
Decomposition of Leaves and Fine Roots in Three Subtropical Plantations in China Affected by Litter Substrate Quality and Soil Microbial Community

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Description
2.2. Experimental Design and Mass Loss Estimation
2.3. Litter Chemical Analysis
2.4. Soil Microbial Community Analysis
2.5. Statistical Analysis
3. Results
3.1. Initial Chemical Properties
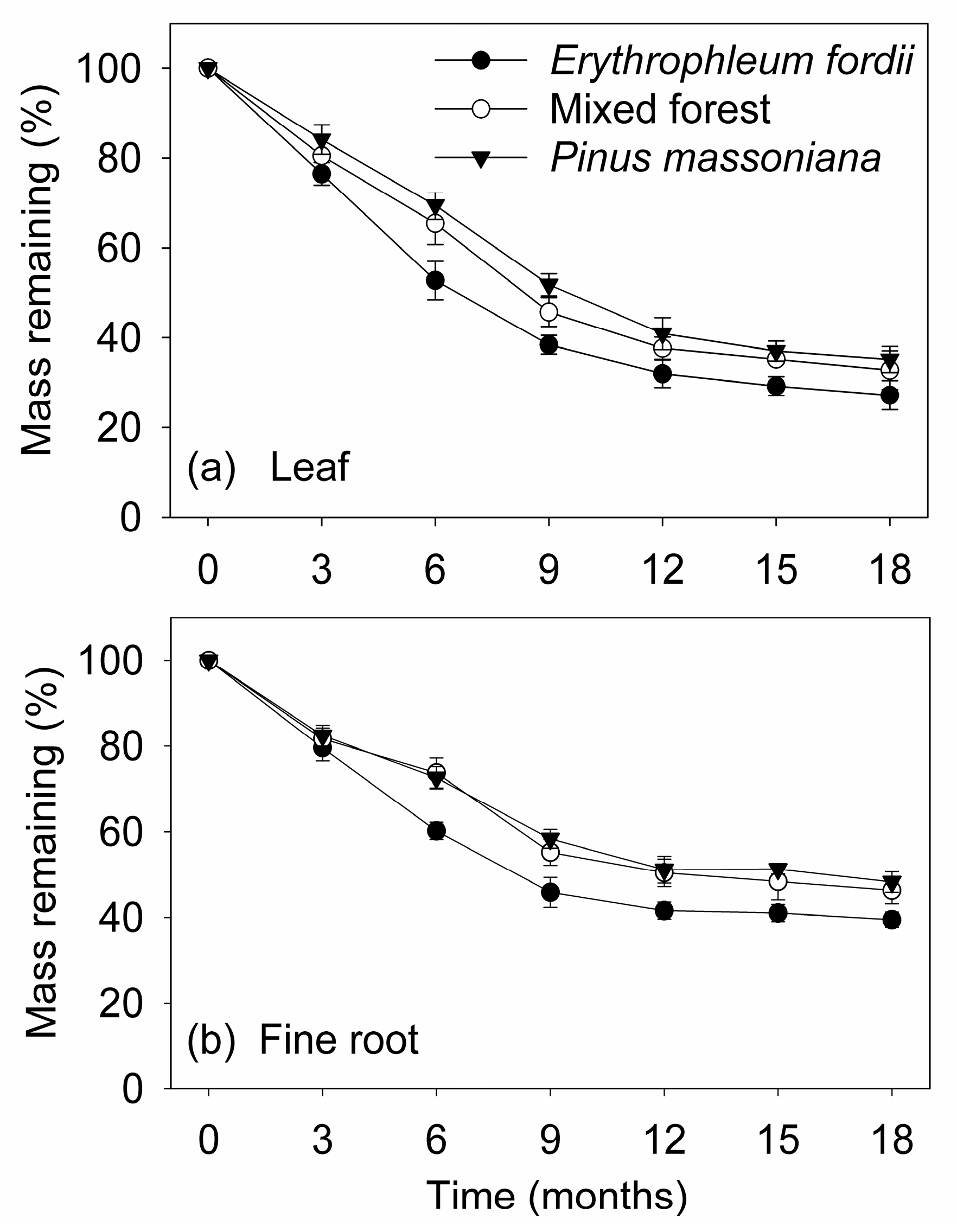
3.2. Dry Mass Loss during Decomposition
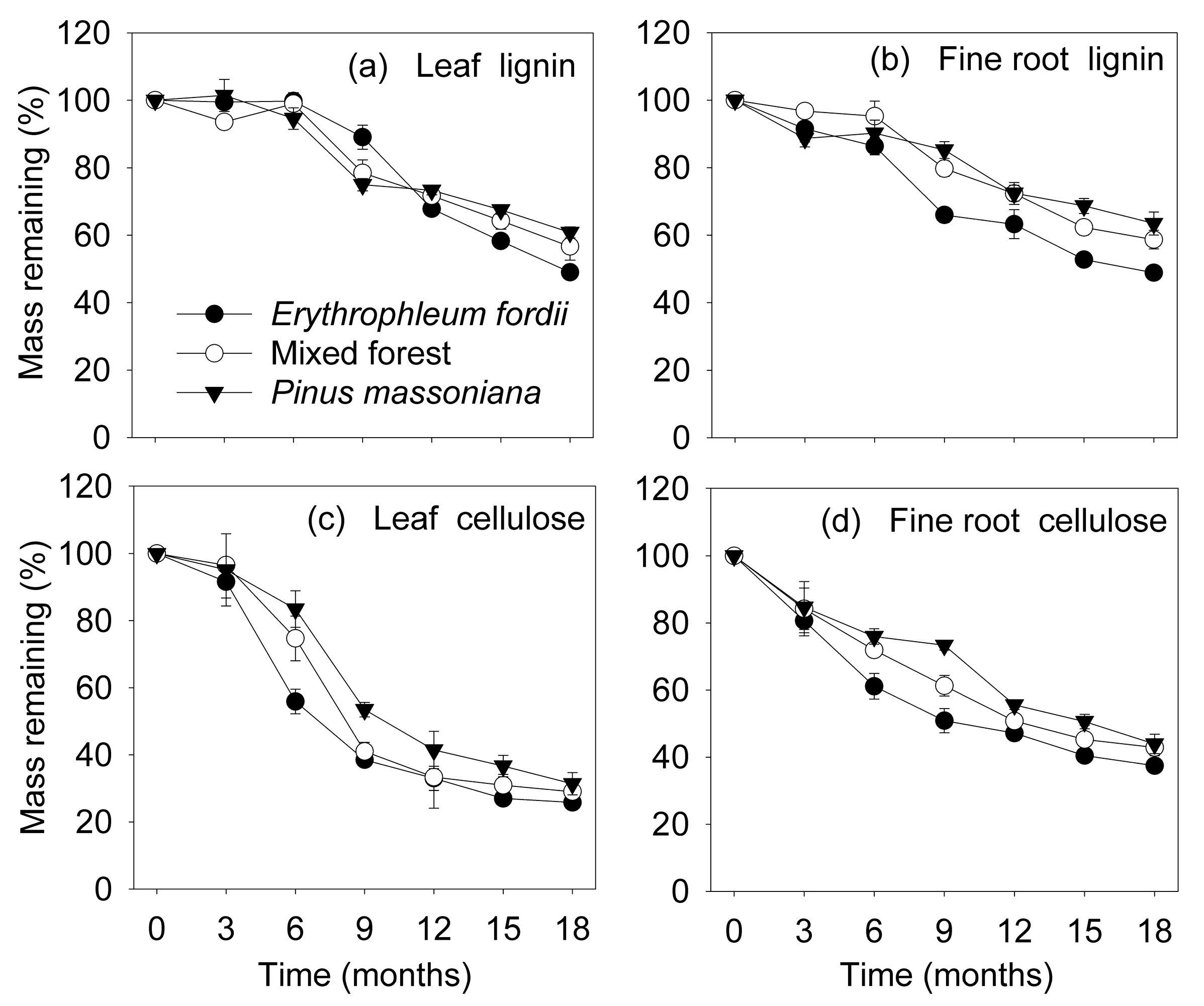
3.3. Organic Chemical Components during Decomposition
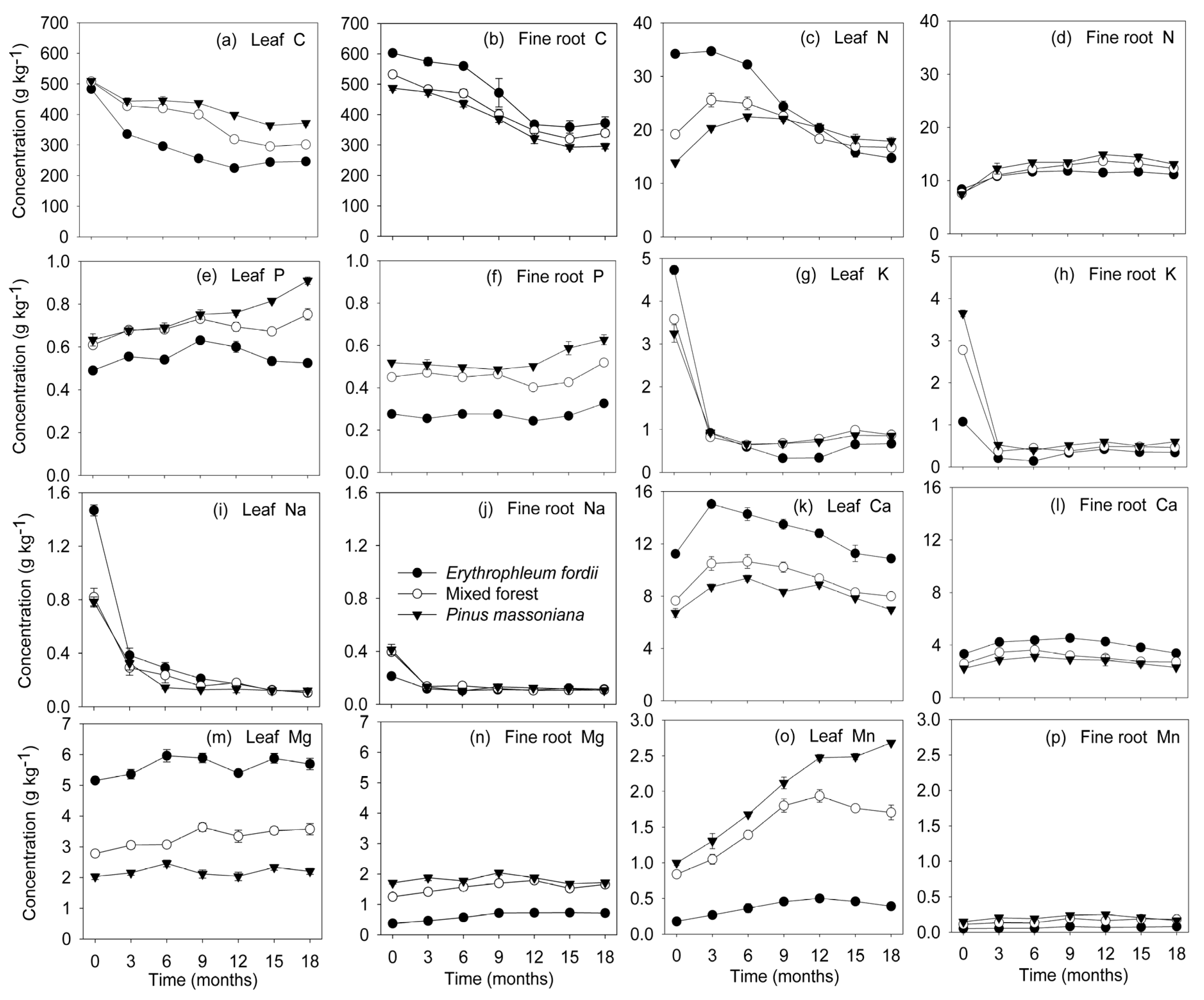
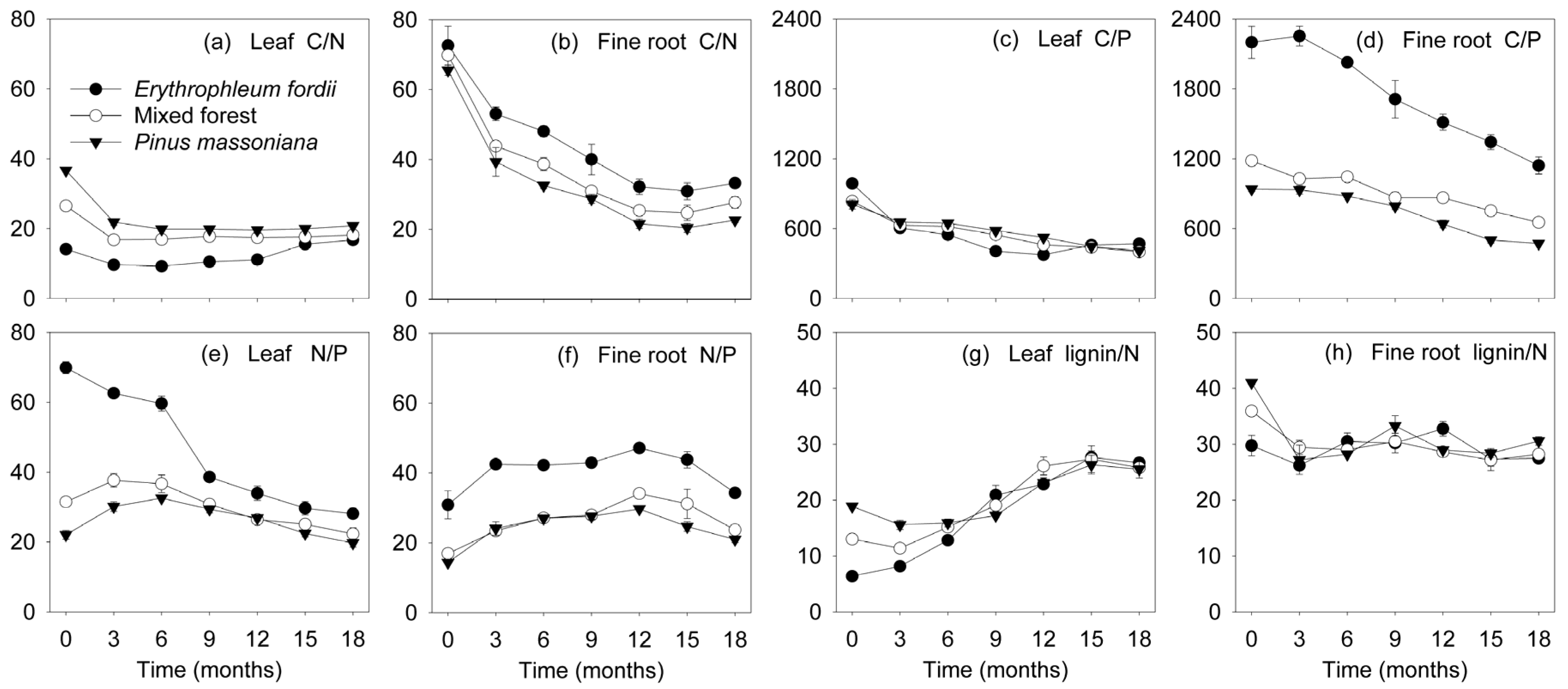
3.4. Nutrient Dynamics during Decomposition
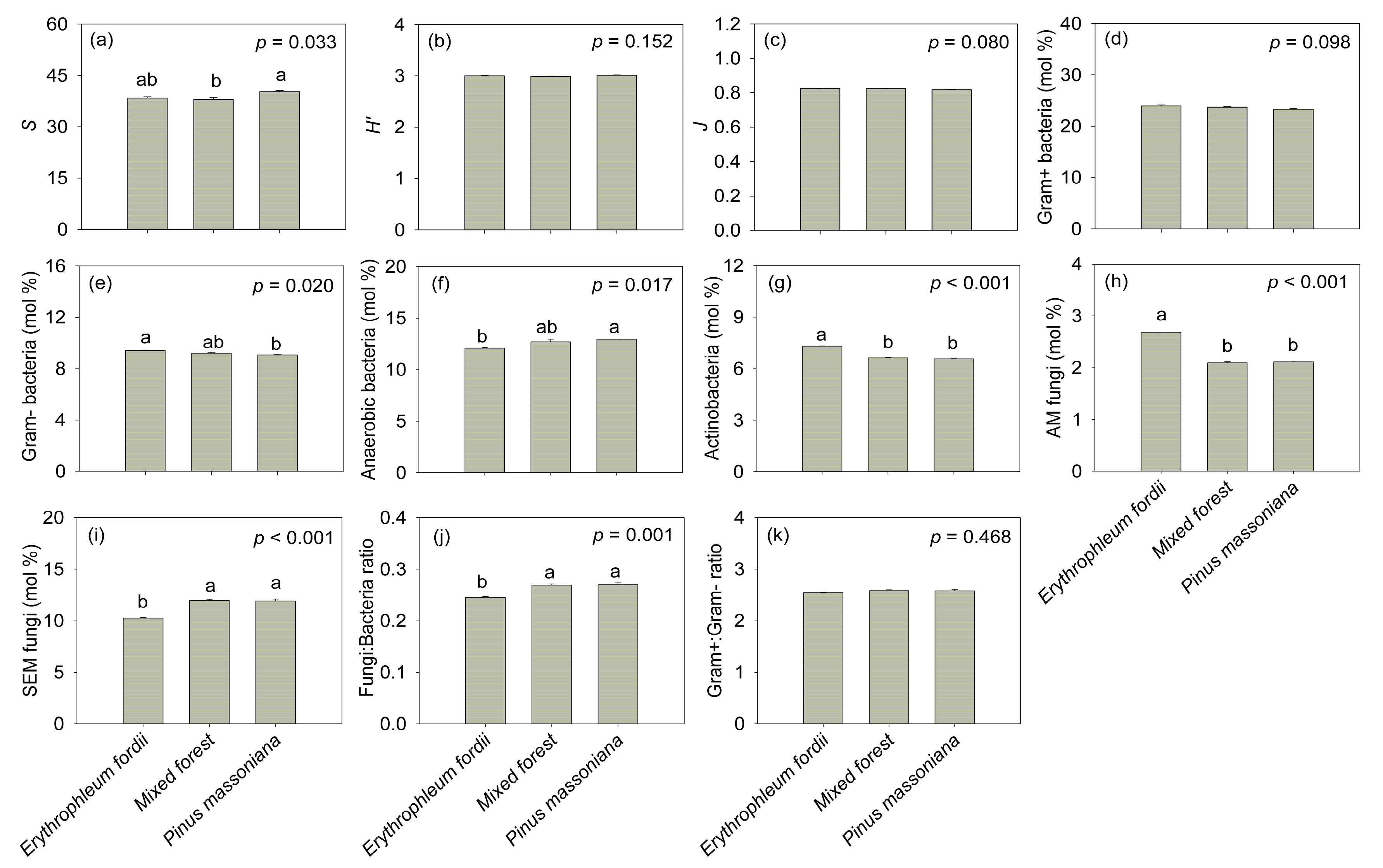
3.5. Soil Microbial Community
4. Discussion
4.1. Decomposition of Leaves and Fine Roots
4.2. Effects of Tree Species on Litter Decomposition
4.3. Nutrient Dynamics during Decomposition
4.4. Soil Microbial Influence on Decomposition
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Swift, M.J.; Heal, O.W.; Anderson, J.M. Decomposition in Terrestrial Ecosystems; University of California Press: Berkeley, CA, USA, 1979. [Google Scholar]
- Norby, R.J.; Ledford, J.; Reilly, C.D.; Miller, N.E.; O’Neill, E.G. Fine-root production dominates response of a deciduous forest to atmospheric CO2 enrichment. Proc. Natl. Acad. Sci. USA 2004, 101, 9689–9693. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Su, J.; Janssens, I.A.; Zhou, G.; Xiao, C. Fine root and litterfall dynamics of three Korean pine (Pinus koraiensis) forests along an altitudinal gradient. Plant Soil 2014, 374, 19–32. [Google Scholar] [CrossRef]
- Ball, B.A.; Carrillo, Y.; Molina, M. The influence of litter composition across the litter-soil interface on mass loss, nitrogen dynamics and the decomposer community. Soil Biol. Biochem. 2014, 69, 71–82. [Google Scholar] [CrossRef]
- Aponte, C.; García, L.V.; Maranon, T. Tree species effect on litter decomposition and nutrient release in mediterranean oak forests changes over time. Ecosystems 2012, 15, 1204–1218. [Google Scholar] [CrossRef] [Green Version]
- Cizungu, L.; Staelens, J.; Huygens, D.; Walangululu, J.; Muhindo, D.; Van Cleemput, O.; Boeckx, P. Litterfall and leaf litter decomposition in a central African tropical mountain forest and Eucalyptus plantation. For. Ecol. Manag. 2014, 326, 109–116. [Google Scholar] [CrossRef]
