Does the Slope of the Self-thinning Line Remain a Constant Value across Different Site Qualities?—An Implication for Plantation Density Management


Abstract
:1. Introduction
2. Materials and Methods
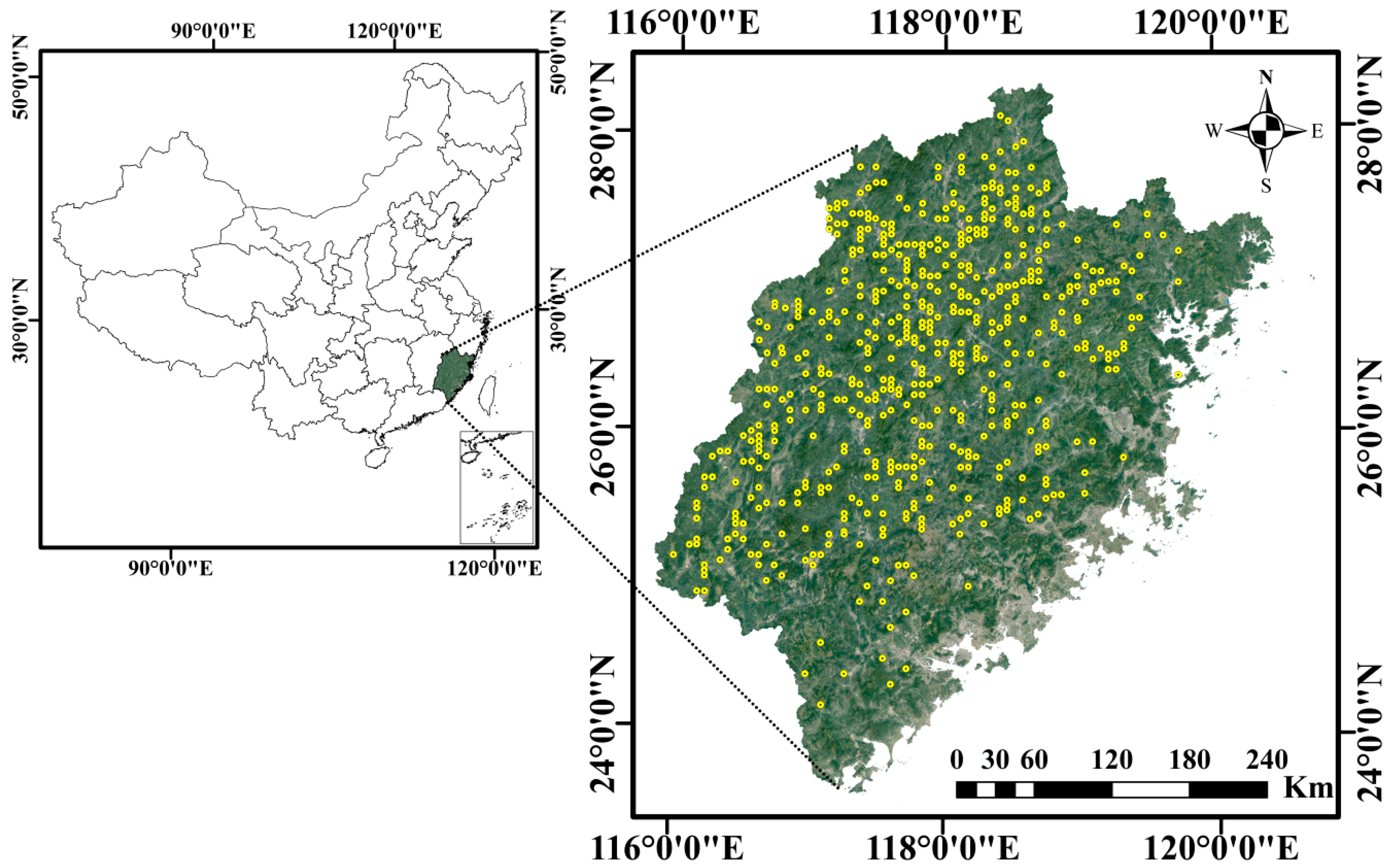
2.1. Data Source
2.2. Site Quality Classification
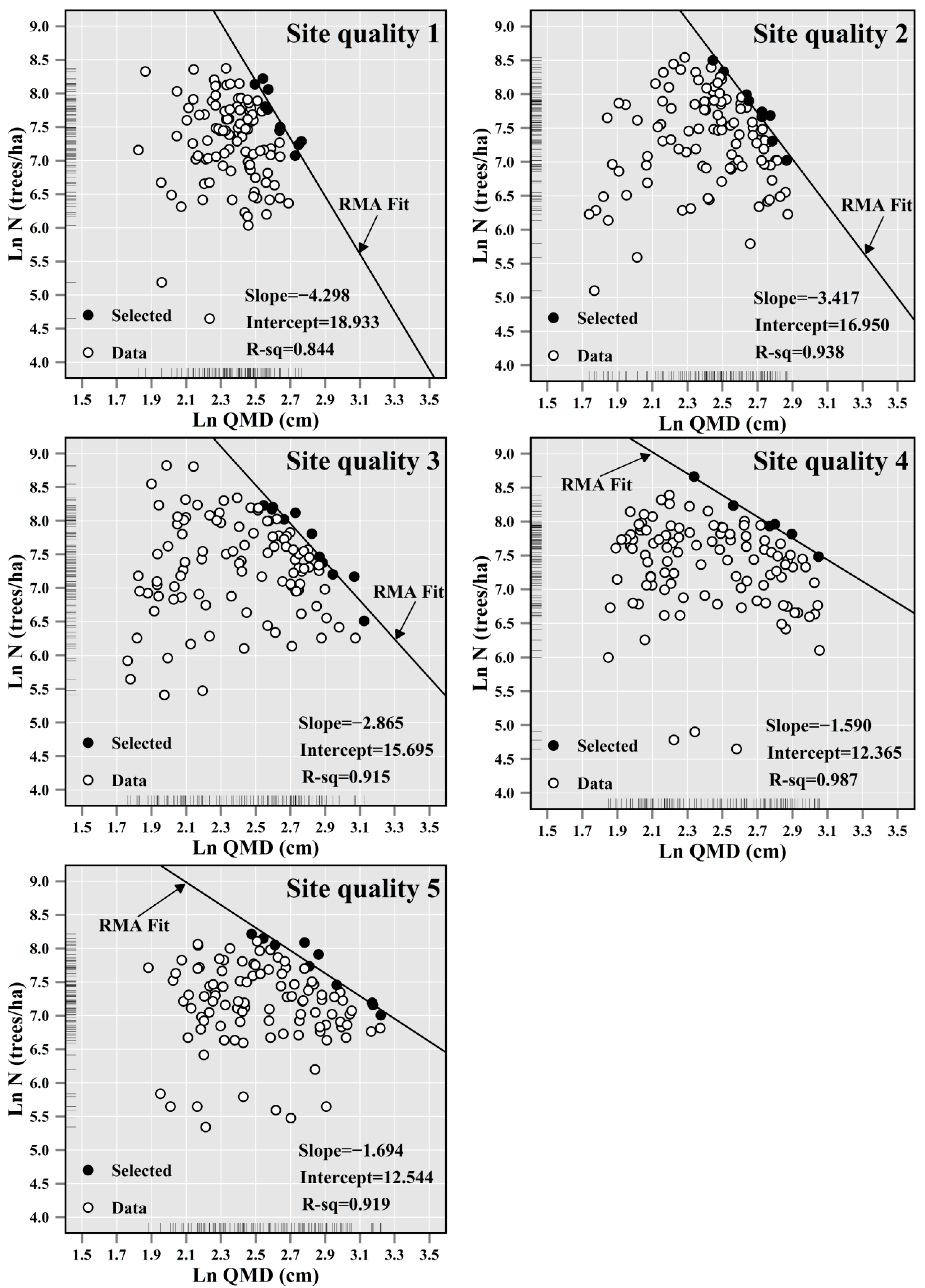
2.3. Selection of Full Stocked Plots and Regression Methods
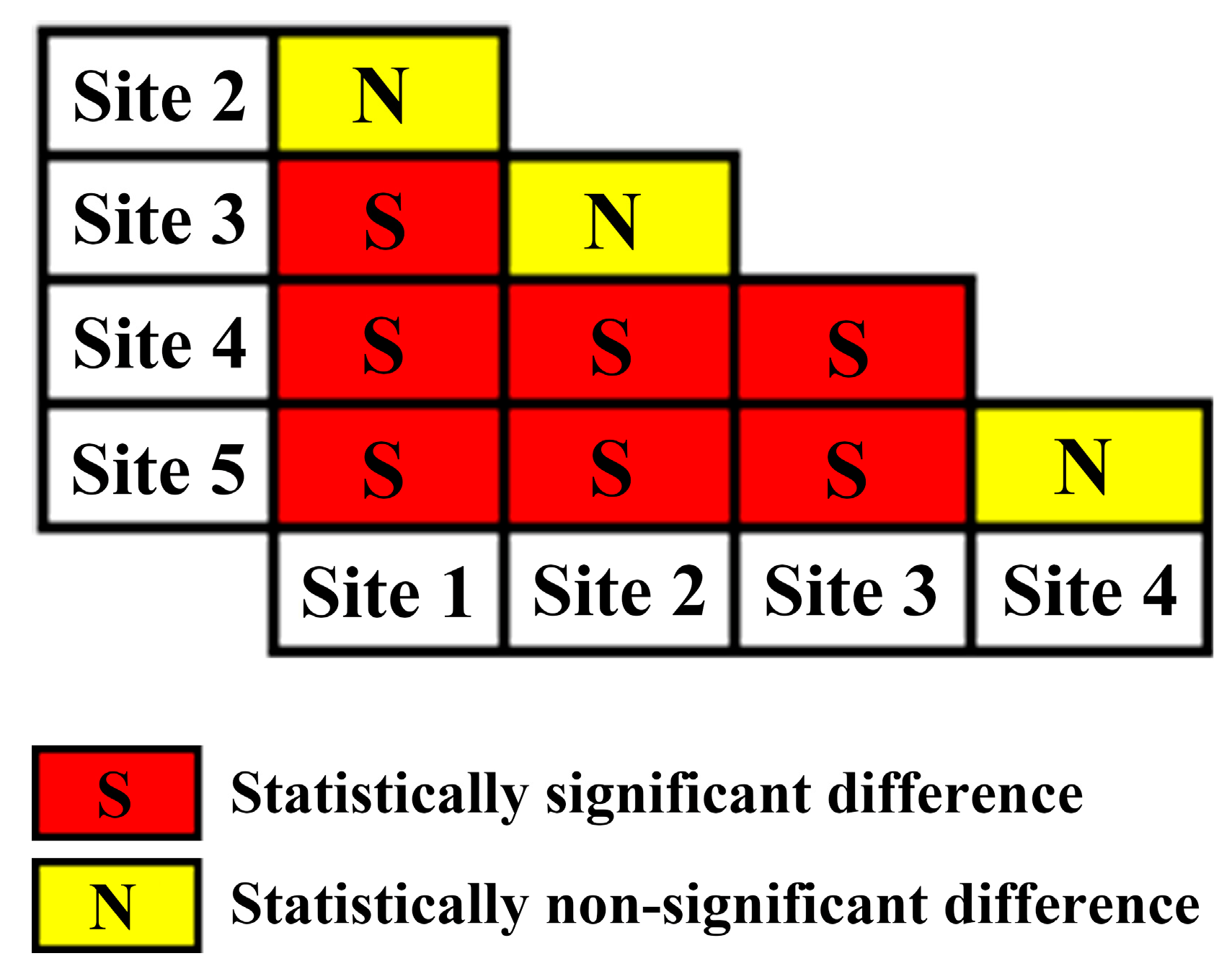
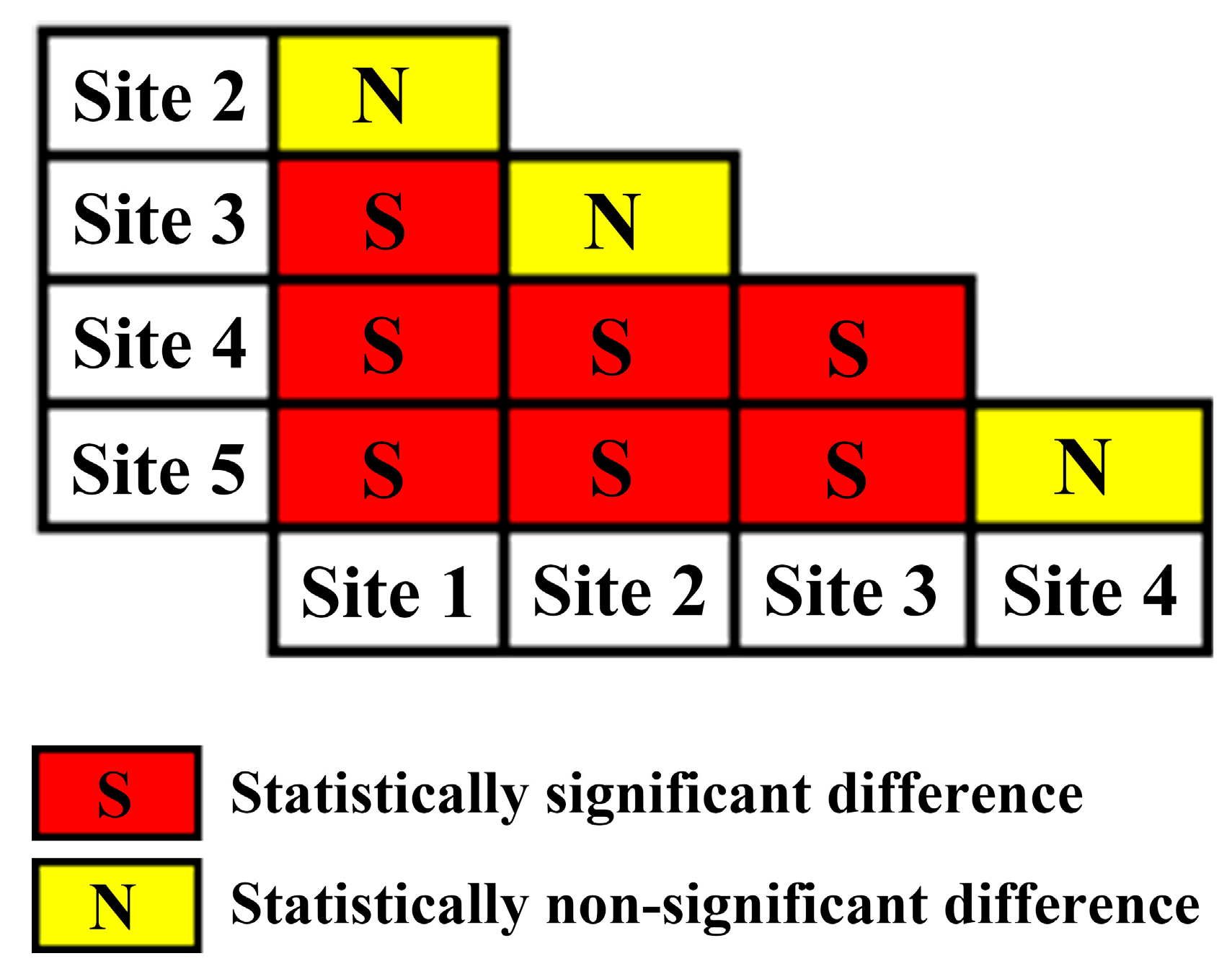
2.4. Significance Test of Slope Difference and Model Evaluation
3. Results
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Cubbage, F.; Harou, P.; Sills, E. Policy instruments to enhance multi-functional forest management. For. Policy Econ. 2007, 9, 833–851. [Google Scholar] [CrossRef]
- Meng, J.; Lu, Y.; Zeng, J. Transformation of a degraded Pinus massoniana plantation into a mixed-species irregular forest: Impacts on stand structure and growth in southern China. Forests 2014, 5, 3199–3221. [Google Scholar] [CrossRef]
- Mantau, U. Recreational and Environmental Markets for Forest Enterprises: A New Approach towards Marketability of Public Goods; CABI Publishing: New York, NY, USA, 2001. [Google Scholar] [CrossRef]
- Kanninen, M. Plantation forests: Global perspectives. In Ecosystem Goods and Services from Plantation Forests; Bauhus, J., van der Meer, P., Kanninen, M., Eds.; Earthscan Press: London, UK, 2010; pp. 1–15. [Google Scholar] [CrossRef]
- Zeng, W.; Tomppo, E.; Healey, S.P.; Gadow, K.V. The national forest inventory in China: History-results-international context. For. Ecosyst. 2015, 2, 1–16. [Google Scholar] [CrossRef]
- Macdicken, K.; Jonsson, Ö.; Piña, L.; Maulo, S.; Adikari, Y.; Garzuglia, M.; Lindquist, E.; Reams, G.; D’Annunzio, R. Global Forest Resources Assessment 2015: How Are the World's Forests Changing? Food And Agriculture Organization of the United Nations: Rome, Italy, 2015; ISBN 9789251088210. [Google Scholar]
