Ecological Factors Influencing Norway Spruce Regeneration on Nurse Logs in a Subalpine Virgin Forest



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Experimental Design and Measured Variables
2.3. Data Analysis
2.4. Detection and Identification of Wood-Decomposing Basidiomycetes
3. Results
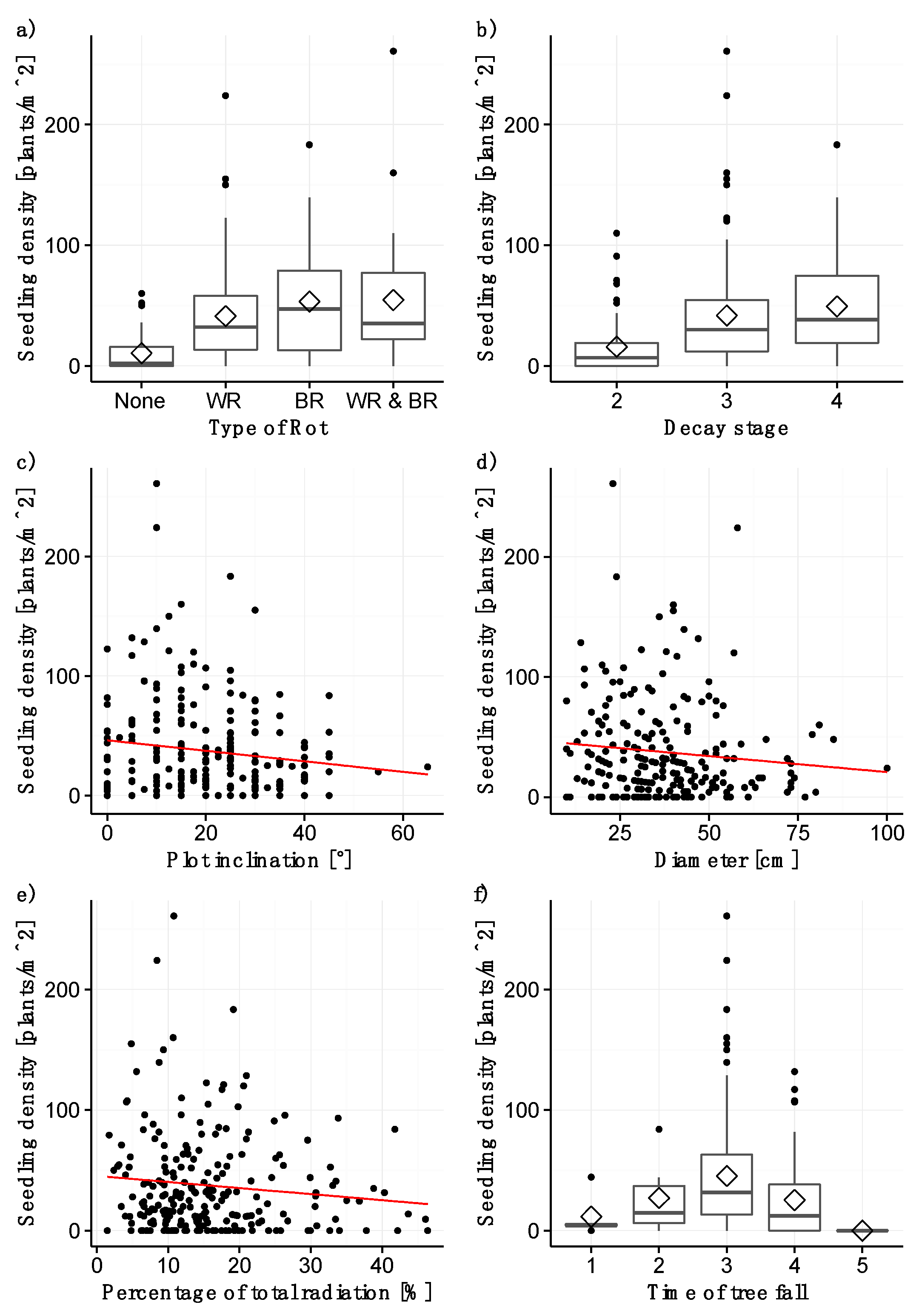
3.1. Seedling Density
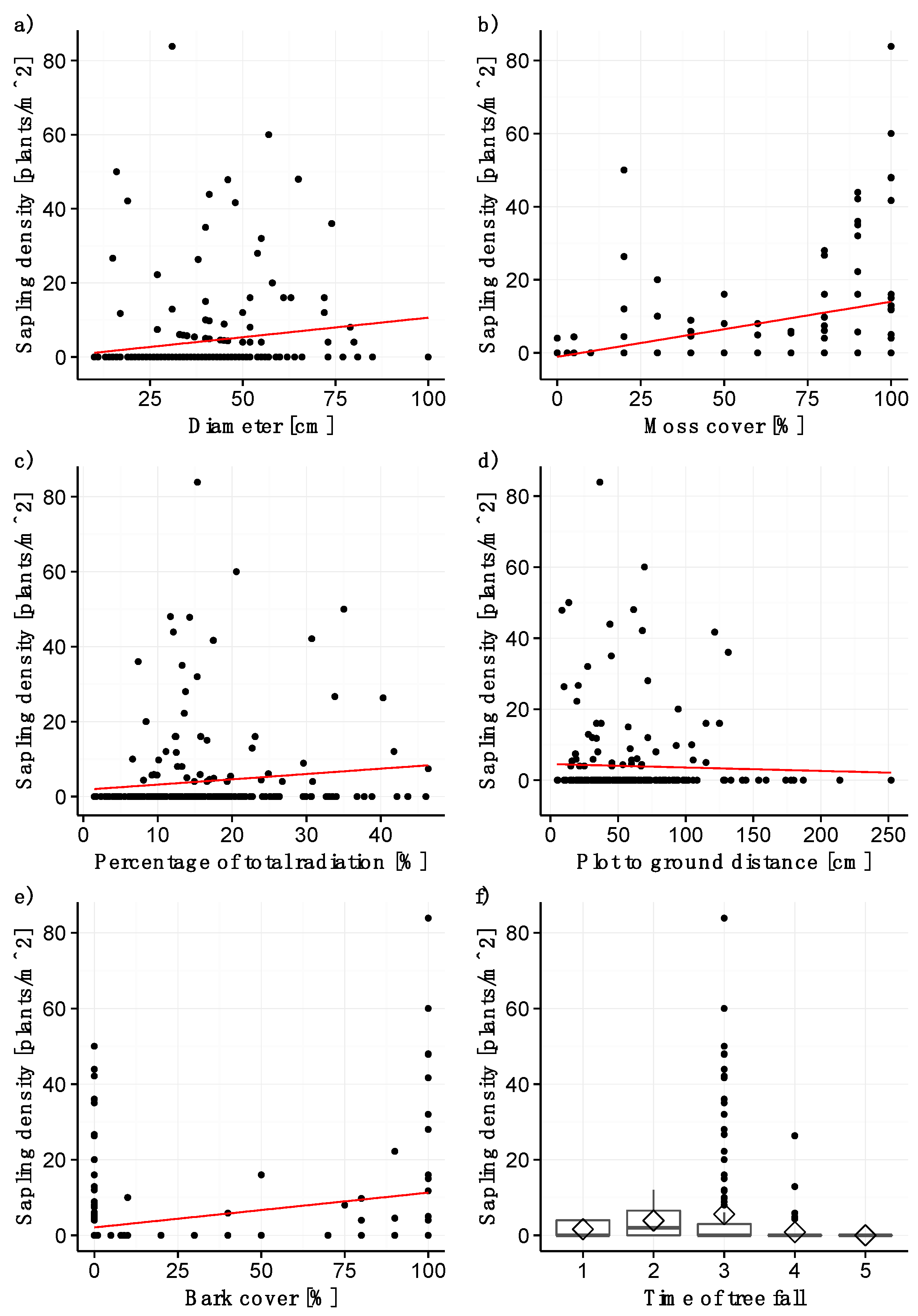
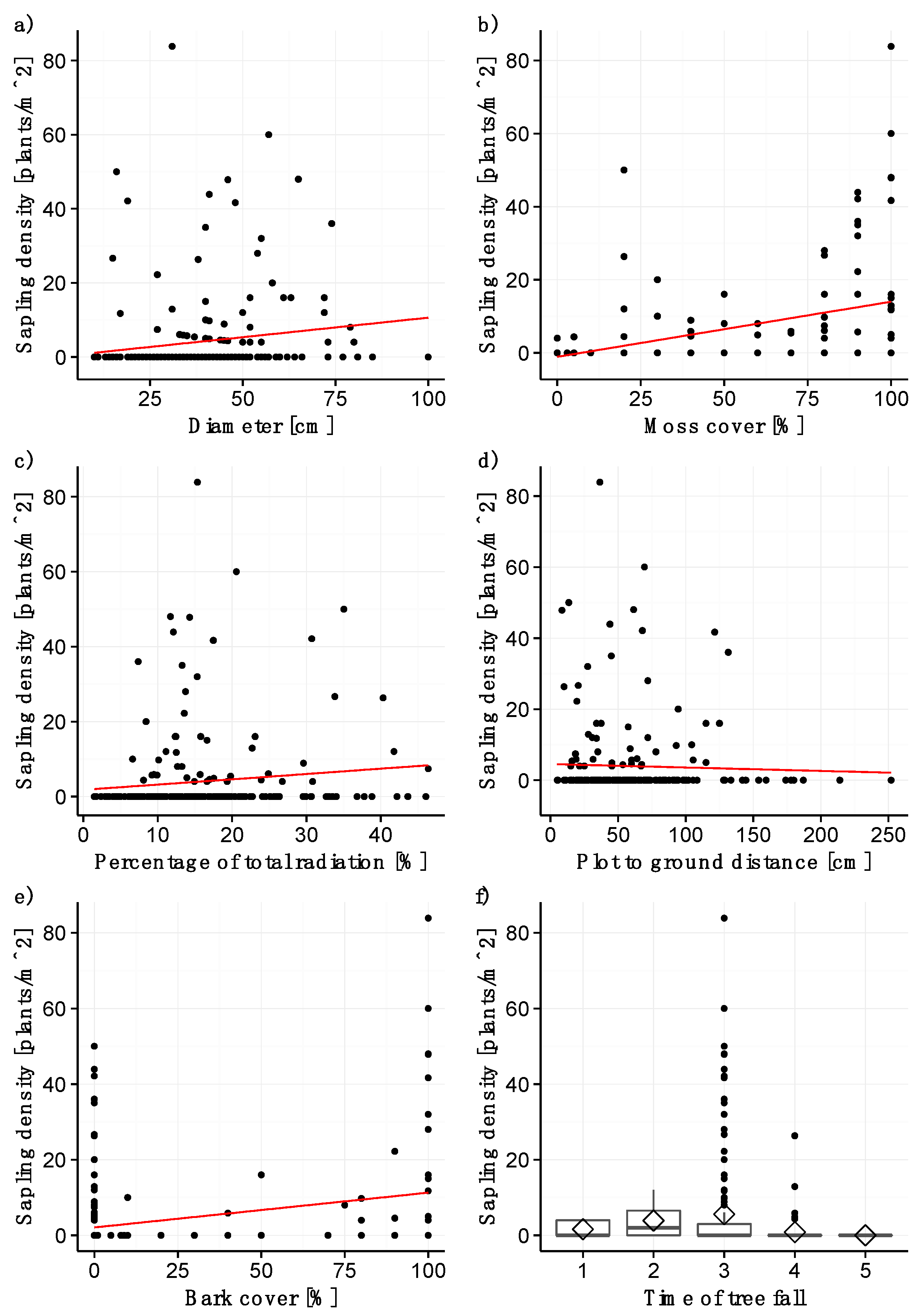
3.2. Sapling Density
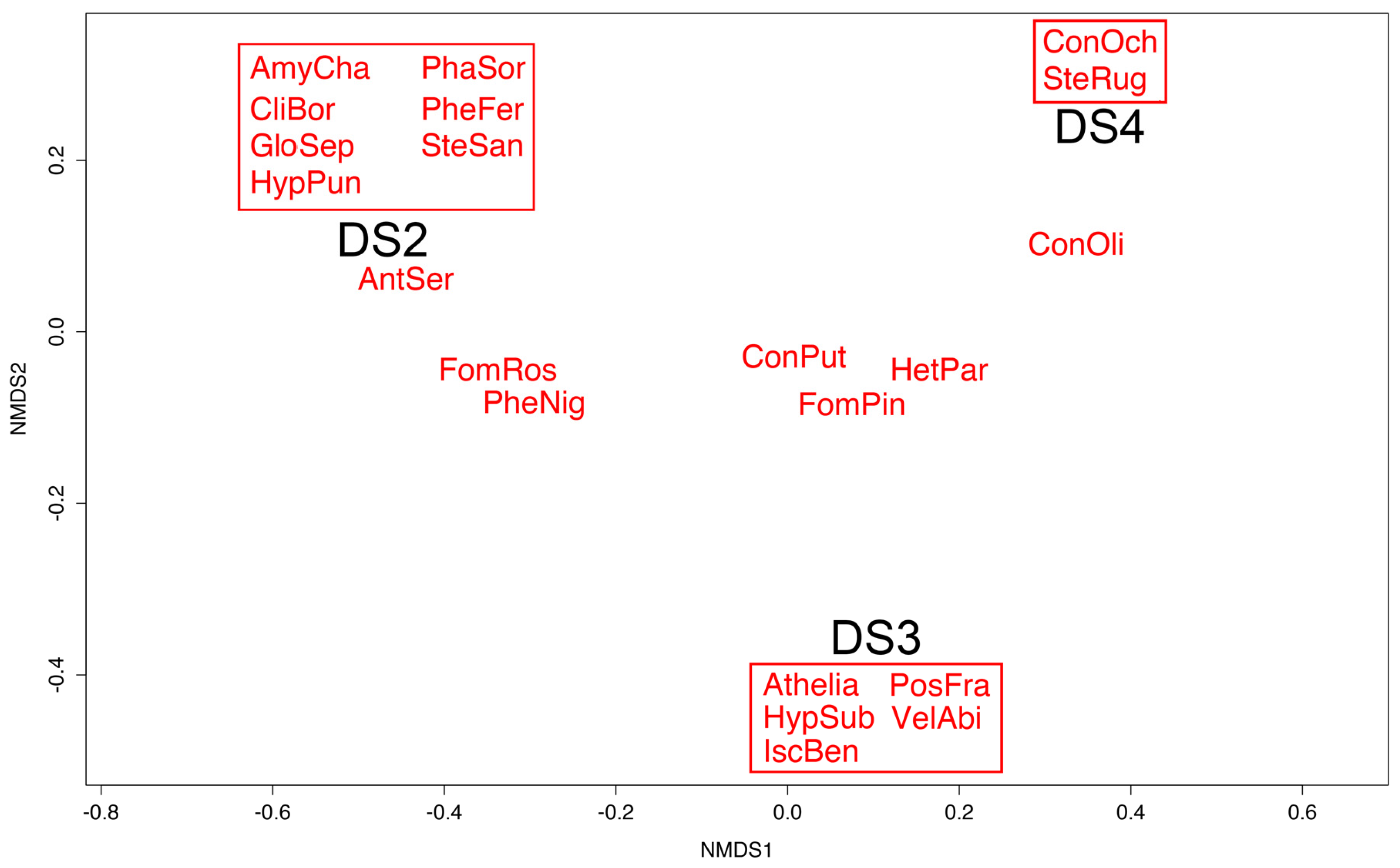
3.3. Polypore Diversity and Decay Stage
4. Discussion
4.1. Factors with a Significant Effect on Both Seedling and Sapling Density
4.2. Factors with a Significant Effect on Seedling Density
4.3. Factors with a Significant Effect on Sapling Density
4.4. Polypore Diversity and Decay Stage
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wohlgemuth, T.; Kull, P.; Wüthrich, H. Disturbance of microsites and early tree regeneration after windthrow in Swiss mountain forests due to the winter storm Vivian 1990. For. Snow Landsc. Res. 2002, 77, 17–47. [Google Scholar]
- Eichrodt, R. Über die Bedeutung von Moderholz für die Natürliche Waldverjüngung im Subalpinen Fichtenwald; Beiheft zu den Zeitschriften des Schweizerischen Forstvereins; Bühler Buchdruck: Zurich, Switzerland, 1970; Volume 45. [Google Scholar]
- Ott, E.; Frehner, M.; Frey, H.-U.; Lüscher, P. Gebirgsnadelwälder; Verlag Paul Haupt Berne: Bern, Switzerland; Stuttgart, Germany; Wien, Austria, 1997; ISBN 3-258-05601-3. [Google Scholar]
- Brang, P. Resistance and elasticity: Promising concepts for the management of protection forests in the European Alps. For. Ecol. Manag. 2001, 145, 107–119. [Google Scholar] [CrossRef]
- Landolt, E. Unsere Alpenflora, 7th ed.; SAC-Verlag: Bern, Switzerland, 2003; ISBN 3-85902-218-0. [Google Scholar]
- Leibundgut, H. Die Natürliche Waldverjüngung; Verlag Paul Haupt Berne: Bern, Switzerland; Stuttgart, Germany, 1984; ISBN 3-258-03415-X. [Google Scholar]
- Svoboda, M.; Fraver, S.; Janda, P.; Bače, R.; Zenáhlíková, J. Natural development and regeneration of a Central European montane spruce forest. For. Ecol. Manag. 2010, 260, 707–714. [Google Scholar] [CrossRef]
- Takahashi, K. Effect of size structure, forest floor type and disturbance regime on tree species composition in a coniferous forest in Japan. J. Ecol. 1994, 82, 769–773. [Google Scholar] [CrossRef]
- Harmon, M.E.; Franklin, J.F.; Swanson, F.J.; Sollins, P.; Gregory, S.V.; Lattin, J.D.; Anderson, N.H.; Cline, S.P.; Aumen, N.G.; Sedell, J.R.; et al. Ecology of coarse woody debris in temperate ecosystems. In Advances in Ecological Research; Elsevier: Amsterdam, The Netherlands, 1986; Volume 15, pp. 133–302. ISBN 978-0-12-013915-6. [Google Scholar]