- Lai, Z.; Zhang, Y.; Liu, J.; Wu, B.; Qin, S.; Fa, K. Fine-root distribution, production, decomposition, and effect on soil organic carbon of three revegetation shrub species in northwest China. For. Ecol. Manag. 2016, 359, 381–388. [Google Scholar] [CrossRef]
- Ma, C.; Xiong, Y.; Li, L.; Guo, D. Root and leaf decomposition become decoupled over time: Implications for below- and above-ground relationships. Funct. Ecol. 2016, 30, 1239–1246. [Google Scholar] [CrossRef]
- Fujii, S.; Takeda, H. Dominant effects of litter substrate quality on the difference between leaf and root decomposition process above- and belowground. Soil Biol. Biochem. 2010, 42, 2224–2230. [Google Scholar] [CrossRef]
- Hobbie, S.E. Temperature and plant species control over litter decomposition in Alaskan tundra. Ecol. Monogr. 1996, 66, 503–522. [Google Scholar] [CrossRef]
- Ostertag, R.; Hobbie, S.E. Early stages of root and leaf decomposition in Hawaiian forests: Effects of nutrient availability. Oecologia 1999, 121, 564–573. [Google Scholar] [CrossRef] [PubMed]
- Bloomfield, J.; Vogt, K.A.; Vogt, D.J. Decay rate and substrate quality of fine roots and foliage of two tropical tree species in the Luquillo Experimental Forest, Puerto Rico. Plant Soil 1993, 150, 233–245. [Google Scholar] [CrossRef]
- Lehmann, J.; Schroth, G.; Zech, W. Decomposition and nutrient release from leaves, twigs and roots of three alley-cropped tree legumes in central Togo. Agrofor. Syst. 1995, 29, 21–36. [Google Scholar] [CrossRef]
- Wang, H.; Liu, S.; Mo, J. Correlation between leaf litter and fine root decomposition among subtropical tree species. Plant Soil 2010, 335, 289–298. [Google Scholar] [CrossRef]
- Couteaux, M.M.; Bottner, P.; Berg, B. Litter decomposition, climate and liter quality. Trends Ecol. Evol. 1995, 10, 63–66. [Google Scholar] [CrossRef]
- Osono, T.; Takeda, H. Decomposition of organic chemical components in relation to nitrogen dynamics in leaf litter of 14 tree species in a cool temperate forest. Ecol. Res. 2005, 20, 41–49. [Google Scholar] [CrossRef]
- Fujimaki, R.; Takeda, H.; Wiwatiwitaya, D. Fine root decomposition in tropical dry evergreen and dry deciduous forests in Thailand. J. For. Res. 2008, 13, 338–346. [Google Scholar] [CrossRef]
- Aerts, R. Climate, leaf litter chemistry and leaf litter decomposition in terrestrial ecosystems: A triangular relationship. Oikos 1997, 79, 439–449. [Google Scholar] [CrossRef]
- Li, S.; Liu, W.; Li, D.; Li, Z.; Song, L.; Chen, K.; Fu, Y. Slower rates of litter decomposition of dominant epiphytes in the canopy than on the forest floor in a subtropical montane forest, southwest China. Soil Biol. Biochem. 2014, 70, 211–220. [Google Scholar] [CrossRef]
