Effect of Vertical Canopy Architecture on Transpiration, Thermoregulation and Carbon Assimilation



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Theory
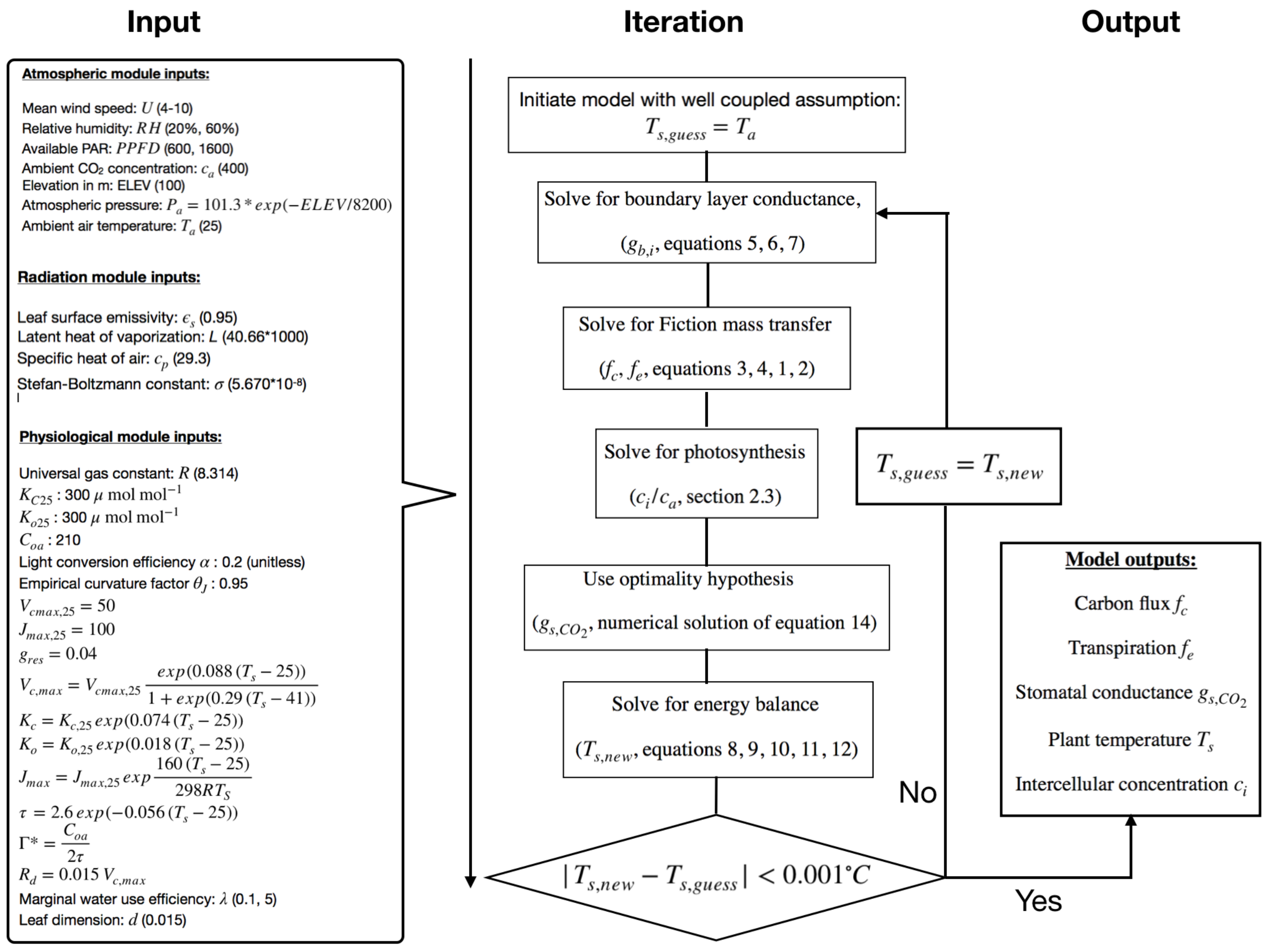
2.1. Fickian Mass Transfer Model
2.2. Boundary Layer Conductance
2.3. Photosynthesis Model
2.4. Leaf Level Energy Balance
2.5. Stomatal Optimization Model
2.6. Algorithm
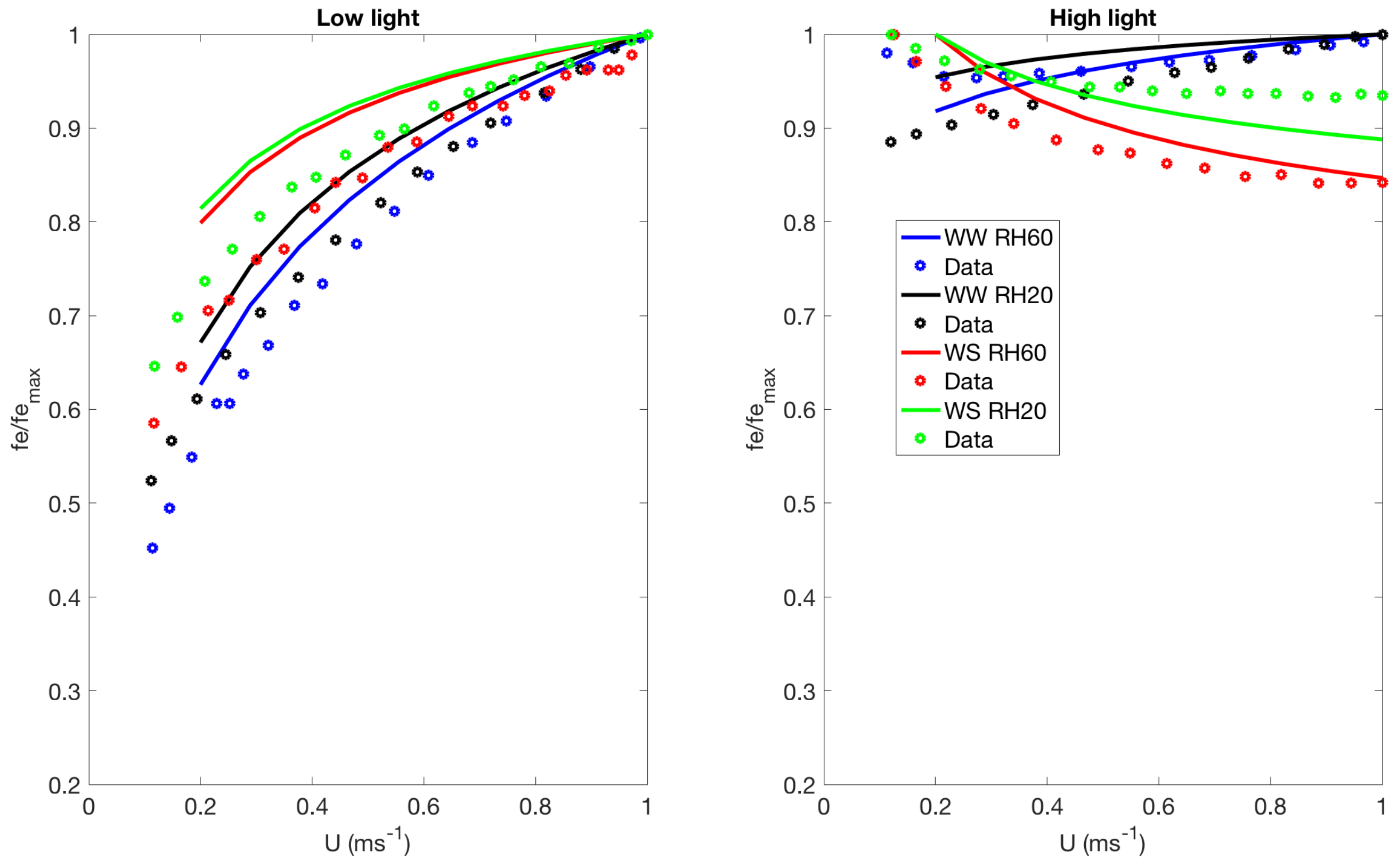
- WW RH20: well watered soil () and high evaporative demand in the atmosphere (relative humidity (RH) = );
- WW RH60: well watered soil () and low evaporative demand in the atmosphere (relative humidity (RH) = );
- WS RH20: water stressed soil () and high evaporative demand in the atmosphere (relative humidity (RH) = );
- WS RH60: water stressed soil () and low evaporative demand in the atmosphere (relative humidity (RH) = ).
