Spatial Upscaling of Soil Respiration under a Complex Canopy Structure in an Old‐Growth Deciduous Forest, Central Japan

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
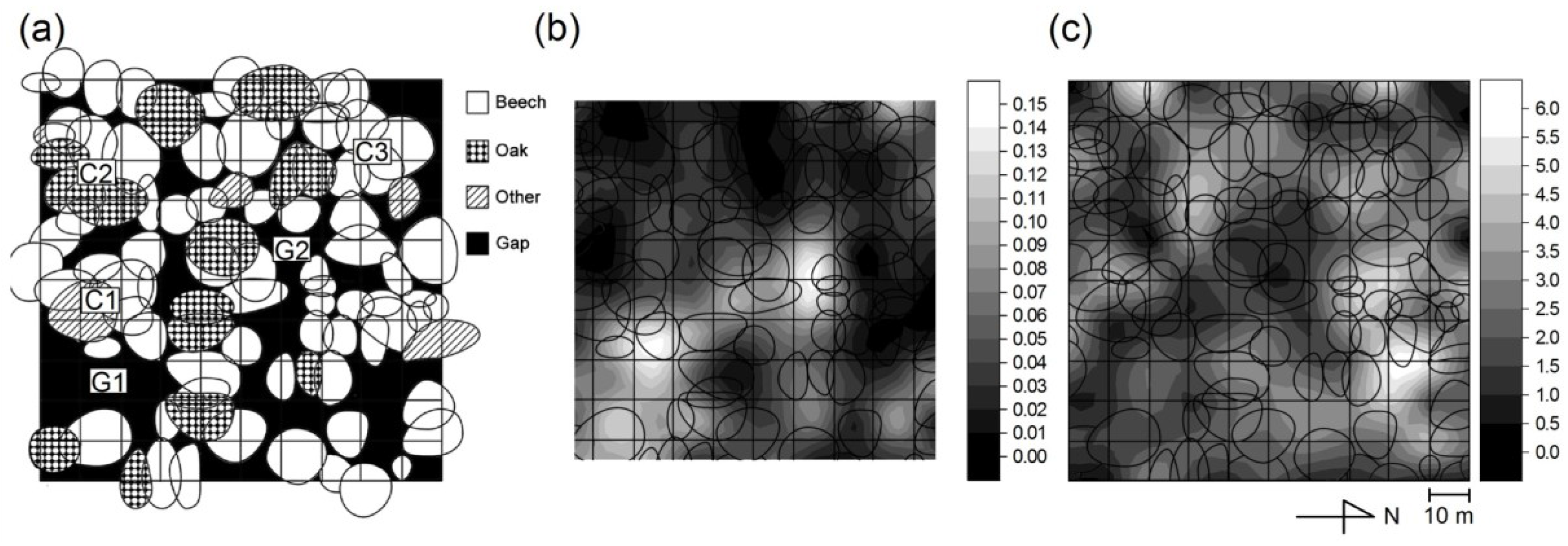
2.1. Study Site
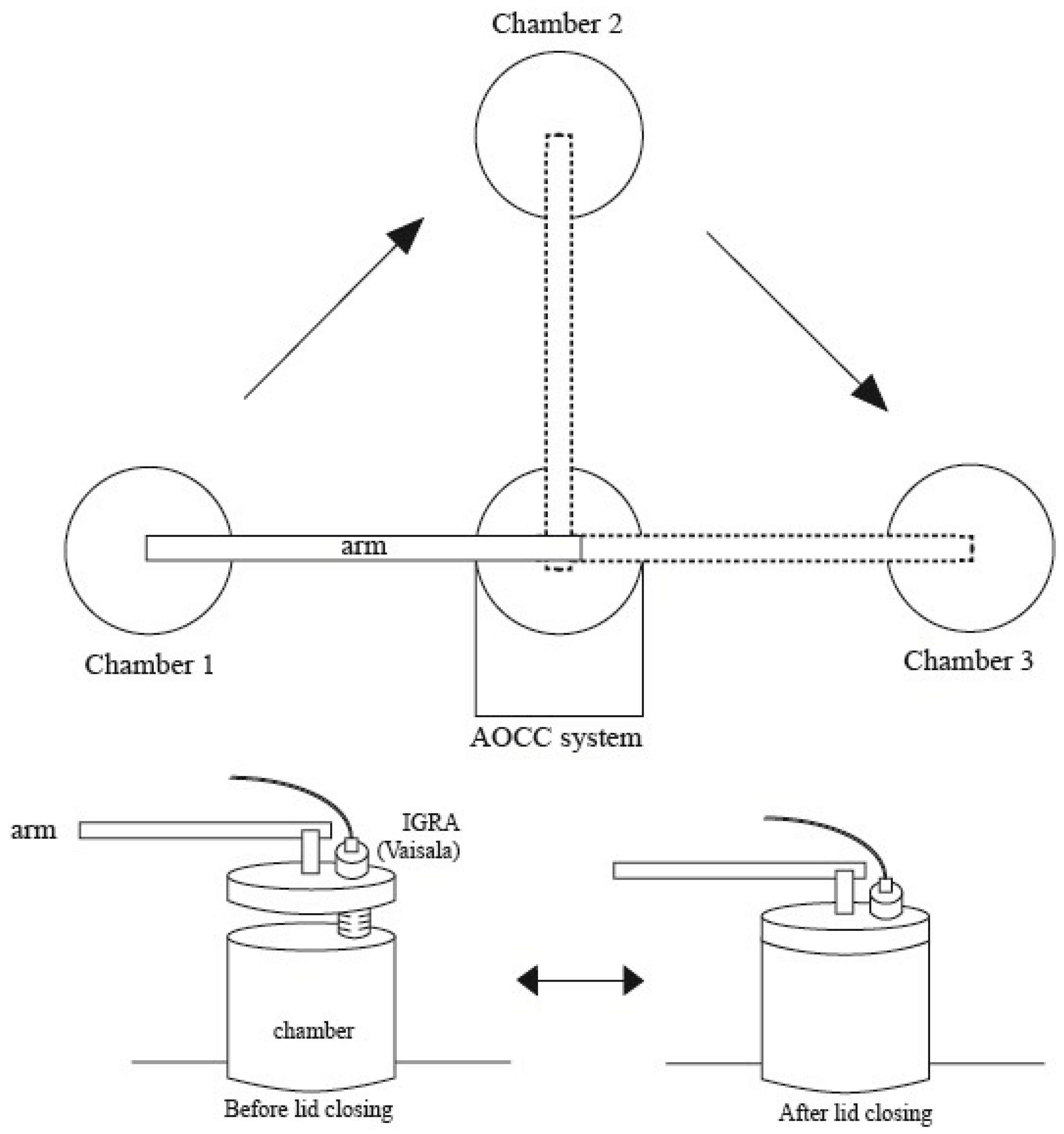
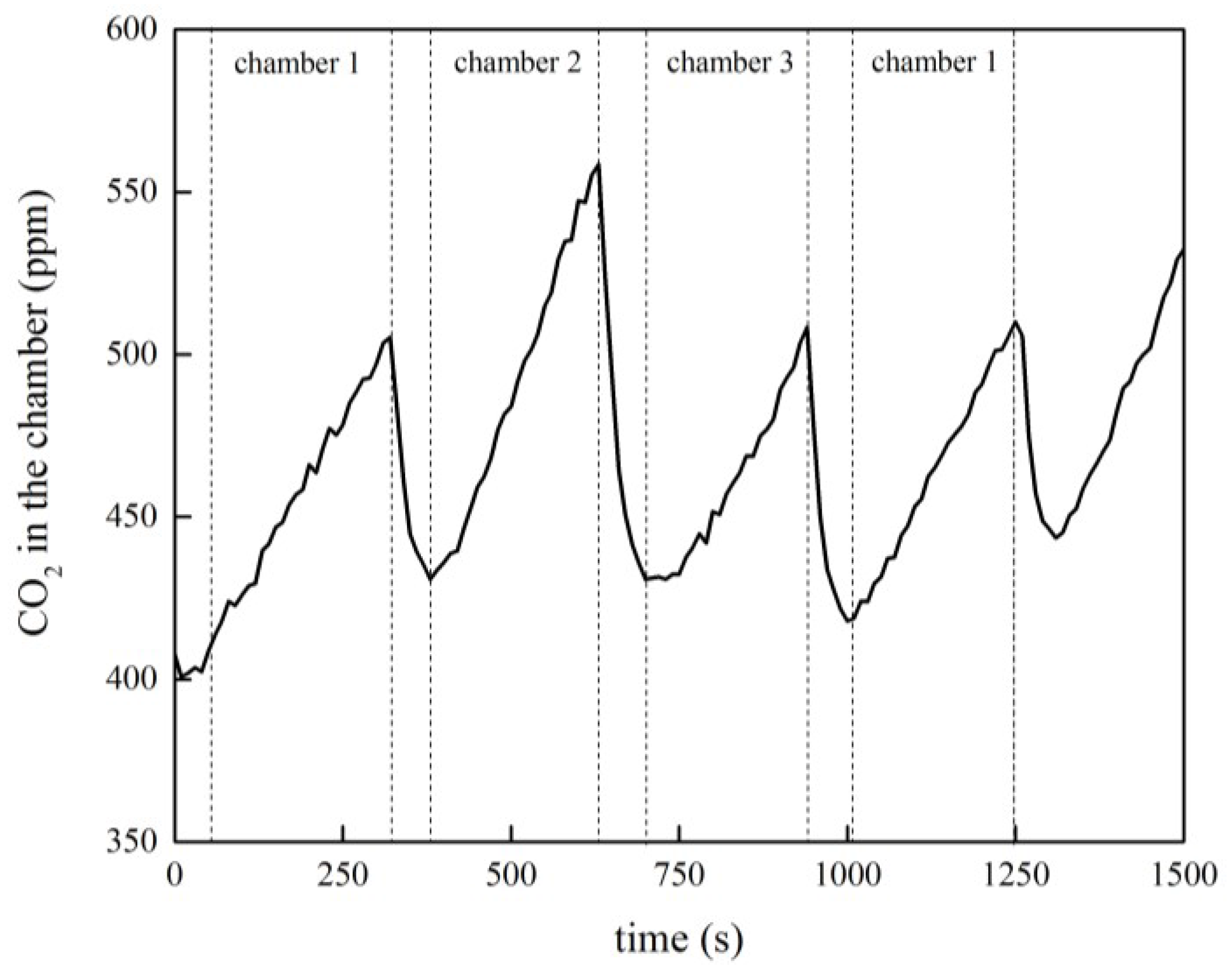
2.2. Measurement of the Soil CO2 Efflux
2.3. Measurement of Soil Temperature and Water Content
2.4. Data Analysis
3. Results
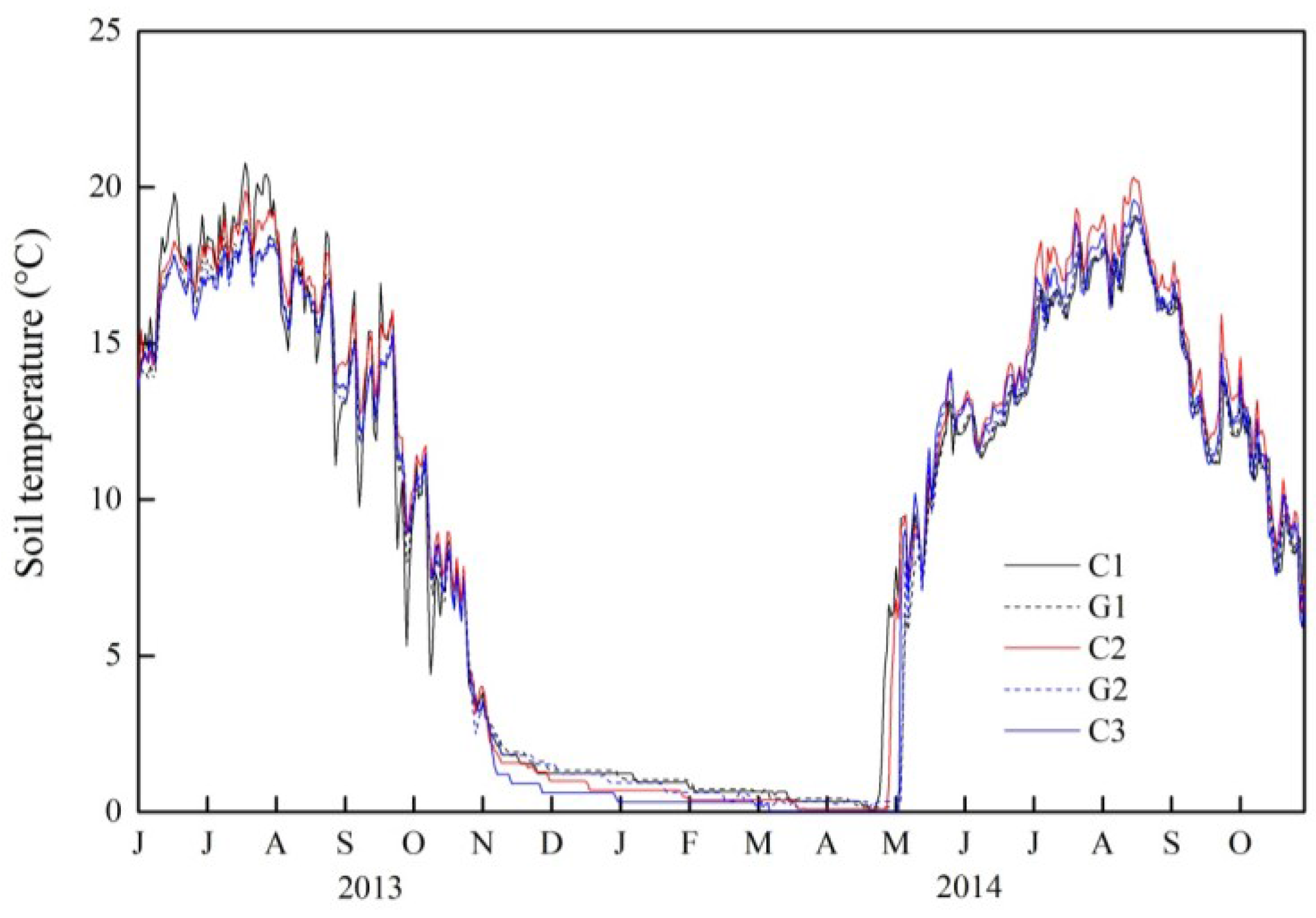
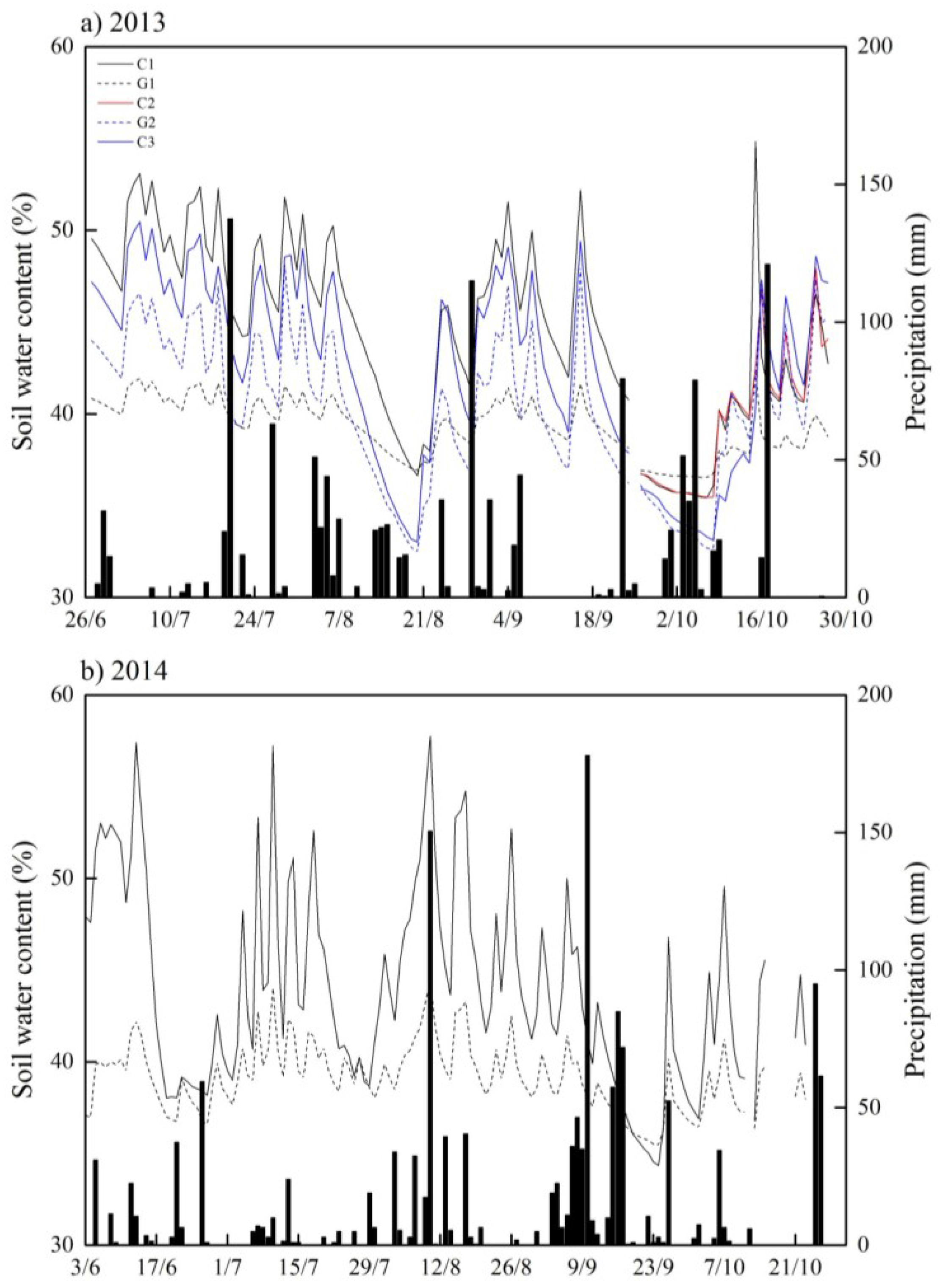
3.1. Temporal Changes in Soil Temperature and Soil Water Content
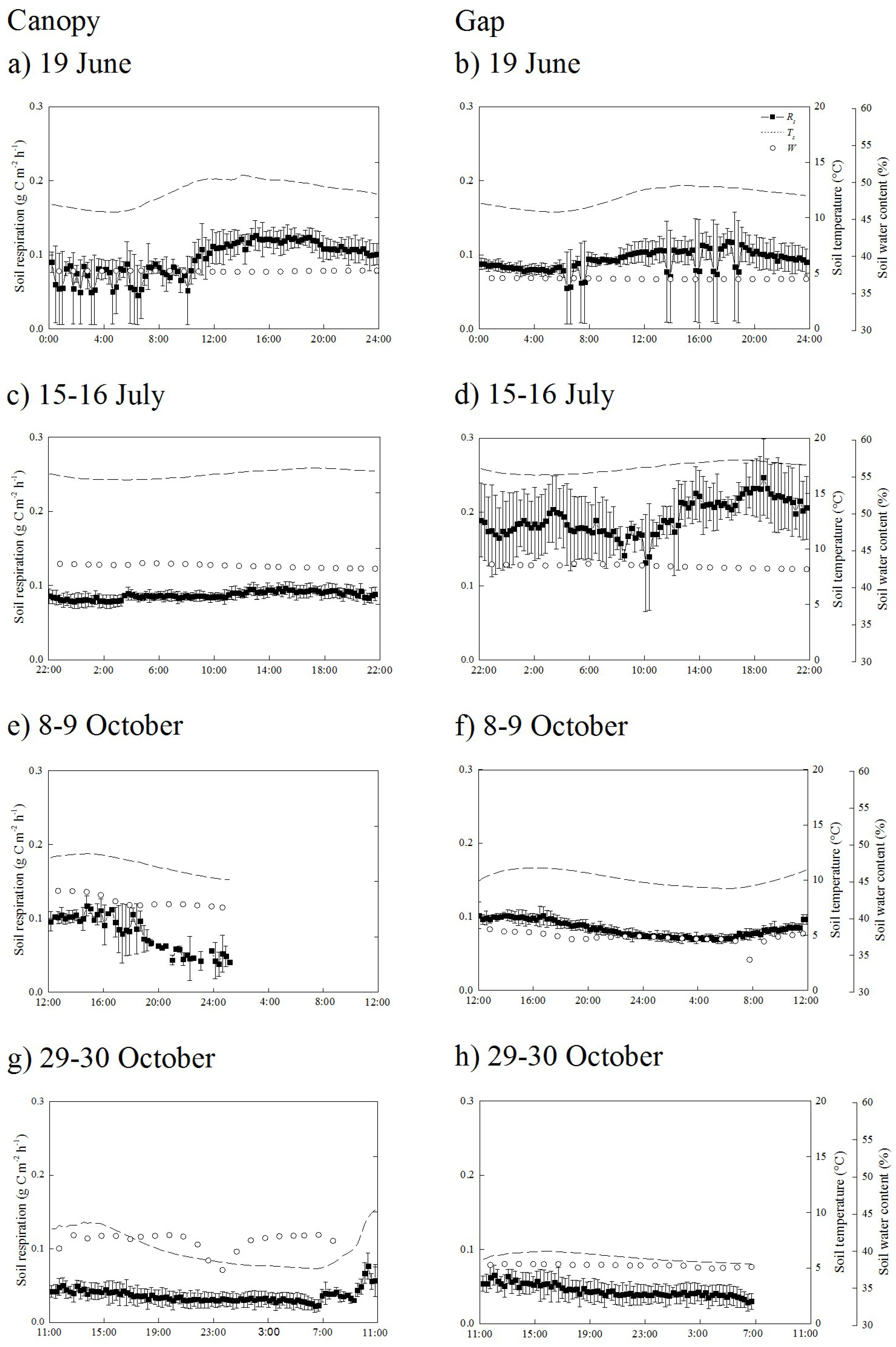
3.2. Diurnal Changes in Soil Temperature, Soil Water Content, and Soil Respiration
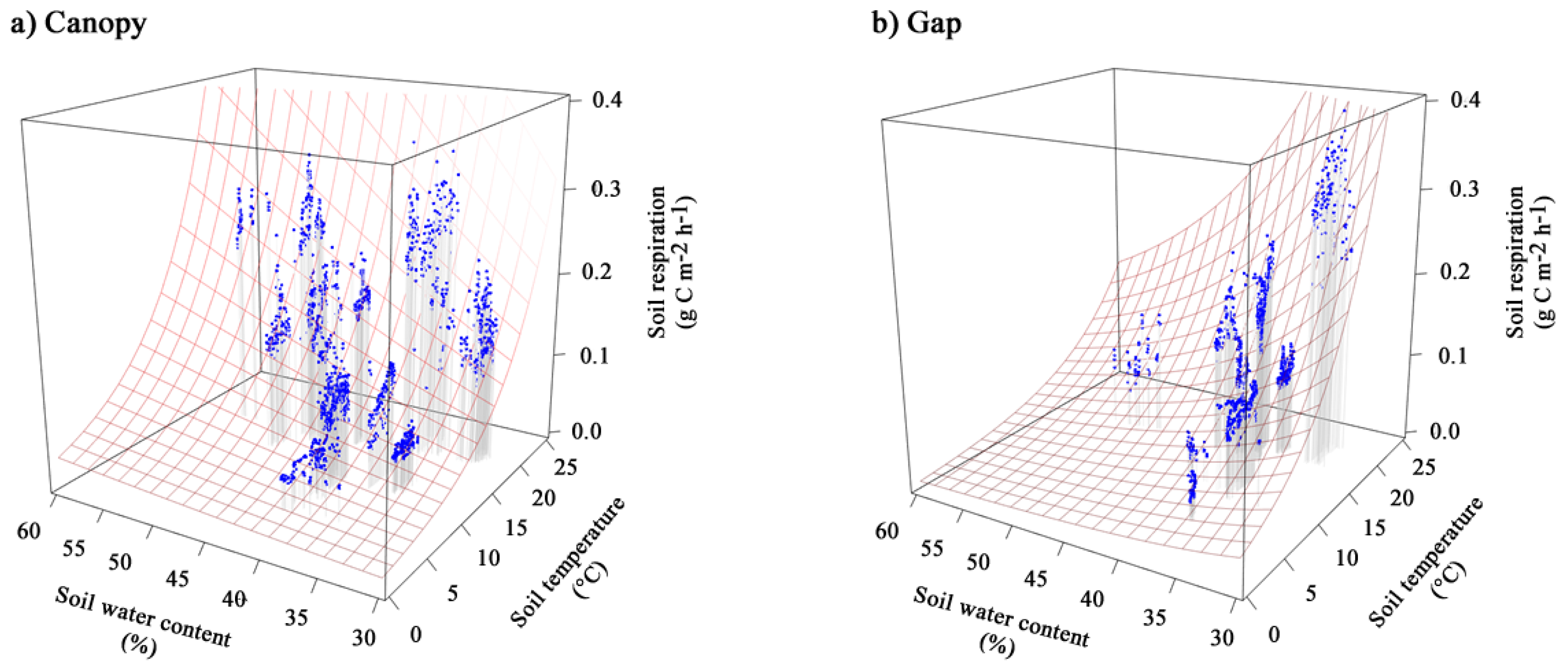
3.3. Effect of Soil Temperature and Soil Water Content on Soil Respiration
3.4. Estimation and Upscaling of Rs to the Whole Site
4. Discussion
4.1. Effect of Soil Temperature and Soil Water Content on Soil Respiration
4.2. Estimation of Soil Respiration in the Old-Growth Forest
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Liu, Q.; Edwards, N.T.; Post, W.M.; Gu, L.; Ledford, J.; Lenhart, S. Temperature-independent diel variation in soil respiration observed from a temperate deciduous forest. Glob. Chang. Biol. 2006, 12, 2136–2145. [Google Scholar] [CrossRef]
