results
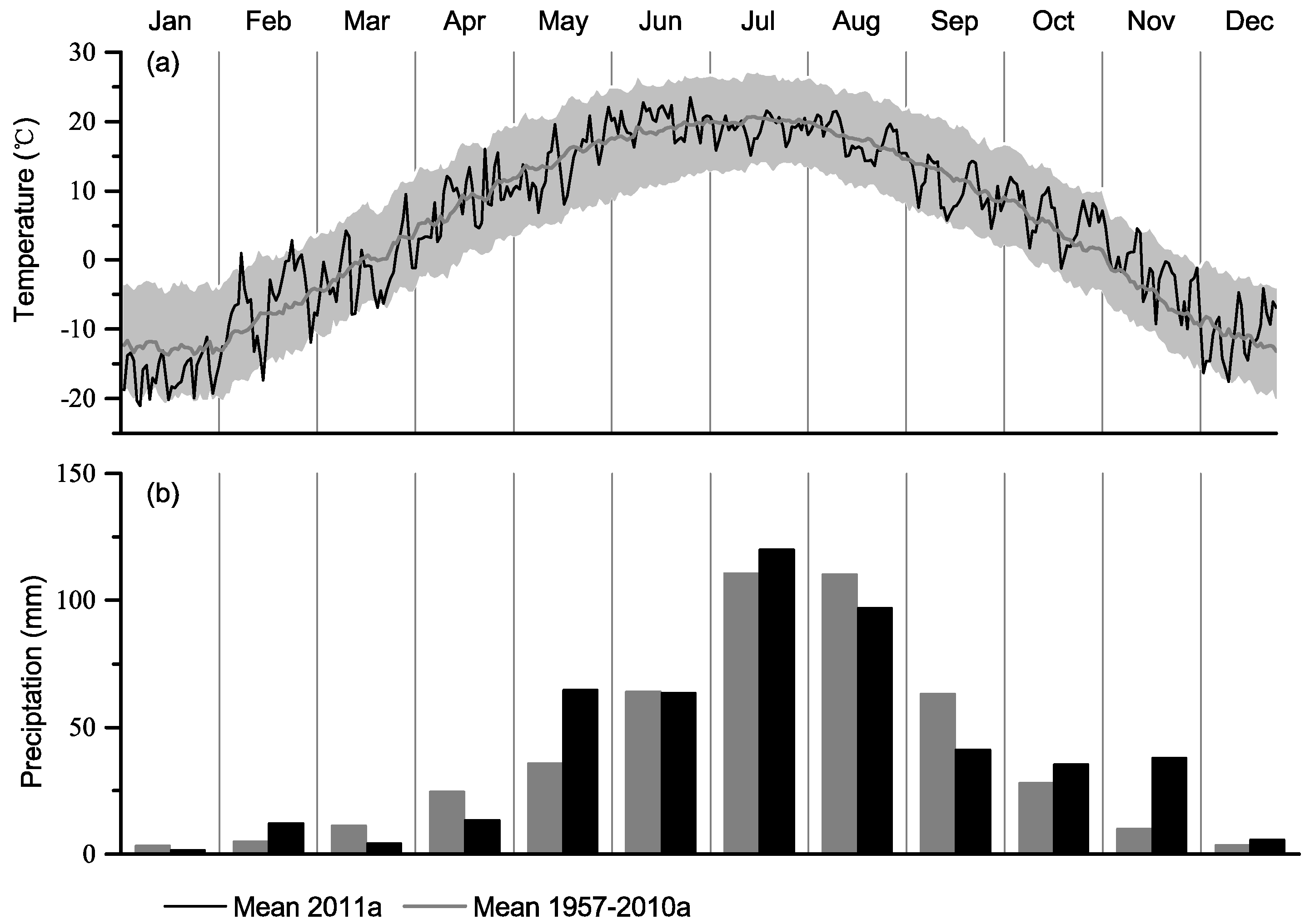
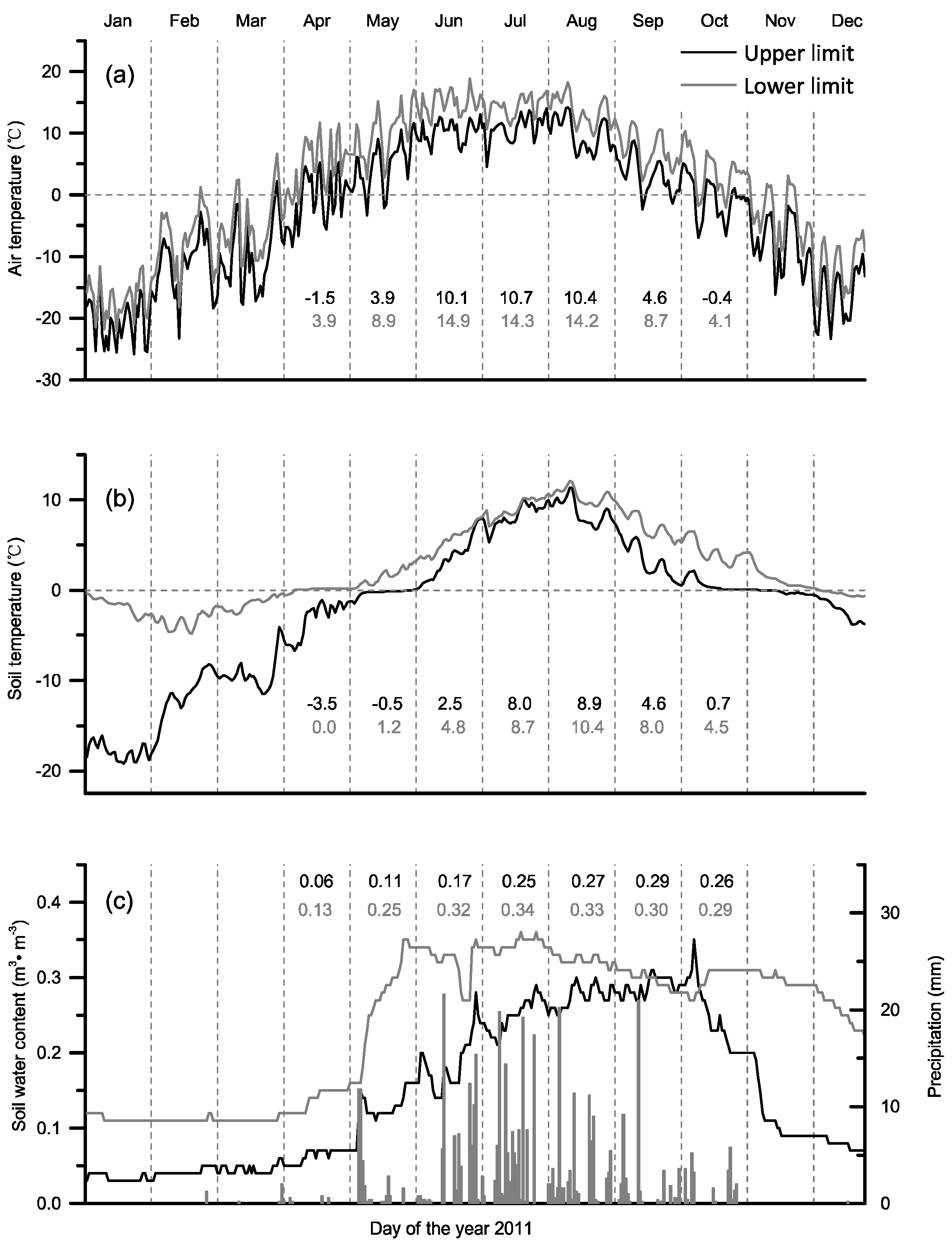
3.1. Temperature and Water Conditions at the Upper and Lower Distribution Limits


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Altitude (m a.s.l.) | Tree Height (m) | DBH (cm) | Age | Ta (°C) | Ts (°C) | SWC (m3·m−3) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| annual | winter | GS | annual | winter | GS | annual | winter | GS | |||||
| Upper distribution limit (Upper) | 2740 | 5.7 ± 1.3 | 17.2 ± 7.3 | 43 ± 3 | −2.1 | −13.5 | 7.9 | −1.7 | −10.9 | 4.7 | 0.15 | 0.05 | 0.22 |
| Lower distribution limit (Lower) | 2040 | 15.3 ± 1.5 | 27.3 ± 5.6 | 34 ± 5 | 2.3 | −9.1 | 12.2 | 2.8 | −0.9 | 6.6 | 0.24 | 0.16 | 0.31 |
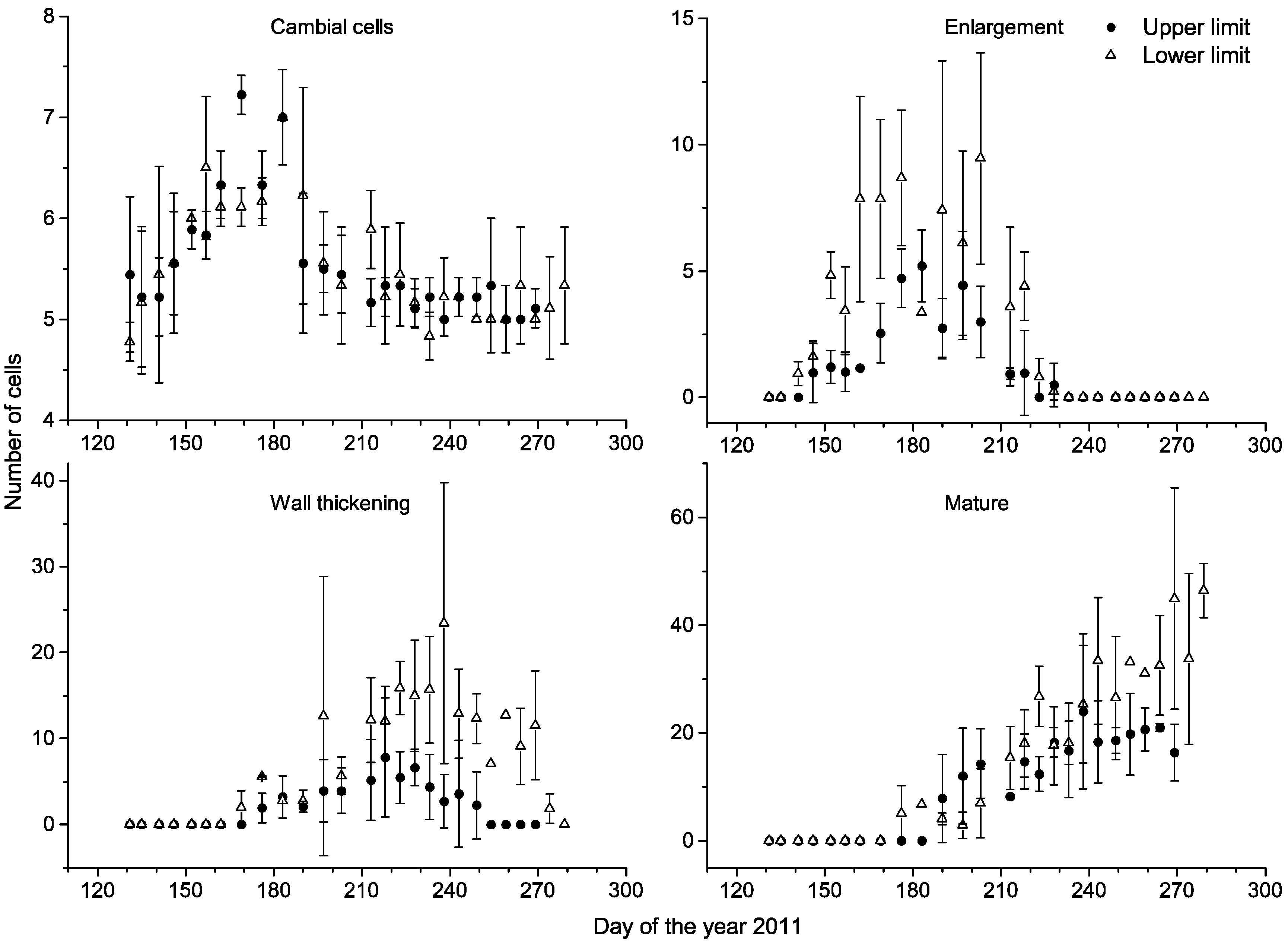
3.2. Cambial Activity and Xylem Differentiation

| Site | Onset | End | Duration | |||||
|---|---|---|---|---|---|---|---|---|
| EN | WT | M | EN | WT | EN | WT | xylogenesis | |
| Upper | 150 ± 3 | 178 ± 4 | 190 ± 0 a | 225 ± 8 | 245 ± 8 a | 75 ± 11 | 67 ± 5 a | 95 ± 12 a |
| Lower | 143 ± 3 | 171 ± 4 | 176 ± 0 b | 225 ± 3 | 277 ± 3 b | 82 ± 0 | 106 ± 4 b | 135 ± 3 b |
| Z | −1.826 | −1.650 | −2.236 | −0.232 | −1.993 | −0.696 | −1.964 | −1.993 |
| P | 0.068 | 0.099 | 0.025 | 0.817 | 0.046 | 0.487 | 0.050 | 0.046 |
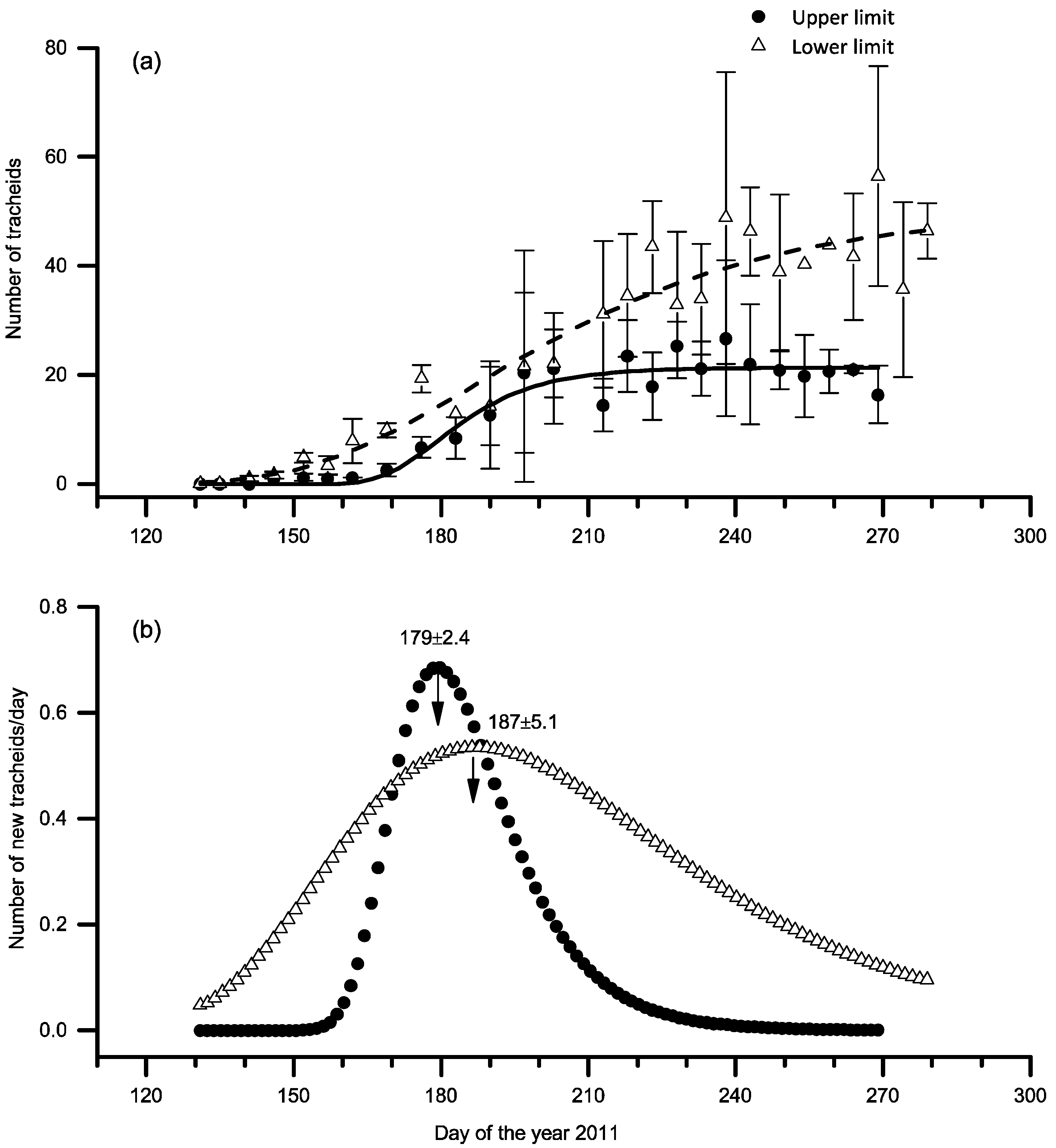
3.3. Xylem Growth Comparison

3.4. Climate-Growth Relationship
| Site | A: Pearson Correlation Coefficients | B: Threshold Temperature | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SWC | P | Ta | Ts | Ta (°C) | Ts (°C) | |||||||||
| min | mean | max | min | mean | max | min | mean | max | min | mean | max | |||
| Upper | 0.546 | 0.374 | 0.685 | 0.660 | 0.631 | 0.951 | 0.945 | 0.935 | 2.8 ± 0.7 a | 6.7 ± 0.5 a | 11.5 ± 0.1 a | −0.07 ± 0.01 | −0.07 ± 0.02 | −0.05 ± 0.04 |
| Lower | 0.497 | 0.492 | 0.831 | 0.805 | 0.736 | 0.924 | 0.924 | 0.921 | 4.5 ± 0.6 b | 9.4 ± 0.8 b | 14.6 ± 0.9 b | 1.23 ± 0.27 | 1.42 ± 0.28 | 1.71 ± 0.29 |
