Overlapping Bark Beetle Outbreaks, Salvage Logging and Wildfire Restructure a Lodgepole Pine Ecosystem

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
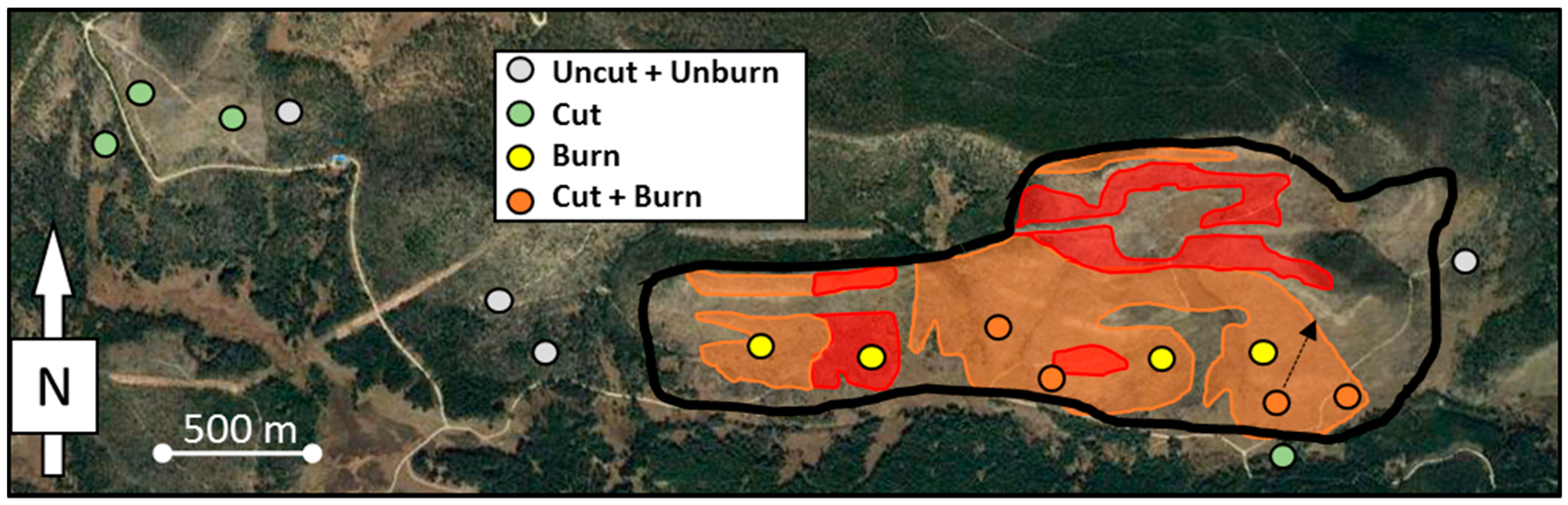
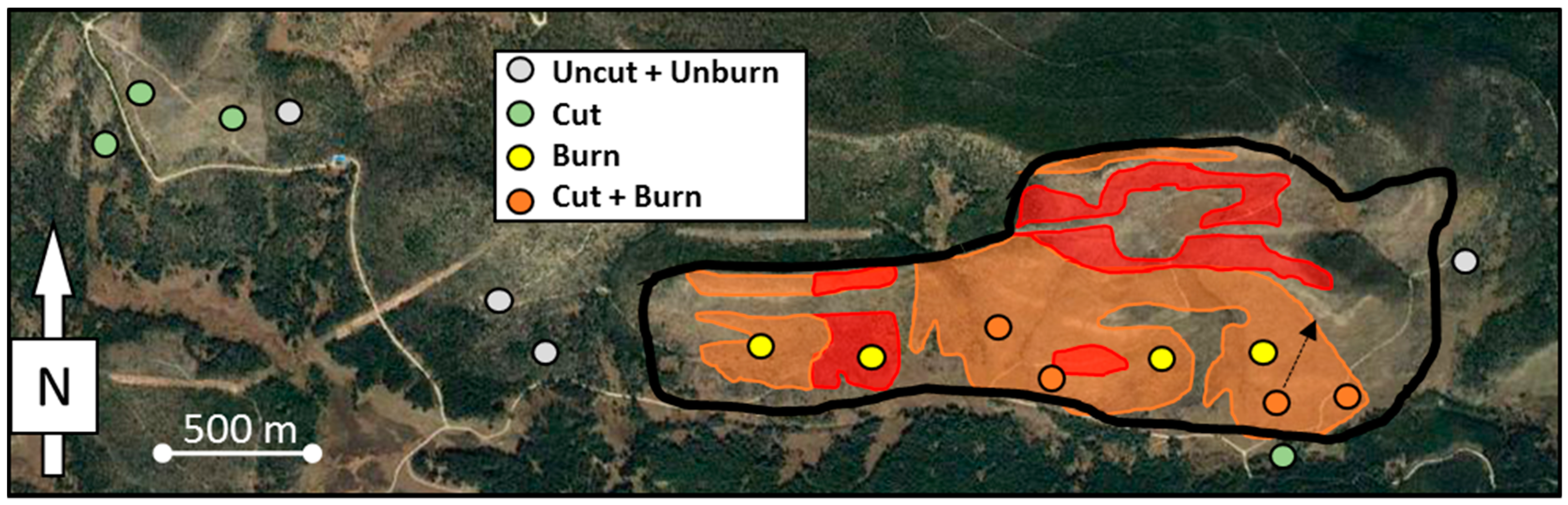
2.1. Study Area
2.2. Sampling and Analysis
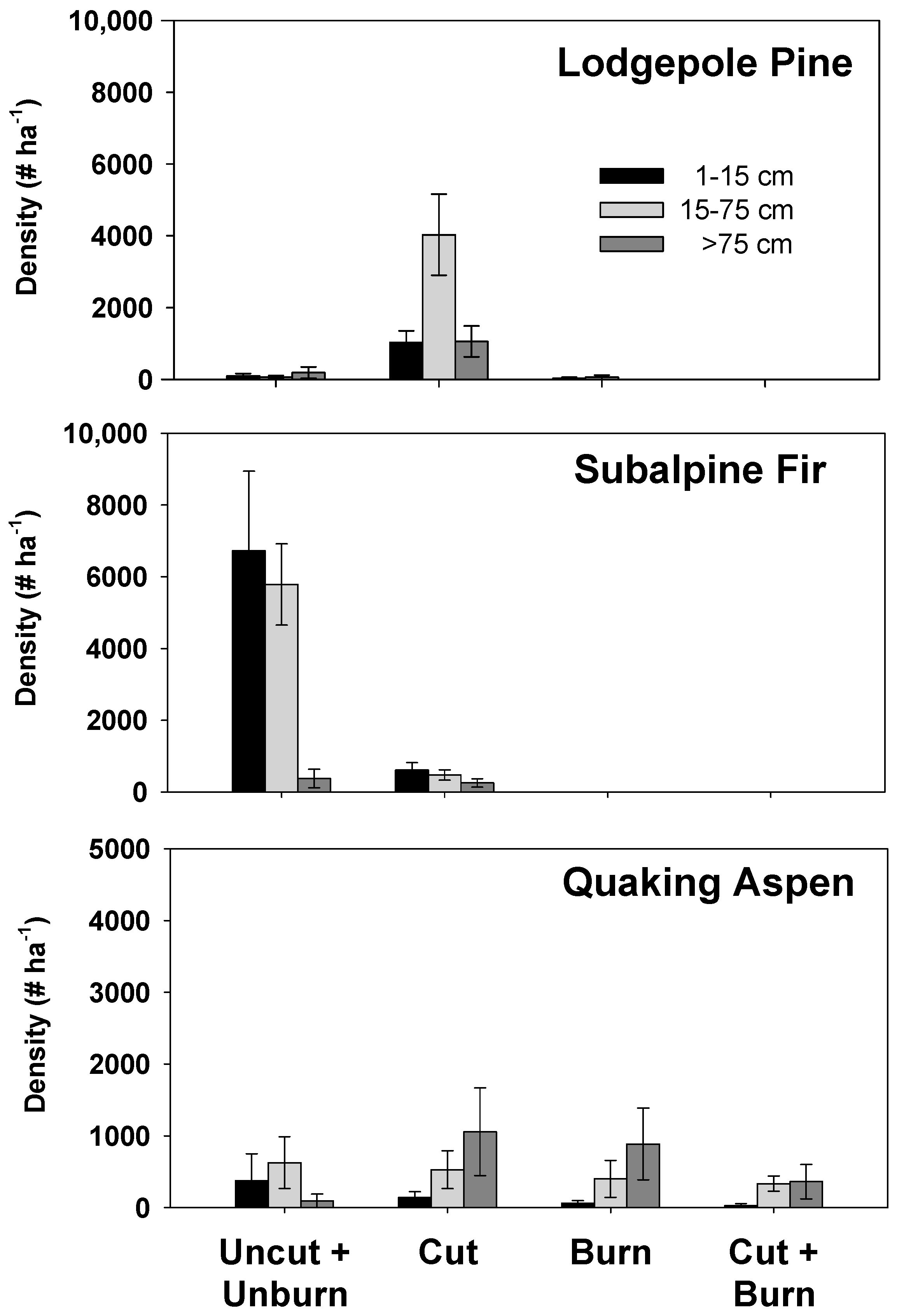
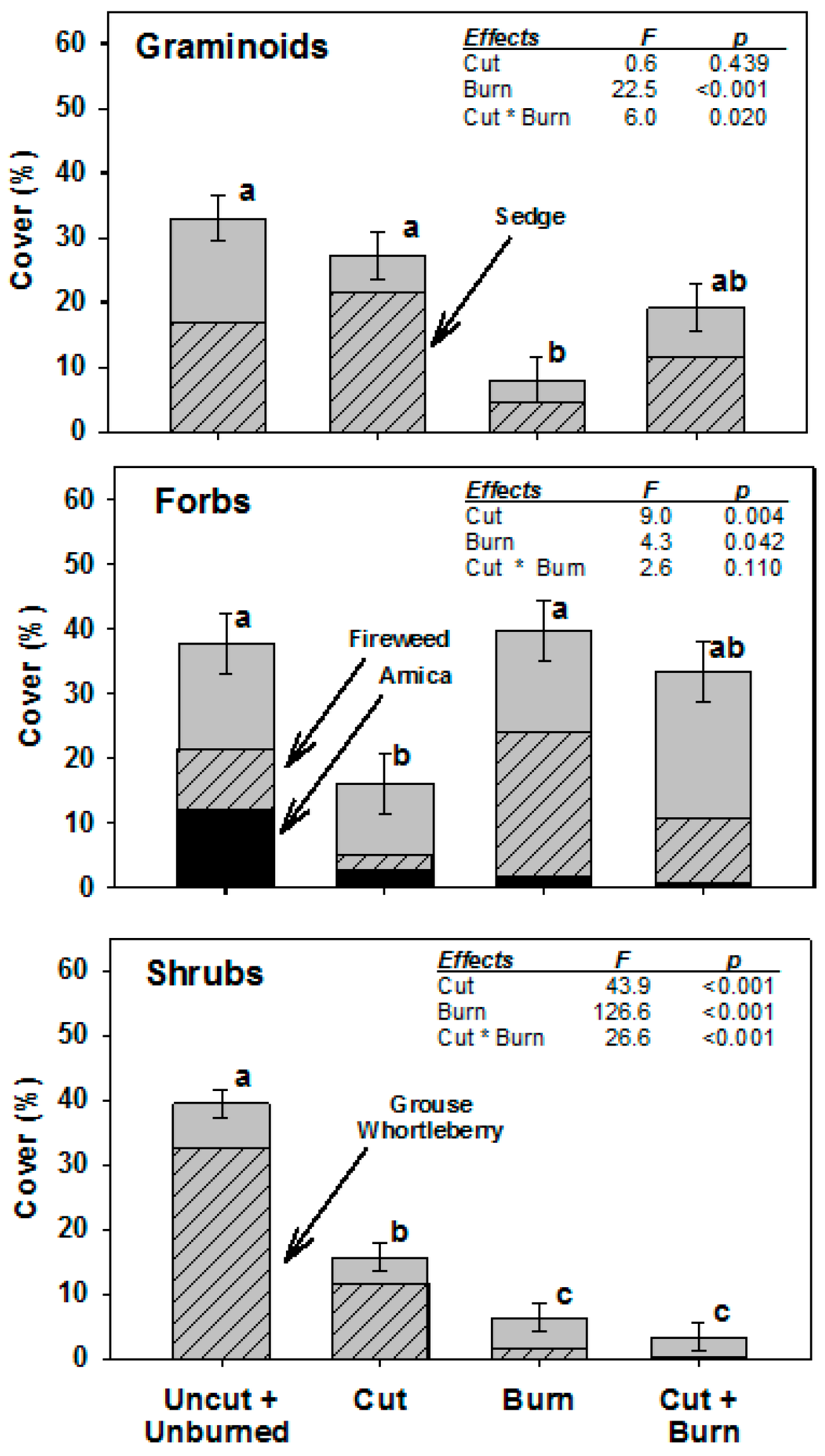
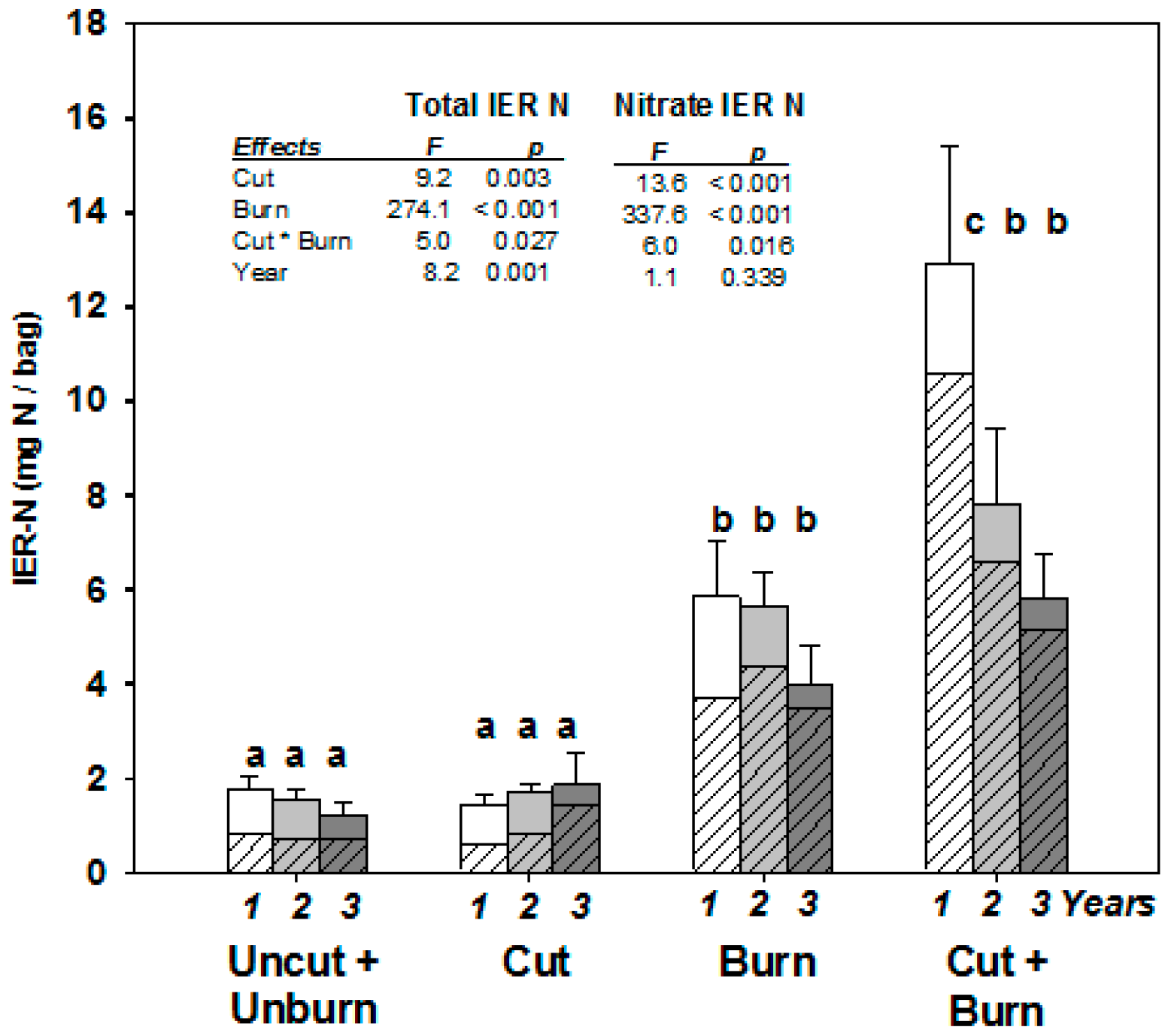
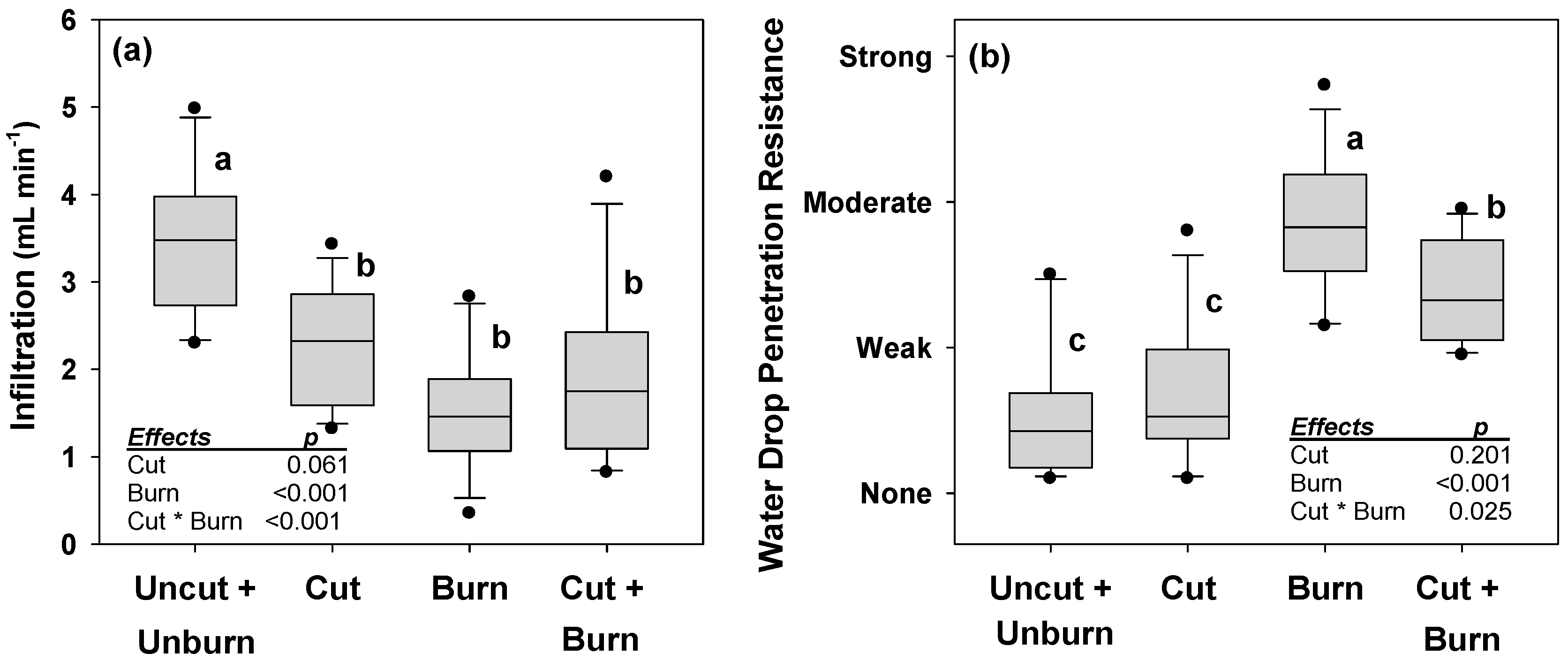
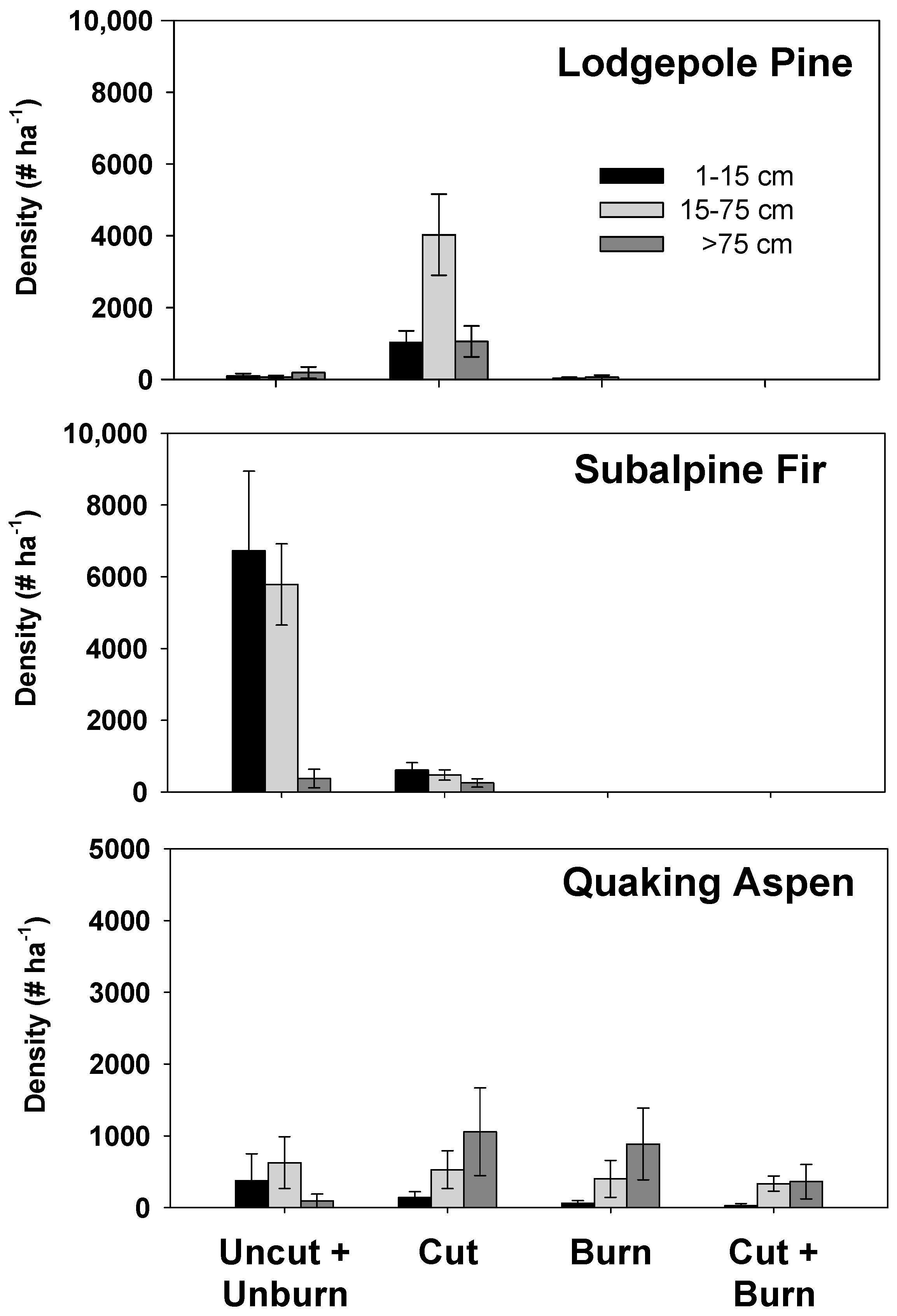
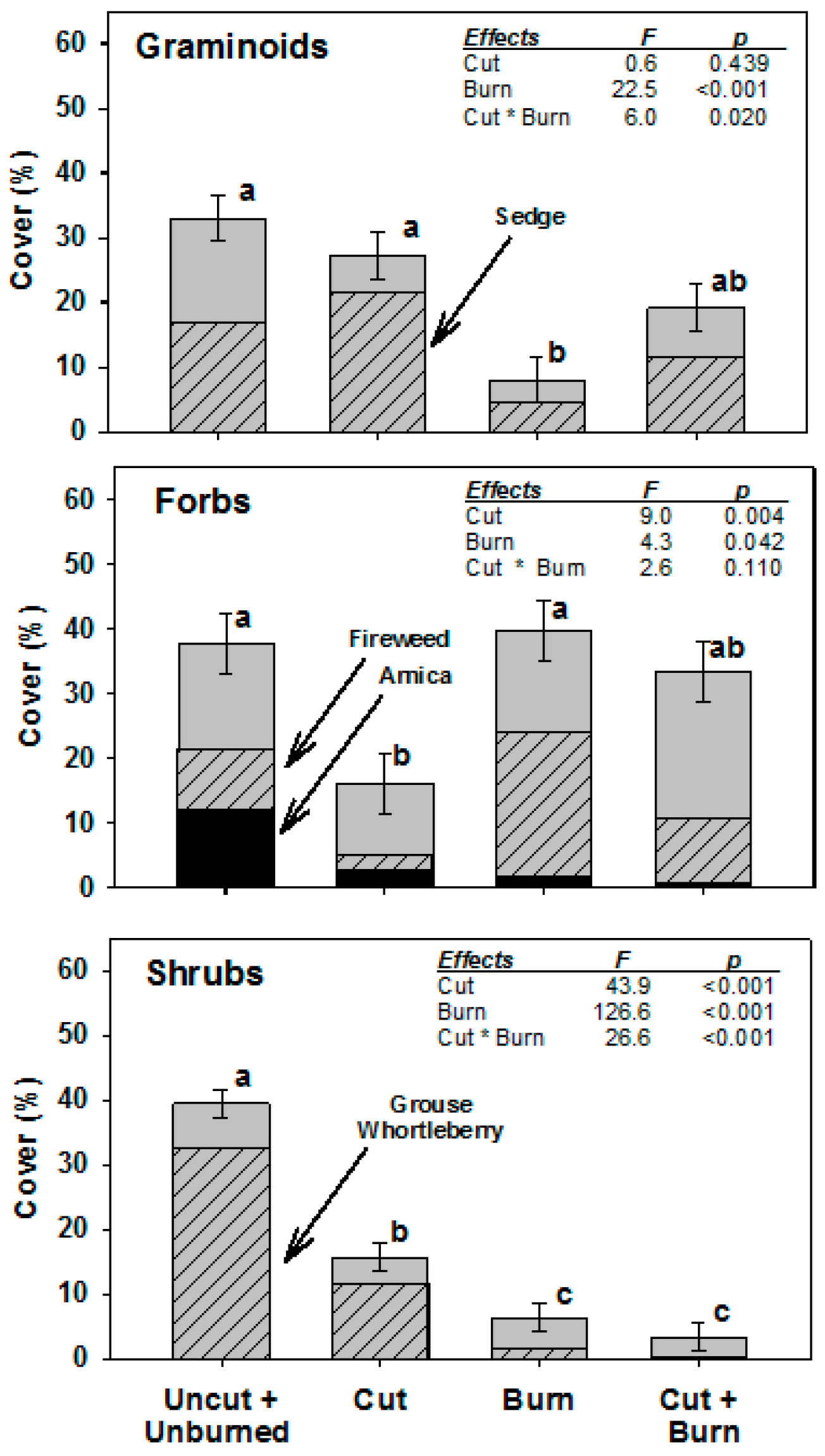
3. Results
4. Discussion
4.1. Overlapping Disturbances
4.2. Implications of Post-Bark Beetle Salvage Logging on Wildfire Effects
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lotan, J.E.; Critchfield, W.B. Pinus contorta Dougl. ex. Loud. lodgepole pine. In Silvics of North America. Volume 1. Conifers; Burns, R.M., Honkala, B.H., Eds.; U.S. USDA: Washington, DC, USA, 1990; Volume 1, pp. 302–315. [Google Scholar]