- Kupferschmid, A.D.; Bugmann, H. Effect of microsites, logs and ungulate browsing on Picea abies regeneration in a mountain forest. For. Ecol. Manag. 2005, 205, 251–265. [Google Scholar] [CrossRef]
- Zielonka, T. When does dead wood turn into a substrate for spruce replacement? J. Veg. Sci. 2006, 17, 739–746. [Google Scholar] [CrossRef]
- Szewczyk, J.; Szwagrzyk, J. Tree regeneration on rotten wood and on soil in old-growth stand. Vegetatio 1996, 122, 37–46. [Google Scholar] [CrossRef]
- Jonsson, B.G.; Hofgaard, A. The structure and regeneration of high-altitude Norway spruce forests: A review of Arnborg (1942, 1943). Scand. J. For. Res. 2011, 26, 17–24. [Google Scholar] [CrossRef]
- Anderson, L.J.; Winterton, A.J. Germination as a determinant of seedling distributions among natural substrates in Picea engelmannii (Pinaceae) and Abies lasiocarpa (Pinaceae). Am. J. Bot. 1996, 83, 112–117. [Google Scholar] [CrossRef]
- Simard, M.-J.; Bergeron, Y.; Sirois, L. Conifer seedling recruitment in a southeastern Canadian boreal forest: The importance of substrate. J. Veg. Sci. 1998, 9, 575–582. [Google Scholar] [CrossRef]
- Narukawa, Y.; Yamamoto, S. Development of conifer seedlings roots on soil and fallen logs in boreal and subalpine coniferous forests of Japan. For. Ecol. Manag. 2003, 175, 131–139. [Google Scholar] [CrossRef]
- Narukawa, Y.; Iida, S.; Tanouchi, H.; Abe, S.; Yamamoto, S. State of fallen logs and the occurrence of conifer seedlings and saplings in boreal and subalpine old-growth forests of central Japan. Ecol. Res. 2003, 18, 267–277. [Google Scholar] [CrossRef]
- Staniaszek-Kik, M.; Żarnowiec, J.; Chmura, D. The vascular plant colonization on decaying Picea abies logs in Karkonosze mountain forest belts: The effects of forest community type, cryptogam cover, log decomposition and forest management. Eur. J. For. Res. 2016, 135, 1145–1157. [Google Scholar] [CrossRef]
- Zielonka, T.; Piątek, G. Norway spruce regeneration on decaying logs in subalpine forests in the Tatra National Park. Pol. Bot. J. 2001, 46, 251–260. [Google Scholar]
- Heinemann, K.; Kitzberger, T. Effects of position, understorey vegetation and coarse woody debris on tree regeneration in two environmentally contrasting forests of north-western Patagonia: A manipulative approach. J. Biogeogr. 2006, 33, 1357–1367. [Google Scholar] [CrossRef]
