Pine Plantations and Invasion Alter Fuel Structure and Potential Fire Behavior in a Patagonian Forest-Steppe Ecotone

 , ,
, ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Study Design
2.3. Fuel Characterization
2.4. Fire Behavior Modeling
2.5. Data Analysis
3. Results
3.1. Fuel Characterization
3.2. Fire Behavior Modeling
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Flannigan, M.D.; Amiro, B.D.; Logan, K.A.; Stocks, B.J.; Wotton, B.M. Forest fires and climate change in the 21ST century. Mitig. Adapt. Strateg. Glob. Chang. 2006, 11, 847–859. [Google Scholar] [CrossRef]
- Cochrane, M.A.; Barber, C.P. Climate change, human land use and future fires in the Amazon. Glob. Chang. Biol. 2009, 15, 601–612. [Google Scholar] [CrossRef]
- Veblen, T.T.; Kitzberger, T.; Donnegan, J. Climatic and human influences on fire regimes in Ponderosa pine forests in the Colorado Front Range. Ecol. Appl. 2000, 10, 1178–1195. [Google Scholar] [CrossRef]
- Blackhall, M.; Raffaele, E.; Paritsis, J.; Tiribelli, F.; Morales, J.M.; Kitzberger, T.; Gowda, J.H.; Veblen, T.T. Effects of biological legacies and herbivory on fuels and flammability traits: A long-term experimental study of alternative stable states. J. Ecol. 2017, 105, 1309–1322. [Google Scholar] [CrossRef]
- D’Antonio, C.M.; Hughes, R.F.; Tunison, J.T. Long-term impacts of invasive grasses and subsequent fire in seasonally dry Hawaiian woodlands. Ecol. Appl. 2011, 21, 1617–1628. [Google Scholar] [CrossRef] [PubMed]
- Brooks, M.L.; D’Antonio, C.M.; Richardson, D.M.; Grace, J.B.; Keeley, J.E.; DiTomaso, J.M.; Hobbs, R.J.; Pellant, M.; Pyke, D. Effects of invasive alien plants on fire regimes. BioScience 2004, 54, 677–688. [Google Scholar] [CrossRef]
- Mandle, L.; Bufford, J.L.; Schmidt, I.B.; Daehler, C.C. Woody exotic plant invasions and fire: Reciprocal impacts and consequences for native ecosystems. Biol. Invasions 2011, 13, 1815–1827. [Google Scholar] [CrossRef]
- Balch, J.K.; Bradley, B.A.; D’Antonio, C.M.; Gómez-Dans, J. Introduced annual grass increases regional fire activity across the arid western USA (1980–2009). Glob. Chang. Biol. 2013, 19, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Veblen, T.T.; Holz, A.; Paritsis, J.; Raffaele, E.; Kitzberger, T.; Blackhall, M. Adapting to global environmental change in Patagonia: What role for disturbance ecology? Austral Ecol. 2011, 36, 891–903. [Google Scholar] [CrossRef]
- Simberloff, D.; Nuñez, M.A.; Ledgard, N.J.; Pauchard, A.; Richardson, D.M.; Sarasola, M.; Van Wilgen, B.W.; Zalba, S.M.; Zenni, R.D.; Bustamante, R.; et al. Spread and impact of introduced conifers in South America: Lessons from other southern hemisphere regions. Austral Ecol. 2010, 35, 489–504. [Google Scholar] [CrossRef]
- Van Wilgen, B.W.; Richardson, D.M. The effects of alien shrub Invasions on vegetation structure and fire behaviour in South African fynbos shrublands: A simulation study. J. Appl. Ecol. 1985, 22, 955–966. [Google Scholar] [CrossRef]
- Nuñez, M.A.; Chiuffo, M.C.; Torres, A.; Paul, T.; Dimarco, R.D.; Raal, P.; Policelli, N.; Moyano, J.; García, R.A.; Van Wilgen, B.W.; et al. Ecology and management of invasive Pinaceae around the world: Progress and challenges. Biol. Invasions 2017, 19, 3099–3120. [Google Scholar] [CrossRef]
- Bava, J.O.; Logercio, G.A.; Salvador, G. Por qué plantar en Patagonia? Estado actual y el rol futuro de los bosques plantados. Ecol. Austral 2015, 21, 101–111. [Google Scholar]
- Richardson, D.M.; Van Wilgen, B.W.; Nuñez, M.A. Alien conifer invasions in South America: Short fuse burning? Biol. Invasions 2008, 10, 573–577. [Google Scholar] [CrossRef]