- State Forestry Administration. Report of Forest Resources in China (2009–2013); China Forestry Press: Beijing, China, 2014; ISBN 9787503874246.
- Drew, T.J.; Flewelling, J.W. Stand density management: An alternative approach and its application to Douglas-fir plantations. For. Sci. 1979, 25, 518–532. [Google Scholar]
- Bormann, B.; Gordon, J. Stand density effects in young red alder plantations: Productivity, photosynthate partitioning, and nitrogen fixation. Ecology 1984, 65, 394–402. [Google Scholar] [CrossRef]
- Castano-Santamaria, J.; Barrio-Anta, M.; Alvarez-Alvarez, P. Regional-scale stand density management diagrams for Pyrenean oak (Quercus pyrenaica Willd.) stands in north-west Spain. Iforest 2013, 6, 113–122. [Google Scholar] [CrossRef]
- Castedo-Dorado, F.; Crecente-Campo, F.; Álvarez-Álvarez, P.; Barrio-Anta, M. Development of a stand density management diagram for radiata pine stands including assessment of stand stability. Forestry 2009, 82, 1–16. [Google Scholar] [CrossRef]
- Zhang, J.; Oliver, W.W.; Powers, R.F. Reevaluating the self-thinning boundary line for ponderosa pine (Pinus ponderosa) forests. Can. J. For. Res. 2013, 43, 963–971. [Google Scholar] [CrossRef]
- Drew, T.J.; Flewelling, J.W. Some recent Japanese theories of yield-density relationships and their application to monterey pine plantations. For. Sci. 1977, 23, 517–534. [Google Scholar]
- White, J. Demographic factors in populations of plants. In Demography and Evolution in Plant Populations; Solbrig, O.T., Ed.; Blackwell Publishing: Oxford, UK, 1980; pp. 21–48. [Google Scholar] [CrossRef]
- R Yoda, K.; Kira, T.; Ogawa, H.; Hozumi, K. Self-thinning in overcrowded pure stands under cultivated and natural conditions. J. Biol. Osaka City Univ. 1963, 14, 107–129. [Google Scholar]
- Westoby, M. The self-thinning rule. Adv. Ecol. Res. 1984, 14, 167–225. [Google Scholar] [CrossRef]
- Zhang, L.; Bi, H.; Gove, J.H.; Heath, L.S. A comparison of alternative methods for estimating the self-thinning boundary line. Can. J. For. Res. 2005, 35, 1507–1514. [Google Scholar] [CrossRef]