3.5. Threshold Temperature
4. Discussion
4.1. Comparison of Xylem Growth Patterns at the Upper and Lower Distribution Limits
4.2. Role of Soil Temperature in Cambial Activity Onset and Its Threshold Values
4.3. Cessation of Xylem Growth
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Fonti, P.; von Arx, G.; García-González, I.; Eilmann, B.; Sass-Klaassen, U.; Gärtner, H.; Eckstein, D. Studying global change through investigation of the plastic responses of xylem anatomy in tree rings. New Phytol. 2010, 185, 42–53. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.G.; Bergeron, Y.; Zhai, L.H.; Denneler, B. Variation in intra-annual radial growth (xylem formation) of Picea mariana (pinaceae) along a latitudinal gradient in western Quebec, Canada. Am. J. Bot. 2011, 98, 792–800. [Google Scholar] [CrossRef] [PubMed]
- Rossi, S.; Morin, H.; Deslauriers, A.; Plourde, P.-Y. Predicting xylem phenology in black spruce under climate warming. Glob. Chang. Biol. 2011, 17, 614–625. [Google Scholar] [CrossRef]
- Gruber, A.; Zimmermann, J.; Wieser, G.; Oberhuber, W. Effects of climate variables on intra-annual stem radial increment in Pinus cembra (L.) along the alpine treeline ecotone. Ann. For. Sci. 2009, 66, 503. [Google Scholar] [CrossRef]
- Rossi, S.; Deslauriers, A.; Anfodillo, T. Assessment of cambial activity and xylogenesis by microsampling tree species: An example at the Alpine timberline. IAWA J. 2006, 27, 383–394. [Google Scholar] [CrossRef]
- Oberhuber, W.; Gruber, A.; Kofler, W.; Swidrak, I. Radial stem growth in response to microclimate and soil moisture in a drought-prone mixed coniferous forest at an inner Alpine site. Eur. J. For. Res. 2014, 133, 467–479. [Google Scholar] [CrossRef] [PubMed]
