1. Introduction
The 5th Assessment of the Intergovernmental Panel on Climate Change, Working Group II [
1] identified with high confidence that under climate change there is a “
Risk of loss of terrestrial and inland water ecosystems, biodiversity, and the ecosystem goods, functions, and services they provide for livelihoods”. Their report indicates that adaptation of our management systems is necessary to reduce impacts on society. The economic value of ecosystem services to society is high; for example, Costanza
et al. [
2] estimated an annual value in 2011 of 124.8 × 10
12 2007 US$. Projected changes to forests under a changing climate include increased disturbance by fire, wind, pests and disease, and changes in survival and growth of species [
3,
4,
5]. However, uncertainty is high as to where and when problems will develop, the degree of mortality and the magnitude of gains or losses in growth and, therefore, the socio-economic consequences. Parts of the western North America have already experienced biophysical, economic and social impacts as a result of a mountain pine beetle (
Dendroctonus ponderosae Hopkins) epidemic [
5] triggered mainly by milder winters as the climate warms [
6].
If we expect a continuous flow of ecological goods and services in the future, then strategies to adapt to climate change are an important consideration for current forest management [
1,
7,
8]. One framework to manage under this uncertainty is to increase forests resilience to disturbance [
9,
10,
11,
12,
13]. In this study we use the resilience definition: “
the capacity of a system to absorb disturbance and reorganize while undergoing change so as to still retain essentially the same function, structure, identity, and feedbacks” [
14]. According to resilience theory, systems can respond to disturbances by growing along similar pathways as prior to the disturbance, or by reorganizing into a qualitatively different form. Resilience theory can be applied to socio-ecological systems such as forestry, where ecosystems and landscapes affect, and are affected by management practices and institutions [
15]. Rist and Moen [
16] describe the value of resilience theory as offering new ways to integrate social, economic and ecological interactions and complexity and thereby improving management’s ability to meet the challenges of climate change. One of the ways resilience theory does that is by inherently acknowledging change, disturbance, and uncertainty rather than the constancy and stability of previous forestry paradigms.
Quantifiable resilience indicators for forest management are the subject of a number of recent papers. DeRose and Long [
17] provide a thoughtful framework and articulate the challenges of defining measurable indicators that are operationally relevant. They recommend structural and compositional stand and landscape indicators that reflect post-disturbance management goals over a clear time horizon. For example, indicators of resilience to spruce beetle could include surviving large trees (stand) and the potential for future spruce dominance (landscape). Seidl
et al. [
18] propose using the historical range of variability of two different ecosystem properties to “
delineates the past basin of attraction of the system”. Together with social acceptance, the historical range of variability of ecosystem properties can be used to determine if altered disturbance rates are likely to exceed socio-ecological resilience. A third approach was taken by Duveneck and Scheller [
19] with a resilience index for post-wildfire landscapes based on the recovery rate of species composition and aboveground biomass. A fourth approach was presented by Dymond
et al. [
20] where a landscape with greater socio-ecological resilience was defined as having higher species diversity (Shannon Diversity Index), post-disturbance green-tree volume (growing stock), post-disturbance harvest rates, annual net revenue, and net present value (NPV), in addition, more resilient landscapes had more consistent harvest rates and annual net revenue. We chose to follow this last approach because, based on our observations, we think these indicators reflect many aspects of the decision making process of forest planners in North America. The species diversity and volume indicators are similar conceptually to the approach of Duveneck and Scheller [
19]. The harvest rate, net revenue, and NPV criteria in particular represent the socio-economic values of private companies, jobs in forest-dependent communities, and rent to land-owners. Only the species diversity indicator is as prescriptive for silviculturalists as recommended by DeRose and Long [
17].
Biological diversity is a component of ecosystem resilience where it provides functional redundancy in key ecological processes [
21,
22]. Biological diversity can contribute to resilience through a bet-hedging effect (insurance hypothesis): buffering the temporal variability of productivity and increasing productivity over time [
23]. A second aspect of resilience is the degree of impact on forest ecosystems due to expected pests, disease, and drought. Changing the species, structural, and age class composition through management is another way to potentially lower the severity of disturbances thereby increasing resilience (e.g., [
12,
13,
24,
25]). Dymond
et al. [
20] identified the potential benefits of greater tree species diversity in increasing socio-ecological resilience through proactive management of an extensive insect outbreak. Their study identified short-term reductions in annual net revenue as a trade-off for higher diversity, more consistent growing stocks, harvest rates and greater net revenue over time. However, we need to know how successful similar management strategies might be under different forest conditions.
Adapting to climate change by increasing species and structural complexity of forests will be strongly influenced by past management activities that have produced the current state of the forest. Templi
et al. [
26] assessed four management strategies to increase both species and structural diversity and reduce susceptibility to disturbance across two landscapes. The key drivers of the provision of ecosystem goods and services were the current conditions of the forest and the past management activities, the timing of climate change impacts, and the adaptive management strategy.