- Vospernik, S.; Sterba, H. Do competition-density rule and self-thinning rule agree? Ann. For. Sci. 2015, 72, 379–390. [Google Scholar] [CrossRef]
- Reineke, L.H. Perfecting a stand-density index for even-aged forests. J. Agric. Res. 1933, 46, 627–638. [Google Scholar]
- Burkhart, H.E.; Tomé, M. Modeling Forest Trees and Stands; Springer Science & Business Media: Berlin, Germany, 2012. [Google Scholar] [CrossRef]
- White, J.; Harper, J.L. Correlated changes in plant size and number in plant populations. J. Ecol. 1970, 58, 467–485. [Google Scholar] [CrossRef]
- Gorham, E. Shoot height; weight and standing crop in relation to density of monospecific plant Stands. Nature 1979, 279, 148–150. [Google Scholar] [CrossRef]
- Jack, S.B.; Long, J.N. Linkages between silviculture and ecology: An analysis of density management diagrams. For. Ecol. Manag. 1996, 86, 205–220. [Google Scholar] [CrossRef]
- Zeide, B. Tolerance and self-tolerance of trees. For. Ecol. Manag. 1985, 13, 149–166. [Google Scholar] [CrossRef]
- Weller, D.E. A reevaluation of the −3/2 power rule of plant self-thinning. Ecol. Monogr. 1987, 57, 23–43. [Google Scholar] [CrossRef]
- Weller, D.E. Self-thinning exponent correlated with allometric measures of plant geometry. Ecology 1987, 68, 813–821. [Google Scholar] [CrossRef]
- Zeide, B. Analysis of the 3/2 power law of self-thinning. For. Sci. 1987, 33, 517–537. [Google Scholar]
- Lonsdale, W.M. The self-thinning rule: Dead or alive? Ecology 1990, 71, 550–579. [Google Scholar] [CrossRef]
- Weller, D.E. Will the real self-thinning rule please stand up?—A Reply to Osawa and Sugita. Ecology 1990, 71, 1204–1207. [Google Scholar] [CrossRef]
- Puettmann, K.J.; Hann, D.W.; Hibbs, D.E. Evaluation of the size-density relationships for pure red alder and Douglas-fir stands. For. Sci. 1993, 39, 7–27. [Google Scholar]
- Hamilton, N.S.; Matthew, C.; Lemaire, G. In defence of the −3/2 boundary rule: A re-evaluation of self-thinning concepts and status. Ann. Bot. 1995, 76, 569–577. [Google Scholar] [CrossRef]