3. Results and Discussions
3.1. Comparison with Published Results
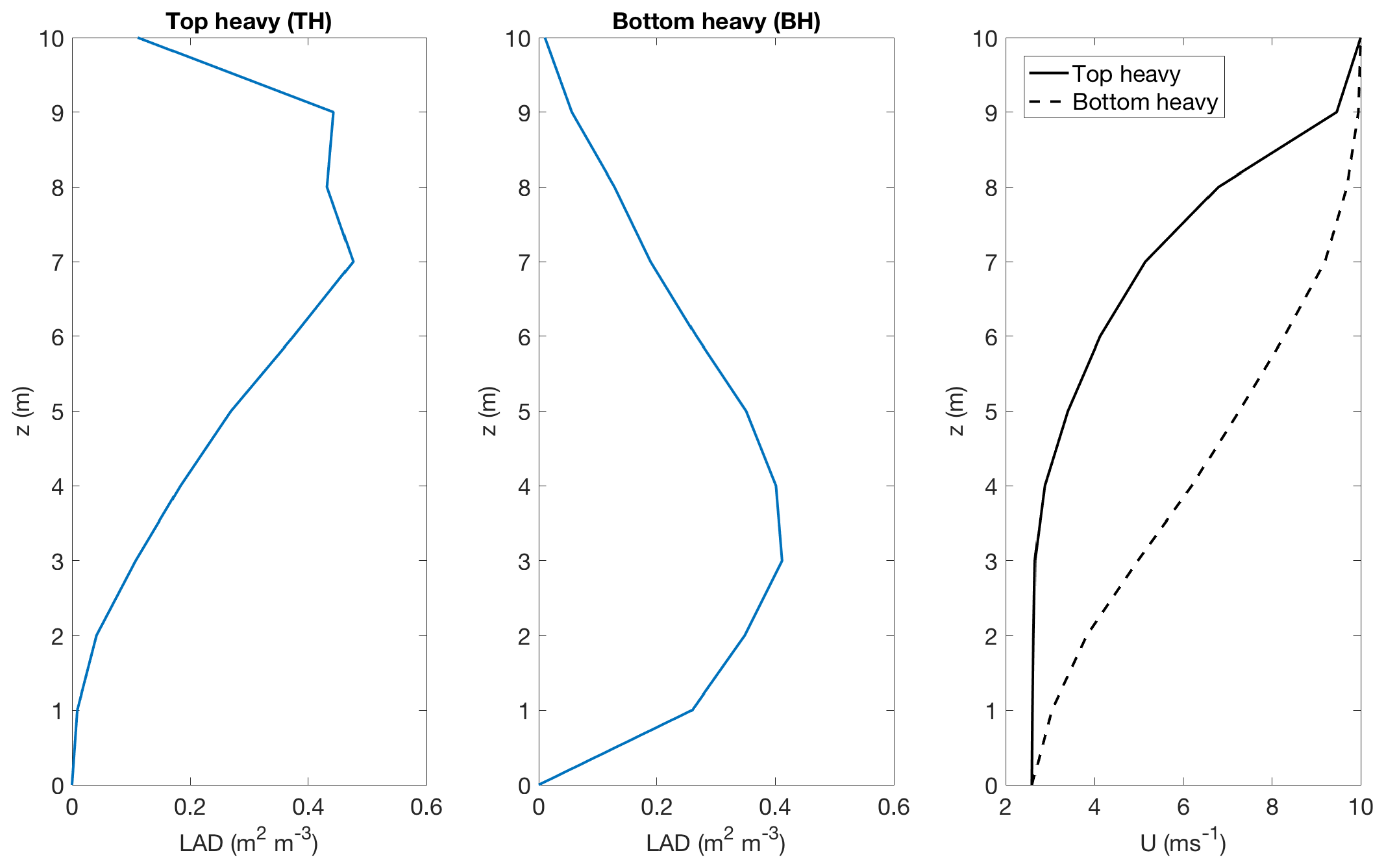
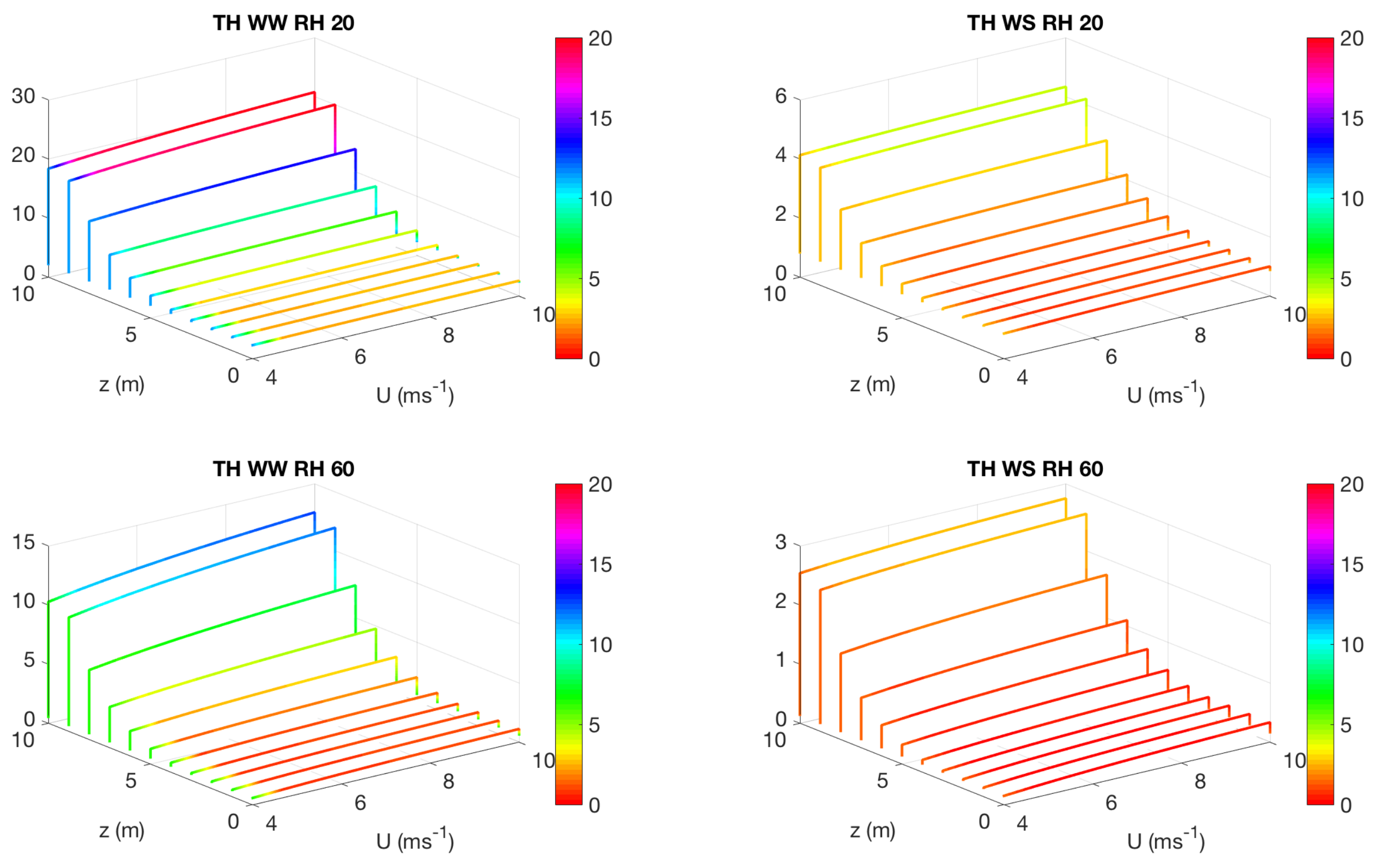
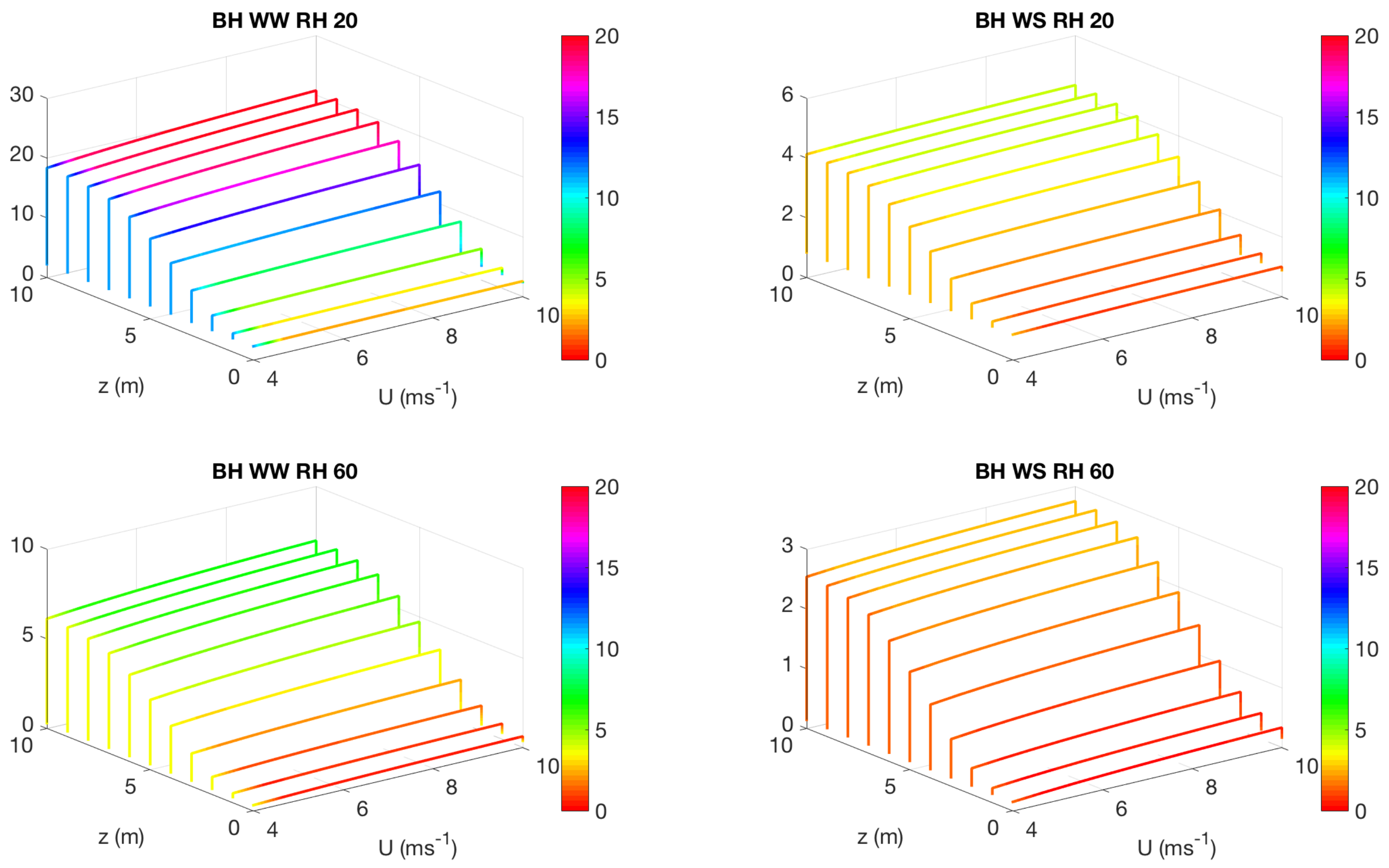
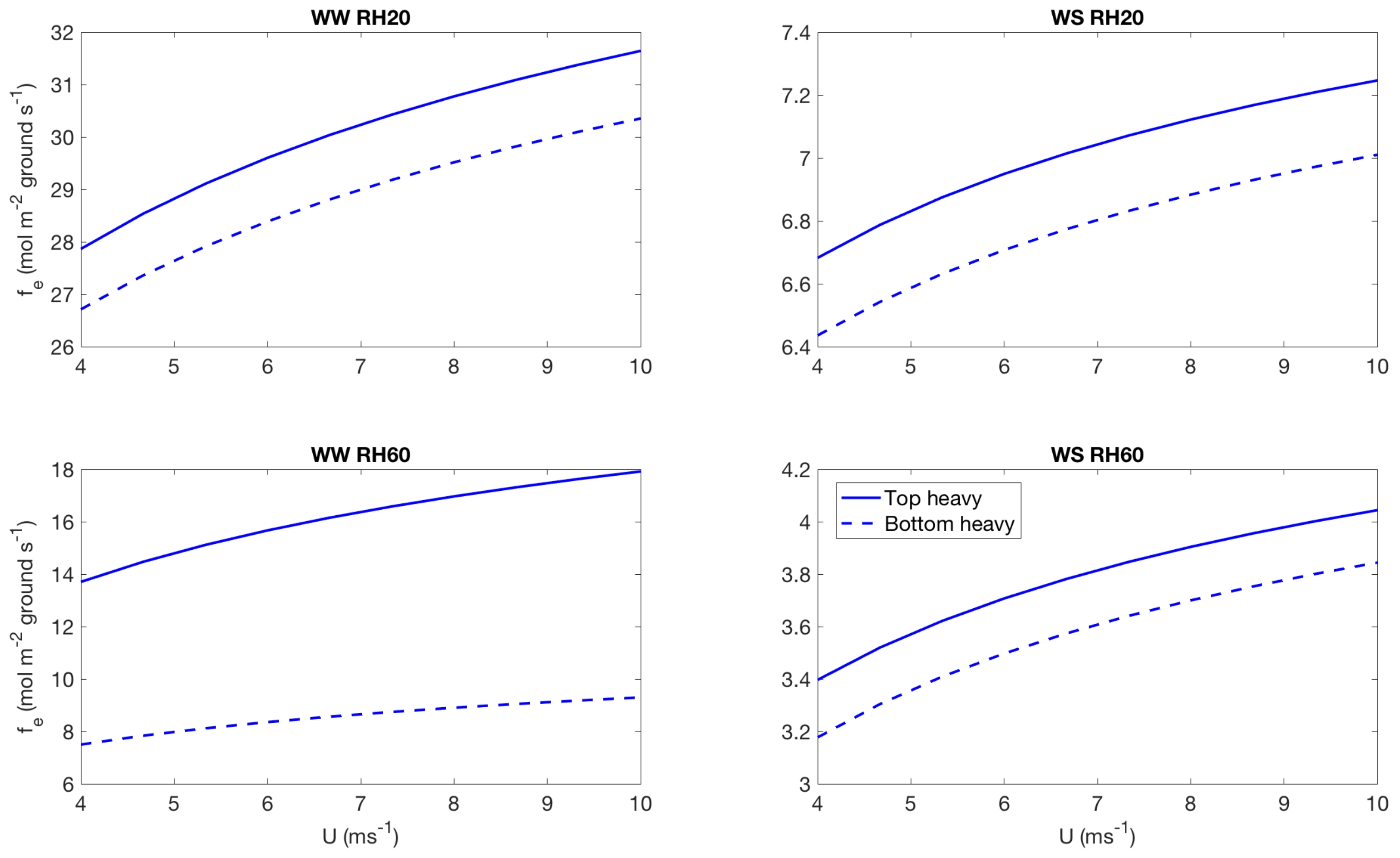
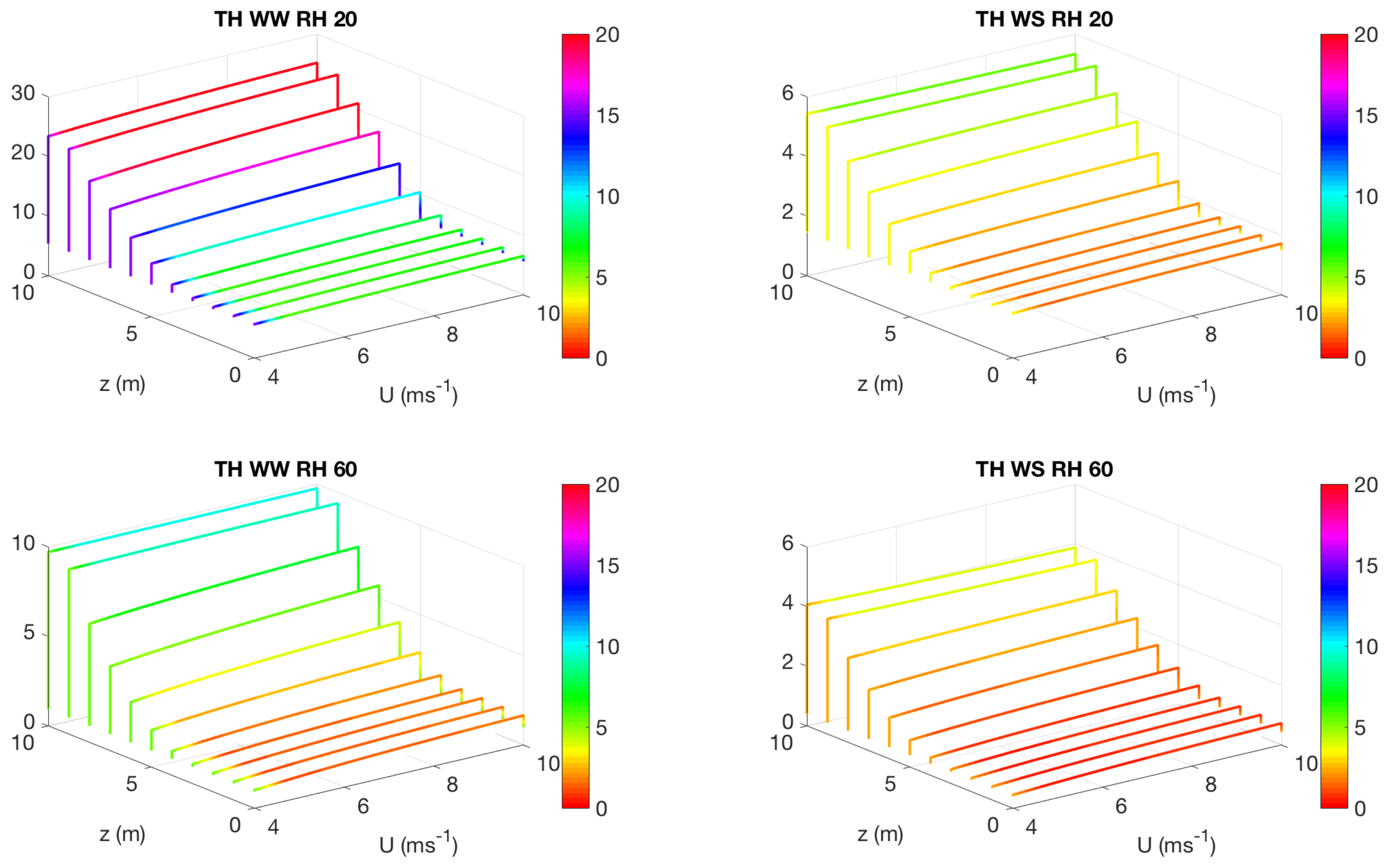
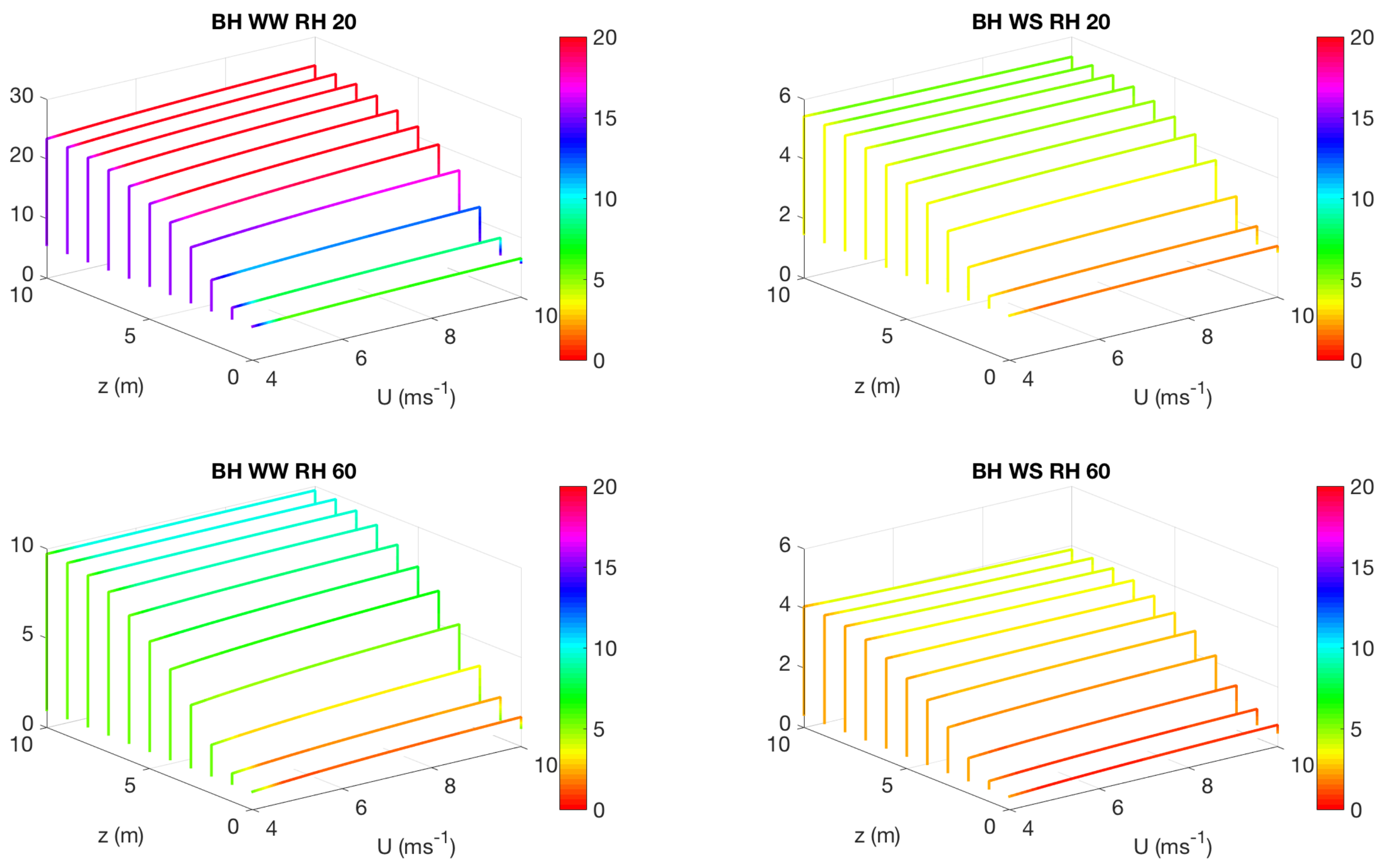
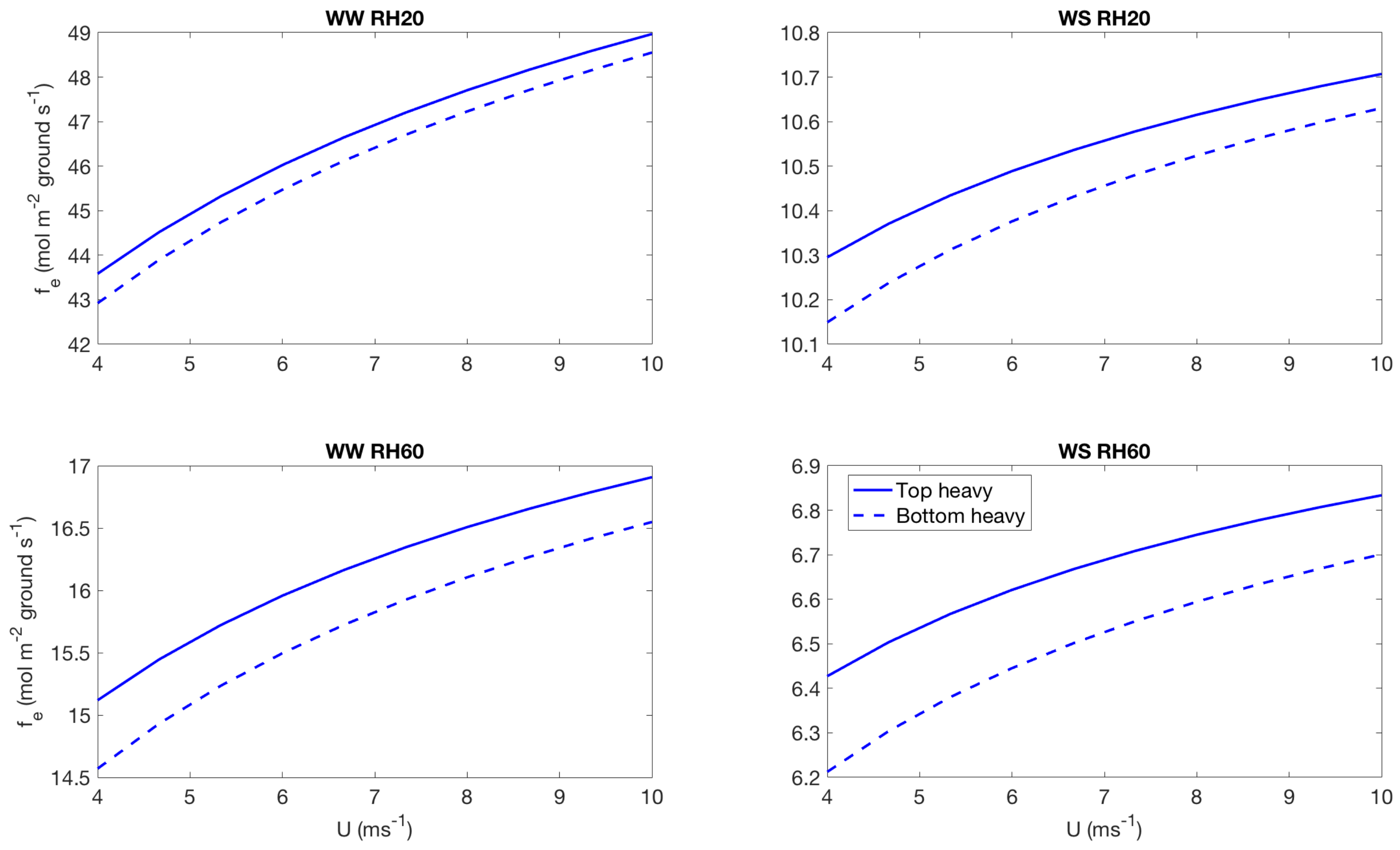
3.2. Effect of Canopy Architecture on Transpiration
- well watered high evaporative demand condition (WW RH20): Transpiration is highest for this condition for both top heavy and bottom heavy scenarios under all light conditions. For the top heavy conditions, the top levels of the canopy undergo higher levels of transpiration and lower levels have significantly less transpiration. For the bottom heavy scenario, the bottom levels undergo considerable transpiration. However canopy level transpirations are higher for the top heavy scenario. Transpiration increases with wind speed for almost all cases both for leaf level and canopy level results, although this is more prominent in the top layers of the canopy. Overall, integrated canopy level transpiration is much higher for the higher light conditions.