- Chmura, D.; Żarnowiec, J.; Staniaszek-Kik, M. Interactions between plant traits and environmental factors within and among montane forest belts: A study of vascular species colonising decaying logs. For. Ecol. Manag. 2016, 379, 216–225. [Google Scholar] [CrossRef]
- Kathke, S.; Bruelheide, H. Interaction of gap age and microsite type for the regeneration of Picea abies. For. Ecol. Manag. 2010, 259, 1597–1605. [Google Scholar] [CrossRef]
- Bače, R.; Svoboda, M.; Pouska, V.; Janda, P.; Červenka, J. Natural regeneration in Central-European subalpine spruce forests: Which logs are suitable for seedling recruitment? For. Ecol. Manag. 2012, 266, 254–262. [Google Scholar] [CrossRef]
- Hillgarter, F. Waldbauliche und Ertragskundliche Untersuchungen im Subalpinen Fichtenurwald Scatlè/Brigels. Ph.D. Thesis, ETH Zurich, Zurich, Switzerland, 1971. [Google Scholar]
- AWN, G. Naturwaldreservat Scatlè. 2011. Available online: http://www.webcitation.org/6x4wxntEI (accessed on 14 December 2012).
- Bugmann, H. The Forest Reserve “Scatlè”: A Brief Overview; ETH Zurich: Zurich, Switzerland, 2010. [Google Scholar]
- Heiri, C.; Hallenbarter, D. Der Urwald Scatlè. In Waldreservate. 50 Jahre Natürliche Waldentwicklung in der Schweiz; Brang, P., Heiri, C., Bugmann, H., Eds.; Haupt: Bern, Switzerland; Stuttgart, Germany; Wien, Austria, 2011; ISBN 978-3-258-07725-3. [Google Scholar]
- Renvall, P. Community structure and dynamics of wood-rotting Basidiomycetes on decomposing co-nifer trunks in northern Finland. Karstenia 1995, 35, 1–52. [Google Scholar] [CrossRef]
- Stahel, W. Statistische Datenanalyse, 5th ed.; Vieweg + Teubner: Wiesbaden, Germany, 2008; ISBN 978-3-8348-0410-5. [Google Scholar]
- Zuur, A.F.; Savaliev, A.A.; Ieno, E.N. Zero Inflated Models and Generalized Linear Mixed Models with R; Highland Statistics Ltd.: Newburgh, NY, USA, 2012; ISBN 0-9571741-1-X. [Google Scholar]
- Dormann, C.F. Parametrische Statistik. In Verteilungen, Maximum Likelihood and GLM in R; Springer: Berlin, Germany, 2017. [Google Scholar]
- Rohner, B.; Bigler, C.; Wunder, J.; Brang, P.; Bugmann, H. Fifty years of natural succession in Swiss forest reserves: Changes in stand structure and mortality rates of oak and beech. J. Veg. Sci. 2012, 23, 892–905. [Google Scholar] [CrossRef]
- Frazer, G. Gap Light Analyzer (GLA): Imaging Software to Extract Canopy Structure and Gap Light Transmission Indices from True-Colour Fisheye Photographs. Users Manual and Program Documentation, version 2.0; Simon Fraser University: Burnaby, BC, Canada; Institute of Ecosystem Studies: Millbrook, NY, USA, 1999. [Google Scholar]