- Zeide, B. Natural thinning and environmental change: An ecological process model. For. Ecol. Manag. 2001, 154, 165–177. [Google Scholar] [CrossRef]
- Morris, E.C. Self-thinning lines differ with fertility level. Ecol. Res. 2002, 17, 17–28. [Google Scholar] [CrossRef]
- Pretzsch, H. A unified law of spatial allometry for woody and herbaceous plants. Plant Biol. 2002, 4, 159–166. [Google Scholar] [CrossRef]
- Solomon, D.S.; Zhang, L. Maximum size–density relationships for mixed softwoods in the northeastern USA. For. Ecol. Manag. 2002, 155, 163–170. [Google Scholar] [CrossRef]
- Pretzsch, H. Stand density and growth of Norway spruce (Picea abies (L.) Karst.) and European beech (Fagus sylvatica L.): Evidence from long-term experimental plots. Eur. J. For. Res. 2005, 124, 193–205. [Google Scholar] [CrossRef]
- Pretzsch, H.; Biber, P. A re-evaluation of Reineke’s rule and stand density index. For. Sci. 2005, 51, 304–320. [Google Scholar]
- Pretzsch, H. Species-specific allometric scaling under self-thinning: Evidence from long-term plots in forest stands. Oecologia 2006, 146, 572–583. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.H.; Zhang, J.G.; Duan, A.G.; Sun, H.G. Review of studies on maximum size-density rules. J. Plant Ecol. 2008, 32, 501–511. [Google Scholar] [CrossRef]
- Osawa, A. Inverse relationship of crown fractal dimension to self-thinning exponent of tree populations: A hypothesis. Can. J. For. Res. 1995, 25, 1608–1617. [Google Scholar] [CrossRef]
- Kikuzawa, K. Theoretical relationships between mean plant size; size distribution and self thinning under one-sided competition. Ann. Bot. 1999, 83, 11–18. [Google Scholar] [CrossRef]
- Li, B.L.; Wu, H.I.; Zou, G. Self-thinning rule: A causal interpretation from ecological field theory. Ecol. Model 2000, 132, 167–173. [Google Scholar] [CrossRef]
- Del Río, M.; Montero, G.; Bravo, F. Analysis of diameter–density relationships and self-thinning in non-thinned even-aged Scots pine stands. For. Ecol. Manag. 2001, 142, 79–87. [Google Scholar] [CrossRef]