- well watered low evaporative demand condition (WW RH60): This scenario always has lower transpiration than the high evaporative demand conditions because of lower demand but has higher transpiration rates compared to the other water stressed conditions because of higher supply of water/moisture. For the top heavy conditions, the top levels of the canopy undergo higher levels of transpiration and lower levels have significantly less transpiration. For the bottom heavy scenario, the bottom levels undergo considerable transpiration. However canopy level transpirations are higher for the top heavy scenario. Transpiration increases with wind speed for almost all cases both for leaf level and canopy level results, however the rate of increase of leaf level transpiration at the top layers are higher than the well watered high evaporative demand condition. Another interesting observation is that for this scenario, the increase of transpiration with wind speed for top heavy and bottom heavy scenarios follows different rates, and the difference is more prominent for the low light conditions. Overall, integrated canopy level transpiration is much higher for the higher light conditions.
- water stressed high evaporative demand condition (WS RH20): This scenario has lower transpiration rates compared to both well watered scenarios because of lower supply of soil moisture. However, the transpiration rate is higher compared to the water stressed, low evaporative demand condition (WS RH60) because of higher atmospheric demand. For the top heavy conditions, the top levels of the canopy undergo higher levels of transpiration and lower levels have significantly less transpiration. For the bottom heavy scenario, the bottom levels undergo considerable transpiration. Canopy level transpirations are higher for the top heavy scenario. Unlike the other scenarios, leaf level transpiration rates remain relatively flat with the increase of wind speed at all levels for the low level condition. For high light conditions, leaf level transpiration actually reduces at the top levels with increase of wind speed. At lower levels, leaf level transpiration still increases with wind speed. Overall canopy level transpiration increases with wind speed but the rate of increase is quite low.
- water stressed low evaporative demand condition (WS RH60): This scenario has lowest transpiration rates among all cases because of lowest supply and lowest demand. For the top heavy