- Sieber, T.N. Pyrenochaeta ligni-putridi sp. nov., a new coelomycete associated with butt rot of Picea abies in Switzerland. Mycol. Res. 1995, 99, 274–276. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press, Inc.: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Queloz, V.; Grünig, C.R.; Berndt, R.; Kowalski, T.; Sieber, T.N.; Holdenrieder, O. Cryptic speciation in Hymenoscyphus albidus. For. Pathol. 2011, 41, 133–142. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Breitenbach, J.; Kränzlin, F. Fungi of Switzerland—Non Gilled Fungi; Mycologia: Lucerne, Switzerland, 1986; Volume 2, ISBN 3-85604-220-2. [Google Scholar]
- Dittrich, S.; Jacob, M.; Bade, C.; Leuschner, C.; Hauck, M. The significance of deadwood for total bryophyte, lichen, and vascular plant diversity in an old-growth spruce forest. Plant Ecol. 2014, 215, 1123–1137. [Google Scholar] [CrossRef]
- Iijima, H.; Shibuya, M.; Saito, H. Effects of surface and light conditions of fallen logs on the emergence and survival of coniferous seedlings and saplings. J. For. Res. 2007, 12, 262–269. [Google Scholar] [CrossRef]
- Zielonka, T.; Piątek, G. The herb and dwarf shrubs colonization of decaying logs in subalpine forest in the Polish Tatra Mountains. Plant Ecol. 2004, 172, 63–72. [Google Scholar] [CrossRef]
- Christy, E.J.; Mack, R.N. Variation in demography of juvenile Tsuga heterophylla across the substratum mosaic. J. Ecol. 1984, 72, 75–91. [Google Scholar] [CrossRef]
- Kimmins, J. Forest Ecology: A Foundation for Sustainable Forest Management and Environmental Ethics in Forestry, 3rd ed.; Pearson Prentice Hall: London, NY, USA, 2004; ISBN 0-13-066258-5. [Google Scholar]
- Schmidt-Vogt, H. Die Fichte. Band 1: Taxonomie, Verbreitung, Morphologie, Ökologie, Waldgesellschaften; Paul Parey: Hamburg/Berlin, Germany, 1987; ISBN 3-490-08216-8. [Google Scholar]
- Pisek, A.; Winkler, E. Licht- und Temperaturabhängigkeit der CO2-Assimilation von Fichte (Picea excelsa Link), Zirbe (Pinus cembra L.) und Sonnenblume (Helianthus annuus L.). Planta 1959, 53, 532–550. [Google Scholar] [CrossRef]
- Harmon, M.E. Effects of bark fragmentation on plant succession on conifer logs in the Picea-Tsuga forests of Olympic National Park, Washington. Am. Midl. Nat. 1989, 121, 112–124. [Google Scholar] [CrossRef]
- Wasser, B.; Frehner, M. Wegleitung Minimale Pflegemassnahmen für Wälder mit Schutzfunktion; Bundesamt für Umwelt, Wald und Landschaft BUWAL: Bern, Switzerland, 1996. [Google Scholar]
- Ostrofsky, A.; Jellison, J.; Smith, K.T.; Shortle, W.C. Changes in cation concentrations in red spruce wood decayed by brown rot and white rot fungi. Can. J. For. Res. 1997, 27, 567–571. [Google Scholar] [CrossRef]