- Turner, M.G.; Romme, W.H.; Gardner, R.H.; Hargrove, W.W. Effects of fire size and pattern on early succession in Yellowstone National Park. Ecol. Monogr. 1997, 67, 411–433. [Google Scholar] [CrossRef]
- Collins, B.J.; Rhoades, C.C.; Hubbard, R.M.; Battaglia, M.A. Tree regeneration and future stand development after bark beetle infestation and harvesting in Colorado lodgepole pine stands. For. Ecol. Manag. 2011, 261, 2168–2175. [Google Scholar] [CrossRef]
- Diskin, M.; Rocca, M.E.; Nelson, K.N.; Aoki, C.F.; Romme, W.H. Forest developmental trajectories in mountain pine beetle disturbed forests of Rocky Mountain National Park, Colorado. Can. J. For. Res. 2011, 41, 782–792. [Google Scholar] [CrossRef]
- Stone, W.E.; Wolfe, M.L. Response of understorey vegetation to variable tree mortality following a mountain pine beetle epidemic in lodgepole pine stands in northern Utah. Vegetatio 1996, 122, 1–12. [Google Scholar] [CrossRef]
- Selmants, P.C.; Knight, D.H. Understory plant species composition 30–50 years after clearcutting in southeastern Wyoming coniferous forests. For. Ecol. Manag. 2003, 185, 275–289. [Google Scholar] [CrossRef]
- Turner, M.G. Disturbance and landscape dynamics in a changing world. Ecology 2010, 91, 2833–2849. [Google Scholar] [CrossRef] [PubMed]
- Fornwalt, P.J.; Rhoades, C.C.; Hubbard, R.M.; Harris, R.L.; Faist, A.M.; Bowman, W.D. Short-term understory plant community responses to salvage logging in beetle-affected lodgepole pine forests. For. Ecol. Manag. 2018, 409, 84–93. [Google Scholar] [CrossRef]
- Romme, W.H.; Whitby, T.G.; Tinker, D.B.; Turner, M.G. Deterministic and stochastic processes lead to divergence in plant communities 25 years after the 1988 Yellowstone fires. Ecol. Monogr. 2016, 86, 327–351. [Google Scholar] [CrossRef]
