Effect of Stand Structure and Number of Sample Trees on Optimal Management for Scots Pine: A Model-Based Study



Abstract
:1. Introduction
2. Materials and Methods
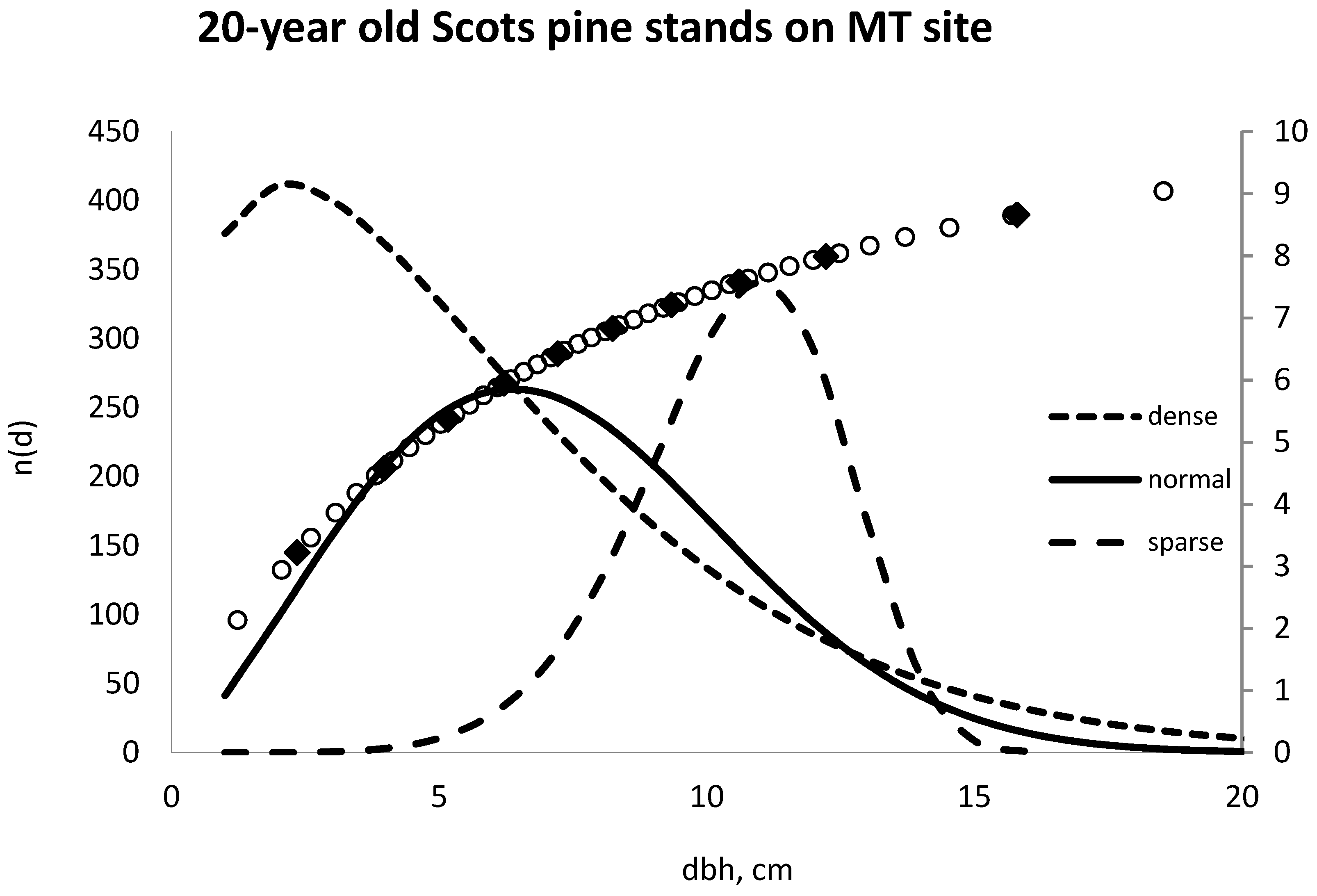
2.1. Initial Stands
2.2. Analyses of the Varying Stand Structures and Sample Sizes
2.3. Stand-level Optimization
3. Results
Optimized Management Schedules
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
| d.d. | G, m2ha−1 | N, ha−1 | dgM, cm | hgM, m | Hdom, m | b0 | b1 | b | c |
|---|---|---|---|---|---|---|---|---|---|
| 1250 | 14.4 | 3500 | 11.3 | 7.9 | 8.7 | 0.951 | 0.310 | 6.250 | 1.320 |
| 1250 | 14.4 | 2276 | 11.3 | 7.9 | 8.7 | 0.979 | 0.307 | 8.176 | 2.300 |
| 1250 | 14.4 | 1600 | 11.3 | 7.9 | 8.6 | 1.017 | 0.302 | 11.310 | 6.460 |
| 1050 | 9.17 | 3500 | 10.0 | 6.6 | 7.0 | 1.093 | 0.325 | 4.486 | 1.119 |
| 1050 | 9.17 | 2150 | 10.0 | 6.6 | 7.0 | 1.129 | 0.321 | 7.130 | 1.753 |
| 1050 | 9.17 | 1200 | 10.0 | 6.6 | 6.9 | 1.174 | 0.316 | 10.158 | 16.396 |
References
- Hyytiäinen, K.; Tahvonen, O.; Valsta, L. Optimum juvenile density, harvesting and stand structure in even-aged Scots pine stands. For. Sci. 2005, 51, 120–133. [Google Scholar]
- Amacher, G.S.; Ollikainen, M.; Koskela, E.A. Economics of Forest Resources; The MIT Press: Cambridge, MA, USA, 2009; p. 448. ISBN 978-0-262-01248-5. [Google Scholar]
- Cao, T. Silvicultural Decisions Based on Simulation-Optimization Systems. Available online: http://www.metla.fi/dissertationes/df103.htm (accessed on 15 May 2018).
- Niinimäki, S.; Tahvonen, O.; Mäkelä, A. Applying a process-based model in Norway spruce management. For. Ecol. Manag. 2012, 265, 102–115. [Google Scholar] [CrossRef]
- Tahvonen, O.; Pihlainen, S.; Niinimäki, S. On the economics of optimal timber production in boreal Scots pine stands. Can. J. For. Res. 2013, 43, 719–730. [Google Scholar] [CrossRef]
- Getz, W.M.; Haight, R.G. Population Harvesting: Demographic Models of Fish, Forest and Animal Resources; Princetown University Press: Princetown, NJ, USA, 1989; p. 408. [Google Scholar]
- Gove, J.H.; Fairweather, S.E. Optimizing the management of uneven-aged forest stands: A stochastic approach. For. Sci. 1992, 38, 623–640. [Google Scholar]
- Haight, R.G. Evaluating the efficiency of even-aged and uneven-aged stand management. For. Sci. 1987, 33, 116–134. [Google Scholar]
- Solberg, B.; Haight, R.G. Analysis of optimal economic management regimes for Picea abies stands using a stage-structured optimal-control model. Scand. J. For. Res. 1991, 6, 559–572. [Google Scholar] [CrossRef]
