
Molecular and Phylogenetic Characterization of Novel Papillomaviruses Isolated from Oral and Anogenital Neoplasms of Japanese Macaques (Macaca fuscata)

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Tumor Specimens
2.2. DNA Extraction and Amplification of Partial Viral Sequences
2.3. Amplification of the Complete MfuPV1 and MfuPV2 Genomes
2.4. Cloning of the Complete MfuPV1 and MfuPV2 Genomes
2.5. Sequencing of the Complete MfuPV1 and MfuPV2 Genomes
2.6. Molecular Analysis of MfuPV1 and MfuPV2
2.7. Phylogenetic Analysis of MfuPV1 and MfuPV2 and Sequence Similarities
2.8. MfuPV1 and MfuPV2 Type-Specific Quantitative Real-Time PCR Assays
2.9. MfuPV1 and MfuPV2 Viral Load Calculation
2.10. Data Availability
3. Results
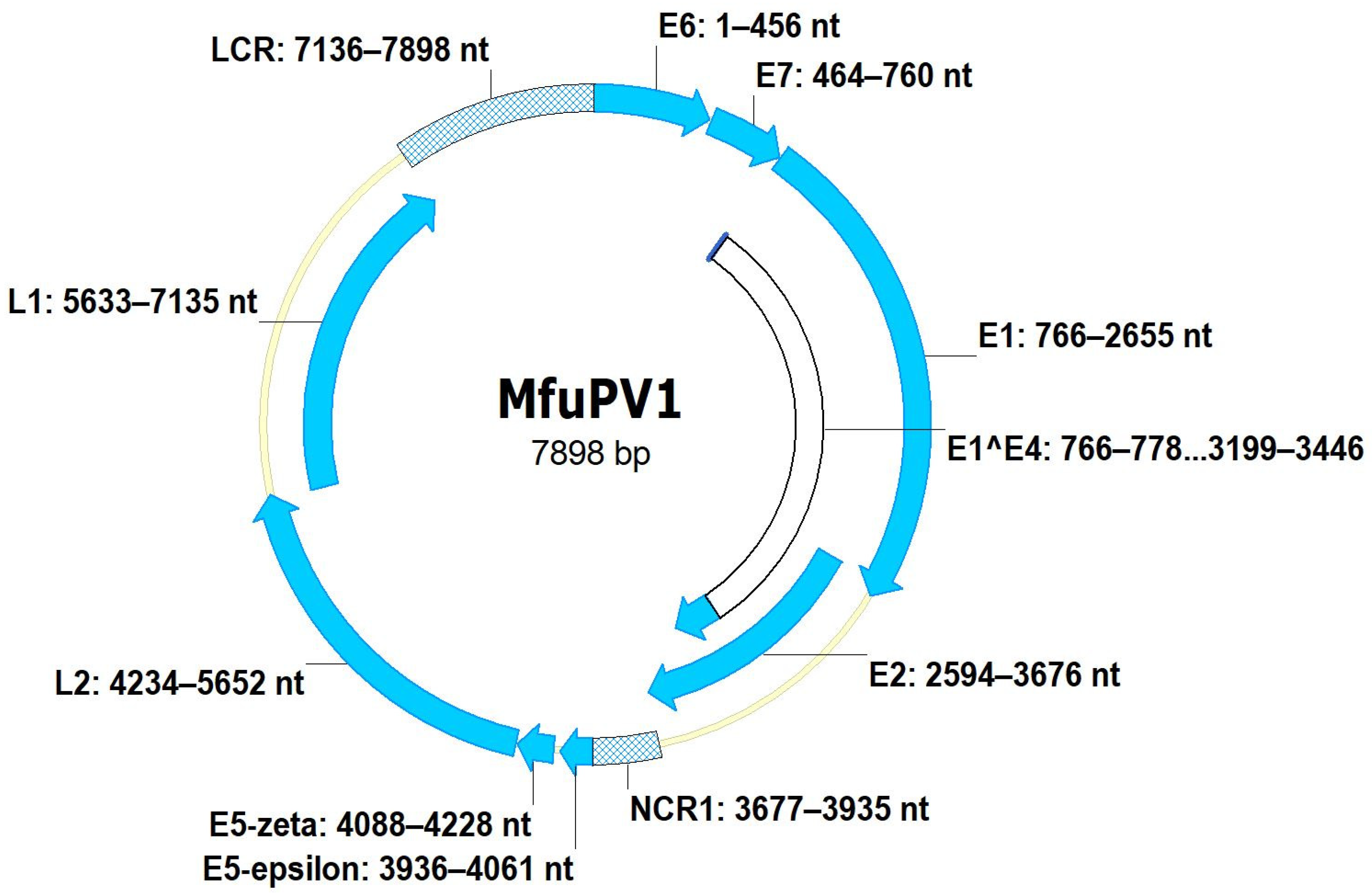
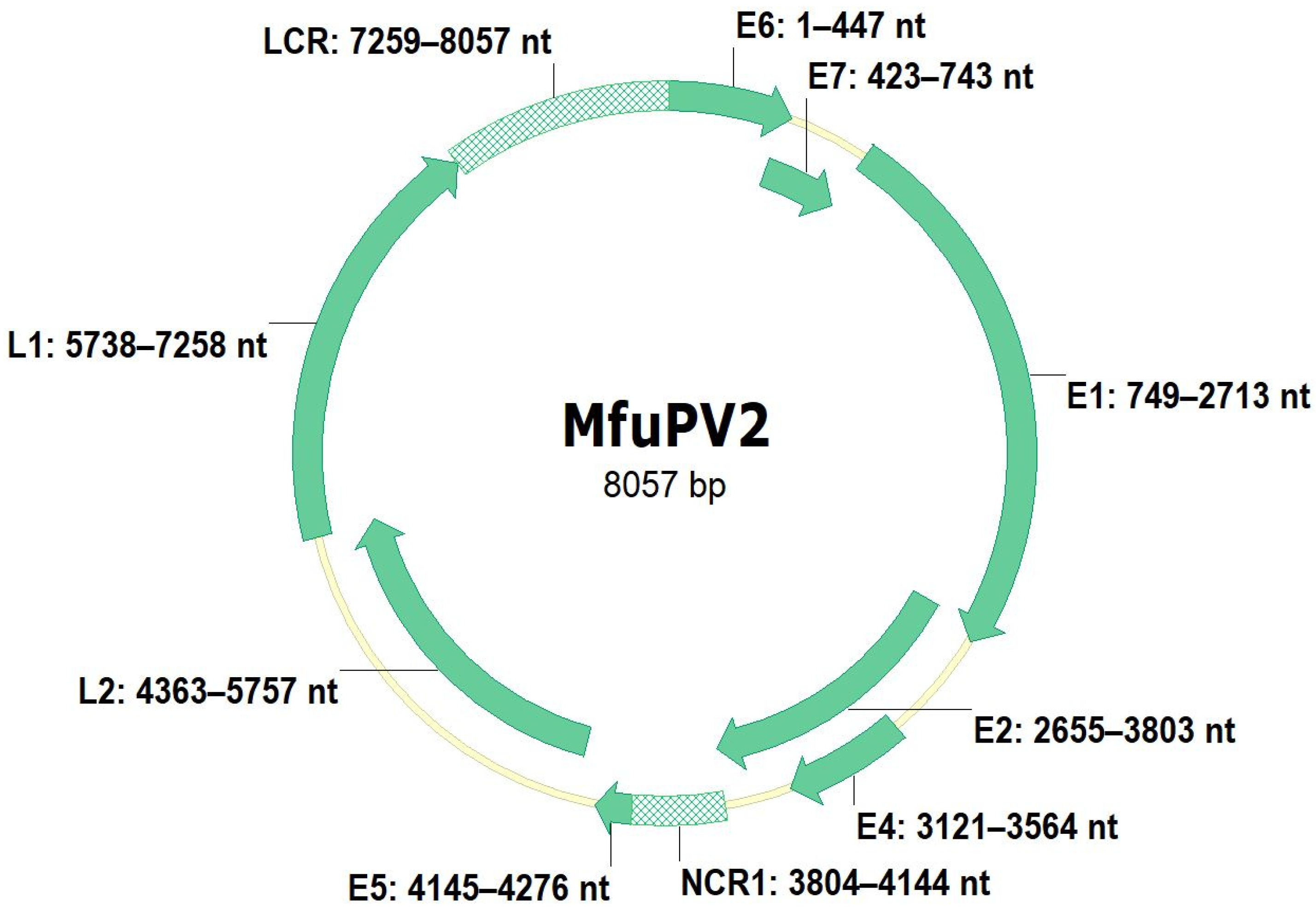
3.1. Genomic Organization of MfuPV1 and MfuPV2
3.2. Characteristics of MfuPV1 and MfuPV2 Proteins
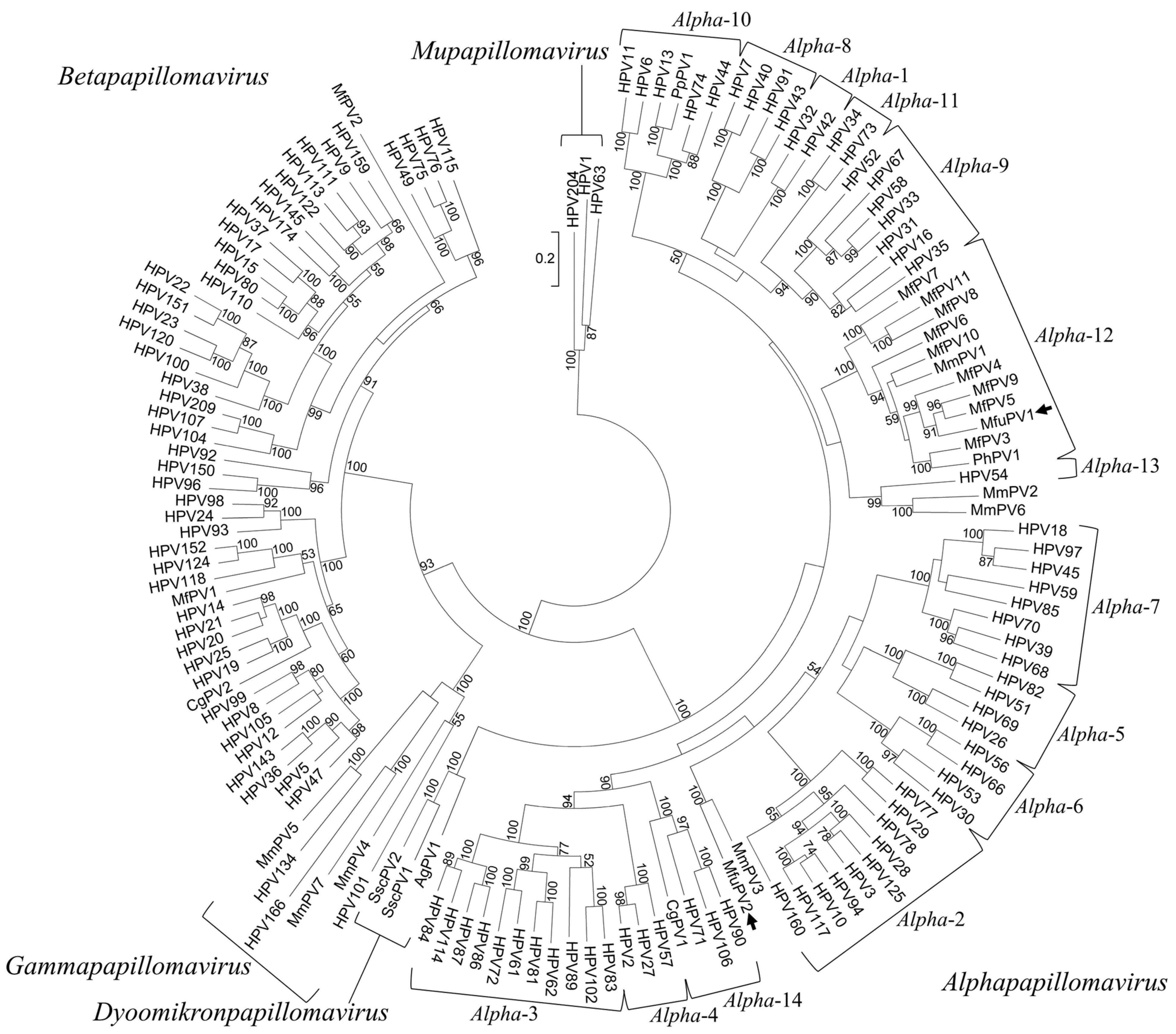
3.3. Phylogenetic Analysis of MfuPV1 and MfuPV2 and Sequence Similarities
3.4. MfuPV1 and MfuPV2 Viral Load
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Villiers, E.M.; Fauquet, C.; Broker, T.R.; Bernard, H.U.; zur Hausen, H. Classification of papillomaviruses. Virology 2004, 324, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Bernard, H.U.; Burk, R.D.; Chen, Z.; van Doorslaer, K.; zur Hausen, H.; de Villiers, E.M. Classification of papillomaviruses (PVs) based on 189 PV types and proposal of taxonomic amendments. Virology 2010, 401, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Doorbar, J.; Egawa, N.; Griffin, H.; Kranjec, C.; Murakami, I. Human papillomavirus molecular biology and disease association. Rev. Med. Virol. 2015, 25, 2–23. [Google Scholar] [CrossRef]
- Van Doorslaer, K.; Dillner, J. The launch of an international animal papillomavirus reference center. Viruses 2019, 11, 55. [Google Scholar] [CrossRef]
- MacLachlan, N.J.; Dubovi, E.J. (Eds.) Chapter 11—Papillomaviridae and Polyomaviridae. In Fenner’s Veterinary Virology, 5th ed.; Academic Press: Cambridge, MA, USA, 2016; pp. 229–243. [Google Scholar]
- Van Doorslaer, K.; Li, Z.; Xirasagar, S.; Maes, P.; Kaminsky, D.; Liou, D.; Sun, Q.; Kaur, R.; Huyen, Y.; McBride, A.A. The Papillomavirus Episteme: A major update to the papillomavirus sequence database. Nucleic Acids Res. 2017, 45, D499–D506. [Google Scholar] [CrossRef]
- Chan, S.Y.; Ostrow, R.S.; Faras, A.J.; Bernard, H.U. Genital papillomaviruses (PVs) and epidermodysplasia verruciformis PVs occur in the same monkey species: Implications for PV evolution. Virology 1997, 228, 213–217. [Google Scholar] [CrossRef]