- Yang, Y.; Titus, S.J. Maximum size–density relationship for constraining individual tree mortality functions. For. Ecol. Manag. 2002, 168, 259–273. [Google Scholar] [CrossRef]
- Skovsgaard, J.P.; Vanclay, J.K. Forest site productivity: A review of the evolution of dendrometric concepts for even-aged stands. Forestry 2008, 81, 13–31. [Google Scholar] [CrossRef]
- Weiskittel, A.R.; Hann, D.W.; Kershaw, J.A., Jr.; Vanclay, J.K. Forest Growth and Yield Modeling; John Wiley & Sons Incorporated: Hoboken, NJ, USA, 2011. [Google Scholar] [CrossRef]
- Lin, K.; Guo, Y.; Yu, X.; Zhang, W.; Chen, Y. The structure characters of undergrowth vegetation composition in the old-growth Chinese fir forest. J. Fujian Coll. For. 1999, 19, 124–128. [Google Scholar] [CrossRef]
- Chen, C.; Wang, S. Ecology of Mixed Plantation Forest; Science Press: Beijing, China, 2004; ISBN 9787030134745. [Google Scholar]
- Wang, Q.; Wang, S.; Huang, Y. Comparisons of litterfall, litter decomposition and nutrient return in a monoculture Cunninghamia lanceolata and a mixed stand in southern China. For. Ecol. Manag. 2008, 255, 1210–1218. [Google Scholar] [CrossRef]
- Wang, W.; Duan, Y.; Zhang, L.; Wang, B.; Li, X. Effects of different rotations on carbon sequestration in Chinese fir plantations. Chin. J. Plant Ecol. 2016, 40, 669–678. [Google Scholar] [CrossRef]
- Yu, X. The research of Chinese fir in China. J. Fujian Coll. For. 1988, 8, 203–220. [Google Scholar] [CrossRef]
- State Forestry Administration. Code of Forest Harvesting (LY/T 1646−2005); State Forestry Administration: Beijing, China, 2005.
- Duan, A.; Zhang, J. Modeling of dominant height growth and building of polymorphic site index equations of Chinese fir plantation. Sci. Silvae Sin. 2004, 40, 13–19. [Google Scholar] [CrossRef]
- Li, Z.; Meng, J. Selection of optimal height–diameter models for Cunninghamia Lanceolata in Fujian Province based on cross–validation analysis. J. Northwest For. Univ. 2016, 31, 92–96. [Google Scholar] [CrossRef]