- Buongiorno, J. Quantifying the implications of transformation from even to uneven-aged forest stands. For. Ecol. Manag. 2001, 51, 121–132. [Google Scholar] [CrossRef]
- Haight, R.G.; Monserud, R.A. Optimizing any-aged management of mixed-species stands.1. Performance of a coordinate-search process. Can. J. For. Res. 1990, 20, 15–25. [Google Scholar] [CrossRef]
- Haight, R.G.; Monserud, R.A.; Chew, J.D. Optimal harvesting with stand density targets: Managing Rocky Mountain conifer stands for multiple forest outputs. For. Sci. 1992, 38, 554–574. [Google Scholar]
- Valsta, L. An optimization model for Norway spruce management based on individual-tree growth models. Acta For. Fenn. 1992, 232, 1–20. [Google Scholar] [CrossRef]
- Wikström, P. Effect of decision variable definition and data aggregation on a search process applied to a single-tree simulator. Can. J. For. Res. 2001, 31, 1057–1066. [Google Scholar] [CrossRef]
- Pukkala, T. Population-based methods in the optimization of stand management. Silva Fenn. 2009, 43, 261–274. [Google Scholar] [CrossRef]
- Wikström, P.; Erikson, L.O. Solving the stand management problem under biodiversity-related considerations. For. Ecol. Manag. 2000, 126, 361–376. [Google Scholar] [CrossRef]
- Mäkelä, A.; del Rio, M.; Hynynen, J.; Hawkings, M.J.; Reyer, C.; Soares, P.; van Oijen, M.; Tome, M. Using stand-scale forest models for estimating indicators for sustainable forest management. For. Ecol. Manag. 2012, 285, 164–178. [Google Scholar] [CrossRef] [Green Version]
- Pretzsch, H. Forest Dynamics, Growth and Yield—From Measurement to Model; Springer: Berlin, Germany, 2009; p. 644. [Google Scholar]
- Weiskittel, A.R.; Hann, D.W.; Kershaw, J.A., Jr.; Vanclay, J.K. Forest Growth and Yield Modeling; John Wiley & Sons Ltd.: Oxford, UK, 2011; p. 430. [Google Scholar]
- Shiver, B.D. Sample sizes and estimation methods for the Weibull distribution for unthinned slash pine plantation diameter distributions. For. Sci. 1988, 34, 809–814. [Google Scholar]
- Kilkki, P.; Maltamo, M.; Mykkänen, R.; Päivinen, R. Use of the Weibull function in the estimation the basal area dbh-distribution. Silva Fenn. 1989, 23, 311–318. [Google Scholar] [CrossRef]
- Picard, N.; Quedraogo, D.; Bar-Hen, A. Choosing classes for size projection matrix models. Ecol. Model. 2010, 221, 2270–2279. [Google Scholar] [CrossRef]
- Hyytiäinen, K.; Tahvonen, O. The effects of legal limits and recommendations on timber production: The case of Finland. For. Sci. 2001, 47, 443–454. [Google Scholar]