conditions, the top levels of the canopy undergo higher levels of transpiration and lower levels have significantly less transpiration. For the bottom heavy scenario, the bottom levels undergo considerable transpiration. Canopy level transpirations are higher for the top heavy scenario. It is interesting to note that for low light conditions, leaf level transpiration at the top layers increase with wind speed but decrease with wind speed at high light conditions. Overall canopy level transpiration still increases with wind speed but at different rates for the top heavy and bottom heavy scenarios.
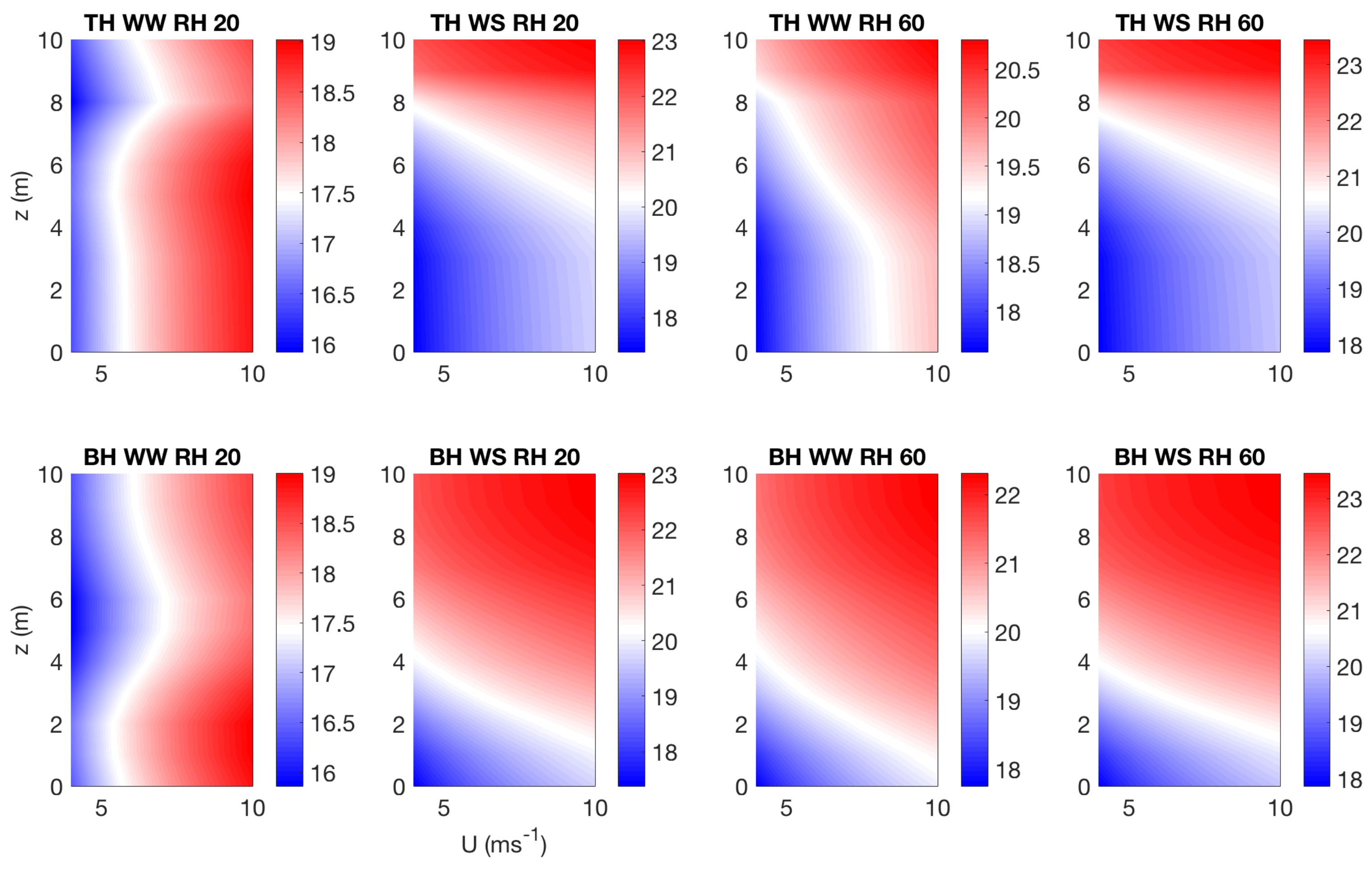
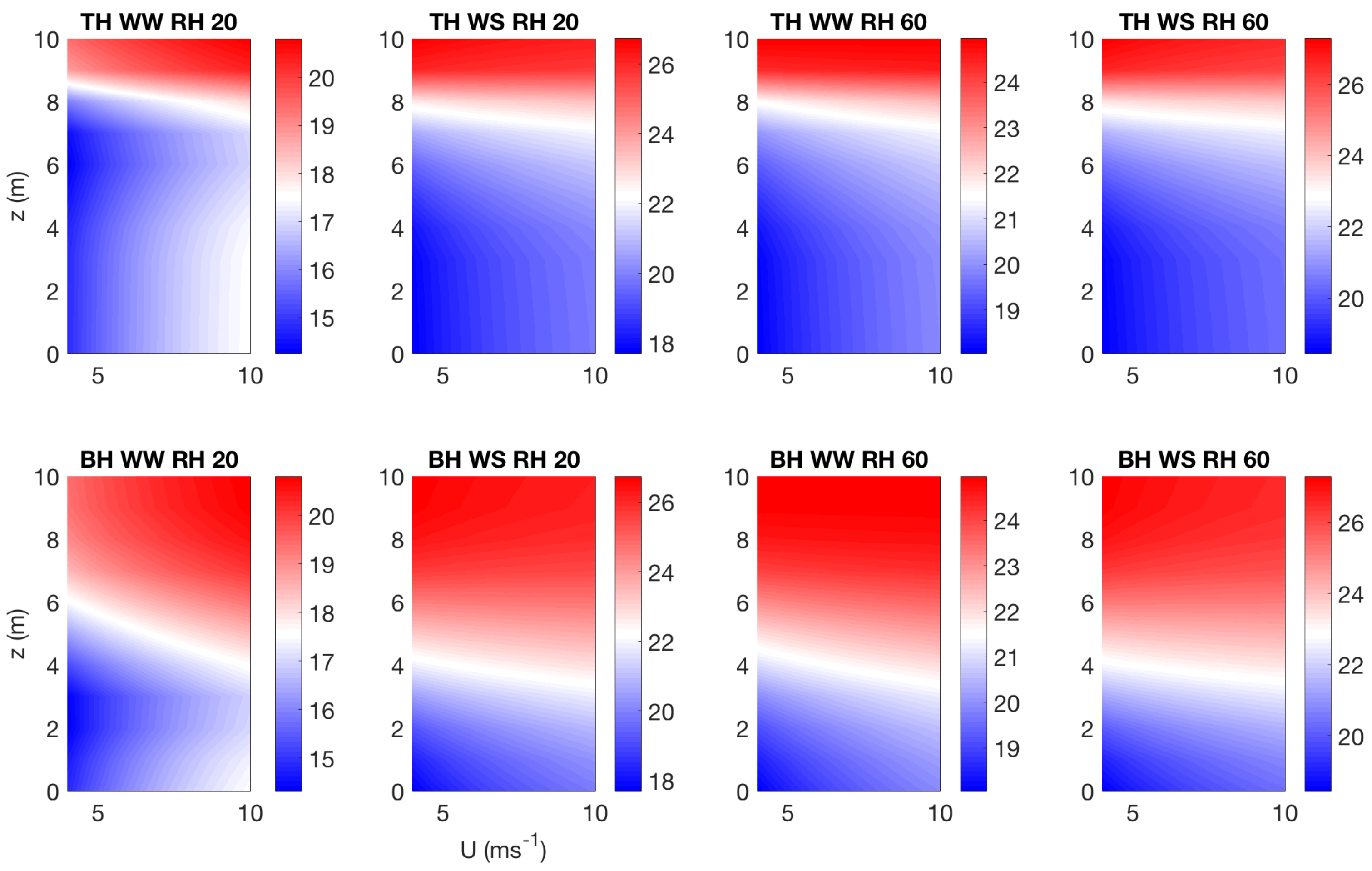
3.3. Effect of Canopy Architecture on Canopy Temperature
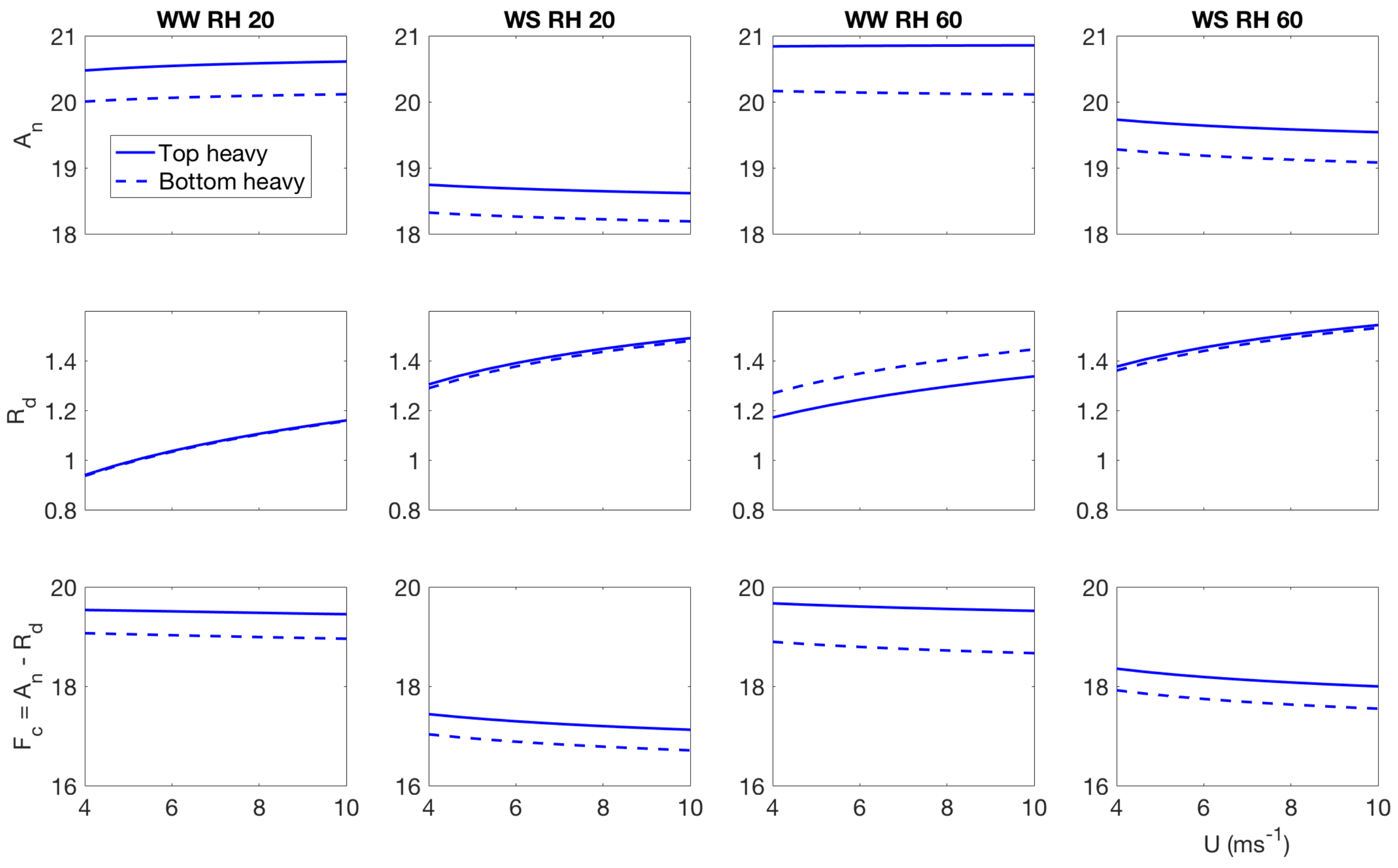
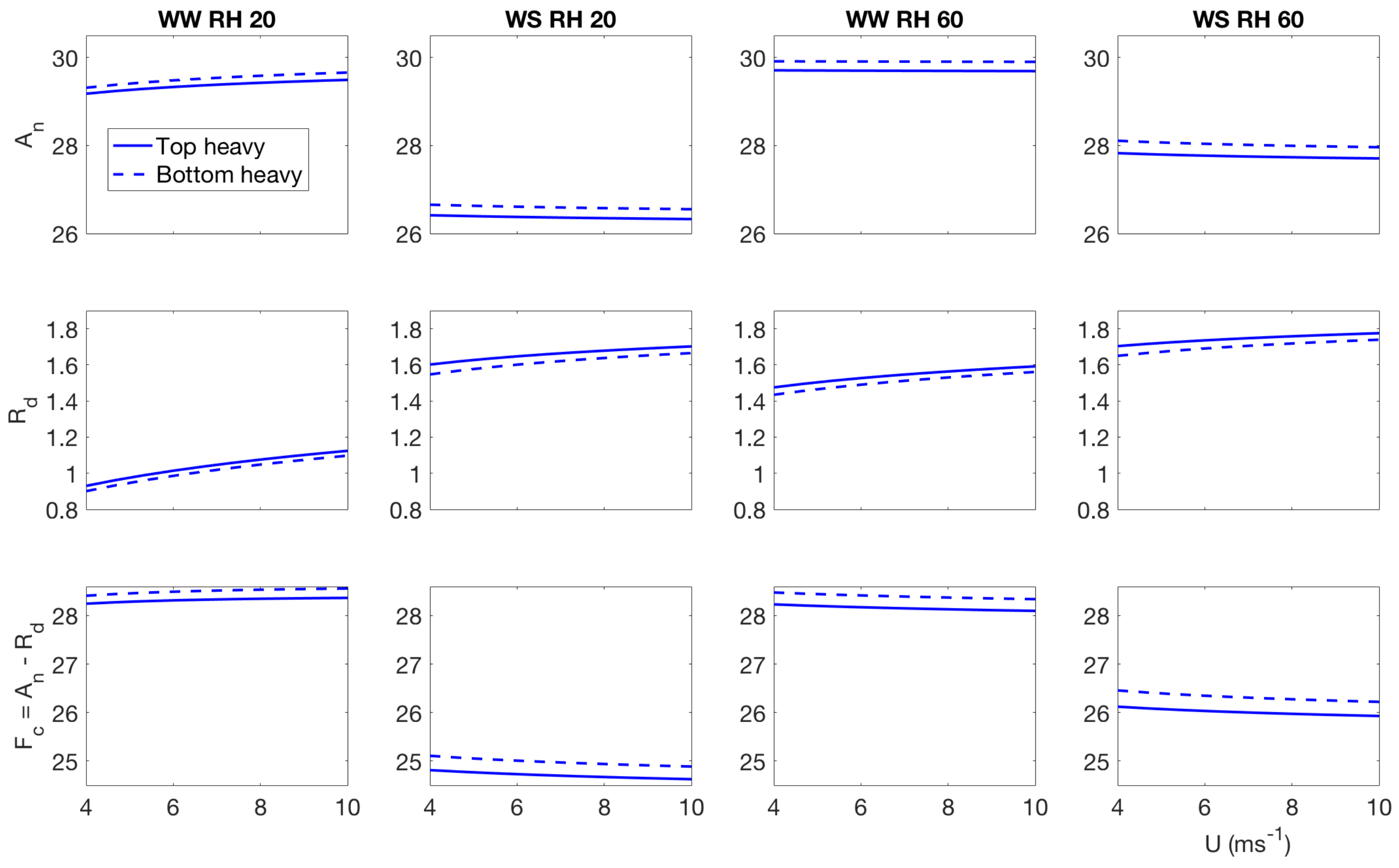
4. Effect of Canopy Architecture on Carbon Exchange
- Well watered, high evaporative demand: carbon assimilation increases slightly with wind speed, however, respiration increases at a faster rate, so effectively net carbon uptake reduces with wind speed at low light condition. The top heavy scenario has higher assimilation and carbon uptake in the low light condition compared to the