- Fukasawa, Y. Effects of wood decomposer fungi on tree seedling establishment on coarse woody debris. For. Ecol. Manag. 2012, 266, 232–238. [Google Scholar] [CrossRef]
- Harvey, A.E.; Jurgensen, M.F.; Larsen, M.J. Seasonal distribution of ectomycorrhizae in a mature Douglas-fir/Larch forest soil in western Montana. For. Sci. 1978, 24, 203–208. [Google Scholar]
- Tedersoo, L.; May, T.W.; Smith, M.E. Ectomycorrhizal lifestyle in fungi: Global diversity, distribution, and evolution of phylogenetic lineages. Mycorrhiza 2010, 20, 217–263. [Google Scholar] [CrossRef] [PubMed]
- Červenka, J.; Bače, R.; Svoboda, M. Stand-replacing disturbance does not directly alter the succession of Norway spruce regeneration on dead wood. J. For. Sci. 2014, 60, 417–424. [Google Scholar] [CrossRef]
- Takahashi, M.; Sakai, Y.; Ootomo, R.; Shiozaki, M. Establishment of tree seedlings and water-soluble nutrients in coarse woody debris in an old-growth Picea abies forest in Hokkaido, northern Japan. Can. J. For. Res. 2000, 30, 1148–1155. [Google Scholar] [CrossRef]
- Laiho, R.; Prescott, C.E. Decay and nutrient dynamics of coarse woody debris in northern coniferous forests: A synthesis. Can. J. For. Res. 2004, 34, 763–777. [Google Scholar] [CrossRef]
- Robert, E.; Brais, S.; Harvey, B.D.; Greene, D. Seedling establishment and survival on decaying logs in boreal mixedwood stands following a mast year. Can. J. For. Res. 2012, 42, 1446–1455. [Google Scholar] [CrossRef]
- Sugita, H.; Nagaike, T. Microsites for seedling establishment of subalpine conifers in a forest with moss-type undergrowth on Mt. Fuji, central Honshu, Japan. Ecol. Res. 2005, 20, 678–685. [Google Scholar] [CrossRef]
- Chmura, D.; Zarnowiec, J.; Staniaszek-Kik, M. Comparison of traits of non-colonized and colonized decaying logs by vascular plant species. IForest-Biogeosci. For. 2018, 11, 11–16. [Google Scholar] [CrossRef]
- Nakamura, T. Effect of bryophytes on survival of conifer seedlings in subalpine forests of central Japan. Ecol. Res. 1992, 7, 155–162. [Google Scholar] [CrossRef]
- Doi, Y.; Mori, A.S.; Takeda, H. Conifer establishment and root architectural responses to forest floor heterogeneity in an old-growth subalpine forest in central Japan. For. Ecol. Manag. 2008, 255, 1472–1478. [Google Scholar] [CrossRef]
- Holeksa, J.; Zielonka, T.; Żywiec, M. Modeling the decay of coarse woody debris in a subalpine Norway spruce forest of the West Carpathians, Poland. Can. J. For. Res. 2008, 38, 415–428. [Google Scholar] [CrossRef]