- Hashimoto, T.; Miura, S.; Ishizuka, S. Temperature controls temporal variation in soil CO2 efflux in a secondary beech forest in Appi Highlands, Japan. J. For. Res. 2009, 14, 44–50. [Google Scholar] [CrossRef]
- Joo, S.J.; Park, S.U.; Park, M.S.; Lee, C.S. Estimation of soil respiration using automated chamber systems in an oak (Quercus mongolica) forest at the Nam-San site in Seoul, Korea. Sci. Total Environ. 2012, 416, 400–409. [Google Scholar] [CrossRef] [PubMed]
- Kishimoto-Mo, A.W.; Yonemura, S.; Uchida, M.; Kondo, M.; Murayama, S.; Koizumi, H. Contribution of soil moisture to seasonal and annual variations of soil CO2 efflux in a humid cool-temperate oak-birch forest in central Japan. Ecol. Res. 2015, 30, 311–325. [Google Scholar] [CrossRef]
- Xu, M.; Qi, Y. Soil-surface CO2 efflux and its spatial and temporal variations in a young ponderosa pine plantation in Northern California. Glob. Chang. Biol. 2001, 7, 667–677. [Google Scholar] [CrossRef]
- Mo, W.; Lee, M.; Uchida, M.; Inatomi, M.; Saigusa, N.; Mariko, S.; Koizumi, H. Seasonal and annual variations in soil respiration in a cool-temperate deciduous broad-leaved forest in Japan. Agric. For. Meteorol. 2005, 134, 81–94. [Google Scholar] [CrossRef]
- Ohtsuka, T.; Shizu, Y.; Nishiwaki, A.; Yashiro, Y.; Koizumi, H. Carbon cycling and net ecosystem production at an early stage of secondary succession in an abandoned coppice forest. J. Plant Res. 2010, 123, 393–401. [Google Scholar] [CrossRef] [PubMed]
- Inoue, T.; Nagai, S.; Inoue, S.; Ozaki, M.; Sakai, S.; Muraoka, H.; Koizumi, H. Seasonal variability of soil respiration in multiple ecosystems under the same physical-geographical environmental conditions in central Japan. For. Sci. Technol. 2012, 8, 52–60. [Google Scholar] [CrossRef]
- Davidson, E.A.; Belk, E.; Boone, R. Soil water content and temperature as independent of confounded factors controlling soil respiration in a temperate mixed hardwood forest. Glob. Chang. Biol. 1998, 4, 217–227. [Google Scholar] [CrossRef]
- Wang, Y.; Hao, Y.; Cui, X.Y.; Zhao, H.; Xu, C.; Zhou, X.; Xu, Z. Responses of soil respiration and its components to drought stress. J. Soil Sediments 2013, 14, 99–109. [Google Scholar] [CrossRef]