- Sharma, M.; Amateis, R.L.; Burkhart, H.E. Top height definition and its effect on site index determination in thinned and unthinned loblolly pine plantations. For. Ecol. Manag. 2002, 168, 163–175. [Google Scholar] [CrossRef]
- Wilson, D.S.; Seymour, R.S.; Maguire, D.A. Density management diagram for northeastern red spruce and balsam fir forests. N. J. Appl. For. 1999, 16, 48–56. [Google Scholar]
- Inoue, A.; Miyake, M.; Nishizono, T. Allometric model of the Reineke equation for Japanese cypress (Chamaecyparis obtusa) and red pine (Pinus densiflora) stands. J. For. Res. 2004, 9, 319–324. [Google Scholar] [CrossRef]
- Bi, H.; Turvey, N.D. A method of selecting data points for fitting the maximum biomass–density line for stands undergoing self-thinning. Aust. J. Ecol. 1997, 22, 356–359. [Google Scholar] [CrossRef]
- Blackburn, T.M.; Lawton, J.H.; Perry, J.N. A method of estimating the slope of upper bounds of plots of body size and abundance in natural animal assemblages. Oikos 1992, 65, 107–112. [Google Scholar] [CrossRef]
- Leduc, D.J. A comparative analysis of the reduced major axis technique of fitting lines to bivariate data. Can. J. For. Res. 1987, 17, 654–659. [Google Scholar] [CrossRef]
- Bohonak, A.J. RMA: Software for Reduced Major Axis Regression, Version 1.17; San Diego State University: San Diego, CA, USA, 2004. Available online: http://www.bio.sdsu.edu/pub/andy/rma.html (accessed on 3 September 2004).
- Sokal, R.R.; Rohlf, F.J. Biometry, 3rd ed.; W.H. Freeman and Company: New York, NY, USA, 1995; ISBN 0716724111. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015; Available online: https://www.r-project.org/ (accessed on 20 June 2015).
- Legendre, P. lmodel2: Model II Regression. In R Package; Version 1.7-2; Legendre, P., Ed.; R Foundation for Statistical Computing: Vienna, Austria, 2014; Available online: http://CRAN.R-project.org/package=lmodel2 (accessed on 24 February 2014).