- Keeley, J.E. Ecology and evolution of pine life histories. Ann. For. Sci. 2012, 69, 445–453. [Google Scholar] [CrossRef]
- Taylor, K.T.; Maxwell, B.D.; McWethy, D.B.; Pauchard, A.; Nuñez, M.A.; Whitlock, C. Pinus contorta invasions increase wildfire fuel loads and may create a positive feedback with fire. Ecology 2017, 98, 678–687. [Google Scholar] [CrossRef] [PubMed]
- Gyenge, J.E.; Fernández, M.E.; Schlichter, T.M. Are differences in productivity between native and exotic trees in N.W. Patagonia related to differences in hydraulic conductance? Trees Struct. Funct. 2008, 22, 483–490. [Google Scholar] [CrossRef]
- Raffaele, E.; Nuñez, M.A.; Relva, M.A. Plantaciones de coníferas exóticas en Patagonia: Los riesgos de plantar sin un manejo adecuado. Ecol. Austral 2015, 25, 89–92. [Google Scholar]
- Godoy, M.M.; Defossé, G.E.; Bianchi, L.O.; Davel, M.M.; Withington, T.E. Fire-caused tree mortality in thinned Douglas-fir stands in Patagonia, Argentina. Int. J. Wildland Fire 2013, 22, 810–814. [Google Scholar] [CrossRef]
- Cóbar-Carranza, A.J.; García, R.A.; Pauchard, A.; Peña, E. Effect of Pinus contorta invasion on forest fuel properties and its potential implications on the fire regime of Araucaria araucana and Nothofagus antarctica forests. Biol. Invasions 2014, 16, 2273–2291. [Google Scholar] [CrossRef]
- Andrews, P.L. Current status and future needs of the BehavePlus Fire Modeling System. Int. J. Wildland Fire 2013, 23, 21–33. [Google Scholar] [CrossRef]
- Andrews, P.L.; Bevins, C.D.; Seli, R.C. BehavePlus Fire Modeling System, Version 4.0: User’s Guide (General Technical Report); USDA Forest Service. Rocky Mountain Research Station: Fort Collins, CO, USA, 2008.
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Ghermandi, L.; Guthmann, N.; Bran, D. Early post-fire succession in northwestern Patagonia grasslands. J. Veg. Sci. 2004, 15, 67–76. [Google Scholar] [CrossRef]
- Defossé, G.E. Conviene seguir fomentando las plantaciones forestales en el norte de la Patagonia? Ecol. Austral 2015, 25, 93–100. [Google Scholar]
- Nuñez, M.A.; Sarasola, M. Invasión de pinos: Una problemática para no descuidarse. Prod. For. 2014, 3, 15–17. [Google Scholar]
- Sarasola, M.M.; Rusch, V.E.; Schlichter, T.M.; Ghersa, C.M. Invasión de coníferas forestales en áreas de estepa y bosques de ciprés de la cordillera en la Región Andino Patagónica. Ecol. Austral 2006, 16, 143–156. [Google Scholar]
- Reiner, A.L.; Vaillant, N.M.; Dailey, S.N. Mastication and prescribed fire influences on tree mortality and predicted fire behavior in Ponderosa pine. West. J. Appl. For. 2012, 27, 36–41. [Google Scholar]
- Nelson, K.N.; Turner, M.G.; Romme, W.H.; Tinker, D.B. Simulated fire behaviour in young, postfire lodgepole pine forests. Int. J. Wildland Fire 2017, 26, 852–865. [Google Scholar] [CrossRef]
- Raffaele, E.; Schlichter, T. Efectos de las plantaciones de pino ponderosa sobre la heterogeneidad de micrositios en estepas del noroeste patagónico. Ecol. Austral 2000, 10, 151–158. [Google Scholar]
- Gosper, C.R.; Yates, C.J.; Prober, S.M.; Wiehl, G. Application and validation of visual fuel hazard assessments in dry Mediterranean-climate woodlands. Int. J. Wildland Fire 2014, 23, 385–393. [Google Scholar] [CrossRef]
- Scott, J.H.; Reinhardt, E.D. Stereo Photo Guide for Estimating Canopy Fuel Characteristics in Conifer Stands; General Technical Report RMRS-GTR-145; USDA Forest Service, Rocky Mountain Research Station: Ft. Collins, CO, USA, 2005.