- Cajander, A.K. The theory of forest types. Acta For. Fenn. 1925, 29, 10. [Google Scholar] [CrossRef]
- Venäläinen, A.; Tuomenvirta, H.; Pirinen, P.; Drebs, A. A Basic Finnish Climate Data Set 1961–2000—Description and Illustrations. Available online: http://www.oulu.fi/oulugis/doc/data_description.pdf (accessed on 20 April 2018).
- Siipilehto, J. Local prediction of stand structure using linear prediction theory in Scots pine-dominated stands in Finland. Silva Fenn. 2011, 45, 669–692. [Google Scholar] [CrossRef]
- Siipilehto, J.; Mehtätalo, L. Parameter recovery vs. parameter prediction for Scots pine stand structures in Finland. Silva Fenn. 2013, 47, 1057. [Google Scholar] [CrossRef]
- Gove, J.H.; Patil, G.P. Modeling the basal area-size distribution of forest stand: A compatible approach. For. Sci. 1998, 44, 285–297. [Google Scholar]
- Päivinen, R. On the estimation of stem diameter distribution and stand characteristics. Folia For. 1980, 442, 1–28, (In Finnish with English summary). [Google Scholar]
- Kilkki, P.; Päivinen, R. Weibull-function in the esimation of basal area dbh-distribution. Silva Fenn. 1986, 20, 149–156. [Google Scholar] [CrossRef]
- Maltamo, M. Comparing basal area diameter distributions estimated by tree species and for the entire growing stock in a mixed stand. Silva Fenn. 1997, 31, 53–65. [Google Scholar] [CrossRef]
- Kangas, A.; Maltamo, M. Percentile based basal area diameter distribution models for Scots pine, Norway spruce and birch species. Silva Fenn. 2000, 34, 371–380. [Google Scholar] [CrossRef]
- VMI11 Maastotyöohje 2012/Koko Suomi. NFI11 Field Work Guidelines 2012/Whole Finland. Available online: http://www.metla.fi/ohjelma/vmi/vmi11-maasto-ohje09-2p.pdf (accessed on 18 October 2018).
- Hynynen, J.; Ojansuu, R.; Hökkä, H.; Salminen, H.; Siipilehto, J.; Haapala, P. Models for Predicting Stand Development in MELA System; Finnish Forest Research Institute: Vantaa, Finland, 2002; Volume 835, p. 116. [Google Scholar]
- Salminen, H.; Lehtonen, M.; Hynynen, J. Reusing legacy FORTRAN in the MOTTI growth and yield simulator. Comput. Electron. Agric. 2005, 49, 103–113. [Google Scholar] [CrossRef]
- Matala, J.; Hynynen, J.; Miina, J.; Ojansuu, R.; Peltola, H.; Sievänen, R.; Väisänen, H.; Kellomäki, S. Comparison of a physiological model and a statistical model for prediction of growth and yield in boreal forests. Ecol. Model. 2003, 161, 95–116. [Google Scholar] [CrossRef]
- Huuskonen, S. Nuorten Männiköiden Kehitys—Taimikonhoito ja Ensiharvennus. Available online: http://www.metla.fi/dissertationes/df62.htm (accessed on 31 March 2018).