- Comeau, P.G.; White, M.; Kerr, G.; Hale, S.E. Maximum density–size relationships for Sitka spruce and coastal Douglas-fir in Britain and Canada. Forestry 2010. [Google Scholar] [CrossRef]
- Cameron, I.R. Evaluation of the density management diagram for coastal Douglas-fir. FRDA Rep. 1988, 426, 311–316. [Google Scholar]
- Bégin, E.; Bégin, J.; Bélanger, L.; Rivest, L.P.; Tremblay, S. Balsam fir self-thinning relationship and its constancy among different ecological regions. Can. J. For. Res. 2001, 31, 950–959. [Google Scholar] [CrossRef]
- Liu, J.; Wei, L.; Wang, C.M.; Wang, G.X.; Wei, X.P. Effect of water deficit on self-thinning line in spring wheat (Triticum aestivum L.) populations. J. Integr. Plant Biol. 2006, 48, 415–419. [Google Scholar] [CrossRef]
- Bai, G.Y.; Zhang, W.; Jia, X.; Wang, N.; Zhou, L.; Xu, S.; Wang, G. Variation in root: Shoot ratios induced the differences between above and belowground mass-density relationships along an aridity gradient. Acta. Oecol. 2010, 36, 393–395. [Google Scholar] [CrossRef]
- Morris, E.; Myerscough, P. Self-thinning and competition intensity over a gradient of nutrient availability. J. Ecol. 1991, 79, 903–923. [Google Scholar] [CrossRef]
- Dewar, R.C. A mechanistic analysis of self-thinning in terms of the carbon balance of trees. Ann. Bot. 1993, 71, 147–159. [Google Scholar] [CrossRef]
- Chu, C.J.; Weiner, J.; Maestre, F.T.; Wang, Y.S.; Morris, C.; Xiao, S.; Yuan, J.L.; Du, G.Z.; Wang, G. Effects of positive interactions; size symmetry of competition and abiotic stress on self-thinning in simulated plant populations. Ann. Bot. 2010. [Google Scholar] [CrossRef] [PubMed]
- Weiskittel, A.; Gould, P.; Temesgen, H. Sources of variation in the self-thinning boundary line for three species with varying levels of shade tolerance. For. Sci. 2009, 55, 84–93. [Google Scholar]
- Chen, H.Y.; Krestov, P.V.; Klinka, K. Trembling aspen site index in relation to environmental measures of site quality at two spatial scales. Can. J. For. Res. 2002, 32, 112–119. [Google Scholar] [CrossRef]
- Ercanli, I.; Gunlu, A.; Altun, L.; Zeki Baskent, E. Relationship between site index of oriental spruce [Picea orientalis (L.) Link] and ecological variables in Maçka, Turkey. Scand. J. For. Res. 2008, 23, 319–329. [Google Scholar] [CrossRef]
- Corona, P.; Scotti, R.; Tarchiani, N. Relationship between environmental factors and site index in Douglas-fir plantations in central Italy. For. Ecol. Manag. 1998, 110, 195–207. [Google Scholar] [CrossRef]
- Reinhardt, E.D. Influence of Site Quality on the Height-Diameter Relationship of Western Larch; Dissertation; University of Montana: Missoula, MT, USA, 1982. [Google Scholar]
- Waring, R.H.; Milner, K.S.; Jolly, W.M.; Phillips, L.; Mcwethy, D. Assessment of site index and forest growth capacity across the Pacific and Inland Northwest USA with a MODIS satellite-derived vegetation index. For. Ecol. Manag. 2006, 228, 285–291. [Google Scholar] [CrossRef]
- Sánchez-González, M.; del Río, M.; Canellas, I.; Montero, G. Distance independent tree diameter growth model for cork oak stands. For. Ecol. Manag. 2006, 225, 262–270. [Google Scholar] [CrossRef]
- Fosbroke, D.E.; Hicks, R.R., Jr. Tree Mortality Following Gypsy Moth Defoliation in Southwestern Pennsylvania; USDA Forest Service General Technical Report NC-North Central Forest Experiment Station: Newtown Square, PA, USA, 1989. [Google Scholar]
- Davidson, C.B.; Gottschalk, K.W.; Johnson, J.E. Tree mortality following defoliation by the European gypsy moth (Lymantria dispar L.) in the United States: A review. For. Sci. 1999, 45, 74–84. [Google Scholar]
- DeBell, D.S.; Harms, W.R.; Whitesell, C.D. Stockability: A major factor in productivity differences between Pinus taeda plantations in Hawaii and the southeastern United States. For. Sci. 1989, 35, 708–719. [Google Scholar]
- Morris, E. Self-thinning in Ocimum basilicum grown at three soil fertility levels with and without mycorrhizal inoculum. Proc. Linn. Soc. New South Wales 1995, 115, 89–107. [Google Scholar]
- Morris, E.C. Density-dependent mortality induced by low nutrient status of the substrate. Ann. Bot. 1999, 84, 95–107. [Google Scholar] [CrossRef]
- Zhang, J.; Ritchie, M.W.; Maguire, D.A.; Oliver, W.W. Thinning ponderosa pine (Pinus ponderosa) stands reduces mortality while maintaining stand productivity. Can. J. For. Res. 2013, 43, 311–320. [Google Scholar] [CrossRef]