- Qi, Y.; Jolly, W.M.; Dennison, P.E.; Kropp, R.C. Seasonal relationships between foliar moisture content, heat content and biochemistry of lodgepole line and big sagebrush foliage. Int. J. Wildland Fire 2016, 25, 574–578. [Google Scholar] [CrossRef]
- Rothermel, R.C. A Mathematical Model for Predicting Fire Spread in Wildland Fuels; INT-115 U.S. Forest Service Research Paper; USDA Forest Service: Ogden, UT, USA, 1972.
- Yospin, G.I.; Bridgham, S.D.; Kertis, J.; Johnson, B.R. Ecological correlates of fuel dynamics and potential fire behavior in former upland prairie and oak savanna. For. Ecol. Manag. 2012, 266, 54–65. [Google Scholar] [CrossRef]
- Diamond, J.M.; Call, C.A.; Devoe, N. Effects of targeted grazing and prescribed burning on community and seed dynamics of a downy brome (Bromus tectorum)–dominated landscape. Invasive Plant Sci. Manag. 2012, 5, 259–269. [Google Scholar] [CrossRef]
- Rothermel, R.C. How to Predict the Spread and Intensity of Forest and Range Fires; General Technical Report INT-143; USDA Forest Service: Ogden, UT, USA, 1983.
- Turner, M.G.; Baker, W.L.; Peterson, C.J.; Peet, R.K. Factors influencing succession: Lessons from large, infrequent natural disturbances. Ecosystems 1998, 1, 511–523. [Google Scholar] [CrossRef]
- Scott, J.H.; Burgan, R.E. Standard Fire Behavior Fuel Models: A Comprehensive Set for Use with Rothermel’s Surface Fire Spread Model; General Technical Report RMRS-GTR-153; USDA Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2005.
- Sagarzazu, M.S.; Defossé, G.E. Study and Analysis of Large Past Fires in Western Chubut Province, Patagonia, Argentina, Deliverable D8.3-1 of the Integrated project “Fire Paradox”, Project no. FP6-018505; European Commission: Brussels, Belgium, 2009; p. 82. [Google Scholar]
- Team, R.C. R: A Language and Environment for Statistical Computing, Version 3.4.1; R Foundation for Statistical Computing: Vienna, Austria, 2017; Available online: http://www.R-project.org (accessed on 4 September 2017).
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; Core-Team, R. Nlme: Linear and nonli Near Mixed Effects Models. R Package Version 3.1-128. 2016. Available online: https://cran.r-project.org/web/packages/nlme/ (accessed on 4 September 2017).