- Parsons, W.F.J.; Knight, D.H.; Miller, S.L. Root gap dynamics in lodgepole pine forest: Nitrogen transformations in gaps of different sizes. Ecol. Appl. 1994, 4, 354–362. [Google Scholar] [CrossRef]
- Griffin, J.M.; Turner, M.G.; Simard, M. Nitrogen cycling following mountain pine beetle disturbance in lodgepole pine forests of Greater Yellowstone. For. Ecol. Manag. 2011, 261, 1077–1089. [Google Scholar] [CrossRef]
- Rhoades, C.C.; McCutchan, J.H.; Cooper, L.A.; Clow, D.W.; Detmer, T.M.; Briggs, J.S.; Stednick, J.D.; Veblen, T.T.; Ertz, R.M.; Likens, G.; et al. Biogeochemistry of beetle kill: Explaining a weak nitrate response. Proc. Natl. Acad. Sci. USA 2013, 110, 1756–1760. [Google Scholar] [CrossRef] [PubMed]
- Colorado State Forest Service (CSFS). 2015 Report on the Health of Colorado’s Forests: 15 Years of Change; Colorado State University: Fort Collins, CO, USA, 2016; Volume 32, Available online: https://csfs.colostate.edu/media/sites/22/2016/02/ForestHealthReport-2015.pdf (accessed on 14 January 2018).