- Esperschütz, J.; Zimmermann, C.; Dümig, A.; Welzl, G.; Buegger, F.; Elmer, M.; Munch, J.C.; Schloter, M. Dynamics of microbial communities during decomposition of litter from pioneering plants in initial soil ecosystems. Biogeosciences 2013, 10, 5115–5124. [Google Scholar] [CrossRef]
- Smyth, C.E.; Macey, D.; Trofymow, J.A. Long-term litter decay in Canadian forests and the influence of soil microbial community and soil chemistry. Soil Biol. Biochem. 2015, 80, 251–259. [Google Scholar] [CrossRef]
- Brant, J.B.; Myrold, D.D.; Sulzman, E.W. Root controls on soil microbial community structure in forest soils. Oecologia 2006, 148, 650–659. [Google Scholar] [CrossRef] [PubMed]
- Wickings, K.; Grandy, A.S.; Reed, S.C.; Cleveland, C.C. The origin of litter chemical complexity during decomposition. Ecol. Lett. 2012, 15, 1180–1188. [Google Scholar] [CrossRef] [PubMed]
- Setälä, H.; Huhta, V. Soil fauna increase Betula pendula growth: Laboratory experiments with coniferous forest floor. Ecology 1991, 72, 665–671. [Google Scholar] [CrossRef]
- Luo, D.; Cheng, R.; Shi, Z.; Wang, W.; Xu, G.; Liu, S. Impacts of nitrogen-fixing and non-nitrogen-fixing tree species on soil respiration and microbial community composition during forest management in subtropical China. Ecol. Res. 2016, 31, 683–693. [Google Scholar] [CrossRef]
- Hobbie, S.E.; Oleksyn, J.; Eissenstat, D.M.; Reich, P.B. Fine root decomposition rates do not mirror those of leaf litter among temperate tree species. Oecologia 2010, 162, 505–513. [Google Scholar] [CrossRef] [PubMed]
- Mo, J.; Brown, S.; Xue, J.; Fang, Y.; Li, Z. Response of litter decomposition to simulated N deposition in disturbed, rehabilitated and mature forests in subtropical China. Plant Soil 2006, 282, 135–151. [Google Scholar] [CrossRef]
- Berg, B.; Berg, M.P.; Bottner, P.; Box, E.; Breymeyer, A.; de Anta, R.C.; Couteaux, M.; Escudero, A.; Gallardo, A.; Kratz, W. Litter mass loss rates in pine forests of Europe and Eastern United States: Some relationships with climate and litter quality. Biogeochemistry 1993, 20, 127–159. [Google Scholar] [CrossRef]
- Ostertag, R.; Marín-Spiotta, E.; Silver, W.L.; Schulten, J. Litterfall and decomposition in relation to soil carbon pools along a secondary forest chronosequence in Puerto Rico. Ecosystems 2008, 11, 701–714. [Google Scholar] [CrossRef]