Other examples of landscape scale forestry studies highlight short term and long term tradeoffs in risk management strategies. Steenberg
et al. [
27] simulated targeting harvesting towards the species expected to be mal-adapted (decreasing productivity) under a changing climate in eastern Canada. That strategy was effective at converting the forest to well-adapted species and maintaining higher stocks of above-ground biomass compared to other management strategies. However, the harvest rate collapsed after the targeted species had been removed. Schou
et al. [
28] used a simulation approach to model the forest response and resulting economic values of management strategies to move away from an even aged monoculture forest. Harvesting the over-mature forest provided a significant cash flow up-front and reduced the variability between the long-term strategies. This made the choice among options less clear but providing for some planning flexibility. Cameron [
12] identified early thinning and planting additional species as options to adapt Sitka spruce stands to a changing climate in Scotland.
Our study builds on the work of Dymond
et al. [
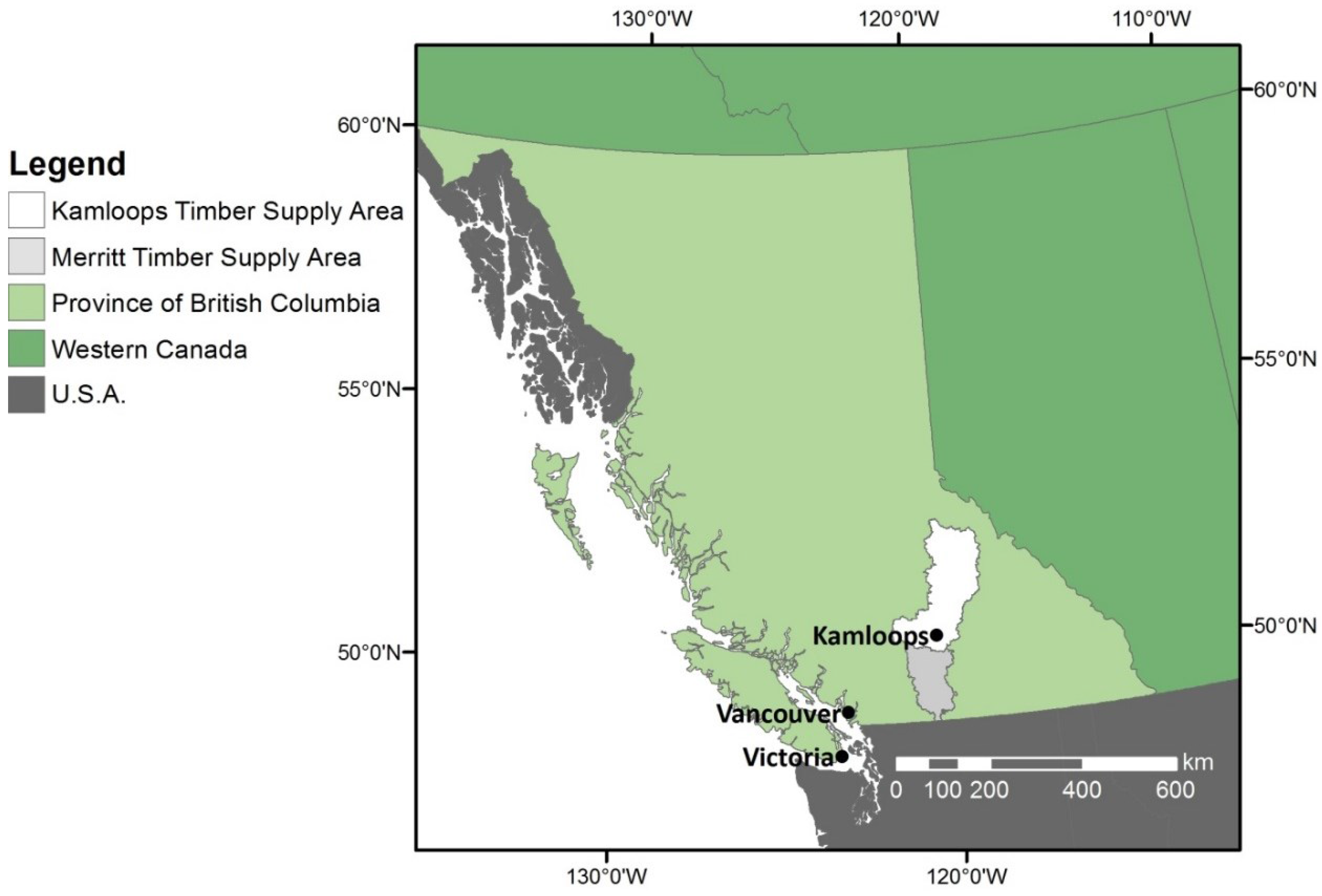
20] noted above. We consider the Kamloops Timber Supply Area (TSA) in southern British Columbia, adjacent to their Merritt TSA study area. The Kamloops TSA has higher diversity of tree and site conditions than Merritt TSA and therefore could have different ecological and economic responses to similar management strategies. Our assessment uses the mountain pine beetle (
Dendroctonus ponderosae) epidemic in a historical retrospective approach to assess two potential management strategies for increasing resilience relative to business as usual. One alternative management strategy increased the species diversity of the tree seedlings being planted. The second alternative strategy decreased the area occupied by a high risk species through targeted harvest and increased species diversity through reforestation. We evaluated socio-ecological resilience using measures of tree species diversity, post-disturbance growing stock, net present value and revenue flow over the management unit as a whole, for the period 1980 to 2060.