- Rossi, S.; Anfodillo, T.; Čufar, K.; Cuny, H.E.; Deslauriers, A.; Fonti, P.; Frank, D.; Gričar, J.; Gruber, A.; King, G.M.; et al. A meta-analysis of cambium phenology and growth: Linear and non-linear patterns in conifers of the northern hemisphere. Ann. Bot. 2013, 112, 1911–1920. [Google Scholar] [CrossRef] [PubMed]
- Begum, S.; Nakaba, S.; Yamagishi, Y.; Oribe, Y.; Funada, R. Regulation of cambial activity in relation to environmental conditions: Understanding the role of temperature in wood formation of trees. Physiol. Plant. 2013, 147, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Begum, S.; Nakaba, S.; Bayramzadeh, V.; Oribe, Y.; Kubo, T.; Funada, R. Temperature responses of cambial reactivation and xylem differentiation in hybrid poplar (Populus sieboldii × P. grandidentata) under natural conditions. Tree Physiol. 2008, 28, 1813–1819. [Google Scholar] [CrossRef] [PubMed]
- Deslauriers, A.; Rossi, S.; Anfodillo, T.; Saracino, A. Cambial phenology, wood formation and temperature thresholds in two contrasting years at high altitude in southern Italy. Tree Physiol. 2008, 28, 863–871. [Google Scholar] [CrossRef] [PubMed]
- Rossi, S.; Deslauriers, A.; Gričar, J.; Seo, J.-W.; Rathgeber, C.B.K.; Anfodillo, T.; Morin, H.; Levanic, T.; Oven, P.; Jalkanen, R. Critical temperatures for xylogenesis in conifers of cold climates. Glob. Ecol. Biogeogr. 2008, 17, 696–707. [Google Scholar] [CrossRef]
- Schmitt, U.; Jalkanen, R.; Eckstein, D. Cambium dynamics of Pinus sylvestris and Betula spp. in the northern boreal forest in Finland. Silva Fenn. 2004, 38, 167–178. [Google Scholar] [CrossRef]
- Rossi, S.; Deslauriers, A.; Anfodillo, T.; Carraro, V. Evidence of threshold temperatures for xylogenesis in conifers at high altitudes. Oecologia 2007, 152, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Swidrak, I.; Gruber, A.; Kofler, W.; Oberhuber, W. Effects of environmental conditions on onset of xylem growth in Pinus sylvestris under drought. Tree Physiol. 2011, 31, 483–493. [Google Scholar] [CrossRef] [PubMed]