- Howard, P.J.A.; Howard, D.M. Microbial decomposition of tree and shrub leaf litter. Oikos 1974, 25, 341–352. [Google Scholar] [CrossRef]
- Berg, B. Decomposition patterns for foliar litter—A theory for influencing factors. Soil Biol. Biochem. 2014, 78, 222–232. [Google Scholar] [CrossRef]
- Van Soest, P.; Wine, R. Determination of lignin and cellulose in acid-detergent fiber with permanganate. J. Assoc. Off. Anal. Chem. 1968, 51, 780–785. [Google Scholar]
- Bossio, D.A.; Scow, K.M.; Gunapala, N.; Graham, K.J. Determinants of soil microbial communities: Effects of agricultural management, season, and soil type on phospholipid fatty acid profiles. Microb. Ecol. 1998, 36, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Frostegård, Å.; Bååth, E. The use of phospholipid fatty acid analysis to estimate bacterial and fungal biomass in soil. Biol. Fert. Soils 1996, 22, 59–65. [Google Scholar] [CrossRef]
- Fichtner, A.; Von Oheimb, G.; Härdtle, W.; Wilken, C.; Gutknecht, J. Effects of anthropogenic disturbances on soil microbial communities in oak forests persist for more than 100 years. Soil Biol. Biochem. 2014, 70, 79–87. [Google Scholar] [CrossRef]
- Frostegård, Å.; Tunlid, A.; Bååth, E. Use and misuse of PLFA measurements in soils. Soil Biol. Biochem. 2011, 43, 1621–1625. [Google Scholar] [CrossRef]
- McClaugherty, C.A.; Pastor, J.; Aber, J.D.; Melillo, J.M. Forest litter decomposition in relation to soil nitrogen dynamics and litter quality. Ecology 1985, 66, 266–275. [Google Scholar] [CrossRef]
- Melillo, J.M.; Aber, J.D.; Muratore, J.F. Nitrogen and lignin control of hardwood leaf litter decomposition dynamics. Ecology 1982, 63, 621–626. [Google Scholar] [CrossRef]
- Krashevska, V.; Sandmann, D.; Maraun, M.; Scheu, S. Consequences of exclusion of precipitation on microorganisms and microbial consumers in montane tropical rainforests. Oecologia 2012, 170, 1067–1076. [Google Scholar] [CrossRef] [PubMed]
- Hättenschwiler, S.; Tiunov, A.V.; Scheu, S. Biodiversity and litter decomposition in terrestrial ecosystems. Annu. Rev. Ecol. Evol. Syst. 2005, 36, 191–218. [Google Scholar] [CrossRef]
- Aber, J.D.; Melillo, J.M.; McClaugherty, C.A. Predicting long-term patterns of mass loss, nitrogen dynamics, and soil organic matter formation from initial fine litter chemistry in temperate forest ecosystems. Can. J. Bot. 1990, 68, 2201–2208. [Google Scholar] [CrossRef]
- Berg, B. Litter decomposition and organic matter turnover in northern forest soils. For. Ecol. Manag. 2000, 133, 13–22. [Google Scholar] [CrossRef]