- Edwards, M.; Krawchuk, M.A.; Burton, P.J. Short-interval disturbance in lodgepole pine forests, British Columbia, Canada: Understory and overstory response to mountain pine beetle and fire. For. Ecol. Manag. 2015, 338, 163–175. [Google Scholar] [CrossRef]
- Harvey, B.J.; Donato, D.C.; Turner, M.G. Recent mountain pine beetle outbreaks, wildfire severity, and postfire tree regeneration in the US Northern Rockies. Proc. Natl. Acad. Sci. USA 2016, 111, 15120–15125. [Google Scholar] [CrossRef] [PubMed]
- Kane, J.M.; Varner, J.M.; Metz, M.R.; van Mantgem, P.J. Characterizing interactions between fire and other disturbances and their impacts on tree mortality in western U.S. Forests. For. Ecol. Manag. 2017, 405, 188–199. [Google Scholar] [CrossRef]
- Pelz, K.A.; Smith, F.W. Thirty year change in lodgepole and lodgepole/mixed conifer forest structure following 1980s mountain pine beetle outbreak in western Colorado, USA. For. Ecol. Manag. 2012, 280, 93–102. [Google Scholar] [CrossRef]
- Kulakowski, D.; Matthews, C.; Jarvis, D.; Veblen, T.T. Compounded disturbances in sub-alpine forests in western Colorado favour future dominance by quaking aspen (Populus tremuloides). J. Veg. Sci. 2013, 24, 168–176. [Google Scholar] [CrossRef]
- Seidl, R.; Donato, D.C.; Raffa, K.F.; Turner, M.G. Spatial variability in tree regeneration after wildfire delays and dampens future bark beetle outbreaks. Proc. Natl. Acad. Sci. USA 2016, 113, 13075–13080. [Google Scholar] [CrossRef] [PubMed]
- Turner, M.G.; Gardner, R.H.; Romme, W.H. Prefire heterogeneity, fire severity, and early postfire plant reestablishment in subalpine forests of Yellowstone National Park, Wyoming. J. Int. Assoc. Wildland Fire 1999, 9, 21–36. [Google Scholar] [CrossRef]
- Harvey, B.J.; Donato, D.C.; Romme, W.H.; Turner, M.G. Fire severity and tree regeneration following bark beetle outbreaks: The role of outbreak stage and burning conditions. Ecol. Appl. 2014, 24, 1608–1625. [Google Scholar] [CrossRef] [PubMed]
- Agne, M.C.; Woolley, T.; Fitzgerald, S. Fire severity and cumulative disturbance effects in the post-mountain pine beetle lodgepole pine forests of the Pole Creek Fire. For. Ecol. Manag. 2016, 366, 73–86. [Google Scholar]
- Meigs, G.W.; Zald, H.S.; Campbell, J.L.; Keeton, W.S.; Kennedy, R.E. Do insect outbreaks reduce the severity of subsequent forest fires? Environ. Res. Lett. 2016, 11, 045008. [Google Scholar] [CrossRef]
- Jolly, W.M.; Parsons, R.A.; Hadlow, A.M.; Cohn, G.M.; McAllister, S.S.; Popp, J.B.; Hubbard, R.M.; Negron, J.F. Relationships between moisture, chemistry, and ignition of Pinus contorta needles during the early stages of mountain pine beetle attack. For. Ecol. Manag. 2012, 269, 52–59. [Google Scholar] [CrossRef]
- Hart, S.J.; Schoennagel, T.; Veblen, T.T.; Chapman, T.B. Area burned in the western United States is unaffected by recent mountain pine beetle outbreaks. Proc. Natl. Acad. Sci. USA 2015, 112, 4375–4380. [Google Scholar] [CrossRef] [PubMed]
- Lynch, H.J.; Renkin, R.A.; Crabtree, R.L.; Moorcroft, P.R. The Influence of previous mountain pine beetle (Dendroctonus ponderosae) activity on the 1988 Yellowstone Fires. Ecosystems 2006, 9, 1318–1327. [Google Scholar] [CrossRef]
- Hubbard, R.M.; Rhoades, C.C.; Elder, K.; Negron, J.F. Changes in transpiration and foliage growth in lodgepole pine trees following mountain pine beetle attack and mechanical girdling. For. Ecol. Manag. 2013, 289, 312–317. [Google Scholar] [CrossRef]
- Teste, F.P.; Lieffers, V.J.; Landhäusser, S.M. Viability of forest floor and canopy seed banks in Pinus contorta var. latifolia (Pinaceae) forests after a mountain pine beetle outbreak. Am. J. Bot. 2011, 98, 630–637. [Google Scholar] [PubMed]
- Aoki, C.F.; Romme, W.H.; Rocca, M.E. Lodgepole pine seed germination following tree death from mountain pine beetle attack in Colorado, USA. Am. Midl. Nat. 2011, 165, 446–451. [Google Scholar] [CrossRef]
- Tinker, D.B.; Romme, W.H.; Hargrove, W.W.; Gardner, R.H.; Turner, M.G. Landscape-scale heterogeneity in lodgepole pine serotiny. Can. J. For. Res. 1994, 24, 897–903. [Google Scholar] [CrossRef]
- Collins, B.J.; Rhoades, C.C.; Underhill, J.; Hubbard, R.M. Post-harvest seedling recruitment following mountain pine beetle infestation of Colorado lodgepole pine stands: A comparison using historic survey records. Can. J. For. Res. 2010, 40, 2452–2456. [Google Scholar] [CrossRef]
- Page, W.G.; Alexander, M.E.; Jenkins, M.J. Wildfire’s resistance to control in mountain pine beetle-attacked lodgepole pine forests. Forest. Chron. 2013, 89, 783–794. [Google Scholar] [CrossRef]
- Pelz, K.A.; Rhoades, C.C.; Hubbard, R.M.; Battaglia, M.A.; Smith, F.W. Species composition influences management outcomes following mountain pine beetle in lodgepole pine-dominated forests. For. Ecol. Manag. 2015, 336, 11–20. [Google Scholar] [CrossRef]
- Hood, P.R.; Nelson, K.N.; Rhoades, C.C.; Tinker, D.B. The effect of salvage logging on surface fuel loads and fuel moisture in beetle-infested lodgepole pine forests. For. Ecol. Manag. 2017, 390, 80–88. [Google Scholar] [CrossRef]
- Griffin, J.M.; Simard, M.; Turner, M.G. Salvage harvest effects on advance tree regeneration, soil nitrogen, and fuels following mountain pine beetle outbreak in lodgepole pine. For. Ecol. Manag. 2013, 291, 228–239. [Google Scholar] [CrossRef]
- Donato, D.C.; Simard, M.; Romme, W.H.; Harvey, B.J.; Turner, M.G. Evaluating post-outbreak management effects on future fuel profiles and stand structure in bark beetle-impacted forests of Greater Yellowstone. For. Ecol. Manag. 2013, 303, 160–174. [Google Scholar] [CrossRef]
- Raffa, K.F.; Aukema, B.H.; Bentz, B.J.; Carroll, A.L.; Hicke, J.A.; Turner, M.G.; Romme, W.H. Cross-scale drivers of natural disturbances prone to anthropogenic amplification: The dynamics of bark beetle eruptions. BioScience 2008, 58, 501–517. [Google Scholar] [CrossRef]
- Alstatt, D.; Miles, R.L. Soil Survey of Grand County Area, Colorado; USDA: Washington, DC, USA, 1983; p. 174.
- Bailey, R.G. Ecoregions: The Ecosystem Geography of the Oceans and the Continents; Springer: New York, NY, USA, 1998. [Google Scholar]
- Tishmack, J.; Mata, S.A.; Schmid, J.M.; Porth, L. Mountain Pine Beetle Emergence from Lodgepole Pine at Different Elevations Near Fraser, CO; USDA: Fort Collins, CO, USA, 2004; p. 13.