- Joh, J.; Hopper, K.; van Doorslaer, K.; Sundberg, J.P.; Jenson, A.B.; Ghim, S.J. Macaca fascicularis papillomavirus type 1: A non-human primate betapapillomavirus causing rapidly progressive hand and foot papillomatosis. J. Gen. Virol. 2009, 90, 987–994. [Google Scholar] [CrossRef]
- Li, J.; Han, K.; Xing, J.; Kim, H.S.; Rogers, J.; Ryder, O.A.; Disotell, T.; Yue, B.; Batzer, M.A. Phylogeny of the macaques (Cercopithecidae: Macaca) based on Alu elements. Gene 2009, 448, 242–249. [Google Scholar] [CrossRef]
- Wood, C.E.; Tannehill-Gregg, S.H.; Chen, Z.; van Doorslaer, K.; Nelson, D.R.; Cline, J.M.; Burk, R.D. Novel Betapapillomavirus associated with hand and foot papillomas in a cynomolgus macaque. Vet. Pathol. 2011, 48, 731–736. [Google Scholar] [CrossRef]
- Chen, Z.; van Doorslaer, K.; DeSalle, R.; Wood, C.E.; Kaplan, J.R.; Wagner, J.D.; Burk, R.D. Genomic diversity and interspecies host infection of Macaca fascicularis papillomaviruses (MfPVs) within the alpha papillomavirus α12 species. Virology 2009, 393, 304–310. [Google Scholar] [CrossRef]
- Long, T.; Wong, P.Y.; Ho, W.C.S.; Burk, R.D.; Chan, P.K.S.; Chen, Z. Complete Genome Sequences of Six Novel Macaca mulatta Papillomavirus Types Isolated from Genital Sites of Rhesus Monkeys in Hong Kong SAR, China. Microbiol. Resour. Announc. 2018, 7, e01414-18. [Google Scholar] [CrossRef]
- Kloster, B.E.; Manias, D.A.; Ostrow, R.S.; Shaver, M.K.; McPherson, S.W.; Rangen, S.R.; Uno, H.; Faras, A.J. Molecular cloning and characterization of the DNA of two papillomaviruses from monkeys. Virology 1988, 166, 30–40. [Google Scholar] [CrossRef]
- National Research Council (US) Committee for the Update of the Guide for the Care and Use of Laboratory Animals. Guide for the Care and Use of Laboratory Animals, 8th ed.; National Academies Press (US): Washington, DC, USA, 2011. [Google Scholar]
- Chouhy, D.; Kocjan, B.J.; Staheli, J.P.; Bolatti, E.M.; Hošnjak, L.; Sagadin, M.; Giri, A.A.; Rose, T.M.; Poljak, M. Detection of novel Betapapillomaviruses and Gammapapillomaviruses in eyebrow hair follicles using a single-tube ‘hanging droplet’ PCR assay with modified pan-PV CODEHOP primers. J. Gen. Virol. 2018, 99, 109–118. [Google Scholar] [CrossRef]
- Rector, A.; Tachezy, R.; Van Ranst, M. A sequence-independent strategy for detection and cloning of circular DNA virus genomes by using multiply primed rolling-circle amplification. J. Virol. 2004, 78, 4993–4998. [Google Scholar] [CrossRef] [PubMed]
- Platt, A.R.; Woodhall, R.W.; George, A.L., Jr. Improved DNA sequencing quality and efficiency using an optimized fast cycle sequencing protocol. Biotechniques 2007, 43, 58–62. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.Y.; Chang, W.C.; Hsu, J.B.; Chang, T.H.; Shien, D.M. GPMiner: An integrated system for mining combinatorial cis-regulatory elements in mammalian gene group. BMC Genom. 2012, 13, S3. [Google Scholar] [CrossRef]