- Turcotte, A.; Morin, H.; Krause, C.; Deslauriers, A.; Thibeault-Martel, M. The timing of spring rehydration and its relation with the onset of wood formation in black spruce. Agric. For. Meteorol. 2009, 149, 1403–1409. [Google Scholar] [CrossRef]
- Goodine, G.K.; Lavigne, M.B.; Krasowski, M.J. Springtime resumption of photosynthesis in balsam fir (Abies balsamea). Tree Physiol. 2008, 28, 1069–1076. [Google Scholar] [CrossRef] [PubMed]
- Körner, C. Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystem, 2nd ed.; Springer: Berlin, Germany, 2003. [Google Scholar]
- Körner, C. The use of ‘altitude’ in ecological research. Trends Ecol. Evol. 2007, 22, 569–574. [Google Scholar] [CrossRef] [PubMed]
- Ruosteenoja, K.; Räisänen, J.; Pirinen, P. Projected changes in thermal seasons and the growing season in Finland. Int. J. Climatol. 2011, 31, 1473–1487. [Google Scholar] [CrossRef]
- Oladi, R.; Pourtahmasi, K.; Eckstein, D.; Bräuning, A. Seasonal dynamics of wood formation in Oriental beech (Fagus orientalis Lipsky) along an altitudinal gradient in the Hyrcanian forest, Iran. Trees-Struct. Funct. 2011, 25, 425–433. [Google Scholar] [CrossRef]
- Takahashi, K.; Okuhara, I. Comparison of climatic effects on radial growth of evergreen broad-leaved trees at their northern distribution limit and co-dominating deciduous broad-leaved trees and evergreen conifers. Ecol. Res. 2011, 27, 125–132. [Google Scholar] [CrossRef]
- Moser, L.; Fonti, P.; Büentgen, U.; Esper, J.; Luterbacher, J.; Franzen, J.; Frank, D. Timing and duration of European larch growing season along altitudinal gradients in the Swiss Alps. Tree Physiol. 2010, 30, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Treml, V.; Kašpar, J.; Kuželová, H.; Gryc, V. Differences in intra-annual wood formation in Picea abies across the treeline ecotone, Giant Mountains, Czech Republic. Trees-Struct. Funct. 2015, 29, 515–526. [Google Scholar] [CrossRef]