- Hobbie, S.E.; Eddy, W.C.; Buyarski, C.R.; Adair, E.C.; Ogdahl, M.L.; Weisenhorn, P. Response of decomposing litter and its microbial community to multiple forms of nitrogen enrichment. Ecol. Monogr. 2012, 82, 389–405. [Google Scholar] [CrossRef]
- Enriquez, S.; Duarte, C.M.; Sand-Jensen, K. Patterns in decomposition rates among photosynthetic organisms: The importance of detritus C: N: P content. Oecologia 1993, 94, 457–471. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Hirata, E. Decomposition patterns of leaf litter of seven common canopy species in a subtropical forest: N and P dynamics. Plant Soil 2005, 273, 279–289. [Google Scholar] [CrossRef]
- Eriksson, K.E.L.; Blanchette, R.; Ander, P. Microbial and Enzymatic Degradation of Wood and Wood Components; Springer Science & Business Media: Berlin, Germany, 2012. [Google Scholar]
- Berg, B.; De Santo, A.V.; Rutigliano, F.A.; Fierro, A.; Ekbohm, G. Limit values for plant litter decomposing in two contrasting soils-influence of litter elemental composition. Acta Oecol. 2003, 24, 295–302. [Google Scholar] [CrossRef]
- Hobbie, S.E.; Reich, P.B.; Oleksyn, J.; Ogdahl, M.; Zytkowiak, R.; Hale, C.; Karolewski, P. Tree species effects on decomposition and forest floor dynamics in a common garden. Ecology 2006, 87, 2288–2297. [Google Scholar] [CrossRef]
- Reich, P.B.; Oleksyn, J.; Modrzynski, J.; Mrozinski, P.; Hobbie, S.E.; Eissenstat, D.M.; Chorover, J.; Chadwick, O.A.; Hale, C.M.; Tjoelker, M.G. Linking litter calcium, earthworms and soil properties: A common garden test with 14 tree species. Ecol. Lett. 2005, 8, 811–818. [Google Scholar] [CrossRef]
- Pérez, C.A.; Hedin, L.O.; Armesto, J.J. Nitrogen mineralization in two unpolluted old-growth forests of contrasting biodiversity and dynamics. Ecosystems 1998, 1, 361–373. [Google Scholar]
- Staff, H.; Berg, B. Accumulation and release of plant nutrients in decomposing Scots pine needle litter. II. Long-term decomposition in a Scots pine forest. Can. J. Bot. 1982, 60, 1561–1568. [Google Scholar] [CrossRef]
- Osono, T.; Azuma, J.; Hirose, D. Plant species effect on the decomposition and chemical changes of leaf litter in grassland and pine and oak forest soils. Plant Soil 2014, 376, 411–421. [Google Scholar] [CrossRef]
- Lisanework, N.; Michelsen, A. Litterfall and nutrient release by decomposition in three plantations compared with a natural forest in the Ethiopian highland. For. Ecol. Manag. 1994, 65, 149–164. [Google Scholar] [CrossRef]