- Irvine, J.; Law, B.E. Contrasting soil respiration in young and old-growth ponderosa pine forests. Glob. Chang. Biol. 2002, 8, 1183–1194. [Google Scholar] [CrossRef]
- Edwards, N.T.; Riggs, J.S. Automated monitoring of soil respiration: A moving chamber design. Soil Sci. Soc. Am. J. 2003, 67, 1266–1271. [Google Scholar] [CrossRef]
- Liang, N.; Inoue, G.; Fujinuma, Y. A multichannel automated chamber system for continuous measurement of forest soil CO2 efflux. Tree Physiol. 2003, 23, 825–832. [Google Scholar] [CrossRef] [PubMed]
- Suh, S.U.; Chun, Y.M.; Chae, N.; Kim, J.; Lim, J.H.; Yokozawa, M.; Lee, M.S.; Lee, J.S. A chamber system with automatic opening and closing for continuously measuring soil respiration based on an open-flow dynamic method. Ecol. Res. 2006, 21, 405–414. [Google Scholar] [CrossRef]
- Nakashizuka, T.; Numata, M. Regeneration process of climax beech forests: I. Structure of a beech forest with the undergrowth of Sasa. Jpn. J. Ecol. 1982, 32, 57–67. [Google Scholar]
- Tanouchi, H.; Yamamoto, S. Structure and regeneration of canopy species in an old-growth evergreen broad-leaved forest in Aya district, Southwestern Japan. Vegetation 1995, 117, 51–60. [Google Scholar] [CrossRef]