- Chapman, T.B.; Veblen, T.T.; Schoennagel, T. Spatiotemporal patterns of mountain pine beetle activity in the southern Rocky Mountains. Ecology 2012, 93, 2175–2185. [Google Scholar] [CrossRef] [PubMed]
- Rhoades, C.C.; Hubbard, R.M.; Elder, K. A Decade of streamwater nitrogen and forest cynamics after a mountain pine beetle outbreak at the Fraser Experimental Forest, Colorado. Ecosystems 2016, 20, 380–392. [Google Scholar] [CrossRef]
- Rhoades, C. Church’s Park Fire Study, Field Data; US Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2018.
- USDA Forest Service (USFS), InciWeb. Church’s Park Fire. Fire and Aviation Management, Washington, DC. 2010. Available online: https://www.fs.fed.us/fire/aviation/av_library/FS%20Special%20Mission%20Airworthiness%20Assurance%20Guide_Final_11_5_2010.pdf (accessed on 15 December 2010).
- Western Regional Climate Center (WRCC). Remote automatic weather station. Western Regional Climate Center, Reno, NV. 2017. Available online: https://www.dri.edu/images/stories/news/media_kits/WRCCFactSheet11_15_3.pdf (accessed on 10 December 2017).
- Moriarty, K. Firefighter Observations on Mountain Pine Beetle Post-Outbreak Lodgepole Pine fires: Expectations, Surprises and Decision-Making. Master’s Thesis, Colorado State University, Fort Collins, CO, USA, 2014. [Google Scholar]
- Parsons, A.; Robichaud, P.R.; Lewis, S.A.; Napper, C.; Clark, J.T. Field Guide for Mapping Post-Fire Soil Burn Severity; USDA: Fort Collins, CO, USA, 2010; p. 49.
- USDA Forest Service (USFS). Church’s Park Fire—Burned Area Emergency Rehabilitation Burn Severity Estimate—Technical Report; USDA Forest Service (USFS): Denver, CO, USA, 2010.
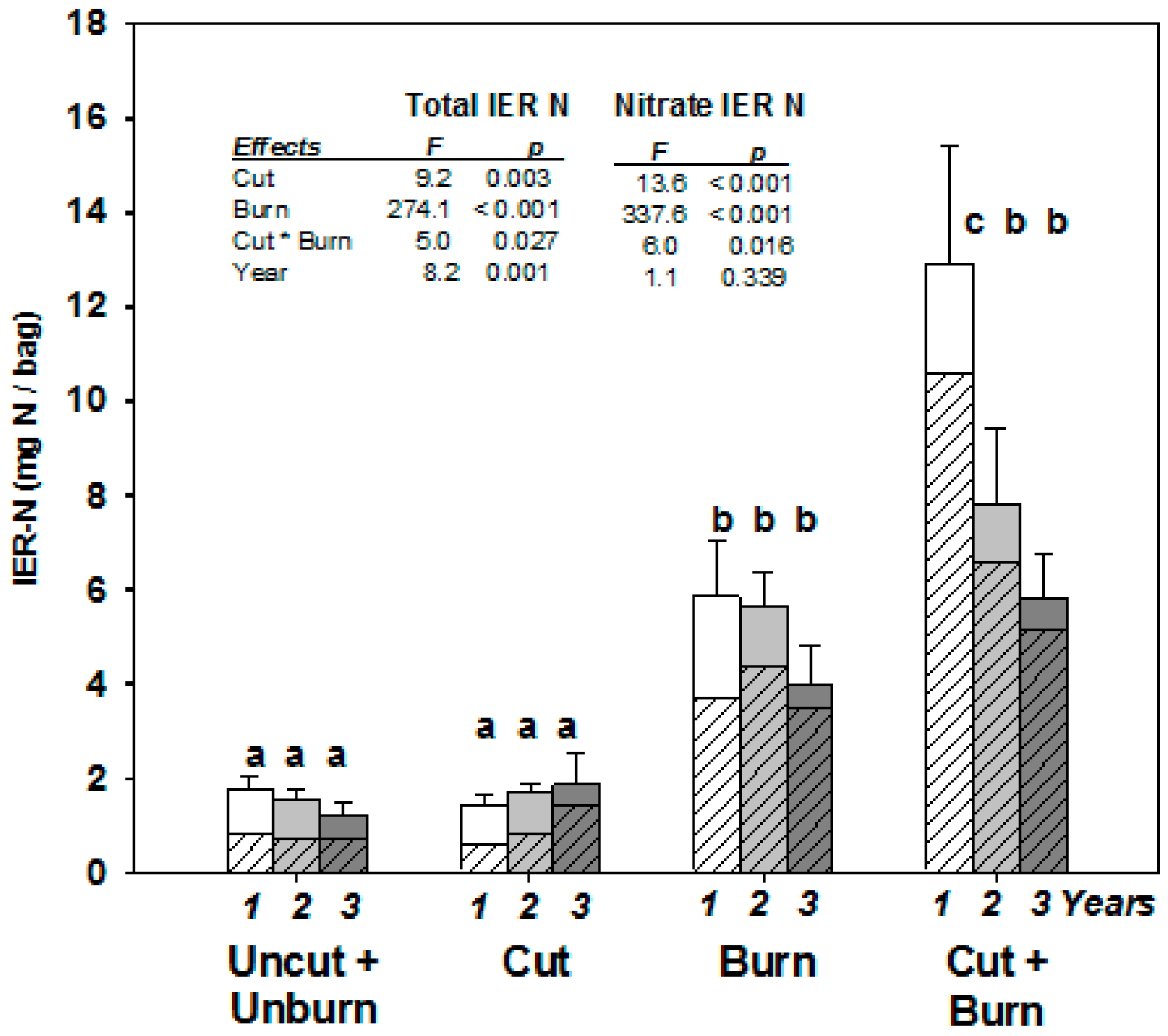
- Binkley, D.; Matson, P. Ion exchange resin bag method for assessing forest soil nitrogen availability. Soil Sci. Soc. Am. J. Abstr. 1983, 47, 1050–1052. [Google Scholar] [CrossRef]
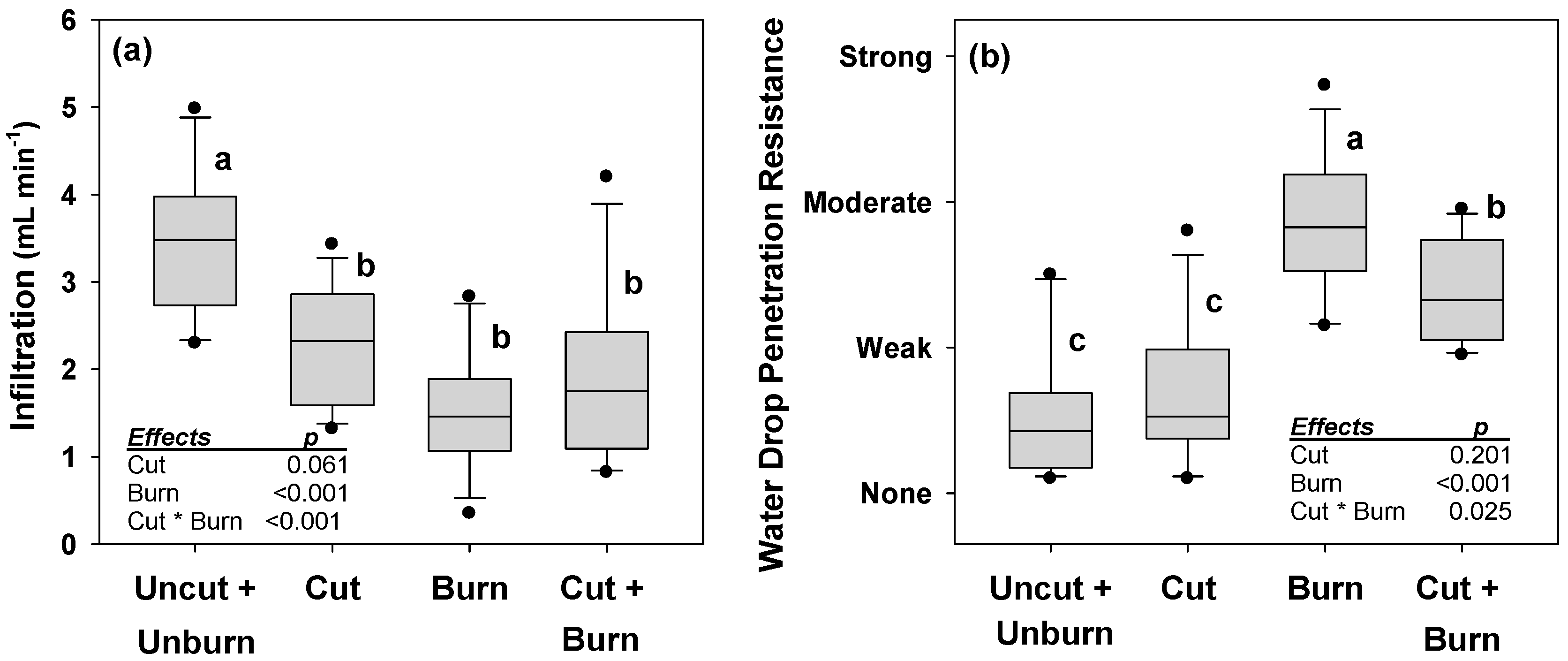
- DeBano, L.F. Water Repellent Soils: A State of the Art; USDA: Berkeley, CA, USA, 1981; p. 21.