- Wu, Z. Vegetation of China; Science Press: Beijing, China, 1980. [Google Scholar]
- Wang, G.X.; Liu, X.Q.; Qiao, J. Forest of Shanxi; China Forestry Press: Beijing, China, 1992. [Google Scholar]
- Jiang, Y.; Zhang, W.T.; Wang, M.C.; Kang, M.Y.; Dong, M.Y. Radial growth of two dominant montane conifer tree species in response to climate change in North-Central China. PLoS ONE 2014, 9, e112537. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z. Vegetation of Shanxi; China Science and Technology Press: Beijing, China, 2001. (In Chinese) [Google Scholar]
- National Soil Survey Office. Annals of Soil Species in China; China Agriculture Press: Beijing, China, 1995; Volume 4. (In Chinese)
- Zhai, L.H.; Bergeron, Y.; Huang, J.G.; Berninger, F. Variation in intra-annual wood formation, and foliage and shoot developement of three major Candian boreal tree species. Am. J. Bot. 2012, 99, 827–837. [Google Scholar] [CrossRef] [PubMed]
- Rossi, S.; Deslauriers, A.; Morin, H. Application of the Gompertz equation for the study of xylem cell development. Dendrochronologia 2003, 21, 33–39. [Google Scholar] [CrossRef]
- Li, X.X.; Liang, E.Y.; Gričar, J.; Prislan, P.; Rossi, S.; Čufar, K. Age dependence of xylogenesis and its climatic sensitivity in Smith fir on the south-eastern Tibetan Plateau. Tree Physiol. 2013, 33, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Rathgeber, C.B.K.; Rossi, S.; Bontemps, J.-D. Cambial activity related to tree size in a mature silver-fir plantation. Ann. Bot. 2011, 108, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Jyske, T.; Mäkinen, H.; Kalliokoski, T.; Nöjd, P. Intra-annual tracheid production of Norway spruce and Scots pine across a latitudinal gradient in Finland. Agric. For. Meteorol. 2014, 194, 241–254. [Google Scholar] [CrossRef]