- Manabe, T.; Nishimura, N.; Miura, M.; Yamamoto, S. Population structure and spatial patterns for trees in a temperate old-growth evergreen broad-leaved forest in Japan. Plant Ecol. 2000, 151, 181–197. [Google Scholar] [CrossRef]
- Miura, M.; Manabe, T.; Nishimura, N.; Yamamoto, S. Forest canopy and community dynamics in a temperate old-growth evergreen broad-leaved forest, south-western Japan: A 7-year study of a 4-ha plot. J. Ecol. 2001, 89, 841–849. [Google Scholar] [CrossRef]
- Matsushita, M.; Hoshino, D.; Yamamoto, S.; Nishimura, N. Twenty-three years and dynamics in an old-growth Chamaecyparis forest in central Japan. J. For. Res. 2014, 19, 134–142. [Google Scholar] [CrossRef]
- Suchewaboripont, V.; Iimura, Y.; Yoshitake, S.; Kato, S.; Komiyama, A.; Ohtsuka, T. Change in biomass of an old-growth beech-oak forest of Mt. Hakusan over a 17-year period. Jpn. J. For. Environ. 2015, 57, 33–42. [Google Scholar]
- Hardiman, B.S.; Bohrer, G.; Gough, C.M.; Vogel, C.S.; Curtis, P.S. The role of canopy structural complexity in wood net primary production of a maturing northern deciduous forest. Ecology 2011, 92, 1818–1827. [Google Scholar] [CrossRef] [PubMed]