4. Discussion
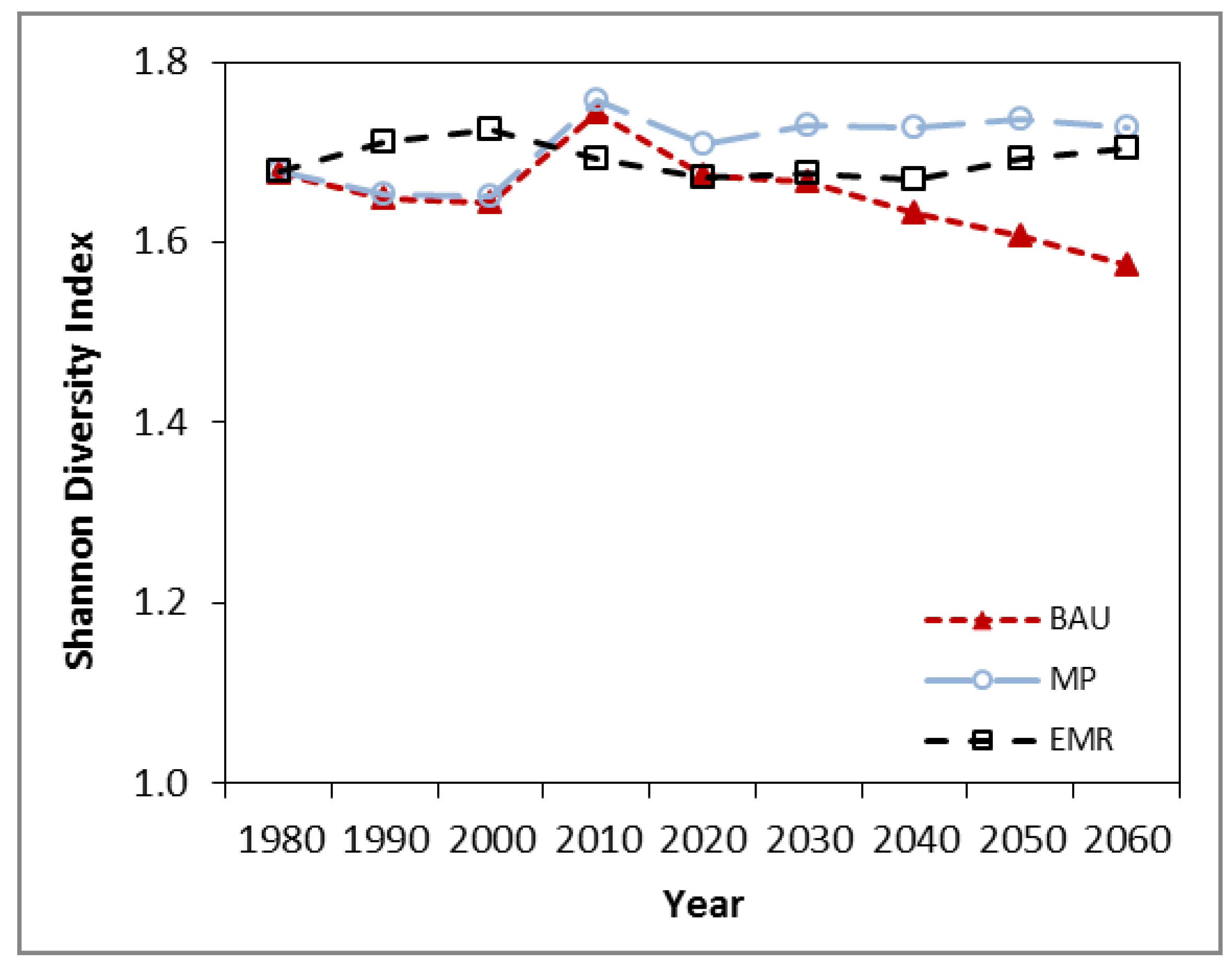
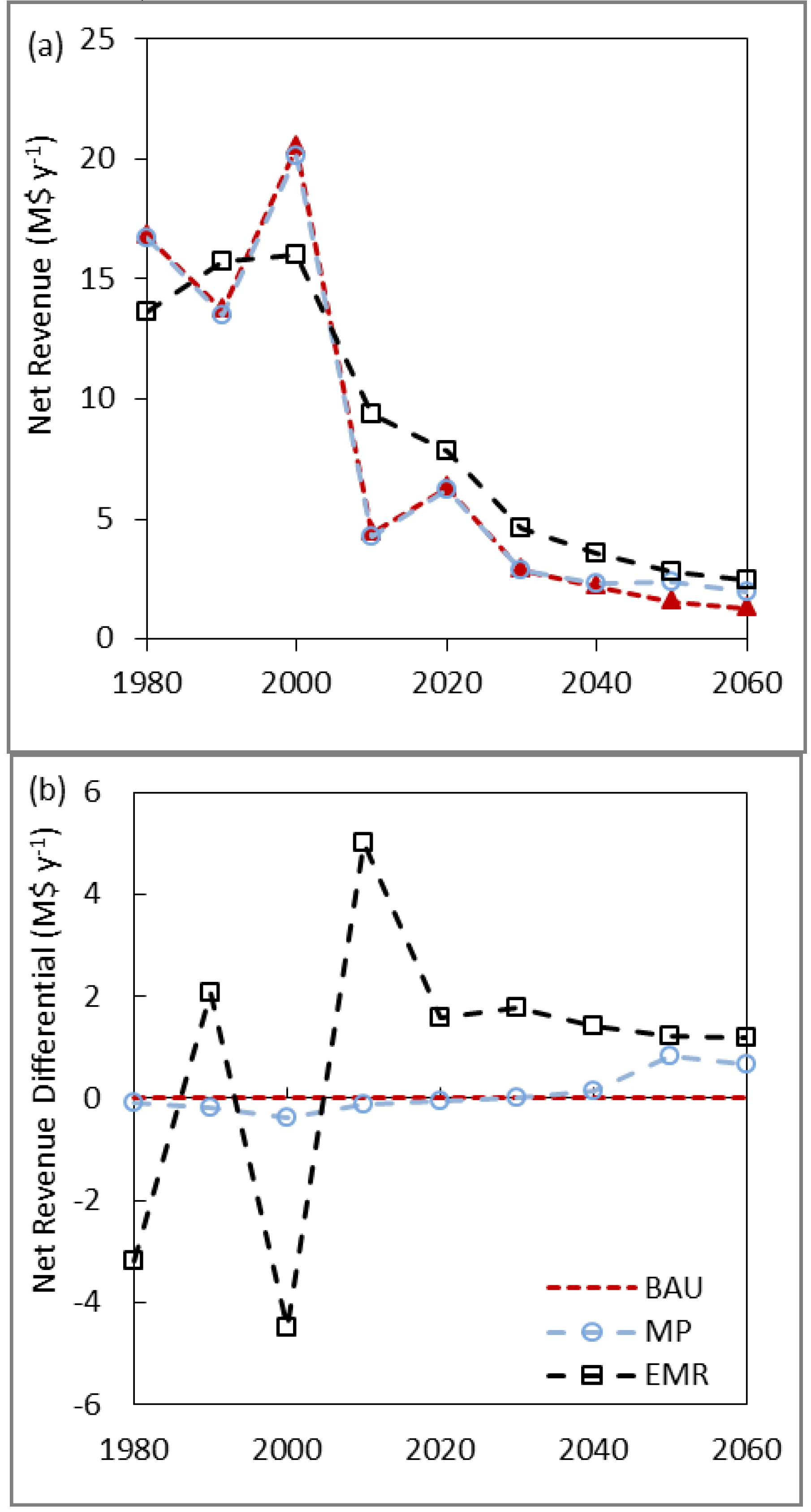
The study results indicated that forest management activities can make a difference to the resilience in the supply of multiple ecological goods and services in the context of natural disturbances. Pro-active management to reduce high risk species while maintaining or increasing diversity reduced the damage of a forest health agent. However, we may be limited in our ability to affect changes on tree species diversity as a mechanism to increase resilience. These limitations may come from the local growing conditions, which may only be able to support a limited number or relative abundance of species, or from policies against introducing non-native species.
Where the effect of climate change on forest pests, diseases or species productivity can be reliably forecast, our study suggests that an adaptive forest management in temperate forests can potentially reduce likelihood of catastrophic ecological and economic instability. The EMR strategy combined the targeted removal of a high risk species with increased partial cutting and increasing the diversity through reforestation (both planted and natural). The combination of these activities was able to reduce the mountain pine beetle impact after only two decades when operating on only 0.5%–0.8% of the forest annually. The strategy also changed landscape-level species mix over the longer term. Greater socio-ecological resilience in the EMR landscape was indicated by having the highest post-disturbance growing stock and highest NPV. The EMR landscape had the highest species diversity for the 1990–2009 decades, but only by a minor amount and was similar to the MP landscape for most of the simulation. Removing the high risk species alone will not necessarily increase resilience. For example, a similar study assessing management options in mitigating the risk of mountain pine beetle found that increasing the harvesting rate and targeting pine did not appreciably change the landscape composition over 20 or 70 years because the stands were replanted with the same species as were harvested [
41].