- Thibeault-Martel, M.; Krause, C.; Morin, H.; Rossi, S. Cambial activity and intra-annual xylem formation in roots and stems of Abies balsamea and Picea mariana. Ann. Bot. 2008, 102, 667–674. [Google Scholar] [CrossRef] [PubMed]
- Deslauriers, A.; Morin, H.; Begin, Y. Cellular phenology of annual ring formation of Abies balsamea in the Quebec boreal forest (Canada). Can. J. For. Res. 2003, 33, 190–200. [Google Scholar] [CrossRef]
- Cuny, H.E.; Rathgeber, C.B.; Lebourgeois, F.; Fortin, M.; Fournier, M. Life strategies in intra-annual dynamics of wood formation: Example of three conifer species in a temperate forest in north-east France. Tree Physiol. 2012, 32, 612–625. [Google Scholar] [CrossRef] [PubMed]
- Jarvis, P.; Linder, S. Constraints to growth of boreal forests. Nature 2000, 405, 904–905. [Google Scholar] [CrossRef] [PubMed]
- Rossi, S.; Girard, M.-J.; Morin, H. Lengthening of the duration of xylogenesis engenders disproportionate increases in xylem production. Glob. Chang. Biol. 2014, 20, 2261–2271. [Google Scholar] [CrossRef] [PubMed]
- Devine, W.D.; Harrington, T.B. Aboveground growth interactions of paired conifer seedlings in close proximity. New For. 2011, 41, 163–178. [Google Scholar] [CrossRef]
- Prislan, P.; Gričar, J.; de Luis, M.; Smith, K.T.; Čufar, K. Phenological variation in xylem and phloem formation in Fagus sylvatica from two contrasting sites. Agric. For. Meteorol. 2013, 180, 142–151. [Google Scholar] [CrossRef]
- Dufour, B.; Morin, H. Climatic control of tracheid production of black spruce in dense mesic stands of eastern Canada. Tree Physiol. 2013, 33, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Gruber, A.; Baumgartner, D.; Zimmermann, J.; Oberhuber, W. Temporal dynamic of wood formation in Pinus cembra along the alpine treeline ecotone and the effect of climate variables. Trees-Struct. Funct. 2009, 23, 623–635. [Google Scholar] [CrossRef]