- Harvey, B.J.; Donato, D.C.; Turner, M.G. High and dry: Post-fire tree seedling establishment in subalpine forests decreases with post-fire drought and large stand-replacing burn patches. Global Ecol. Biogeogr. 2016. [Google Scholar] [CrossRef]
- Lotan, J.E.; Brown, J.K.; Neuenschwander, L.F. Role of fire in lodgepole pine forests. In Lodgepole Pine: The Species and Its Management; Baumgartner, D.M., Krebill, R.G., Arnott, J.T., Weetman, G.F., Eds.; Washington State University: Spokane, WA, USA, 1985; pp. 133–152. [Google Scholar]
- Wright, M.; Rocca, M. Do post-fire mulching treatments affect regeneration in serotinous lodgepole pine. Fire Ecology 2017, 13. [Google Scholar] [CrossRef]
- Lotan, J.E. The role of cone serotiny in lodgepole pine forests. In Management of Lodgepole Pine Ecosystems: Symposium Proceedings; Washington State University, Cooperative Extension Service, College of Agriculture: Pullman, WA, USA, 1975; pp. 471–495. [Google Scholar]
- Schoennagel, T.; Turner, M.G.; Romme, W.H. The influence of fire interval and serotiny on postfire lodgepole pine density in Yellowstone National Park. Ecology 2003, 84, 2967–2978. [Google Scholar] [CrossRef]
- Teste, F.P.; Lieffers, V.J.; Landhäusser, S.M. Seed release in serotinous lodgepole pine forests after mountain pine beetle outbreak. Ecol. Appl. 2011, 21, 150–162. [Google Scholar] [CrossRef] [PubMed]
- Amman, G.D.; Baker, B.H. Mountain pine beetle influence on lodgepole pine stand structure. J. For. 1972, 70, 204–209. [Google Scholar]
- Page, W.G.; Jenkins, M.J. Mountain pine beetle-induced changes to selected lodgepole pine fuel complexes within the intermountain region. For. Sci. 2007, 53, 507–518. [Google Scholar]
- Berner, L.T.; Law, B.E.; Meddens, A.J.H.; Hicke, J.A. Tree mortality from fires, bark beetles, and timber harvest during a hot and dry decade in the western United States (2003–2012). Env. Res. Lett. 2017, 12, 065005. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Foster, D.R.; Franklin, J.F.; Hunter, M.L.; Noss, R.F.; Schmiegelow, F.A.; Perry, D. Salvage harvesting policies after natural disturbance. Science 2004, 80, 1303. [Google Scholar] [CrossRef] [PubMed]
- Leverkus, A.B.; Benayas, J.M.R.; Castro, J.; Boucher, D.; Brewer, S.; Collins, B.M.; Donato, D.; Fraver, S.; Kishchuk, B.E.; Lee, E.-J.; et al. Salvage logging effects on regulating and supporting ecosystem services—A systematic map. Forests 2018. under review. [Google Scholar]
- Jenkins, M.J.; Page, W.G.; Hebertson, E.G.; Alexander, M.E. Fuels and fire behavior dynamics in bark beetle-attacked forests in Western North America and implications for fire management. For. Ecol. Manag. 2012, 275, 23–34. [Google Scholar] [CrossRef]
- Collins, B.J.; Rhoades, C.C.; Battaglia, M.A.; Hubbard, R.M. The effects of bark beetle outbreaks on forest development, fuel loads and potential fire behavior in salvage logged and untreated lodgepole pine forests. For. Ecol. Manag. 2012, 284, 260–268. [Google Scholar] [CrossRef]
- Francos, M.; Pereira, P.; Mataix-Solera, J.; Arcenegui, V.; Alcañiz, M.; Úbeda, X. How clear-cutting affects fire severity and soil properties in a Mediterranean ecosystem. For. Ecol. Manag. 2018, 206, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Donato, D.C.; Fontaine, J.B.; Campbell, J.L.; Robinson, W.D.; Kauffman, J.B.; Law, B.E. Post-wildfire logging hinders regeneration and increases fire risk. Science 2006, 311, 352. [Google Scholar] [CrossRef] [PubMed]
- Rhoades, C.C.; Fornwalt, P.J.; Paschke, M.W.; Shanklin, A.; Jonas, J.L. Recovery of small pile burn scars in conifer forests of the Colorado Front Range. For. Ecol. Manag. 2015, 347, 180–187. [Google Scholar] [CrossRef]
- Clow, D.W.; Rhoades, C.; Briggs, J.; Caldwell, M.; Lewis, W.M. Responses of soil and water chemistry to mountain pine beetle induced tree mortality in Grand County, Colorado, USA. Appl. Geochem. 2011, 26, S174–S178. [Google Scholar] [CrossRef]
- Huber, C. Long lasting nitrate leaching after bark beetle attack in the highlands of the Bavarian Forest National Park. J. Env. Qual. 2005, 34, 1772–1779. [Google Scholar] [CrossRef] [PubMed]
- Rhoades, C.C.; McCutchan, J.H.; Cooper, L.A.; Clow, D.; Detmer, T.M.; Briggs, J.S.; Stednick, J.D.; Veblen, T.T.; Ertz, R.M.; Likens, G.E.; et al. Biogeochemistry of beetle-killed forests: Explaining a weak nitrate response. Proc. Natl. Acad. Sci. USA 2013, 110, 1756–1760. [Google Scholar] [CrossRef] [PubMed]
- USDA Forest Service (USFS). Revision of the Land Resource Management Plan. Arapaho and Roosevelt National Forests and Pawnee National Grassland, Fort Collins, Colo; USDA Forest Service (USFS): Denver, CO, USA, 1997.