- Mäkinen, H.; Hynynen, J.; Isomäki, A. Intensive management of Scots pine stands in southern Finland: First empirical results and simulated further development. For. Ecol. Manag. 2005, 215, 37–50. [Google Scholar] [CrossRef]
- Ahtikoski, A.; Salminen, H.; Hökkä, H.; Kojola, S.; Penttilä, T. Optimising stand management on peatlands: The case of northern Finland. Can. J. For. Res. 2012, 42, 247–259. [Google Scholar] [CrossRef]
- Ahtikoski, A.; Salminen, H.; Ojansuu, R.; Hynynen, J.; Kärkkäinen, K.; Haapanen, M. Optimizing stand management involving the effect of genetic gain: Preliminary results for Scots pine in Finland. Can. J. For. Res. 2013, 43, 1–7. [Google Scholar] [CrossRef]
- Äijälä, O.; Koistinen, A.; Sved, J.; Vanhatalo, K.; Väisänen, P. Metsänhoidon Suositukset (Silvicultural recommendations); The Forestry Development Centre Tapio: Helsinkin, Finland, 2014; ISBN 978-952-6612-32-4. (In Finnish) [Google Scholar]
- Miina, J.; Pukkala, T.; Hotanen, J.-P.; Salo, K. Optimizing the joint production of timber and bilberries. For. Ecol. Manag. 2010, 259, 2065–2071. [Google Scholar] [CrossRef]
- Cao, T.; Valsta, L.; Mäkelä, A. A comparison of carbon assessment methods for optimizing timber production and carbon sequestration in Scots pine stands. For. Ecol. Manag. 2010, 260, 1726–1734. [Google Scholar] [CrossRef]
- Charbonneau, P.; Knapp, B. A User’s Guide to PIKAIA 1.0; NCAR Technical Note 418+IA; High Altitude Observatory, National Center for Atmospheric Research: Boulder, CO, USA, 1995. [Google Scholar]
- Metcalfe, T.S.; Charbonneau, P. Stellar structure modeling using a parallel genetic algorithm for objective global optimization. J. Comput. Phys. 2003, 185, 176–193. [Google Scholar] [CrossRef] [Green Version]
- Li, M.J.; Chen, D.S.; Wang, F.; Li, Y.; Zhou, Y.; Lang, J.L. Optimizing emission inventory for chemical transport models by using genetic algorithm. Atmos. Environ. 2010, 44, 3926–3934. [Google Scholar] [CrossRef]
- Hadi, M.R.H.S.; Gonzalez-Andujar, J.L. Comparison of fitting weed seedling emergence models with nonlinear regression and genetic algorithm. Comput. Electron. Agric. 2009, 65, 19–25. [Google Scholar] [CrossRef]
- Statistics Finland 2013. Citing Online Sources: Cost-of-living, Appendix Table 2. Available online: http://www.stat.fi (accessed on 21 April 2014).