- Prestridge, D.S. SIGNAL SCAN: A computer program that scans DNA sequences for eukaryotic transcriptional elements. Comput. Appl. Biosci. 1991, 7, 203–206. [Google Scholar] [CrossRef]
- Liu, H.; Han, H.; Li, J.; Wong, L. DNAFSMiner: A web-based software toolbox to recognize two types of functional sites in DNA sequences. Bioinformatics 2005, 21, 671–673. [Google Scholar] [CrossRef]
- Bornberg-Bauer, E.; Rivals, E.; Vingron, M. Computational approaches to identify leucine zippers. Nucleic Acids Res. 1998, 26, 2740–2746. [Google Scholar] [CrossRef]
- Hošnjak, L.; Kocjan, B.J.; Pirš, B.; Seme, K.; Poljak, M. Characterization of two novel Gammapapillomaviruses, HPV179 and HPV184, isolated from common warts of a renal-transplant recipient. PLoS ONE 2015, 10, e0119154. [Google Scholar] [CrossRef]
- Kocjan, B.J.; Hošnjak, L.; Račnik, J.; Zadravec, M.; Bakovnik, N.; Cigler, B.; Ummelen, M.; Hopman, A.H.N.; Gale, N.; Švara, T.; et al. Molecular characterization, prevalence and clinical relevance of Phodopus sungorus papillomavirus type 1 (PsuPV1) naturally infecting Siberian hamsters (Phodopus sungorus). J. Gen. Virol. 2017, 98, 2799–2809. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Bao, Y.; Chetvernin, V.; Tatusova, T. Improvements to pairwise sequence comparison (PASC): A genome-based web tool for virus classification. Arch. Virol. 2014, 159, 3293–3304. [Google Scholar] [CrossRef] [PubMed]
- Kocjan, B.J.; Poljak, M.; Seme, K. Universal ProbeLibrary based real-time PCR assay for detection and confirmation of human papillomavirus genotype 52 infections. J. Virol. Methods 2010, 163, 492–494. [Google Scholar] [CrossRef] [PubMed]
- Gregory, T.R. Animal Genome Size Database—Detailed Record for Macaca Fuscata. 2020. Available online: http://www.genomesize.com/result_species.php?id=4403 (accessed on 22 September 2020).
- Longworth, M.S.; Laimins, L.A. Pathogenesis of human papillomaviruses in differentiating epithelia. Microbiol. Mol. Biol. Rev. 2004, 68, 362–372. [Google Scholar] [CrossRef] [PubMed]
- Thomas, M.; Dasgupta, J.; Zhang, Y.; Chen, X.; Banks, L. Analysis of specificity determinants in the interactions of different HPV E6 proteins with their PDZ domain containing substrates. Virology 2008, 376, 371–378. [Google Scholar] [CrossRef]
- Liu, X.; Clements, A.; Zhao, K.; Marmorstein, R. Structure of the human Papillomavirus E7 oncoprotein and its mechanism for inactivation of the retinoblastoma tumor suppressor. J. Biol. Chem. 2006, 281, 578–586. [Google Scholar] [CrossRef]