- Hardiman, B.S.; Gough, C.M.; Halperin, A.; Hofmeister, K.L.; Nave, L.E.; Bohrer, G.; Curtis, P.S. Maintaining high rates of carbon storage in old forests: A mechanism lining canopy structure to forest function. For. Ecol. Manag. 2013, 298, 111–119. [Google Scholar] [CrossRef]
- Hojjati, S.M.; Lamersdorf, N.P. Effect of canopy composition on soil CO2 emission in a mixed spruce-beech forest at Solling, Central Germany. J. For. Res. 2010, 21, 461–464. [Google Scholar] [CrossRef]
- Suchewaboripont, V.; Ando, M.; Iimura, Y.; Yoshitake, S.; Ohtsuka, T. The effect of canopy structure on soil respiration in an old-growth beech-oak forest in central Japan. Ecol. Res. 2015, 30, 867–877. [Google Scholar] [CrossRef]
- Tang, J.; Baldocchi, D.D. Spatial-temporal variation in soil respiration in an oak-grass savanna ecosystem in California and its partitioning into autotrophic and heterotrophic components. Biogeochemistry 2005, 73, 183–207. [Google Scholar] [CrossRef]
- Geological Survey of Japan, AIST (Ed.) Catalog of Eruptive Events during the Last 10,000 Years in Japan, Version 2.2. Available online: https://gbank.gsj.jp/volcano/eruption/index.html (accessed on 29 December 2016).