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Agee, J.K.; Skinner, C.N. Basic principles of forest fuel reduction treatments. For. Ecol. Manag. 2005, 211, 83–96. [Google Scholar] [CrossRef]
- Fulé, P.Z.; Crouse, J.E.; Roccaforte, J.P.; Kalies, E.L. Do thinning and/or burning treatments in western USA ponderosa or Jeffrey pine-dominated forests help restore natural fire behavior? For. Ecol. Manag. 2012, 269, 68–81. [Google Scholar] [CrossRef]
- Scott, J.H.; Reinhardt, E.D. Estimating canopy fuels in conifer forests. Fire Manag. Today 2002, 62, 45–50. [Google Scholar]
- Sarasola, M.M.; Diez, J.P.; Jaque, N.F.; Nuñez, M.A. Desarrollo de protocolos para la prevención, monitoreo y control de las invasiones de coníferas introducidas en el noroeste patagónico: Análisis de efectividad y costos. In Investigación Forestal 2011–2915 Los Proyectos de Investigación Aplicada; UCAR, Ministerio de Agroindustria: Buenos Aires, Argentina, 2016. [Google Scholar]
- González, S.; Ghermandi, L.; Pelaez, D. Growth and reproductive post-fire responses of two shrubs in semiarid Patagonian grasslands. Int. J. Wildland Fire 2015, 24, 809–818. [Google Scholar] [CrossRef]
- Nuñez, M.A.; Raffaele, E. Afforestation causes changes in post-fire regeneration in native shrubland communities of northwestern Patagonia, Argentina. J. Veg. Sci. 2007, 18, 827–834. [Google Scholar] [CrossRef]
- Franzese, J.; Raffaele, E. Fire as a driver of pine invasions in the Southern Hemisphere: A review. Biol. Invasions 2017, 19, 2237–2246. [Google Scholar] [CrossRef]
- Bravo-Monasterio, P.; Pauchard, A.; Fajardo, A. Pinus contorta invasion into treeless steppe reduces species richness and alters species traits of the local community. Biol. Invasions 2016, 18, 1883–1894. [Google Scholar] [CrossRef]