Differences in species composition, climate, site conditions and productivity will require different adaptation strategies. Therefore, it is useful to compare the results of our analysis with those for the neighbouring Merritt TSA [
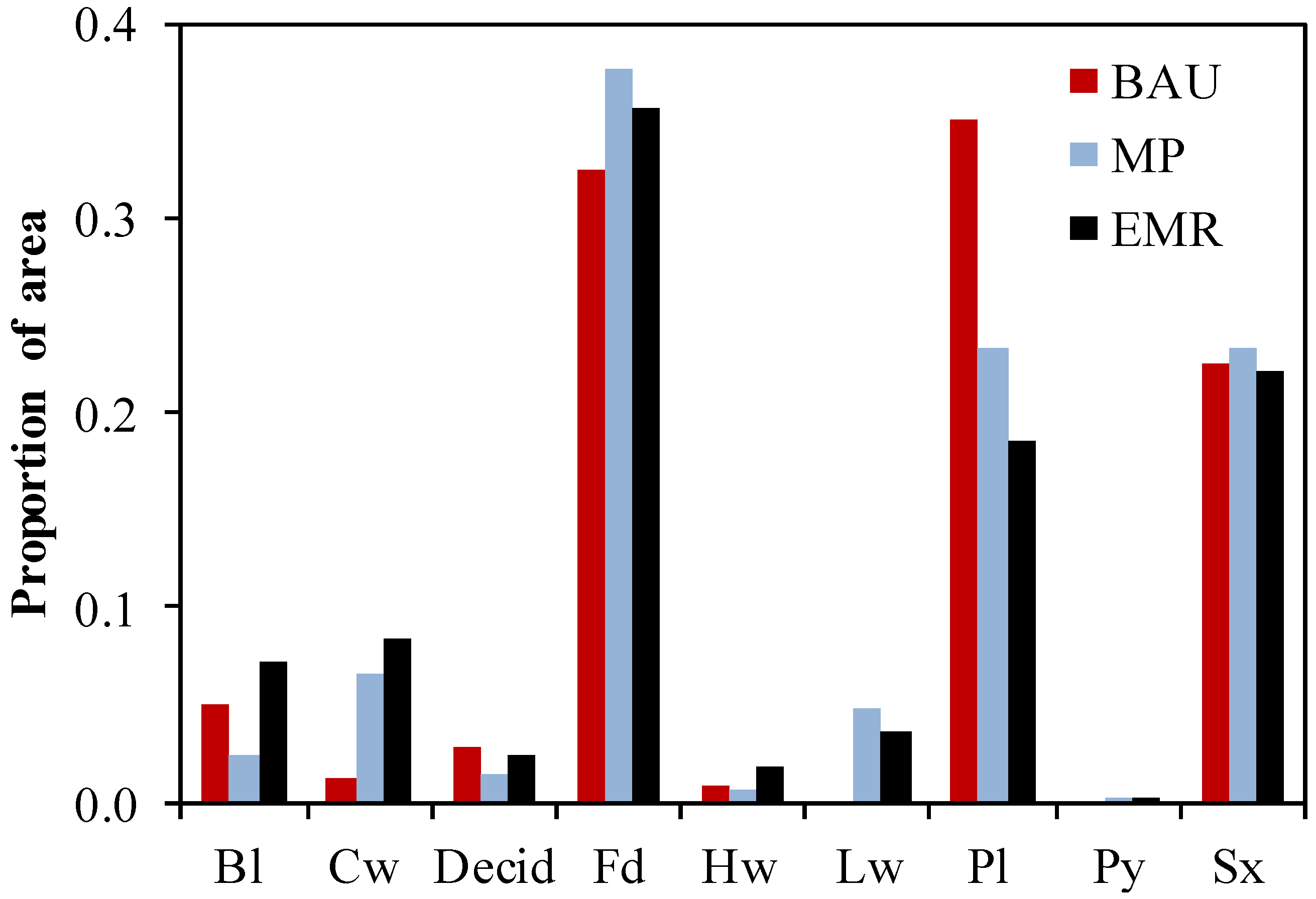
20]. In both studies, the EMR strategies resulted in the greatest ecological resilience (growing stock), highest post-disturbance harvest rates and NPV over the 80 years. Also, the MP and EMR strategies resulted in more old growth by the end of the simulation. The biggest difference between the two studies was that in our study, the EMR landscape did not require a surge in harvest rates in response to the threat from mountain pine beetle. Furthermore, in Merritt TSA management activities substantially reduce the diversity under BAU, and increase it in the MP and EMR strategies, whereas in the current study the diversity indices changed by more modest amounts. Another difference was that the MP strategy in our study was economically viable whereas in the Merritt TSA it was not. These three differences in results are due to differences in the starting conditions of the landscapes, the growing conditions within each landscape, and the BAU approach to silviculture as defined by local forest managers. In the 1980 inventory, the Merritt TSA was overwhelmingly dominated by lodgepole pine and had low tree species diversity compared to our study area. The growing conditions across most of the Merritt TSA landscape are compatible with lodgepole pine and it tends to have higher growth rates than the other tree species. Therefore, BAU forest management has a high proportion of lodgepole pine being planted. The growing conditions in our study area are more variable than in the Merritt TSA, and species other than pine may have similar or higher growth rates. Therefore BAU regeneration in our study area had a better evenness among species being planted than in Merritt TSA. Despite this, both studies show the benefits of strategies designed to increase diversity.