- Fradet-Turcotte, A.; Moody, C.; Laimins, L.A.; Archambault, J. Nuclear export of human papillomavirus type 31 E1 is regulated by Cdk2 phosphorylation and required for viral genome maintenance. J. Virol. 2010, 84, 11747–11760. [Google Scholar] [CrossRef]
- Bergvall, M.; Melendy, T.; Archambault, J. The E1 proteins. Virology 2013, 445, 35–56. [Google Scholar] [CrossRef] [PubMed]
- McBride, A.A. The papillomavirus E2 proteins. Virology 2013, 445, 57–79. [Google Scholar] [CrossRef]
- Landschulz, W.H.; Johnson, P.F.; McKnight, S.L. The leucine zipper: A hypothetical structure common to a new class of DNA binding proteins. Science 1988, 4860, 1759–1764. [Google Scholar] [CrossRef] [PubMed]
- Doorbar, J. The E4 protein; structure, function and patterns of expression. Virology 2013, 445, 80–98. [Google Scholar] [CrossRef] [PubMed]
- Mamoor, S.; Onder, Z.; Karanam, B.; Kwak, K.; Bordeaux, J.; Crosby, L.; Roden, R.B.S.; Moroianu, J. The high risk HPV16 L2 minor capsid protein has multiple transport signals that mediate its nucleocytoplasmic traffic. Virology 2012, 422, 413–424. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.W.; Roden, R.B. L2, the minor capsid protein of papillomavirus. Virology 2013, 445, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Bronnimann, M.P.; Chapman, J.A.; Park, C.K.; Campos, S.K. A transmembrane domain and GxxxG motifs within L2 are essential for papillomavirus infection. J. Virol. 2013, 87, 464–473. [Google Scholar] [CrossRef]
- Zhou, J.; Doorbar, J.; Sun, X.Y.; Crawford, L.V.; McLean, C.S.; Frazer, I.H. Identification of the nuclear localization signal of human papillomavirus type 16 L1 protein. Virology 1991, 185, 625–632. [Google Scholar] [CrossRef]
- Nelson, L.M.; Rose, R.C.; LeRoux, L.; Lane, C.; Bruya, K.; Moroianu, J. Nuclear import and DNA binding of human papillomavirus type 45 L1 capsid protein. J. Cell. Biochem. 2000, 79, 225–238. [Google Scholar] [CrossRef]
- Bernard, H.U. Regulatory elements in the viral genome. Virology 2013, 445, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Belluti, S.; Rigillo, G.; Imbriano, C. Transcription Factors in Cancer: When Alternative Splicing Determines Opposite Cell Fates. Cells 2020, 9, 760. [Google Scholar] [CrossRef] [PubMed]
- Gloss, B.; Bernard, H.U. The E6/E7 promoter of human papillomavirus type 16 is activated in the absence of E2 proteins by a sequence-aberrant Sp1 distal element. J. Virol. 1990, 64, 5577–5584. [Google Scholar] [CrossRef]
- Bravo, I.G.; Alonso, A. Mucosal human papillomaviruses encode four different E5 proteins whose chemistry and phylogeny correlate with malignant or benign growth. J. Virol. 2004, 78, 13613–13626. [Google Scholar] [CrossRef] [PubMed]