- Braun, A.C.; Troeger, D.; Garcia, R.; Aguayo, M.; Barra, R.; Vogt, J. Assessing the impact of plantation forestry on plant biodiversity: A comparison of sites in Central Chile and Chilean Patagonia. Glob. Ecol. Conserv. 2017, 10, 159–172. [Google Scholar] [CrossRef]
- Victoria Lantschner, M.; Rusch, V.; Hayes John, P. Influences of pine plantations on small mammal assemblages of the Patagonian forest-steppe ecotone. Mammalia 2011, 75, 24–255. [Google Scholar] [CrossRef]
- Premoli, A.; Quiroga, P.; Gardner, M. Araucaria Araucana. The IUCN Red List of Threatened Species 2013: e.T31355A2805113. 2013. Available online: http://dx.doi.org/10.2305/IUCN.UK.2013-1.RLTS.T31355A2805113.en (accessed on 10 October 2017).
- Liu, Y.; Stanturf, J.; Goodrick, S. Trends in global wildfire potential in a changing climate. For. Ecol. Manag. 2010, 259, 685–697. [Google Scholar] [CrossRef]
- Holz, A.; Paritsis, J.; Mundo, I.A.; Veblen, T.T.; Kitzberger, T.; Williamson, G.J.; Aráoz, E.; Bustos-Schindler, C.; González, M.E.; Grau, H.R.; et al. Southern Annular Mode drives multicentury wildfire activity in southern South America. Proc. Natl. Acad. Sci. USA 2017, 114, 9552–9557. [Google Scholar] [CrossRef] [PubMed]
- De Torres Curth, M.D.; Biscayart, C.; Ghermandi, L.; Pfister, G. Wildland-urban interface fires and socioeconomic conditions: A case study of a northwestern Patagonia city. Environ. Manag. 2012, 49, 876–891. [Google Scholar] [CrossRef] [PubMed]
- Van Wilgen, B.W.; Forsyth, G.G.; Prins, P. The management of fire-adapted ecosystems in an urban setting: The case of Table Mountain National Park, South Africa. Ecol. Soc. 2012, 17. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
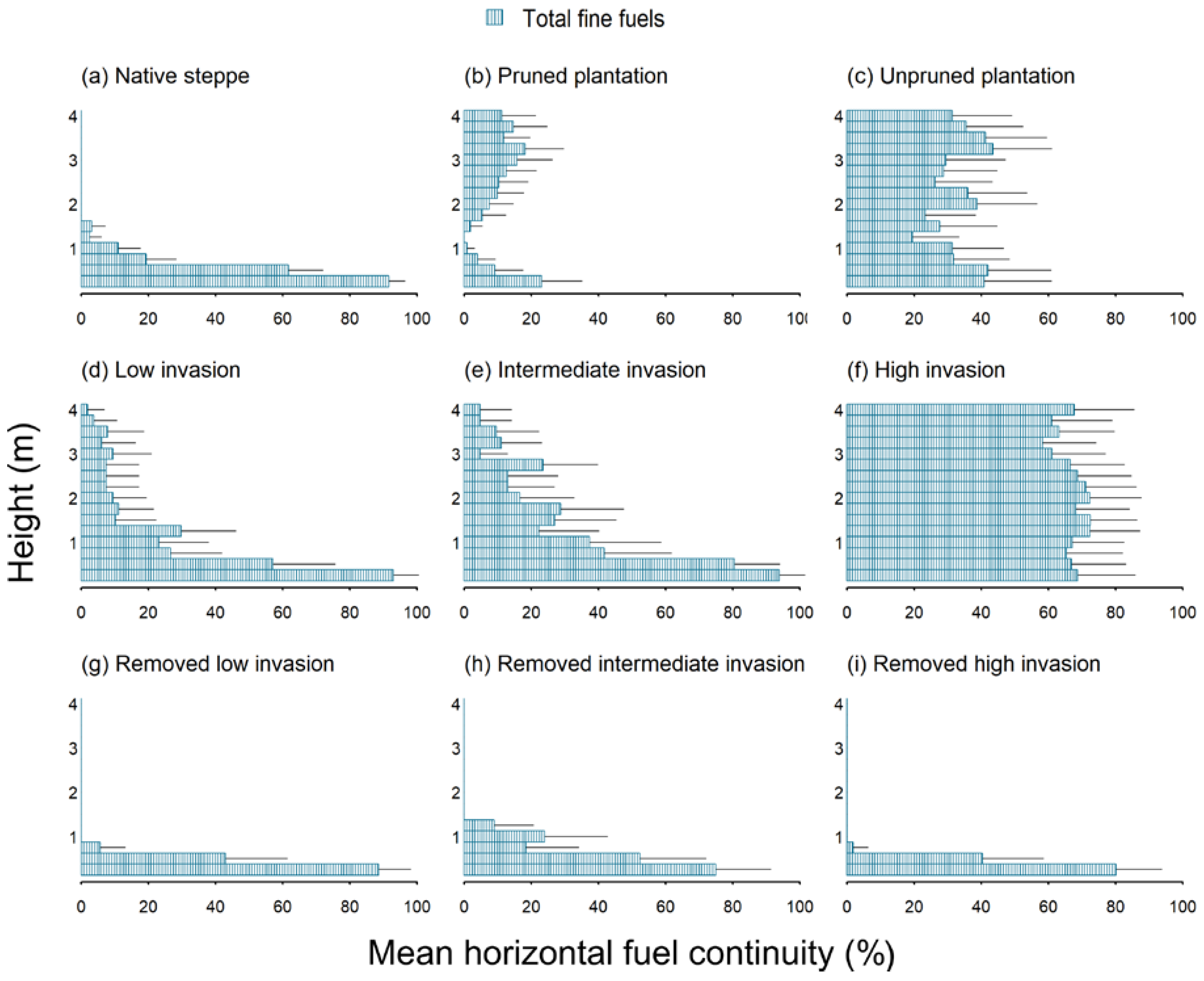{kind=link}
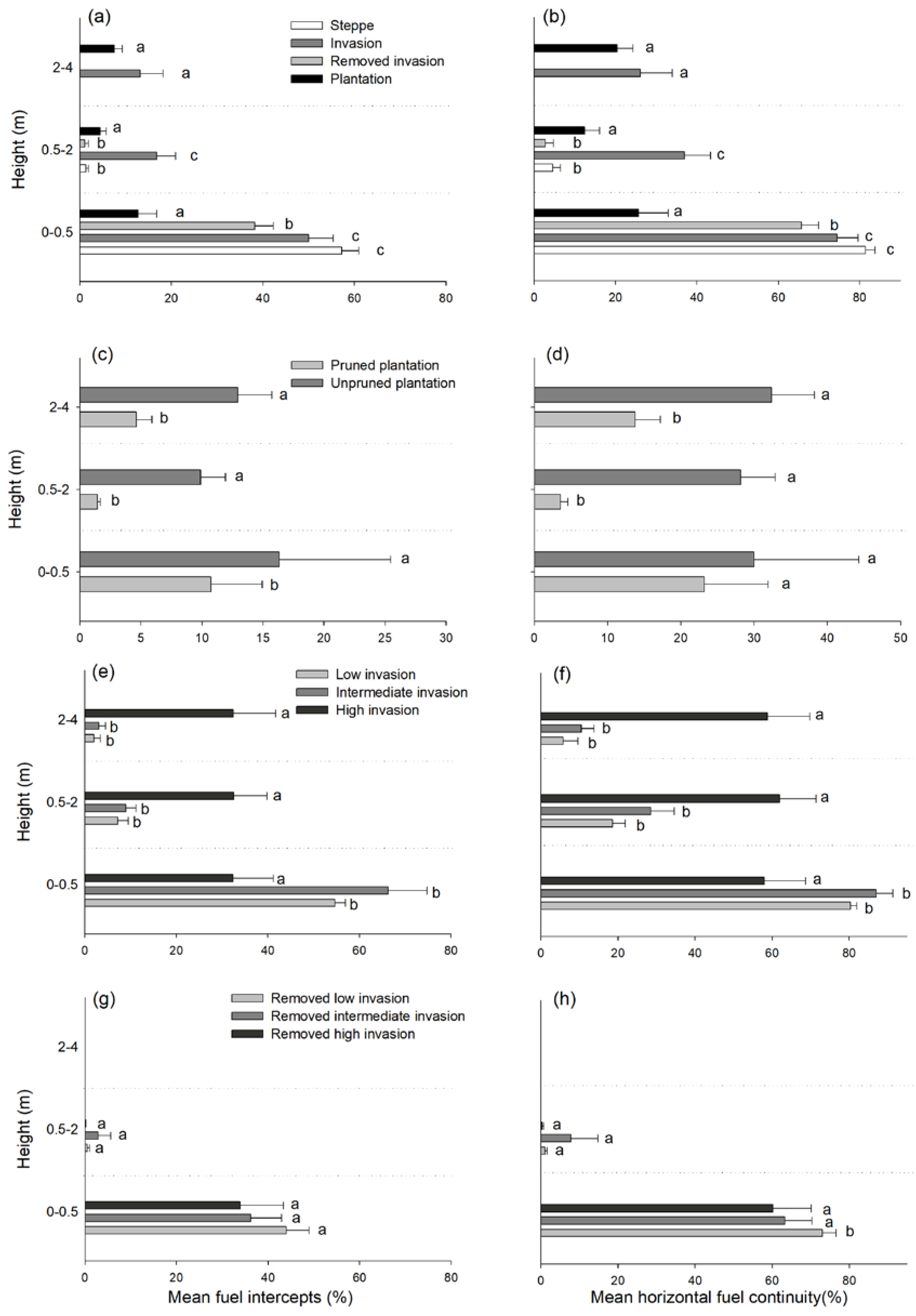{kind=link}
| Vegetation Condition | Control | Low Invasion | Intermediate Invasion | High Invasion | Pruned | Unpruned | Total |