- Kato, S.; Komiyama, A. Distribution patterns of understory trees and diffuse light under the canopy of a beech forest. Jpn. J. Ecol. 1999, 49, 1–10. [Google Scholar]
- Runkle, J.R. Gap regeneration in some old-growth forests of the eastern United States. Ecology 1981, 62, 1041–1051. [Google Scholar] [CrossRef]
- Hirota, M.; Zhang, P.; Gu, S.; Shen, H.; Kuriyama, T.; Li, Y.; Tang, Y. Small-scale variation in ecosystem CO2 fluxes in an alpine meadow depends on plant biomass and species richness. J. Plant Res. 2010, 123, 531–541. [Google Scholar] [CrossRef] [PubMed]
- Abbas, F.; Fares, A.; Fares, S. Field calibrations of soil moisture sensors in a forested watershed. Sensors 2011, 11, 6354–6369. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.S.; Chen, G.S.; Guo, J.F.; Xie, J.S.; Wang, X.G. Soil respiration and carbon balance in a subtropical native forest and two managed plantations. Plant Ecol. 2007, 193, 71–84. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. lme4: Linear Mixed-Effects Models Using Eigen and S4. R Package Version 1.1-7. 2014. Available online: http://CRAN.R-project.org/package=lme4 (accessed on 22 April 2016).
- Epron, D.; Farque, L.; Lucot, É.; Badot, P.M. Soil CO2 efflux in a beech forest: Dependence on soil temperature and soil water content. Ann. For. Sci. 1999, 56, 221–226. [Google Scholar] [CrossRef]
- Qi, Y.; Xu, M. Separating the effects of moisture and temperature on soil CO2 efflux in a coniferous forest in the Sierra Nevada mountains. Plant Soil 2001, 237, 15–23. [Google Scholar] [CrossRef]
- Søe, A.R.B.; Buchmann, N. Spatial and temporal variations in soil respiration in relation to stand structure and soil parameters in an unmanaged beech forest. Tree Physiol. 2005, 25, 1427–1436. [Google Scholar] [CrossRef] [PubMed]
- Knohl, A.; Søe, A.R.B.; Kutsch, W.L.; Göckede, M.; Buchmann, N. Representative estimates of soil and ecosystem respiration in an old beech forest. Plant Soil 2008, 302, 189–202. [Google Scholar] [CrossRef]
- Tomotsune, M.; Masuda, R.; Yoshitake, S.; Anzai, T.; Koizumi, H. Seasonal and inter-annual variations in contribution ratio of heterotrophic respiration to soil respiration in a cool-temperate deciduous forest. J. Geogr. 2013, 122, 745–754. [Google Scholar] [CrossRef]
- Maier, C.A.; Kress, L. Soil CO2 evolution and root respiration in 11 year-old loblolly pine (Pinus taeda) plantations as affected by moisture and nutrient. Can. J. For. Res. 2000, 30, 347–359. [Google Scholar] [CrossRef]
- Dörr, H.; Münnich, K.O. Annual variation in soil respiration in selected areas of the temperate zone. Tellus 1987, 39, 114–121. [Google Scholar] [CrossRef]
- Boone, R.D.; Nadelhoffer, K.J.; Canary, J.D.; Kaye, J.P. Roots exert a strong influence on the temperature sensitivity of soil respiration. Nature 1998, 396, 570–572. [Google Scholar] [CrossRef]
- Liski, J.; Ilvesniemi, H.; Mäkelä, A.; Westman, C.J. CO2 emissions from soil in response to climatic warming are overestimated: The decomposition of old soil organic matter is tolerant of temperature. Ambio 1999, 28, 171–174. [Google Scholar]
- Liang, N.; Hirano, T.; Zheng, Z.-M.; Tang, J.; Fujinuma, Y. Soil CO2 efflux of a larch forest in Northern Japan. Biogeosciences 2010, 7, 3447–3457. [Google Scholar] [CrossRef]
- Vesterdal, L.; Elberling, B.; Christiansen, J.R.; Callesen, I.; Schmidt, I.K. Soil respiration and rates of soil carbon turnover differ among six common European tree species. For. Ecol. Manag. 2012, 264, 185–196. [Google Scholar] [CrossRef]
- Luan, J.; Liu, S.; Wang, J.; Zhu, X. Factors affecting spatial variation of annual apparent Q10 of soil respiration in two warm temperate forests. PLoS ONE 2013, 8, e64167. [Google Scholar] [CrossRef] [PubMed]