- Lange, A.; Mills, R.E.; Lange, C.J.; Stewart, M.; Devine, S.E.; Corbett, A.H. Classical nuclear localization signals: Definition, function, and interaction with importin alpha. J. Biol. Chem. 2007, 282, 5101–5105. [Google Scholar] [CrossRef] [PubMed]
- Chook, Y.M.; Süel, K.E. Nuclear import by karyopherin-βs: Recognition and inhibition. Biochim. Biophys. Acta 2011, 1813, 1593–1606. [Google Scholar] [CrossRef]
- Zou, N.; Lin, B.Y.; Duan, F.; Lee, K.Y.; Jin, G.; Guan, R.; Yao, G.; Lefkowitzt, E.J.; Broker, T.R.; Chow, L.T. The hinge of the human papillomavirus type 11 E2 protein contains major determinants for nuclear localization and nuclear matrix association. J. Virol. 2000, 74, 3761–3770. [Google Scholar] [CrossRef]
- Florin, L.; Becker, K.A.; Lambert, C.; Nowak, T.; Sapp, C.; Strand, D.; Streeck, R.E.; Sapp, M. Identification of a dynein interacting domain in the papillomavirus minor capsid protein L2. J. Virol. 2006, 80, 6691–6696. [Google Scholar] [CrossRef]
- Chen, Z.; DeSalle, R.; Schiffman, M.; Herrero, R.; Wood, C.E.; Ruiz, J.C.; Clifford, G.M.; Chan, P.K.S.; Burk, R.D. Niche adaptation and viral transmission of human papillomaviruses from archaic hominins to modern humans. PLoS Pathog. 2018, 14, e1007352. [Google Scholar] [CrossRef]
- Delius, H.; Saegling, B.; Bergmann, K.; Shamanin, V.; de Villiers, E.M. The genomes of three of four novel HPV types, defined by differences of their L1 genes, show high conservation of the E7 gene and the URR. Virology 1998, 240, 359–365. [Google Scholar] [CrossRef][Green Version]
- Kovanda, A.; Kocjan, B.J.; Potočnik, M.; Poljak, M. Characterization of a novel cutaneous human papillomavirus genotype HPV-125. PLoS ONE 2011, 6, e22414. [Google Scholar] [CrossRef][Green Version]
- Van Doorslaer, K. Evolution of the Papillomaviridae. Virology 2013, 445, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Faust, H.; Eldenhed Alwan, E.; Roslin, A.; Wennerberg, J.; Forslund, O. Prevalence of human papillomavirus types, viral load and physical status of HPV16 in head and neck squamous cell carcinoma from the South Swedish Health Care Region. J. Gen. Virol. 2016, 97, 2949–2956. [Google Scholar] [CrossRef][Green Version]
- Ball, S.L.; Winder, D.M.; Vaughan, K.; Hanna, N.; Levy, J.; Sterling, J.C.; Stanley, M.A.; Goon, P.K. Analyses of human papillomavirus genotypes and viral loads in anogenital warts. J. Med. Virol. 2011, 83, 1345–1350. [Google Scholar] [CrossRef] [PubMed]
- Breznik, V.; Fujs Komloš, K.; Hošnjak, L.; Luzar, B.; Kavalar, R.; Miljković, J.; Poljak, M. Determination of Causative Human Papillomavirus Type in Tissue Specimens of Common Warts Based on Estimated Viral Loads. Front. Cell. Infect. Microbiol. 2020, 10, 4. [Google Scholar] [CrossRef]