|---|---|---|---|---|---|---|---|
| Native steppe | 14 | - | - | - | - | - | 14 |
| Pine plantation | - | - | - | - | 9 | 5 | 14 |
| Pine invasion | - | 5 | 4 | 5 | - | - | 14 |
| Removed pine invasion | - | 5 | 4 | 5 | - | - | 14 |
| Site/fuel variables | Steppe | Pruned Plantation | Unprunned Plantation | High Invasion | Removed High Invasion |
|---|---|---|---|---|---|
| Dominant Species | M. spinosum/P. speciosa/A. splendens | P. ponderosa | P. ponderosa | P. contorta | P. speciosa |
| Tree age (years) | - | ~30 | ~26 | ~3 to 29 | - |
| Tree density (ind/ha) | - | 1220 | 1330 | 8400 | - |
| Basal Area (m²/ha) | - | 85.4 | 59.7 | 25.1 | - |
| 1 h fuel load (ton/ha) | 0.45 | 1.61 | 1.41 | 1.22 | 0.96 |
| 10 h fuel load (ton/ha) | 0 | 0.31 | 0.45 | 0.05 | 0.09 |
| 100 h fuel load (ton/ha) | 0 | 0 | 0 | 0 | 0 |
| Live herbaceous fuel load (ton/ha) | 0.32 | 0 | 0 | 0.03 | 0.08 |
| Live woody fuel load (ton/ha) | 0.61 | 0 | 0 | 0.02 | 0 |
| Fuel bed depth (m) | 0.35 | 0.11 | 0.09 | 0.05 | 0.08 |
| Canopy height (m) | - | 13.8 | 12.3 | 11.2 | - |
| Canopy base height (m) | - | 3.25 | 1.62 | 0.27 | - |
| Canopy bulk density (kg/m³) | - | 0.16 | 0.16 | 0.18 | - |
| Mid-season live woody moisture (%) | 80 | 100 | 100 | 100 | - |
| Mid-season foliar/herbaceous moisture (%) | 140/62/nd | 135 | 135 | 135 | 62 |
| Fire Behavior Variables | Steppe | Pruned Plantation | Unpruned Plantation | High Invasion | Removed High Invasion | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| High | Extreme | High | Extreme | High | Extreme | High | Extreme | High | Extreme | |
| Surface rate of spread (m/min) | 5.9 | 11.2 | 16.6 | 38.8 | 11.4 | 24 | 7.4 | 17.1 | 7.7 | 16.5 |
| Surface flame length (m) | 0.6 | 1.0 | 1.6 | 2.6 | 1.3 | 2.0 | 0.9 | 1.6 | 0.6 | 1.4 |