- O’Connell, A. Nutrient dynamics in decomposing litter in karri (Eucalyptus diversicolor F. Muell.) forests of south-western Australia. J. Ecol. 1988, 76, 1186–1203. [Google Scholar] [CrossRef]
- Sharma, E.; Ambasht, R.S. Litterfall, decomposition and nutrient release in an age sequence of Alnus nepalensis plantation stands in the eastern Himalaya. J. Ecol. 1987, 75, 997–1010. [Google Scholar] [CrossRef]
- Berg, B.; Steffen, K.T.; McClaugherty, C. Litter decomposition rate is dependent on litter Mn concentrations. Biogeochemistry 2007, 82, 29–39. [Google Scholar] [CrossRef]
- Strickland, M.; Lauber, C.; Fierer, N.; Bradford, M.A. Testing the functional signifcance of microbial community composition. Ecology 2009, 90, 441–451. [Google Scholar] [CrossRef] [PubMed]
- Krumins, J.A.; Long, Z.T.; Steiner, C.F.; Morin, P.J. Indirect effects of food web diversity and productivity on bacterial community function and composition. Funct. Ecol. 2006, 20, 514–521. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis; Academic Press: Cambridge, MA, USA, 1996. [Google Scholar]
- Went, F.W.; Stark, N. Mycorrhiza. BioScience 1968, 18, 1035–1039. [Google Scholar] [CrossRef]
- Hodge, A.; Campbell, C.D.; Fitter, A.H. An arbuscular mycorrhizal fungus accelerates decomposition and acquires nitrogen directly from organic material. Nature 2001, 413, 297–299. [Google Scholar] [CrossRef] [PubMed]
- Pritsch, K.; Garbaye, J. Enzyme secretion by ECM fungi and exploitation of mineral nutrients from soil organic matter. Ann. For. Sci. 2011, 68, 25–32. [Google Scholar] [CrossRef]
- Norris, C.E.; Quideau, S.A.; Macey, D.E. Processing of 13C glucose in mineral soil from aspen, spruce and novel ecosystems in the Athabasca oil sands region. Appl. Soil Ecol. 2013, 71, 24–32. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical Property (g kg−1) | Leaf | Fine Root | Litter Type | Plantation Type | ||||
|---|---|---|---|---|---|---|---|---|
| Erythrophleum fordii | Mixed Forest | Pinus massoniana | Erythrophleum fordii | Mixed Forest | Pinus massoniana | |||
| C | 483.6 ± 6.89 | 508.0 ± 5.06 | 509.7 ± 5.90 | 602.7 ± 8.48 | 532.5 ± 9.44 | 487.4 ± 7.55 | p < 0.05 | p = 0.130 |
| N | 34.2 ± 0.16 | 19.2 ± 0.33 | 13.9 ± 0.22 | 8.38 ± 0.52 | 7.62 ± 0.17 | 7.44 ± 0.05 | p < 0.001 | p < 0.01 |
| P | 0.49 ± 0.01 | 0.61 ± 0.01 | 0.63 ± 0.03 | 0.28 ± 0.02 | 0.45 ± 0.01 | 0.52 ± 0.00 | p < 0.001 | p < 0.001 |
| K | 4.73 ± 0.11 | 3.58 ± 0.02 | 3.25 ± 0.21 | 1.07 ± 0.04 | 2.78 ± 0.07 | 3.65 ± 0.08 | p < 0.05 | p = 0.604 |