- Siipilehto, J.; Sarkkola, S.; Mehtätalo, L. Comparing regression estimation techniques when predicting diameter distributions of Scots pine on drained peatlands. Silva Fenn. 2007, 41, 333–349. [Google Scholar] [CrossRef]
- Holopainen, M.; Mäkinen, A.; Rasinmäki, J.; Hyytiäinen, K.; Bayazidi, S.; Pietilä, I. Comparison of various sources of uncertainty in stand-level net present value estimates. For. Policy Econ. 2010, 12, 377–386. [Google Scholar] [CrossRef]
- Pukkala, T.; Miina, J. Optimising the management of heterogeneous stand. Silva Fenn. 2005, 39, 525–538. [Google Scholar] [CrossRef]
- Burk, T.J.; Newberry, J.D. A simple algorithm for moment-based recovery of Weibull distribution parameters. For. Sci. 1984, 30, 329–332. [Google Scholar]
- Cao, Q.V. Predicting parameters of a Weibull function for modeling diameter distribution. For. Sci. 2004, 50, 682–685. [Google Scholar]
- Newton, P.F.; Lei, Y.; Zhang, S.Y. A parameter recovery model for estimating black spruce diameter distribution within the context of a stand density management diagram. For. Chron. 2004, 80, 349–357. [Google Scholar] [CrossRef]
- Palahi, M.; Pukkala, T.; Trasobares, A. Modelling the diameter distribution of Pinus sylvestris, Pinus nigra and Pinus halepensis forest stands in Catalonia using the truncated Weibull function. Forestry 2006, 79, 553–562. [Google Scholar] [CrossRef]
- Liu, C.; Zhang, S.Y.; Beaulieu, J.; Prégent, G.; Zhang, S.Y. Application and comparison of six methods for predicting parameters of the Weibull function in unthinned Picea glauca plantations. Scan. J. For. Res. 2009, 24, 67–75. [Google Scholar] [CrossRef]
- Arias-Rodill, M.; Pukkala, T.; Gonzalez-Gonzalez, J.M.; Barrio-Anta, M.; Dieguez-Aranda, U. Use of depth-first search and direct search methods to optimize even-aged stand management: A case study involving maritime pine in Asturias (northwest Spain). Can. J. For. Res. 2015, 45, 1269–1279. [Google Scholar] [CrossRef]
- Arias-Rodill, M.; Dieguez-Aranda, U.; Vazquez-Mendez, M.E. A differentiable optimization model for the management of single-species, even-aged stands. Can. J. For. Res. 2017, 47, 506–514. [Google Scholar] [CrossRef] [Green Version]
- Ying-Hua, C. Adopting co-evolution and constraint-satisfaction concept on genetic algorithms to solve supply chain network design problems. Expert Syst. Appl. 2010, 37, 6919–6930. [Google Scholar] [CrossRef]
- Seidl, R.; Rammer, W.; Jäger, D.; Currie, W.S.; Lexer, M.J. Assessing trade-offs between carbon sequestration and timber production within a framework of multi-purpose forestry in Austria. For. Ecol. Manag. 2007, 248, 64–79. [Google Scholar] [CrossRef]
- Hynynen, J.; Salminen, H.; Ahtikoski, A.; Huuskonen, S.; Ojansuu, R.; Siipilehto, J.; Lehtonen, M.; Eerikäinen, K. Long-term impacts of forest management on biomass supply and forest resource development: A scenario analysis for Finland. Eur. J. For. Res. 2015, 134, 415–431. [Google Scholar] [CrossRef]
- Siipilehto, J.; Lindeman, H.; Vastaranta, M.; Yu, X.; Uusitalo, J. Reliability of the predicted stand structure for clear-cut stands using optional methods: Airborne laser scanning-based methods, smartphone-based forest inventory application Trestima and pre-harvest measurement tool EMO. Silva Fenn. 2016, 50, 1568. [Google Scholar] [CrossRef]
- Ojansuu, R.; Mäkinen, H.; Heinonen, J. Including variation in branch and tree properties improves timber grade estimates in Scots pine stands. Can. J. For. Res. 2018, 48, 542–553. [Google Scholar] [CrossRef]