- Botting, R.S.; DeLong, C. Macrolichen and bryophyte responses to coarse woody debris characteristics in sub-boreal spruce forest. For. Ecol. Manag. 2009, 258, S85–S94. [Google Scholar] [CrossRef]
- Allmér, J.; Vasiliauskas, R.; Ihrmark, K.; Stenlid, J.; Dahlberg, A. Wood-inhabiting fungal communities in woody debris of Norway spruce (Picea abies (L.) Karst.), as reflected by sporocarps, mycelial isolations and T-RFLP identification: Wood-inhabiting fungal communities in woody debris of Norway spruce. FEMS Microbiol. Ecol. 2006, 55, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Rajala, T.; Peltoniemi, M.; Pennanen, T.; Mäkipää, R. Fungal community dynamics in relation to substrate quality of decaying Norway spruce (Picea abies (L.) Karst.) logs in boreal forests. FEMS Microbiol. Ecol. 2012, 81, 494–505. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, J.M.; Work, T.T. Linking deadwood-associated beetles and fungi with wood decomposition rates in managed black spruce forests. Can. J. For. Res. 2012, 42, 1477–1490. [Google Scholar] [CrossRef]
- Schmidt, O. Wood and Tree Fungi; Springer: Berlin/Heidelberg, Germany, 2006; ISBN 978-3-540-32138-5. [Google Scholar]
- Lindblad, I. Wood-inhabiting fungi on fallen logs of Norway spruce: Relations to forest management and substrate quality. Nord. J. Bot. 1998, 18, 243–255. [Google Scholar] [CrossRef]
- Penttilä, R.; Siitonen, J.; Kuusinen, M. Polypore diversity in managed and old-growth boreal Picea abies forests in southern Finland. Biol. Conserv. 2004, 117, 271–283. [Google Scholar] [CrossRef]
- Jönsson, M.T.; Edman, M.; Jonsson, B.G. Colonization and extinction patterns of wood-decaying fungi in a boreal old-growth Picea abies forest: Colonization-extinction dynamics of wood-decaying fungi. J. Ecol. 2008, 96, 1065–1075. [Google Scholar] [CrossRef]
- Høiland, K.; Bendiksen, E. Biodiversity of wood-inhabiting fungi in a boreal coniferous forest in Sør-Trøndelag County, Central Norway. Nord. J. Bot. 1996, 16, 643–659. [Google Scholar] [CrossRef]
- Petrillo, M.; Cherubini, P.; Sartori, G.; Abiven, S.; Ascher, J.; Bertoldi, D.; Camin, F.; Barbero, A.; Larcher, R.; Egli, M. Decomposition of Norway spruce and European larch coarse woody debris (CWD) in relation to different elevation and exposure in an Alpine setting. iForest-Biogeosci. For. 2016, 9, 154–164. [Google Scholar] [CrossRef]
- Lombardi, F.; Cherubini, P.; Tognetti, R.; Cocozza, C.; Lasserre, B.; Marchetti, M. Investigating biochemical processes to assess deadwood decay of beech and silver fir in Mediterranean mountain forests. Ann. For. Sci. 2013, 70, 101–111. [Google Scholar] [CrossRef]