| Transition ratio to crown fire | - | - | 0.49 | 1.45 | 0.89 | 2.35 | 6.66 | 20.19 | - | - |
| Crown rate of spread (m/min) | - | - | 9.9 | 32.1 | 9.9 | 32.1 | 9.9 | 32.1 | - | - |
| Surface heat per unit area (kJ/m2) | 1033 | 1331 | 2549 | 3264 | 2306 | 2946 | 1870 | 2461 | 1478 | 1906 |
| Canopy heat per unit area (kJ/m2) | 0 | 0 | 31435 | 31435 | 31822 | 31822 | 36638 | 36638 | 0 | 0 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paritsis, J.; Landesmann, J.B.; Kitzberger, T.; Tiribelli, F.; Sasal, Y.; Quintero, C.; Dimarco, R.D.; Barrios-García, M.N.; Iglesias, A.L.; Diez, J.P.; et al. Pine Plantations and Invasion Alter Fuel Structure and Potential Fire Behavior in a Patagonian Forest-Steppe Ecotone. Forests 2018, 9, 117. https://doi.org/10.3390/f9030117
Paritsis J, Landesmann JB, Kitzberger T, Tiribelli F, Sasal Y, Quintero C, Dimarco RD, Barrios-García MN, Iglesias AL, Diez JP, et al. Pine Plantations and Invasion Alter Fuel Structure and Potential Fire Behavior in a Patagonian Forest-Steppe Ecotone. Forests. 2018; 9(3):117. https://doi.org/10.3390/f9030117
Chicago/Turabian StyleParitsis, Juan, Jennifer B. Landesmann, Thomas Kitzberger, Florencia Tiribelli, Yamila Sasal, Carolina Quintero, Romina D. Dimarco, María N. Barrios-García, Aimé L. Iglesias, Juan P. Diez, and et al. 2018. "Pine Plantations and Invasion Alter Fuel Structure and Potential Fire Behavior in a Patagonian Forest-Steppe Ecotone" Forests 9, no. 3: 117. https://doi.org/10.3390/f9030117
APA StyleParitsis, J., Landesmann, J. B., Kitzberger, T., Tiribelli, F., Sasal, Y., Quintero, C., Dimarco, R. D., Barrios-García, M. N., Iglesias, A. L., Diez, J. P., Sarasola, M., & Nuñez, M. A. (2018). Pine Plantations and Invasion Alter Fuel Structure and Potential Fire Behavior in a Patagonian Forest-Steppe Ecotone. Forests, 9(3), 117. https://doi.org/10.3390/f9030117