- Nair, S.; Pillai, M.R. Human papillomavirus and disease mechanisms: Relevance to oral and cervical cancers. Oral. Dis. 2005, 11, 350–359. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Macaque Species (Genus Macaca) | PV Type | PV Genus | Sample Origin | Reference |
|---|---|---|---|---|
| cynomolgus macaque (M. fascicularis) | MfPV1, -2 | Beta-PV | palmoplantar skin papillomas | [8,10] |
| MfPV3, -4, -5, -8 | Alpha-PV | cervical intraepithelial neoplasia | [11] | |
| MfPV6, -7, -9, -10, -11 | Alpha-PV | exfoliated cervical cytology samples | [11] | |
| rhesus macaque (M. mulatta) | MmPV1 | Alpha-PV | lymph node metastasis of a penile squamous cell carcinoma | [13] |
| MmPV2 | Alpha-PV | exfoliated vaginal cells | [12] | |
| MmPV3, -6 | Alpha-PV | swabs of the penile surface | [12] | |
| MmPV4, -5, -7 | Gamma-PV | swabs of the penile surface | [12] | |
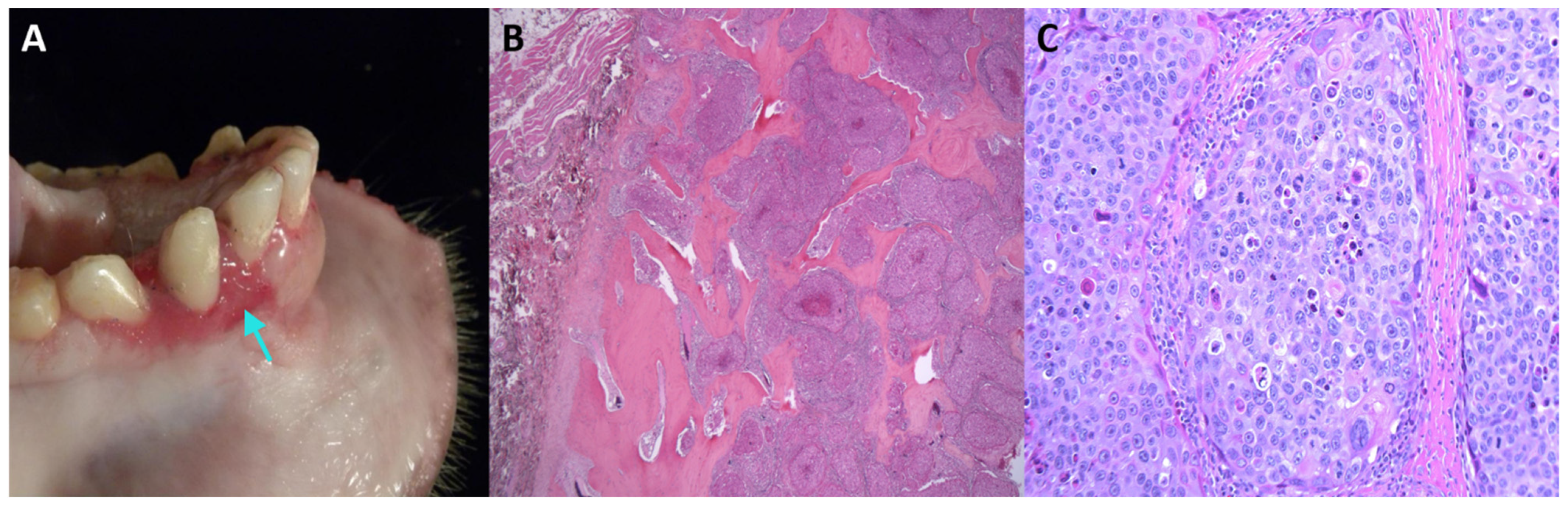
| Japanese macaque (M. fuscata) | MfuPV1 | Alpha-PV | oral squamous cell carcinoma | this study |
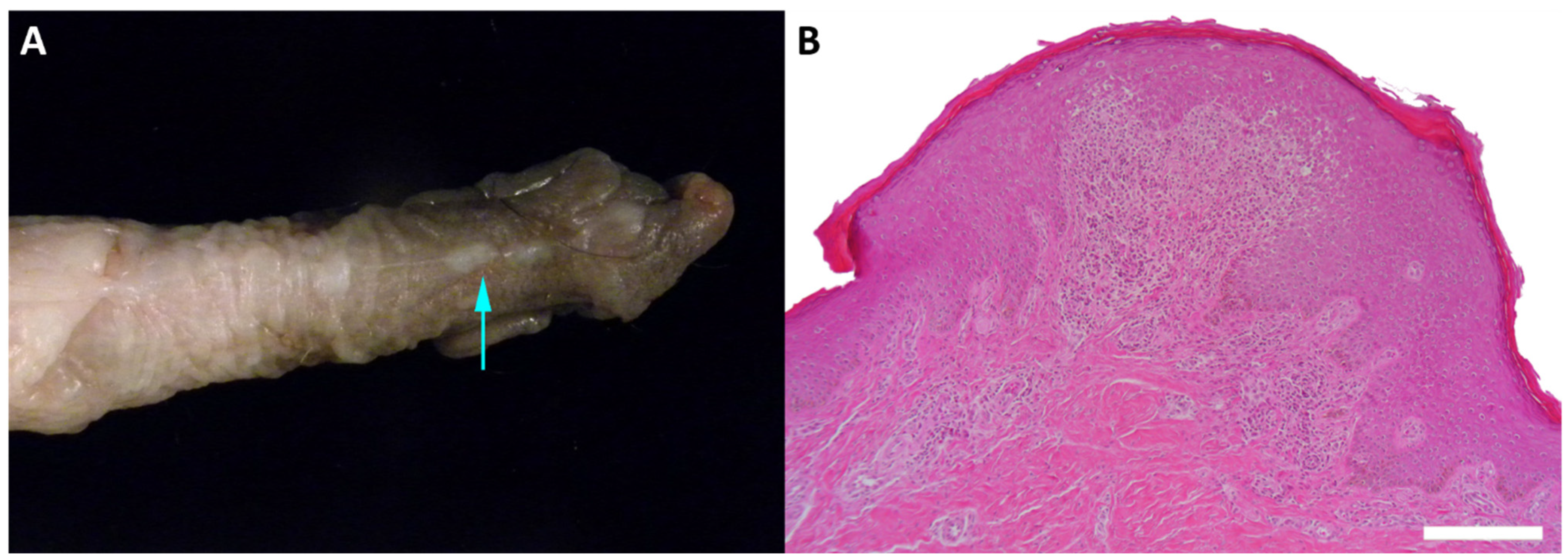
| MfuPV2 | Alpha-PV | benign penile lesion | this study |
| Genomic Region | Motifs and Domains (Consensus Sequence) | MfuPV1 | MfuPV2 |
|---|---|---|---|
| E6 | Zinc-finger domain (CxxC(x)29CxxC) [30] | aa 30–66; | aa 29–65; |
| aa 103–139 | aa 102–138 | ||
| PDZ-binding motif (xT/SxV/L) [31] | aa 148–151 | aa 145–148 | |
| E7 | pRB-binding site (LxCxE) [32] | aa 22–26 | aa 23–27 |
| Zinc-finger domain (CxxC(x)29CxxC) [30] | aa 58–94 | aa 69–105 | |
| E1 | Bipartite NLS [33,34] | aa 85–87…120–123 a | aa 93–95…128–131 |
| NES (Lx(2–3)Lx2(L/I/V)x(L/I)) [34] | aa 106–115 | aa 114–123 | |
| Cyclin-binding motif (RxL) [33,34] | aa 124–126 | aa 132–134 | |
| Cdk-phosphorylation site ((S/T)P) [33] | aa 91–92; | aa 99–100; | |
| aa 95–96; | aa 103–104; | ||
| aa 107–108 | aa 115–116 | ||
| ATP-binding site (Gx4GK(T/S)) [34] | aa 457–464 | aa 482–489 | |
| E2 | NLS (KRxR) [8,35] | aa 242–245 | aa 244–247 |
| Leucine-zipper domain (Lx6Lx6Lx6L) [36] | aa 287–308 | aa 306–327 | |
| E1^E4/E4 | “Leucine-cluster” motif (LLxLL) [37] | aa 14–18 | aa 60–64 a |
| Polyadenylation site (AATAAA) [11] | nt 4224–4229 | nt 4352–4357 | |
| L2 | nNLS [38] | aa 1–11 | aa 1–11 |
| cNLS [38] | aa 455–460 | aa 448–452 | |
| Furin cleavage motif (RxK/RR) [39] | aa 8–11; | aa 8–11; | |
| aa 20–23 | absent | ||
| Transmembrane-like domain (GxxxG motifs) [40] | aa 56–78 | aa 44–66 | |
| NRS [38] | aa 303–323 | aa 292–312 | |
| L1-binding domain (PxxPxxPxxP) [39] | aa 425–434 | aa 414–423 | |
| L1 | Monopartite NLS [41,42] | aa 494–499 | aa 500–505 |