The comparison between Merritt and Kamloops TSA results also provide insight into the more general applicability of these study results to temperate and boreal forest adaptive management. Where diversity is currently low, the management strategies were more successful in increasing that diversity and therefore resilience to disturbance. This result is consistent with modelling in the boreal-temperate forests where there was a positive relationship between resistance and diversity within the low diversity areas [
19].
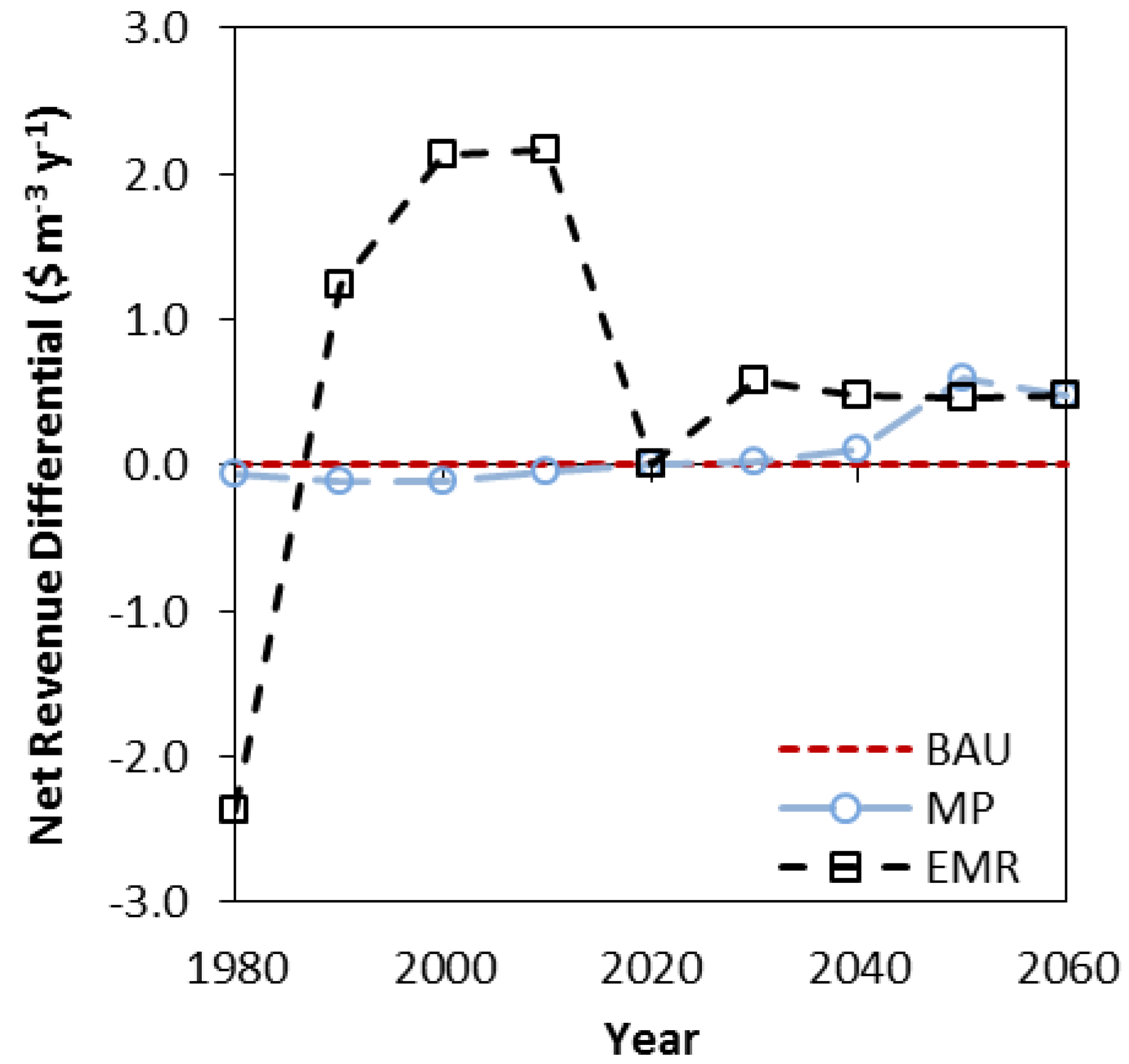
The EMR strategy was clearly the best in terms of NPV over the 80 year simulation and had the lowest per cubic meter costs initially. These results were a surprise since there was no surge in harvesting in 2000–2009, and typically, diversification strategies have poorer economic outcomes [
42]. However, there were near term opportunity costs associated with achieving these longer term outcomes. The EMR strategy produced lower net revenues in the first decade, though there were lower per cubic metre costs as well.