| Na | 1.47 ± 0.04 | 0.82 ± 0.07 | 0.78 ± 0.04 | 0.21 ± 0.01 | 0.40 ± 0.01 | 0.41 ± 0.04 | p < 0.001 | p = 0.182 |
| Ca | 11.2 ± 0.06 | 7.66 ± 0.21 | 6.72 ± 0.33 | 3.33 ± 0.12 | 2.58 ± 0.12 | 2.23 ± 0.07 | p < 0.001 | p < 0.001 |
| Mg | 5.15 ± 0.08 | 2.78 ± 0.04 | 2.03 ± 0.10 | 0.37 ± 0.02 | 1.25 ± 0.02 | 1.71 ± 0.04 | p < 0.01 | p = 0.331 |
| Mn | 0.18 ± 0.00 | 0.84 ± 0.02 | 1.00 ± 0.04 | 0.05 ± 0.00 | 0.11 ± 0.00 | 0.15 ± 0.00 | p < 0.001 | p < 0.01 |
| Lignin | 218.3 ± 0.34 | 250.1 ± 5.21 | 262.4 ± 1.26 | 247.5 ± 0.02 | 273.6 ± 0.02 | 305.2 ± 0.04 | p < 0.001 | p < 0.001 |
| Cellulose | 172.8 ± 5.48 | 133.7 ± 5.20 | 125.7 ± 9.39 | 140.7 ± 1.35 | 140.8 ± 2.27 | 135.4 ± 5.94 | p = 0.418 | p < 0.05 |
| C/N | 14.1 ± 0.21 | 26.5 ± 0.71 | 36.7 ± 0.23 | 72.6 ± 5.55 | 69.9 ± 0.49 | 65.5 ± 1.45 | p < 0.001 | p = 0.138 |
| C/P | 987.7 ± 14.9 | 834.0 ± 18.4 | 808.6 ± 40.5 | 2199 ± 138 | 1182 ± 36.6 | 940.2 ± 20.8 | p < 0.01 | p < 0.01 |
| N/P | 69.9 ± 1.75 | 31.5 ± 0.15 | 22.1 ± 1.25 | 30.9 ± 3.98 | 16.9 ± 0.51 | 14.3 ± 0.13 | p < 0.001 | p < 0.001 |
| Lignin/N | 6.38 ± 0.03 | 13.0 ± 0.21 | 18.9 ± 0.35 | 29.8 ± 1.84 | 35.9 ± 0.75 | 41.0 ± 0.37 | p < 0.001 | p < 0.001 |
| Litter Type | Asymptotic Function | |||
|---|---|---|---|---|
| m | Std. Err. | kA (d−1) | R2adj | |
| Erythrophleum fordii | ||||
| Leaf | 97.96 | 4.33 | 0.24 | 0.97 |
| Fine root | 95.65 | 5.37 | 0.17 | 0.92 |
| Mixed forest | ||||
| Leaf | 99.21 | 3.51 | 0.20 | 0.97 |
| Fine root | 96.74 | 3.85 | 0.14 | 0.94 |
| Pinus massoniana | ||||
| Leaf | 99.60 | 2.85 | 0.19 | 0.98 |
| Fine root | 96.27 | 3.64 | 0.13 | 0.94 |
| Remaining Mass (%) | Leaf | Fine Root | ||||
|---|---|---|---|---|---|---|
| Erythrophleum fordii | Mixed Forest | Pinus massoniana | Erythrophleum fordii | Mixed Forest | Pinus massoniana | |
| Dry mass | 27.1 ± 1.30 | 32.7 ± 0.75 | 35.1 ± 1.10 | 39.5 ± 1.01 | 46.3 ± 1.82 | 48.3 ± 1.39 |
| Lignin | 49.0 ± 1.33 | 56.7 ± 4.08 | 60.8 ± 1.65 | 48.9 ± 0.58 | 58.7 ± 2.79 | 63.5 ± 3.40 |
| Cellulose | 25.8 ± 2.20 | 29.0 ± 0.78 | 31.4 ± 3.32 | 37.5 ± 1.16 | 42.9 ± 2.44 | 44.0 ± 2.90 |
| C | 13.9 ± 0.79 | 19.5 ± 0.83 | 25.6 ± 0.78 | 24.5 ± 2.19 | 29.5 ± 1.00 | 29.3 ± 1.07 |
| N | 11.7 ± 0.21 | 28.7 ± 2.21 | 45.3 ± 2.41 | 53.1 ± 3.79 | 75.1 ± 6.63 | 85.0 ± 2.34 |
| P | 29.1 ± 0.85 | 40.4 ± 0.73 | 50.7 ± 3.11 | 46.9 ± 1.96 | 53.4 ± 2.51 | 58.5 ± 37.4 |
| K | 3.87 ± 0.12 | 8.04 ± 0.40 | 9.34 ± 0.91 | 1.29 ± 3.05 | 7.71 ± 040 | 7.93 ± 0.53 |
| Na | 1.94 ± 0.02 | 4.35 ± 0.46 | 5.36 ± 0.23 | 21.4 ± 2.49 | 12.7 ± 0.11 | 12.5 ± 0.84 |