| Bipartite NLS [41,42] | aa 481–482...494–495 | aa 487–488...500–501 | |
| LCR | Polyadenylation site (AATAAA) [11,20] | nt 7273–7278 | nt 7405–7410 |
| Polyadenylation site (ATTAAA) [20] | absent | nt 7444–7449 | |
| E2-binding site (ACC(N)6GGT) [30] | nt 7436–7447; | nt 7606–7617; | |
| nt 7752–7763; | nt 7915–7926; | ||
| nt 7829–7840; | nt 7994–8005; | ||
| nt 7844–7855 | nt 8009–8020 a | ||
| E1-binding region [8,34] | nt 7789–7806 | nt 7954–7971 | |
| TATA box (TATAAA) [43] | nt 7859–7864 | nt 8024–8029 | |
| C/EBPbeta-binding site (T(T/G)NNGNAA(T/G)) [43,44] | nt 7189–7197 a | nt 7780–7788 | |
| YY1-binding site (CCGCCATNTT) [43] | nt 7398–7408 a | absent | |
| AP-1-binding site (TGANTCA) [43] | nt 7556–7562 | nt 7650–7656 | |
| NFI/CTF-binding site (TTGGC) [43] | nt 7624–7628; | nt 7161–7165; | |
| nt 7667–7671 | nt 7719–7723 | ||
| SP-1-binding site (NGGNGN) [43,45] | nt 7822–7827 | nt 7987–7992 |
| Genomic Region | Pairwise Similarity with MfuPV1 a (%) | MfPV5 a | MfPV9 a | MfPV4 a | MmPV1 a | MfPV10 a |
|---|---|---|---|---|---|---|
| E6 | nucleotide sequence | 80.3 | 80.5 | 78.1 | 64.9 b | 75.7 |
| amino acid sequence | 80.3 | 79.6 | 80.3 | 63.6 | 73.7 | |
| E7 | nucleotide sequence | 72.0 b | 71.5 | 75.0 | 66.1 | 69.9 |
| amino acid sequence | 58.9 | 68.0 | 68.0 | 57.9 | 65.4 | |
| E1 | nucleotide sequence | 86.0 | 85.4 | 80.4 | 79.2 | 79.6 |
| amino acid sequence | 88.9 | 88.7 | 82.1 | 82.0 | 82.9 | |
| E2 | nucleotide sequence | 79.6 | 79.1 | 77.4 b | 72.7 | 76.9 |
| amino acid sequence | 73.4 | 72.1 | 72.1 | 64.8 | 69.9 | |
| E1^E4 | nucleotide sequence | 72.7 | 75.4 | 69.5 | 69.7 | 68.9 |
| amino acid sequence | 62.2 | 65.9 | 54.7 | 52.3 | 53.8 | |
| E5-epsilon | nucleotide sequence | 85.7 | 82.5 | 78.0 | 67.4 | 74.4 |
| amino acid sequence | 85.7 | 85.7 | 72.7 | 69.8 | 76.7 | |
| E5-zeta | nucleotide sequence | 80.1 | 76.7 | 68.6 c | 64.6 | 68.8 |
| amino acid sequence | 74.5 | 74.0 | 54.9 | 56.3 | 53.2 | |
| L2 | nucleotide sequence | 79.2 | 77.7 | 74.1 | 72.8 | 73.6 |
| amino acid sequence | 84.5 | 80.7 | 77.5 | 75.9 | 78.5 | |
| L1 | nucleotide sequence | 83.4 | 82.9 | 79.3 | 79.0 | 78.7 |
| amino acid sequence | 90.2 | 90.2 | 87.9 | 85.7 | 88.4 | |
| whole genome | nucleotide sequence | 80.5 | 78.7 | 76.6 | 72.5 | 75.7 |
| amino acid sequence | / | / | / | / | / |
| Genomic Region | Pairwise Similarity with MfuPV2 a (%) | MmPV3 a | HPV160 b | HPV117 b | HPV77 b | HPV29 b |
|---|---|---|---|---|---|---|
| E6 | nucleotide sequence | 69.8 | 56.4 | 56.6 | 57.4 | 58.5 |
| amino acid sequence | 62.4 | 46.3 | 46.3 | 48.3 | 49.0 | |
| E7 | nucleotide sequence | 68.8 | 58.3 | 43.6 c | 59.8 | 59.8 |