While the ability to reduce costs may be an incentive for forestry companies in North America or Europe to initiate or participate in a program of climate change adaptation, there is a need for them to have a reasonable return on investment in the near term. In contrast, the public who value the state-owned forests as natural capital for both current and future generations may wish to take a much longer term view and assume a lower discount rate [
36]. Consequently, policy makers and stakeholders need to weigh the potential for lost output within a context of higher risk of future infestations or other natural disaster associated with climate change. The sensitivity analysis indicates that the results of our analysis are robust to changes in productivity and log prices.
The economic analysis assumed that log quality and uses are undifferentiated and any change in species mix would not affect the structure of the manufacturing sector, which had a greater level of veneer and plywood production in the 1980s than today in the Kamloops TSA. Any costs associated with shifting production away from veneer into lumber are not factored into this analysis, nor is the market’s ability to absorb a greater volume of lumber or resulting price changes. This change in the supply characteristics could affect the pricing regime, placing less emphasis, thus value, on the logs most suited for veneer or other higher valued products. A change in the price of pine vis-à-vis other species would affect the outcome of our economic analysis, but would still retain the benefit of lower risk.
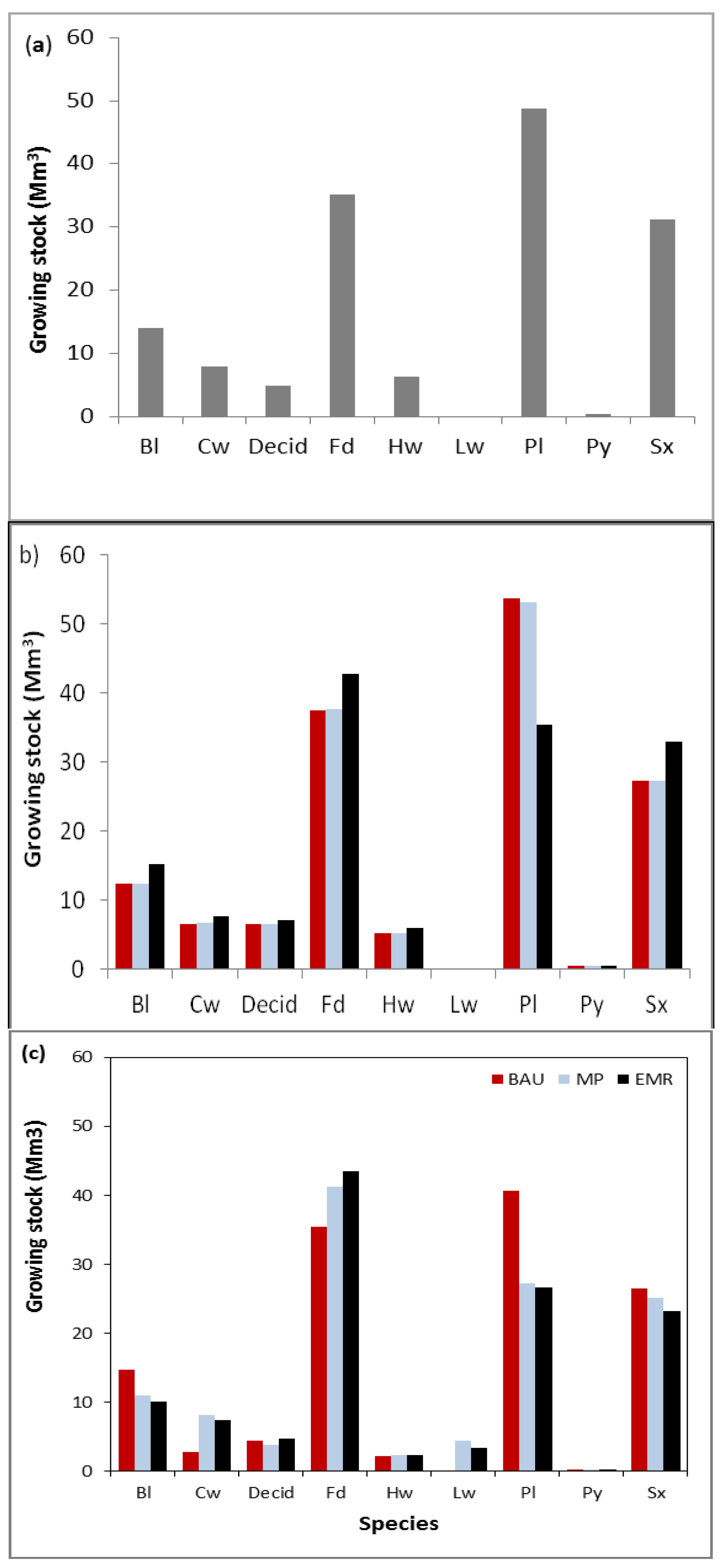
Under the EMR and MP strategies, the future amount of high risk pine decreased, and the amount of Douglas-fir increased (
Figure 3). Although it never reaches the degree of dominance that pine had in 1980, this and the limited increase in diversity led us to consider the potential risks to Douglas-fir from pests and climate change maladaptation.