- Bütler, R.; Patty, L.; Le Bayon, R.-C.; Guenat, C.; Schlaepfer, R. Log decay of Picea abies in the Swiss Jura Mountains of central Europe. For. Ecol. Manag. 2007, 242, 791–799. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Variable | Unit | Minimum | Median | Mean | Maximum |
|---|---|---|---|---|---|
| Average sunshine duration in June | Min/day | 1.47 | 76.07 | 95.63 | 361.19 |
| Bark cover | % | 0 | 0 | 20.74 | 100 |
| Vegetation coverage | % | 0 | 0 | 9.34 | 130 |
| Canopy openness | % | 1.22 | 9.76 | 9.95 | 23.7 |
| Log diameter | cm | 10 | 26 | 37.23 | 100 |
| Moss cover | % | 0 | 20 | 33.73 | 100 |
| Moss height | cm | 0 | 1.5 | 1.63 | 8 |
| No. of individuals | Count | 0 | 6 | 8.99 | 61 |
| No. of saplings >10 cm | Count | 0 | 0 | 0.79 | 15 |
| No. of seedlings | Count | 0 | 4 | 6.3 | 56 |
| Plot inclination | ° | 0 | 20 | 19.99 | 65 |
| Plot to ground distance | cm | 5 | 50 | 57.87 | 252 |
| Tallest individual | cm | 0 | 2.3 | 7.71 | 135 |
| Total radiation | % | 1.47 | 13.95 | 15.68 | 46.38 |
| Seedling Density | Sapling Density | |||
|---|---|---|---|---|
| Deviance | 275.1 | 170.9 | ||
| Performance | r = 0.62 | r = 0.72 | ||
| Robustness | r = 0.21 | r = 0.66 | ||
| Variable | p | Estimate | p | Estimate |
| White rot | 8.08 × 10−7 (***) | 7.217 × 10−1 | - | - |
| Brown rot | 1.26 × 10−5 (***) | 7.897 × 10−1 | - | - |
| White & brown rot | 3.13 × 10−7 (***) | 8.995 × 10−1 | - | - |
| Decay stage 3 | 0.0266 (*) | 2.898 × 10−1 | - | - |
| Decay stage 4 | 0.0043 (**) | 3.982 × 10−1 | - | - |
| Plot inclination | 0.0004 (***) | −1.118 × 10−1 | - | - |
| Diameter | 0.0002 (***) | 8.969 × 10−1 | 9.62 × 10−10 (***) | 3.687 × 10−1 |
| Moss cover | - | <2 × 10−16 (***) | 3.230 × 10−1 | |
| % of total radiation | 0.0689 (.) | −8.986 × 10−1 | 1.68 × 10−8 (***) | 7.601 × 10−1 |
| Time of tree fall: 1977–1989 | 0.1275 (n.s.) | 5.988 × 10−1 | 0.0031 (**) | 2.189 |
| Bark cover | - | - | 0.0288 (*) | 5.419 × 10−1 |
| Distance plot to ground | - | - | 0.0030 (**) | −9.195 × 10−1 |
| Species | Abbrev. | Identity | GenBank Accession Number of Closest Match | GenBank Accession Number | Isolation Frequency | Fruiting Body Frequency | Type of Rot |
|---|---|---|---|---|---|---|---|
| Amylostereum chailletii | AmyCha | 1 | GQ422470 | KT943921 | 2 | - | unknown |
| Amylostereum aerolatum | AmyAer | - | - | - | - | 1 | white |
| Antrodia serialis | AntSer | 0.99 | FM872469 | KT943942 KT943943 | 16 | - | brown |