| amino acid sequence | 67.0 | 52.3 | 44.9 | 56.1 | 56.1 | |
| E1 | nucleotide sequence | 73.7 | 67.7 | 65.7 | 69.1 | 68.6 |
| amino acid sequence | 75.0 | 66.7 | 65.4 | 68.7 | 66.6 | |
| E2 | nucleotide sequence | 69.3 | 63.6 | 61.9 | 62.8 | 61.1 |
| amino acid sequence | 60.5 | 56.4 | 51.7 | 54.8 | 54.9 | |
| E4 | nucleotide sequence | 42.9 c | 54.0 d | - e | 56.1 d | 40.8 c |
| amino acid sequence | 33.3 | 35.2 | - | 34.3 | 19.6 | |
| E5 | nucleotide sequence | 50.0 f | 47.6 c | 47.2 c | 48.6 d | 51.4 d |
| amino acid sequence | 42.6 | 26.5 | 20.4 | 38.5 | 22.2 | |
| L2 | nucleotide sequence | 70.4 | 63.7 | 63.7 | 63.4 | 65.4 |
| amino acid sequence | 73.2 | 63.9 | 63.7 | 63.2 | 64.6 | |
| L1 | nucleotide sequence | 77.1 | 69.7 | 68.4 | 68.1 | 67.8 |
| amino acid sequence | 82.9 | 72.0 | 72.0 | 72.4 | 71.0 | |
| whole genome | nucleotide sequence | 68.7 | 58.5 | 57.1 | 58.4 | 58.8 |
| amino acid sequence | / | / | / | / | / |
| PV Type | Sample | Concentration of Viral DNA (Copies/µL) | Concentration of Genomic DNA (ng/µL) | Concentration of Genomic DNA (Cell/µL) | Viral Load (Copies/Cell) |
|---|---|---|---|---|---|
| MfuPV1 | oral squamous cell carcinoma | 1.013 × 106 | 10.133 | 1.423 × 103 | 711.876 |
| MfuPV2 | benign penile lesion | 3.04 × 104 | 20.867 | 2.931 × 103 | 10.372 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skubic, L.; Hošnjak, L.; Staheli, J.P.; Dyen, M.R.; Ducore, R.M.; Colgin, L.M.A.; Lewis, A.D.; Poljak, M. Molecular and Phylogenetic Characterization of Novel Papillomaviruses Isolated from Oral and Anogenital Neoplasms of Japanese Macaques (Macaca fuscata). Viruses 2021, 13, 630. https://doi.org/10.3390/v13040630
Skubic L, Hošnjak L, Staheli JP, Dyen MR, Ducore RM, Colgin LMA, Lewis AD, Poljak M. Molecular and Phylogenetic Characterization of Novel Papillomaviruses Isolated from Oral and Anogenital Neoplasms of Japanese Macaques (Macaca fuscata). Viruses. 2021; 13(4):630. https://doi.org/10.3390/v13040630
Chicago/Turabian StyleSkubic, Lucijan, Lea Hošnjak, Jeannette P. Staheli, Michael R. Dyen, Rebecca M. Ducore, Lois M. A. Colgin, Anne D. Lewis, and Mario Poljak. 2021. "Molecular and Phylogenetic Characterization of Novel Papillomaviruses Isolated from Oral and Anogenital Neoplasms of Japanese Macaques (Macaca fuscata)" Viruses 13, no. 4: 630. https://doi.org/10.3390/v13040630
APA StyleSkubic, L., Hošnjak, L., Staheli, J. P., Dyen, M. R., Ducore, R. M., Colgin, L. M. A., Lewis, A. D., & Poljak, M. (2021). Molecular and Phylogenetic Characterization of Novel Papillomaviruses Isolated from Oral and Anogenital Neoplasms of Japanese Macaques (Macaca fuscata). Viruses, 13(4), 630. https://doi.org/10.3390/v13040630