- Crouch, G.L. Aspen regeneration after commercial clearcutting in southwestern Colorado. J. Forest. 1983, 81, 316–319. [Google Scholar]
- Bigler, C.; Kulakowski, D.; Veblen, T. Multiple disturbance interactions and drought influence fire severity in Rocky Mountain subalpine forests. Ecology 2005, 86, 3018–3029. [Google Scholar] [CrossRef]
- Abatzoglou, J.T.; Williams, A.P. Impact of anthropogenic climate change on wildfire across western US forests. Proc. Natl. Acad. Sci. USA 2016, 113, 11770–11775. [Google Scholar] [CrossRef] [PubMed]
- Stevens-Rumann, C.S.; Kemp, K.B.; Higuera, P.E.; Harvey, B.J.; Rother, M.T.; Donato, D.C.; Morgan, P.; Veblen, T.T. Evidence for declining forest resilience to wildfires under climate change. Ecol. Lett. 2017. [Google Scholar] [CrossRef] [PubMed]
- Natural Resources Conservation Service (NRCS), National Water and Climate Center, Snowtel Site: CO-335. 2017. Available online: https://wcc.sc.egov.usda.gov/nwcc/site?sitenum=335 (accessed on 6 December 2017).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Surface Cover | -------------------- Woody Fuel Cover -------------------- | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Year | Condition/Label | Organic | Mineral | 1 and 10-h | 100-h | 1000-h | Total Fuel | ||||||||||||
| 2012 | Uncut + Unburn (UU) | 86.9 | 2.3 | a | 2.4 | 0.8 | c | 15.0 | 1.9 | a | 1.5 | 0.4 | b | 5.8 | 1.5 | ab | 22.5 | 2.5 | b |
| Cut (C) | 71.4 | 3.5 | ab | 12.5 | 2.4 | c | 24.8 | 4.8 | a | 6.5 | 0.9 | a | 11.2 | 2.8 | a | 43.9 | 6.6 | a | |
| Burn (B) | 36.6 | 5.7 | c | 52.6 | 5.3 | a | 3.1 | 1.6 | b | 0.9 | 0.2 | b | 2.9 | 1.5 | b | 8.8 | 1.7 | b | |
| Cut + Burn (CB) | 55.5 | 5.7 | b | 31.9 | 5.4 | b | 4.1 | 1.6 | b | 1.2 | 0.3 | b | 5.4 | 1.4 | ab | 10.9 | 2.4 | b | |
| 2013 | Uncut + Unburn (UU) | 87.2 | 2.3 | a | 1.5 | 0.3 | c | 9.5 | 1.8 | ab | 0.9 | 0.3 | b | 7.3 | 1.5 | a | 18.4 | 1.8 | ab |
| Cut (C) | 59.0 | 4.2 | bc | 14.8 | 3.7 | bc | 15.1 | 2.6 | a | 4.5 | 0.7 | a | 10.7 | 3.2 | a | 31.2 | 5.3 | a | |
| Burn (B) | 47.8 | 4.9 | c | 44.6 | 5.5 | a | 3.0 | 1.2 | b | 1.2 | 0.3 | b | 3.1 | 1.4 | a | 9.6 | 1.5 | b | |
| Cut + Burn (CB) | 64.9 | 4.9 | bc | 27.1 | 5.5 | ab | 4.0 | 1.0 | b | 1.8 | 0.4 | b | 4.0 | 1.0 | a | 10.9 | 2.1 | b | |
| 2014 | Uncut + Unburn (UU) | 85.3 | 0.9 | a | 2.0 | 0.6 | c | 11.5 | 1.8 | b | 2.2 | 0.5 | b | 4.2 | 1.4 | ab | 20.1 | 1.9 | b |
| Cut (C) | 53.2 | 4.8 | b | 15.3 | 2.6 | c | 21.8 | 3.5 | a | 5.7 | 1.2 | a | 10.8 | 2.5 | a | 42.3 | 5.8 | a | |
| Burn (B) | 31.8 | 3.3 | c | 60.8 | 3.8 | a | 3.4 | 0.4 | c | 1.1 | 0.3 | b | 2.6 | 0.7 | b | 11.4 | 1.4 | b | |
| Cut + Burn (CB) | 44.3 | 4.9 | bc | 46.6 | 5.1 | b | 3.6 | 0.6 | c | 1.4 | 0.6 | b | 4.8 | 1.7 | ab | 10.7 | 2.2 | b | |
| Effects | F | p | F | p | F | p | F | p | F | p | F | p | |||||||
| Cut | 4.9 | 0.029 | 1.0 | 0.317 | 13.8 | <0.001 | 53.7 | <0.001 | 14.5 | <0.001 | 27.4 | <0.001 | |||||||
| Burn | 123.0 | <0.001 | 270.3 | <0.001 | 101.9 | <0.001 | 47.5 | <0.001 | 19.5 | <0.001 | 102.8 | <0.001 | |||||||
| Cut * Burn | 72.7 | <0.001 | 42.7 | <0.001 | 9.9 | 0.002 | 35.1 | <0.001 | 3.3 | 0.071 | 23.6 | <0.001 | |||||||
| Date | 8.5 | <0.001 | 6.0 | 0.004 | 3.1 | 0.051 | 1.01 | 0.368 | 0.2 | 0.800 | 1.9 | 0.151 | |||||||
| Year | Condition | Graminoid | Forb | Shrub | Total | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2012 | Uncut + Unburn | 43.7 | 4.7 | a | 34.8 | 4.3 | a | 39.7 | 3.7 | a | 118.2 | 5.9 | a |
| Cut | 33.6 | 4.2 | a | 16.8 | 5.6 | a | 12.9 | 2.4 | b | 63.2 | 7.4 | b | |
| Burn | 1.2 | 0.3 | b | 20.0 | 3.1 | a | 2.3 | 0.4 | c | 23.6 | 3.1 | c | |
| Cut + Burn | 13.9 | 3.4 | b | 35.3 | 7.4 | a | 0.7 | 0.3 | c | 49.8 | 9.2 | bc | |
| 2013 | Uncut + Unburn | 32.8 | 5.8 | a | 20.8 | 3.9 | ab | 38.2 | 3.0 | a | 91.8 | 8.5 | a |
| Cut | 26.2 | 3.5 | ab | 13.1 | 3.5 | b | 11.7 | 2.1 | b | 51.0 | 5.5 | b | |
| Burn | 3.9 | 1.2 | c | 28.9 | 4.4 | a | 4.6 | 0.9 | c | 37.5 | 3.9 | b | |
| Cut + Burn | 13.9 | 2.5 | bc | 30.0 | 4.3 | a | 1.8 | 0.6 | c | 45.6 | 4.5 | b | |
| 2014 | Uncut + Unburn | 33.3 | 3.9 | a | 37.5 | 5.1 | a | 39.8 | 3.2 | a | 110.5 | 7.4 | a |
| Cut | 27.1 | 4.3 | ab | 16.0 | 5.0 | b | 15.7 | 2.6 | b | 58.7 | 6.2 | b | |
| Burn | 6.1 | 1.3 | c | 39.0 | 4.7 | a | 6.3 | 1.1 | c | 51.3 | 5.4 | b | |
| Cut + Burn | 18.5 | 3.4 | bc | 32.7 | 3.6 | ab | 3.3 | 0.9 | c | 54.5 | 5.4 | b | |
| Effects | F | p | F | p | F | p | F | p | |||||
| Cut | 1.0 | 0.331 | 6.2 | 0.014 | 179.0 | <0.001 | 30.2 | <0.001 | |||||
| Burn | 150.4 | <0.001 | 11.2 | 0.001 | 466.2 | <0.001 | 111.5 | <0.001 | |||||
| Cut * Burn | 27.2 | <0.001 | 11.1 | 0.001 | 120.4 | <0.001 | 70.3 | <0.001 | |||||
| Date | 1.2 | 0.308 | 3.4 | 0.038 | 2.0 | 0.145 | 4.7 | 0.011 | |||||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rhoades, C.C.; Pelz, K.A.; Fornwalt, P.J.; Wolk, B.H.; Cheng, A.S. Overlapping Bark Beetle Outbreaks, Salvage Logging and Wildfire Restructure a Lodgepole Pine Ecosystem. Forests 2018, 9, 101. https://doi.org/10.3390/f9030101
Rhoades CC, Pelz KA, Fornwalt PJ, Wolk BH, Cheng AS. Overlapping Bark Beetle Outbreaks, Salvage Logging and Wildfire Restructure a Lodgepole Pine Ecosystem. Forests. 2018; 9(3):101. https://doi.org/10.3390/f9030101
Chicago/Turabian StyleRhoades, Charles C., Kristen A. Pelz, Paula J. Fornwalt, Brett H. Wolk, and Antony S. Cheng. 2018. "Overlapping Bark Beetle Outbreaks, Salvage Logging and Wildfire Restructure a Lodgepole Pine Ecosystem" Forests 9, no. 3: 101. https://doi.org/10.3390/f9030101
APA StyleRhoades, C. C., Pelz, K. A., Fornwalt, P. J., Wolk, B. H., & Cheng, A. S. (2018). Overlapping Bark Beetle Outbreaks, Salvage Logging and Wildfire Restructure a Lodgepole Pine Ecosystem. Forests, 9(3), 101. https://doi.org/10.3390/f9030101