- Watkinson, A. Density-dependence in single-species populations of plants. J. Theor. Biol. 1980, 83, 345–357. [Google Scholar] [CrossRef]
- Morris, E. Effect of localized placement of nutrients on root competition in self-thinning populations. Ann. Bot. 1996, 78, 353–364. [Google Scholar] [CrossRef]
- Bloom, A.J.; Chapin, F.S.; Mooney, H.A. Resource limitation in plants—An economic analogy. Ann. Rev. Ecol. Syst. 1985, 16, 363–392. [Google Scholar] [CrossRef]
- Weiner, J. Allocation, plasticity and allometry in plants. Perspect. Plant. Ecol. 2004, 6, 207–215. [Google Scholar] [CrossRef]
- Zhang, W.P.; Jia, X.; Bai, Y.Y.; Wang, G.X. The difference between above-and below-ground self-thinning lines in forest communities. Ecol. Res. 2011, 26, 819–825. [Google Scholar] [CrossRef]
- Deng, J.M.; Wang, G.X.; Morris, E.C.; Wei, X.P.; Li, D.X.; Chen, B.M.; Zhao, C.M.; Liu, J.; Wang, Y. Plant mass–density relationship along a moisture gradient in north-west China. J. Ecol. 2006, 94, 953–958. [Google Scholar] [CrossRef]
- Bi, H. Stochastic frontier analysis of a classic self-thinning experiment. Austral Ecol. 2004, 29, 408–417. [Google Scholar] [CrossRef]
- Kumar, B.M.; Long, J.N.; Kumar, P. A density management diagram for teak plantations of Kerala in peninsular India. For. Ecol. Manag. 1995, 74, 125–131. [Google Scholar] [CrossRef]
- Luis, J.F.S.; Fonseca, T.F. The allometric model in the stand density management of Pinus pinaster Ait. In Portugal. Ann. For. Sci. 2004, 61, 807–814. [Google Scholar] [CrossRef]
- Shaw, J.D.; Long, J.N. A density management diagram for longleaf pine stands with application to red-cockaded woodpecker habitat. South J. Appl. For. 2007, 31, 28–38. [Google Scholar]
- Vacchiano, S.G.; Derose, R.J.; Shaw, J.D.; Svoboda, M.; Motta, R. A density management diagram for Norway spruce in the temperate European montane region. Eur. J. For. Res. 2013, 132, 535–549. [Google Scholar] [CrossRef]
- Tang, S.; Meng, F.R.; Meng, C.H. The impact of initial stand density and site index on maximum stand density index and self-thinning index in a stand self-thinning model. For. Ecol. Manag. 1995, 75, 61–68. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Variable | Mean | S.D. | Min. | Max. |
|---|---|---|---|---|
| Age (a) | 19 | 10 | 4 | 46 |
| Density (stems/ha) | 1801 | 1047 | 104 | 6791 |
| Quadratic mean diameter (cm) | 12.1 | 3.8 | 5.7 | 25.0 |
| Elevation (m) | 483 | 222 | 43 | 1350 |
| Soil depth (cm) | 93.8 | 18.8 | 9.0 | 160.0 |
| Humus depth (cm) | 9.5 | 5.2 | 0 | 42.0 |
| Litter depth (cm) | 3.8 | 5.5 | 0 | 30.0 |
| Slope (°) | 25.9 | 7.1 | 3.0 | 46.0 |
| Azimuth (°) | 194.8 | 104.4 | 45.0 | 360.0 |
| Site index (m) | 22.8 | 5.2 | 8.3 | 42.2 |
| Site Quality Categories (SI Interval) | No. of Plots (Selected) 1 | Slope (β) | Standard Error | Confidence Interval 2 | RMSE | R2 |
|---|---|---|---|---|---|---|
| 1 (8.32, 18.51) | 110 (10) | −4.298 | 0.600 | (−5.898, −3.132) | 0.154 | 0.844 |
| 2 (18.53, 21.30) | 110 (9) | −3.417 | 0.320 | (−4.257, −2.743) | 0.109 | 0.938 |
| 3 (21.35, 23.48) | 111 (11) | −2.865 | 0.279 | (−3.564, −2.304) | 0.157 | 0.915 |
| 4 (23.49, 26.24) | 111 (6) | −1.590 | 0.090 | (−1.860, −1.360) | 0.041 | 0.987 |
| 5 (26.27, 42.16) | 111 (10) | −1.694 | 0.171 | (−2.132, −1.346) | 0.125 | 0.919 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ge, F.; Zeng, W.; Ma, W.; Meng, J. Does the Slope of the Self-thinning Line Remain a Constant Value across Different Site Qualities?—An Implication for Plantation Density Management. Forests 2017, 8, 355. https://doi.org/10.3390/f8100355
Ge F, Zeng W, Ma W, Meng J. Does the Slope of the Self-thinning Line Remain a Constant Value across Different Site Qualities?—An Implication for Plantation Density Management. Forests. 2017; 8(10):355. https://doi.org/10.3390/f8100355
Chicago/Turabian StyleGe, Fangxing, Weisheng Zeng, Wu Ma, and Jinghui Meng. 2017. "Does the Slope of the Self-thinning Line Remain a Constant Value across Different Site Qualities?—An Implication for Plantation Density Management" Forests 8, no. 10: 355. https://doi.org/10.3390/f8100355
APA StyleGe, F., Zeng, W., Ma, W., & Meng, J. (2017). Does the Slope of the Self-thinning Line Remain a Constant Value across Different Site Qualities?—An Implication for Plantation Density Management. Forests, 8(10), 355. https://doi.org/10.3390/f8100355