- Dong, M.Y.; Jiang, Y.; Zhang, W.T.; Yang, Y.G.; Yang, H.C. Effect of alpine treeline conditions on the response of the stem radial variation of Picea meyeri Rebd. et Wils to environmental factors. Pol. J. Ecol. 2011, 59, 729–739. [Google Scholar]
- Kirdyanov, A.; Hughes, M.; Vaganov, E.; Schweingruber, F.; Silkin, P. The importance of early summer temperature and date of snow melt for tree growth in the Siberian Subarctic. Trees-Struct. Funct. 2003, 17, 61–69. [Google Scholar] [CrossRef]
- Alvarez-Uria, P.; Körner, C. Low temperature limits of root growth in deciduous and evergreen temperate tree species. Funct. Ecol. 2007, 21, 211–218. [Google Scholar] [CrossRef]
- Körner, C. Carbon limitation in trees. J. Ecol. 2003, 91, 4–17. [Google Scholar] [CrossRef]
- Schenker, G.; Lenz, A.; Körner, C.; Hoch, G. Physiological minimum temperatures for root growth in seven common European broad-leaved tree species. Tree Physiol. 2014, 34, 302–313. [Google Scholar] [CrossRef] [PubMed]
- Bergh, J.; Linder, S. Effects of soil warming during spring on photosynthetic recovery in boreal Norway spruce stands. Glob. Chang. Biol. 1999, 5, 245–253. [Google Scholar] [CrossRef]
- Leng, W.F.; He, H.S.; Liu, H.J. Response of larch species to climate changes. J. Plant Ecol. 2008, 1, 203–205. [Google Scholar] [CrossRef]
- Wang, Z.; Yang, B.; Deslauriers, A.; Bräuning, A. Intra-annual stem radial increment response of Qilian juniper to temperature and precipitation along an altitudinal gradient in northwestern China. Trees-Struct. Funct. 2015, 29, 25–34. [Google Scholar] [CrossRef]
- Dufour, B.; Morin, H. Tracheid production phenology of Picea mariana and its relationship with climatic fluctuations and bud development using multivariate analysis. Tree Physiol. 2010, 30, 853–865. [Google Scholar] [CrossRef] [PubMed]