| Ca | 26.3 ± 0.39 | 34.3 ± 2.49 | 36.6 ± 2.45 | 40.4 ± 2.95 | 49.0 ± 1.93 | 49.9 ± 1.16 |
| Mg | 30.1 ± 1.60 | 42.2 ± 3.53 | 38.1 ± 0.37 | 75.7 ± 2.15 | 61.5 ± 1.64 | 48.7 ± 2.34 |
| Mn | 59.4 ± 0.84 | 66.5 ± 3.95 | 94.5 ± 3.88 | 61.4 ± 3.46 | 74.7 ± 3.92 | 53.7 ± 1.76 |
| Litter Decay | Factor | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Fixed Effect | Random Effect | ||||||||
| Plantation Type | Litter Type | Time | |||||||
| Num. d.f. | F-Value | p-Value | Num. d.f. | F-Value | p-Value | Estimates | Std. Err. | Wald Z | |
| Dry mass | 2 | 62.966 | 0.013 | 1 | 332.844 | 0.002 | 72.008 | 102.682 | 0.701 |
| Lignin | 2 | 28.687 | <0.001 | 1 | 2.443 | 0.144 | 57.582 | 81.578 | 0.706 |
| Cellulose | 2 | 569.811 | 0.001 | 1 | 3379.03 | <0.001 | 40.041 | 59.682 | 0.671 |
| C | 2 | 19.753 | 0.007 | 1 | 38.391 | 0.003 | 123.823 | 175.675 | 0.705 |
| N | 2 | 146.613 | <0.001 | 1 | 593.284 | <0.001 | 65.290 | 94.215 | 0.693 |
| P | 2 | 30.759 | 0.004 | 1 | 0.476 | 0.529 | 175.069 | 248.144 | 0.706 |
| K | 2 | 0.283 | 0.756 | 1 | 1.461 | 0.238 | 0.292 | 0.630 | 0.464 |
| Na | 2 | 1.343 | 0.285 | 1 | 85.441 | <0.001 | 4.389 | 6.747 | 0.650 |
| Ca | 2 | 69.423 | <0.001 | 1 | 230.415 | <0.001 | 88.551 | 125.928 | 0.703 |
| Mg | 2 | 0.241 | 0.791 | 1 | 18.312 | 0.002 | 89.507 | 127.637 | 0.701 |
| Mn | 2 | 11.660 | <0.001 | 1 | 33.851 | <0.001 | 41.871 | 60.447 | 0.693 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, D.; Cheng, R.; Shi, Z.; Wang, W. Decomposition of Leaves and Fine Roots in Three Subtropical Plantations in China Affected by Litter Substrate Quality and Soil Microbial Community. Forests 2017, 8, 412. https://doi.org/10.3390/f8110412
Luo D, Cheng R, Shi Z, Wang W. Decomposition of Leaves and Fine Roots in Three Subtropical Plantations in China Affected by Litter Substrate Quality and Soil Microbial Community. Forests. 2017; 8(11):412. https://doi.org/10.3390/f8110412
Chicago/Turabian StyleLuo, Da, Ruimei Cheng, Zuomin Shi, and Weixia Wang. 2017. "Decomposition of Leaves and Fine Roots in Three Subtropical Plantations in China Affected by Litter Substrate Quality and Soil Microbial Community" Forests 8, no. 11: 412. https://doi.org/10.3390/f8110412
APA StyleLuo, D., Cheng, R., Shi, Z., & Wang, W. (2017). Decomposition of Leaves and Fine Roots in Three Subtropical Plantations in China Affected by Litter Substrate Quality and Soil Microbial Community. Forests, 8(11), 412. https://doi.org/10.3390/f8110412