The forest health agent that historically causes damage to Douglas-fir in B.C. is the western spruce budworm (WSB)
Choristoneura occidentalis [
43]. Unlike mountain pine beetle, which kills trees within a single year, WSB is a defoliator and multiple years or chronic defoliation is generally required to cause mortality. That type of repeated defoliation occurs within some of the hottest and driest sites within the study area (see
Table S1, Interior Douglas Fir Zone) and can cause 4% to 17% mortality of Douglas-fir over 10 years [
44]. Within this climate zone, smaller Douglas-fir trees within multistoried stands are the most susceptible to WSB (7% to 24% mortality over 10 years). These multi-story stands can be created through partial harvesting followed by natural regeneration. In our study, 18% of the area was modelled with natural regeneration of Douglas-fir in the high hazard sites in all three management strategies. Therefore, the alternative management strategies did not increase the amount of multi-story Douglas-fir stands on the landscape. However, the total volume of Douglas-fir on hot, dry sites in 2060 was lower in the BAU landscape (14 Mm
3) compared to the MP and EMR landscapes (15 Mm
3). Recall that the total growing stock also differed between strategies. As a proportion of total volume in 2060, the amount of Douglas-fir on hot, dry sites varied from 11% under BAU to 12.5% under MP and EMR. That difference is likely well within the uncertainty of this projection. Nonetheless, whether these trade-offs in forest health risks are acceptable will depend on the stakeholders.
Climate change also poses a risk to Douglas-fir productivity. Although growth rates may increase on sites that are not moisture limited, the hot, dry sites most susceptible to WSB are also those where Douglas-fir growth is already moisture limited [
33]. Climatic moisture deficits for these areas calculated using the ClimateWNA spatial software [
45] are projected to increase by 5% to 30% by the mid-century in response to warming and decrease in summer precipitation. This will increase drought stress, increase susceptibility to disease and reduce productivity for Douglas-fir, and many other species [
5]. Adaptive strategies to maintain resilience will require the planting of provenances with greater drought tolerance, particularly where alternate species may not be available [
46].
Invasive species and climate change may bring novel pathogens into managed forest ecosystems [
47,
48]. Our study has focused on known threats, but new agents could negate any potential resilience gain from a shift in management. The theoretical way to reduce the risks from unknown pathogens is to increase diversification as described in the insurance hypothesis [
23]. This is one of the reasons why our management strategies were designed to increase diversity, rather than solely removing pine. Future research could incorporate a range of expected and unexpected pathogens in assessing management implications. Furthermore, monitoring of forest health will need to be sensitive to damage from novel pathogens. As our Kamloops TSA results showed, a tailored management strategy can reduce the damage of a forest health agent while increasing ecological goods and services and the stability of timber revenue. However, there are multiple objectives in forest policy in North America including retaining old-growth forests to protect habitat, biodiversity, and aesthetic values. For example, in the Kamloops TSA, old growth forests help support three herds of woodland caribou as well as 51 other species of special concern under the Species at Risk Act [
49]. By maintaining a larger growing stock, our results indicate that pro-active policies intended to increase forest resilience and reduce forest health damage could also support species at risk.
In addition to local socio-economic impacts, changes caused from the beetle outbreak and subsequent forest management response may have a global effect. The tree mortality impact of the beetle and surge in harvesting led to increased carbon emissions and decreased sinks resulting in affected forests becoming net emitters of carbon to the atmosphere [
50]. Reduction in carbon sinks due to a variety of pathogens have been documented around the world (e.g., [
51]). Further impacts of climate change may prolong that condition, delaying the forests returning to being a net sink (e.g., [
52]). However, if natural resource managers implement strategies that increase resilience and reduce the impact of future forest health outbreaks we could help maintain forest carbon stocks and mitigate climate change through adaptation.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