- Duchesne, L.; Houle, D.; D’Orangeville, L. Influence of climate on seasonal patterns of stem increment of balsam fir in a boreal forest of Quebec, Canada. Agric. For. Meteorol. 2012, 162, 108–114. [Google Scholar] [CrossRef]
- Gričar, J.; Čufar, K.; Oven, P.; Schmitt, U. Differentiation of terminal latewood tracheids in silver fir trees during autumn. Ann. Bot. 2005, 95, 959–965. [Google Scholar] [CrossRef] [PubMed]
- Lupi, C.; Morin, H.; Deslauriers, A.; Rossi, S. Xylem phenology and wood production: Resolving the chicken-or-egg dilemma. Plant Cell Environ. 2010, 33, 1721–1730. [Google Scholar] [CrossRef] [PubMed]
- Rossi, S.; Morin, H.; Deslauriers, A. Causes and correlations in cambium phenology: Towards an integrated framework of xylogenesis. J. Exp. Bot. 2012, 63, 2117–2126. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, Y.; Zhang, Y.; Guo, Y.; Kang, M.; Wang, M.; Wang, B. Intra-Annual Xylem Growth of Larix principis-rupprechtii at Its Upper and Lower Distribution Limits on the Luyashan Mountain in North-Central China. Forests 2015, 6, 3809-3827. https://doi.org/10.3390/f6113809
Jiang Y, Zhang Y, Guo Y, Kang M, Wang M, Wang B. Intra-Annual Xylem Growth of Larix principis-rupprechtii at Its Upper and Lower Distribution Limits on the Luyashan Mountain in North-Central China. Forests. 2015; 6(11):3809-3827. https://doi.org/10.3390/f6113809
Chicago/Turabian StyleJiang, Yuan, Yiping Zhang, Yuanyuan Guo, Muyi Kang, Mingchang Wang, and Biao Wang. 2015. "Intra-Annual Xylem Growth of Larix principis-rupprechtii at Its Upper and Lower Distribution Limits on the Luyashan Mountain in North-Central China" Forests 6, no. 11: 3809-3827. https://doi.org/10.3390/f6113809
APA StyleJiang, Y., Zhang, Y., Guo, Y., Kang, M., Wang, M., & Wang, B. (2015). Intra-Annual Xylem Growth of Larix principis-rupprechtii at Its Upper and Lower Distribution Limits on the Luyashan Mountain in North-Central China. Forests, 6(11), 3809-3827. https://doi.org/10.3390/f6113809