- Jia, S.; Akiyama, T.; Mo, W.; Inatomi, M.; Koizumi, H. Temporal and spatial variability of soil respiration in a cool-temperate broad-leaved forest 1. Measurement of spatial variance and factor analysis. Jpn. J. Ecol. 2003, 53, 13–22. [Google Scholar]
- Scott-Denton, L.E.; Sparks, K.L.; Monson, R.K. Spatial and temporal controls of soil respiration rate in a high-elevation, subalpine forest. Soil Biol. Biochem. 2003, 35, 525–534. [Google Scholar] [CrossRef]
- Saiz, G.; Green, C.; Butterbach-Bahl, K.; Kiese, R.; Avitabile, V.; Farrell, E.P. Seasonal and spatial variability of soil respiration in four Sitka spruce stands. Plant Soil 2006, 287, 161–176. [Google Scholar] [CrossRef]
- Ngao, J.; Epron, D.; Delpierre, N.; Bréda, N.; Granier, A.; Longdoz, B. Spatial variability of soil CO2 efflux linked to soil parameters and ecosystem characteristics in a temperate beech forest. Agric. For. Meteorol. 2012, 154–155, 136–146. [Google Scholar] [CrossRef]
- Stoyan, H.; De-Polli, H.; Böhm, S.; Robertson, G.P.; Paul, E.A. Spatial heterogeneity of soil respiration and related properties at the plant scale. Plant Soil 2000, 222, 203–214. [Google Scholar] [CrossRef]
- Adachi, M.; Bekku, Y.S.; Rashidah, W.; Okuda, T.; Koizumi, H. Differences in soil respiration between different tropical ecosystems. Appl. Soil Ecol. 2006, 34, 258–265. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | Formula | AIC | ∆AIC | Deviance | df | Dispersion parameter |
|---|---|---|---|---|---|---|
| Canopy | ||||||
| 1 | −5167.4 | 3152.4 | 609.3 | 2025 | 0.301 | |
| 2 | −7236.2 | 1083.5 | 226.2 | 2024 | 0.112 | |
| 3 | −5270.1 | 3049.6 | 580.0 | 2024 | 0.287 | |
| 4 | −8319.7 | 0.0 | 133.4 | 2023 | 0.066 | |
| Gap | ||||||
| 1 | −3714.9 | 1551.9 | 473.5 | 1457 | 0.325 | |
| 2 | −4865.5 | 401.3 | 221.0 | 1456 | 0.125 | |
| 3 | −3964.1 | 1302.7 | 401.8 | 1456 | 0.276 | |
| 4 | −5266.8 | 0.0 | 168.6 | 1455 | 0.116 | |
| Factor | Estimate | Standard Error | t Value | p Value |
|---|---|---|---|---|
| Canopy | ||||
| (Intercept) | −10.2589 | 0.1646 | −62.33 | <0.001 |
| Ts | 0.1349 | 0.0015 | 88.11 | <0.001 |
| log W | 1.6838 | 0.0430 | 39.20 | <0.001 |
| Gap | ||||
| (Intercept) | 4.6302 | 0.3816 | 12.14 | <0.001 |
| Ts | 0.1284 | 0.0023 | 55.09 | <0.001 |
| log W | −2.3719 | 0.1038 | −22.86 | <0.001 |
| Model | Estimated Rs (g·C·m−2) | ||
|---|---|---|---|
| Growing Season | Snow Season | ||
| Ts | |||
| All | Rs = 0.0198e0.1260Ts | 437.4 | 153.7 |
| Canopy | Rs = 0.0207e0.1258Ts | 507.7 | 111.8 |
| Gap | Rs = 0.0177e0.1296Ts | 451.8 | 97.0 |
| Total | 493.4 | 108.0 | |
| Ts & W | |||
| Canopy | Rs = 3.5044 × 10−5e0.1349TsW1.684 | 541.4 | No data |
| Gap | Rs = 102.5405e0.1284TsW−2.372 | 384.6 | No data |
| Total | 501.3 | ||
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suchewaboripont, V.; Ando, M.; Yoshitake, S.; Iimura, Y.; Hirota, M.; Ohtsuka, T. Spatial Upscaling of Soil Respiration under a Complex Canopy Structure in an Old‐Growth Deciduous Forest, Central Japan. Forests 2017, 8, 36. https://doi.org/10.3390/f8020036
Suchewaboripont V, Ando M, Yoshitake S, Iimura Y, Hirota M, Ohtsuka T. Spatial Upscaling of Soil Respiration under a Complex Canopy Structure in an Old‐Growth Deciduous Forest, Central Japan. Forests. 2017; 8(2):36. https://doi.org/10.3390/f8020036
Chicago/Turabian StyleSuchewaboripont, Vilanee, Masaki Ando, Shinpei Yoshitake, Yasuo Iimura, Mitsuru Hirota, and Toshiyuki Ohtsuka. 2017. "Spatial Upscaling of Soil Respiration under a Complex Canopy Structure in an Old‐Growth Deciduous Forest, Central Japan" Forests 8, no. 2: 36. https://doi.org/10.3390/f8020036
APA StyleSuchewaboripont, V., Ando, M., Yoshitake, S., Iimura, Y., Hirota, M., & Ohtsuka, T. (2017). Spatial Upscaling of Soil Respiration under a Complex Canopy Structure in an Old‐Growth Deciduous Forest, Central Japan. Forests, 8(2), 36. https://doi.org/10.3390/f8020036