bottom heavy scenario. In the higher light condition, the bottom heavy scenario has higher carbon assimilation and higher net carbon uptake compared to the top heavy scenario. That said, respiration for the top heavy scenario is higher. More interestingly, the net assimilation and carbon uptake increases with wind speed unlike the low light condition. The difference of behavior can be attributed to different temperature response—which is one of the main driver of the stomatal conductance of carbon dioxide and respiration. Also important to recall the contrasting nature of carbon uptake and water release. The stomates open to maximize carbon gain and minimize water loss. However, when the stomatal apertures are large to satisfy the high transpiration rates, they also gain more carbon. However, these results indicate that under low light conditions, the contrasting effects are still there but in higher light conditions, the stomatal opening is not sufficient enough to impose opposing effects of water loss and carbon gain.
- Water stressed, high evaporative demand: this scenario has the lowest carbon assimilation and uptake, which reduce with wind speed under high and low light conditions. For the low light condition, the top heavy scenario has higher carbon assimilation, respiration and uptake. However, for the higher light condition, the bottom heavy scenario has higher carbon assimilation and uptake with the exception of respiration.
- well watered low evaporative demand: This scenario also has higher rates of carbon assimilation and net uptake and they reduce with increasing wind speed under both high and low light conditions. This scenario also displays the highest differences between the top heavy and bottom heavy scenarios. Like the other cases, the top heavy case has higher carbon assimilation and uptake than the bottom heavy scenario with the exception of respiration. The trend completely reverses under higher light condition.
- water stressed low evaporative demand Under this condition, carbon assimilation and net carbon uptake reduce with wind speed, while the respiration increases in wind speed for both light environments. However, the rate of respiration increase are different under different light conditions. Also interestingly, under low light environment, the top heavy case uptakes more carbon, while under high light conditions, the bottom heavy case uptakes more carbon.