| Stand Density | Loc | Dr, % | NPVMax of the Optimal Stand Management, €/ha | NPVMax in Relation to That of n = 10, % | NPVMax in Relation to That of Average Density, % | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| n: 10 | 20 | 40 | 20 | 40 | 10 | 20 | 40 | |||
| Sparse | SF | 3 | 6281 | 6310 | 6336 | 0.5 | 0.9 | −2.4 | −1.2 | −0.3 |
| 4 | 4522 | 4593 | 4611 | 1.6 | 2.0 | −2.6 | 0.0 | 1.0 | ||
| CF | 3 | 3170 | 3225 | 3219 | 1.7 | 1.5 | 3.3 | 5.9 | 6.7 | |
| 4 | 2083 | 2067 | 2065 | −0.8 | −0.9 | 3.7 | 3.8 | 5.5 | ||
| Average | SF | 3 | 6435 | 6389 | 6353 | −0.7 | −1.3 | |||
| 4 | 4644 | 4591 | 4567 | −1.1 | −1.7 | |||||
| CF | 3 | 3069 | 3046 | 3018 | −0.7 | −1.7 | ||||
| 4 | 2009 | 1992 | 1957 | −0.8 | −2.6 | |||||
| Dense | SF | 3 | 5827 | 5878 | 6006 | 0.9 | 3.1 | −9.4 | −8.0 | −5.5 |
| 4 | 4123 | 4157 | 4277 | 0.8 | 3.7 | −11.2 | −9.5 | −6.3 | ||
| CF | 3 | 2513 | 2626 | 2616 | 4.5 | 4.1 | −18.1 | −13.8 | −13.3 | |
| 4 | 1571 | 1623 | 1691 | 3.3 | 7.6 | −21.8 | −18.5 | −13.6 | ||
| Stand Density | Loc | Dr % | First Thinning: Timing (Years)/ Removal Volume (m3/ha) | Intermediate Thinnings: Number of Thinnings/Removal Volume | Clear-cutting, Years/Pulpwood/Logs (m3/ha) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| n: 10 | 20 | 40 | 10 | 20 | 40 | 10 | 20 | 40 | |||
| Sparse | SF | 3 | 29/49.4 | 29/49.8 | 29/47.1 | 3/215.4 | 3/216.1 | 3/211.9 | 64/65/182 | 64/66/182 | 64/65/182 |
| 4 | 29/49.7 | 29/50.9 | 29/47.5 | 3/215.8 | 3/215.0 | 3/210.9 | 61/62/179 | 63/65/181 | 63/64/180 | ||
| CF | 3 | 40/45.0 | 38/46.4 | 38/45.9 | 3/180.2 | 2/115.6 | 2/115.8 | 75/47/162 | 78/51/180 | 78/51/179 | |
| 4 | 38/45.0 | 38/46.5 | 38/46.5 | 2/118.5 | 2/115.5 | 2/115.6 | 74/46/166 | 77/51/180 | 77/50/178 | ||
| Average | SF | 3 | 30/57.3 | 30/57.3 | 30/44.6 | 4/315.0 | 3/209.5 | 3/210.2 | 71/58/167 | 70/94/235 | 69/67/220 |
| 4 | 30/53.1 | 30/57.4 | 30/57.6 | 4/298.0 | 4/312.0 | 4/309.4 | 70/62/169 | 71/58/168 | 71/58/167 | ||
| CF | 3 | 40/45.0 | 40/45.9 | 42/45.0 | 3/180.2 | 3/183.6 | 2/116.6 | 83/75/131 | 80/67/117 | 80/73/162 | |
| 4 | 40/45.2 | 40/47.3 | 40/52,3 | 3/181.0 | 3/187.0 | 3/190.7 | 80/75/117 | 80/69/115 | 80/75/108 | ||
| Dense | SF | 3 | 32/42.4 | 31/47.3 | 32/54.9 | 3/216.8 | 4/284.0 | 4/298.4 | 70/74/197 | 71/74/165 | 72/74/166 |
| 4 | 31/42.3 | 31/55.5 | 32/54.3 | 3/213.4 | 3/208.0 | 4/299.1 | 70/81/201 | 70/114/203 | 72/74/166 | ||
| CF | 3 | 53/68.2 | 50/65.5 | 42/45.3 | 2/139.3 | 3/214.6 | 2/112.2 | 84/112/106 | 97/78/137 | 80/118/121 | |
| 4 | 53/65.9 | 50/65.4 | 42/48.6 | 2/136.6 | 2/132.0 | 2/112.0 | 81/105/96 | 84/120/112 | 80/129/116 | ||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahtikoski, A.; Siipilehto, J.; Salminen, H.; Lehtonen, M.; Hynynen, J. Effect of Stand Structure and Number of Sample Trees on Optimal Management for Scots Pine: A Model-Based Study. Forests 2018, 9, 750. https://doi.org/10.3390/f9120750
Ahtikoski A, Siipilehto J, Salminen H, Lehtonen M, Hynynen J. Effect of Stand Structure and Number of Sample Trees on Optimal Management for Scots Pine: A Model-Based Study. Forests. 2018; 9(12):750. https://doi.org/10.3390/f9120750
Chicago/Turabian StyleAhtikoski, Anssi, Jouni Siipilehto, Hannu Salminen, Mika Lehtonen, and Jari Hynynen. 2018. "Effect of Stand Structure and Number of Sample Trees on Optimal Management for Scots Pine: A Model-Based Study" Forests 9, no. 12: 750. https://doi.org/10.3390/f9120750