| Athelia sp. | - | 0.98 | GU187537 | KT943944 | 1 | - | unknown |
| Climacocystis borealis | CliBor | 0.99 | JN710527 | KT943915 | 3 | - | white |
| Conferticium ochraceum | ConOch | 0.99 | AY781254 | KT943933 | 1 | - | unknown |
| Coniophora olivacea | ConOli | 0.99 | AJ345009 | KT943916 | 2 | - | brown |
| Coniophora puteana | ConPut | 1 | AM946631 | KT943922 KT943923 KT943924 | 7 | - | brown |
| Fomitopsis pinicola | FomPin | 1 | JX501306 | KT943917 KT943918 KT943919 KT943920 | 33 | 14 | brown |
| Fomitopsis rosea | FomRos | 0.99 | DQ491410 | KT943945 KT943946 | 11 | 8 | brown |
| Gloeophyllum sepiarium | GloSep | 1 | JN182924 | KT943931 | 1 | 2 | brown |
| Heterobasidion parviporum | HetPar | 1 | FJ903330 | KT943925 KT943926 | 5 | - | white |
| Hypochnicium punctulatum | HypPun | 1 | AF429412 | KT943936 | 1 | - | unknown |
| Hypochnicium subrigescens | HypSub | 0.99 | AF429427 | KT943939 | 1 | - | unknown |
| Ischnoderma benzoinum | IscBen | 1 | JQ518274 | KT943932 | 1 | - | white |
| Phanerochaete sordida | PhaSor | 1 | FJ228210 | KT943938 KT943940 | 4 | - | unknown |
| Phellinus ferrugineovelutinus | PheFer | 0.98 | AY558618 | KT943948 | 2 | - | unknown |
| Phellopilus nigrolimitatus | PheNig | 0.95 | JQ358813 | KT943927 KT943928 | 3 | - | white |
| Postia fragilis | PosFra | 1 | JF950573 | KT943930 | 1 | - | brown |
| Stereum rugosum | SteRug | 1 | FN539052 | KT943935 | 1 | - | white |
| Stereum sanguinolentum | SteSan | 0.99 | EU673086 | KT943941 | 2 | 1 | white |
| Trichaptum abietinum | TriAbi | - | - | - | - | 2 | white |
| Veluticeps abietina | VelAbi | 0.99 | EU118619 | KT943929 | 1 | - | unknown |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stroheker, S.; Weiss, M.; Sieber, T.N.; Bugmann, H. Ecological Factors Influencing Norway Spruce Regeneration on Nurse Logs in a Subalpine Virgin Forest. Forests 2018, 9, 120. https://doi.org/10.3390/f9030120
Stroheker S, Weiss M, Sieber TN, Bugmann H. Ecological Factors Influencing Norway Spruce Regeneration on Nurse Logs in a Subalpine Virgin Forest. Forests. 2018; 9(3):120. https://doi.org/10.3390/f9030120
Chicago/Turabian StyleStroheker, Sophie, Marc Weiss, Thomas N. Sieber, and Harald Bugmann. 2018. "Ecological Factors Influencing Norway Spruce Regeneration on Nurse Logs in a Subalpine Virgin Forest" Forests 9, no. 3: 120. https://doi.org/10.3390/f9030120
APA StyleStroheker, S., Weiss, M., Sieber, T. N., & Bugmann, H. (2018). Ecological Factors Influencing Norway Spruce Regeneration on Nurse Logs in a Subalpine Virgin Forest. Forests, 9(3), 120. https://doi.org/10.3390/f9030120