5. Conclusions
6. Data Availability
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| Symbol | Description | Unit |
| Ambient oxygen concentration | ||
| H | Sensible heat flux | |
| J | Electron transport rate | |
| Electron transport rate at light saturation | ||
| Normalized at | ||
| Michaelis constant for fixation | ||
| Michaelis constant for inhibition | ||
| L | Latent heat for water vaporization | |
| Latent heat flux | ||
| Photosynthetically active radiation | ||
| Atmospheric pressure | ||
| Net radiation | ||
| Absorbed radiation | ||
| Emitted longwave radiation | ||
| R | Universal gas constant | |
| Daytime mitochondrial respiration rate | ||
| Relative humidity | % | |
| Air temperature | ||
| Leaf surface temperature | ||
| U | Mean wind speed | |
| Vapor pressure deficit | ||
| Maximum carboxylation capacity under light saturated conditions | ||
| Normalized at | ||
| Ambient concentration | ||
| Intercellular concentration | ||
| Capacity of dry air at constant pressure | ||
| d | Characteristic leaf dimension | |
| Ambient water vapor concentration | ||
| Intercellular water vapor concentration | ||
| Carbon assimilation rate | ||
| Transpiration rate | ||
| Laminar boundary layer conductance for | ||
| Laminar boundary layer conductance for water vapor | ||
| Nocturnal residual conductance of water vapor | ||
| Stomatal conductance for | ||
| Stomatal conductance for water vapor | ||
| Total conductance for | ||
| Total conductance for water vapor | ||
| compensation point | ||
| Leaf surface emissivity | Dimensionless | |
| Stefan-Boltzmann constant | ||
| Marginal water use efficiency |
References
- Chambers, J.; Davies, S.; Koven, C.; Kueppers, L.; Leung, R.; McDowell, N.; Norby, R.; Rogers, A. Next Generation Ecosystem Experiment (NGEE) Tropics; US DOE NGEE Tropics White Paper; US Department of Energy, Office of Biological and Environmental Research: Washington DC, USA, 2014. [Google Scholar]
- Alves, I.; Perrier, A.; Pereira, L. Aerodynamic and surface resistances of complete cover crops: How good is the “big leaf”? Trans. ASAE 1998, 41, 345–351. [Google Scholar] [CrossRef]
- Banerjee, T.; De Roo, F.; Mauder, M. Explaining the convector effect in canopy turbulence by means of large-eddy simulation. Hydrol. Earth Syst. Sci. 2017, 21, 2987–3000. [Google Scholar] [CrossRef]
- Yu, M.H.; Ding, G.D.; Gao, G.L.; Sun, B.P.; Zhao, Y.Y.; Wan, L.; Wang, D.Y.; Gui, Z.Y. How the plant temperature links to the air temperature in the desert plant Artemisia Ordosica. PLoS ONE 2015, 10, e0135452. [Google Scholar] [CrossRef] [PubMed]
- Michaletz, S.T.; Weiser, M.D.; McDowell, N.G.; Zhou, J.; Kaspari, M.; Helliker, B.R.; Enquist, B.J. The energetic and carbon economic origins of leaf thermoregulation. Nat. Plants 2016, 2, 16129. [Google Scholar] [CrossRef] [PubMed]
- Campbell, G.S.; Norman, J.M. An Introduction to Environmental Biophysics; Springer Science & Business Media: New York, NY, USA, 2012. [Google Scholar]
- Cassiani, M.; Katul, G.; Albertson, J. The effects of canopy leaf area index on airflow across forest edges: Large-eddy simulation and analytical results. Bound.-Layer Meteorol. 2008, 126, 433–460. [Google Scholar] [CrossRef]
- Banerjee, T.; Katul, G.; Fontan, S.; Poggi, D.; Kumar, M. Mean flow near edges and within cavities situated inside dense canopies. Bound.-Layer Meteorol. 2013, 149, 19–41. [Google Scholar] [CrossRef]
- Katul, G.G.; Mahrt, L.; Poggi, D.; Sanz, C. One-and two-equation models for canopy turbulence. Bound.-Layer Meteorol. 2004, 113, 81–109. [Google Scholar] [CrossRef]
- Bailey, B.; Overby, M.; Willemsen, P.; Pardyjak, E.; Mahaffee, W.; Stoll, R. A scalable plant-resolving radiative transfer model based on optimized GPU ray tracing. Agric. For. Meteorol. 2014, 198, 192–208. [Google Scholar] [CrossRef]
- Woodward, F. Global change: Translating plant ecophysiological responses to ecosystems. Trends Ecol. Evol. 1990, 5, 308–311. [Google Scholar] [CrossRef]
- McCarthy, H.R.; Oren, R.; Finzi, A.C.; Ellsworth, D.S.; Kim, H.S.; Johnsen, K.H.; Millar, B. Temporal dynamics and spatial variability in the enhancement of canopy leaf area under elevated atmospheric CO2. Glob. Chang. Biol. 2007, 13, 2479–2497. [Google Scholar] [CrossRef]
- Katul, G.G.; Oren, R.; Manzoni, S.; Higgins, C.; Parlange, M.B. Evapotranspiration: A process driving mass transport and energy exchange in the soil-plant-atmosphere-climate system. Rev. Geophys. 2012, 50. [Google Scholar] [CrossRef]
- Repo, T.; Hänninen, H.; Kellomäki, S. The effects of long-term elevation of air temperature and CO2 on the frost hardiness of Scots pine. Plant Cell Environ. 1996, 19, 209–216. [Google Scholar] [CrossRef]
- Lutze, J.; Roden, J.; Holly, C.; Wolfe, J.; Egerton, J.; Ball, M. Elevated atmospheric CO2 promotes frost damage in evergreen tree seedlings. Plant Cell Environ. 1998, 21, 631–635. [Google Scholar] [CrossRef]
- Long, S.P.; Ainsworth, E.A.; Leakey, A.D.; Nösberger, J.; Ort, D.R. Food for thought: Lower-than-expected crop yield stimulation with rising CO2 concentrations. Science 2006, 312, 1918–1921. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.; Hanson, P.J.; Mac Post, W.; Kaiser, D.P.; Yang, B.; Nemani, R.; Pallardy, S.G.; Meyers, T. The 2007 eastern US spring freeze: Increased cold damage in a warming world. BioScience 2008, 58, 253–262. [Google Scholar] [CrossRef]
- Huang, C.W.; Chu, C.R.; Hsieh, C.I.; Palmroth, S.; Katul, G.G. Wind-induced leaf transpiration. Adv. Water Resour. 2015, 86, 240–255. [Google Scholar] [CrossRef]
- Launiainen, S.; Katul, G.G.; Kolari, P.; Vesala, T.; Hari, P. Empirical and optimal stomatal controls on leaf and ecosystem level CO2 and H2O exchange rates. Agric. For. Meteorol. 2011, 151, 1672–1689. [Google Scholar] [CrossRef]
- Siqueira, M.; Katul, G.; Sampson, D.; Stoy, P.; Juang, J.Y.; McCarthy, H.; Oren, R. Multiscale model intercomparisons of CO2 and H2O exchange rates in a maturing southeastern US pine forest. Glob. Chang. Biol. 2006, 12, 1189–1207. [Google Scholar] [CrossRef]
- Lai, C.T.; Katul, G.; Oren, R.; Ellsworth, D.; Sch, K. Modeling CO2 and water vapor turbulent flux distributions within a forest canopy. J. Geophys. Res 2000, 105, 26333–26351. [Google Scholar] [CrossRef]
- Farquhar, G.V.; Caemmerer, S.V.; Berry, J. A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta 1980, 149, 78–90. [Google Scholar] [CrossRef] [PubMed]
- Vico, G.; Manzoni, S.; Palmroth, S.; Weih, M.; Katul, G. A perspective on optimal leaf stomatal conductance under CO2 and light co-limitations. Agric. For. Meteorol. 2013, 182, 191–199. [Google Scholar] [CrossRef]
- Katul, G.; Manzoni, S.; Palmroth, S.; Oren, R. A stomatal optimization theory to describe the effects of atmospheric CO2 on leaf photosynthesis and transpiration. Ann. Bot. 2010, 105, 431–442. [Google Scholar] [CrossRef] [PubMed]
- Yi, C. Momentum transfer within canopies. J. Appl. Meteorol. Climatol. 2008, 47, 262–275. [Google Scholar] [CrossRef]
- Kim, H.S.; Palmroth, S.; Thérézien, M.; Stenberg, P.; Oren, R. Analysis of the sensitivity of absorbed light and incident light profile to various canopy architecture and stand conditions. Tree Physiol. 2011, 31, 30–47. [Google Scholar] [CrossRef] [PubMed]
- Russell, E.S.; Liu, H.; Thistle, H.; Strom, B.; Greer, M.; Lamb, B. Effects of thinning a forest stand on sub-canopy turbulence. Agric. For. Meteorol. 2018, 248, 295–305. [Google Scholar] [CrossRef]













© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Banerjee, T.; Linn, R. Effect of Vertical Canopy Architecture on Transpiration, Thermoregulation and Carbon Assimilation. Forests 2018, 9, 198. https://doi.org/10.3390/f9040198
Banerjee T, Linn R. Effect of Vertical Canopy Architecture on Transpiration, Thermoregulation and Carbon Assimilation. Forests. 2018; 9(4):198. https://doi.org/10.3390/f9040198
Chicago/Turabian StyleBanerjee, Tirtha, and Rodman Linn. 2018. "Effect of Vertical Canopy Architecture on Transpiration, Thermoregulation and Carbon Assimilation" Forests 9, no. 4: 198. https://doi.org/10.3390/f9040198
APA StyleBanerjee, T., & Linn, R. (2018). Effect of Vertical Canopy Architecture on Transpiration, Thermoregulation and Carbon Assimilation. Forests, 9(4), 198. https://doi.org/10.3390/f9040198