1. Introduction
Caprine arthritis encephalitis virus (CAEV) is a lentivirus in the family Retroviridae that infects goats and is closely related to maedi-visna virus (MVV) of sheep [
1,
2]. As a result of their genetic similarities, these lentiviruses are collectively referred to as small ruminant lentiviruses or SRLVs. CAEV is a monocyte/macrophage-tropic virus that is not associated with immunodeficiency, generally infects dairy goats and was first reported as a retrovirus in 1980 [
3,
4]. Since this time, multiple studies have focused on defining the gross and microscopic lesions of naturally occurring and experimental SRLV infections in small ruminants [
5,
6,
7]. CAEV is known to cause progressive inflammatory syndromes in a variety of tissues including the joint synovium in animals older than 12 months of age (caprine arthritis) or, less often, encephalomyelitis in 2–4-month-old goat kids. CAEV infection can also result in interstitial pneumonia or indurative mastitis (“hard udder”), lesions similar to those observed in sheep infected with MVV [
8]. Histologically, these lesions are characterized by interstitial mononuclear infiltration often featuring lympho-follicular hyperplasia and fibrosis [
9,
10,
11].
CAEV transcripts, in the absence of inflammatory lesions, have been reported in the renal tubular epithelium of goats naturally infected with CAEV (in situ hybridization) [
12]. In these animals, CAE viral transcripts were also identified in a wide variety of tissues including the brain, spinal cord, lung, joint synovium, mammary gland, liver, spleen, lymph node, thyroid follicles and intestinal enterocytes. In sheep, inflammatory renal lesions associated with the deposition of MVV antigen and amplifiable proviral SRLV DNA have been reported in nine animals naturally infected with SRLVs [
13]. These sheep had lesions consistent with interstitial nephritis characterized by infiltrates of lymphocytes, plasma cells and macrophages within the renal interstitium. Lymphoid follicle formation within the interstitium, a feature of SRLV pneumonia, was also reported in the renal tissues of these animals. The affected sheep in this study also had SRLV-associated pulmonary lesions (interstitial pneumonia).
Here we describe the lesional features and their association with CAEV proviral DNA and viral antigen deposition in six goats necropsied at the California Animal Health Food Safety Laboratory (CAHFS), Davis branch, and the University of California, Davis, Veterinary Medical Teaching Hospital (VMTH). In addition to renal lesions, these goats had multisystemic CAEV inflammatory syndromes affecting the joint synovium, lung, mammary gland and/or brain. Four of the six animals also had histological evidence of cardiac lesions. Despite the frequent occurrence of SRLVs and long-term investigation, CAEV-associated renal inflammation and SRLV-associated arterial lesions have not been previously reported in the peer-reviewed literature.
2. Materials and Methods
2.1. Animals
All of the goats included in this study were derived from farms/flocks in Central and Northern California, USA. A database search was conducted using the following keywords: goat, caprine arthritis encephalitis virus, CAE and small ruminant lentivirus. Goats A, E and F were identified prospectively as a CAHFS case in 2019 (A) or UC Davis VMTH cases in 2020 (E) or 2021 (F). Goats B and C were identified retrospectively from amongst all the caprine cases submitted to the CAHFS Laboratory System from 1990 to 2019 (801 animals total). Goat D was identified retrospectively from the UC Davis VMTH database using the same search terms defined above.
2.2. Pathology and Immunohistochemistry
Goats A, B and C were serologically tested for antibodies to SRLVs using the Small Ruminant Lentivirus Antibody Test Kit, cELISA (VMRD, Pullman, Washington) and were subsequently euthanized for quality of life issues. Goat D had a one-year history of progressive lameness affecting the left stifle and right carpal joints. Due to a deterioration in quality of life, euthanasia was elected. Goat E was treated for respiratory distress and pyrexia for 12 days using antibiotics and corticosteroids. As a result of progressive clinical deterioration in the face of therapy, euthanasia was recommended (goats E and F). Goats D and E were serologically tested with the VMRD cELISA postmortem using extracellular fluid obtained from frozen/thawed splenic tissue or serum, respectively. Goats A–E were adult does while goat F was a wether.
A complete necropsy was performed for each animal and all gross abnormalities were recorded. A complete set of tissues including lung, joint synovium, mammary gland, brain, heart and kidney were obtained for ancillary diagnostic studies. Tissues were immediately fixed in 10% buffered formalin for a minimum of 24 h, trimmed, embedded in paraffin and routinely processed for histological examination. Select tissues with lesions consistent with CAEV-associated inflammation were further evaluated by an immunohistochemistry (IHC) assay to detect CAEV antigen (MAb CAEV-63, CAEV-Co IgG1, CAEP10A1, VMRD, Pullman, WA). All of the immunohistochemical stains were performed at the CAHFS Davis laboratory. Each IHC stain was performed in parallel with known CAEV-positive caprine control tissue and an irrelevant isotype-control antibody as the negative control.
2.3. DNA Isolation, Amplification and Sequencing
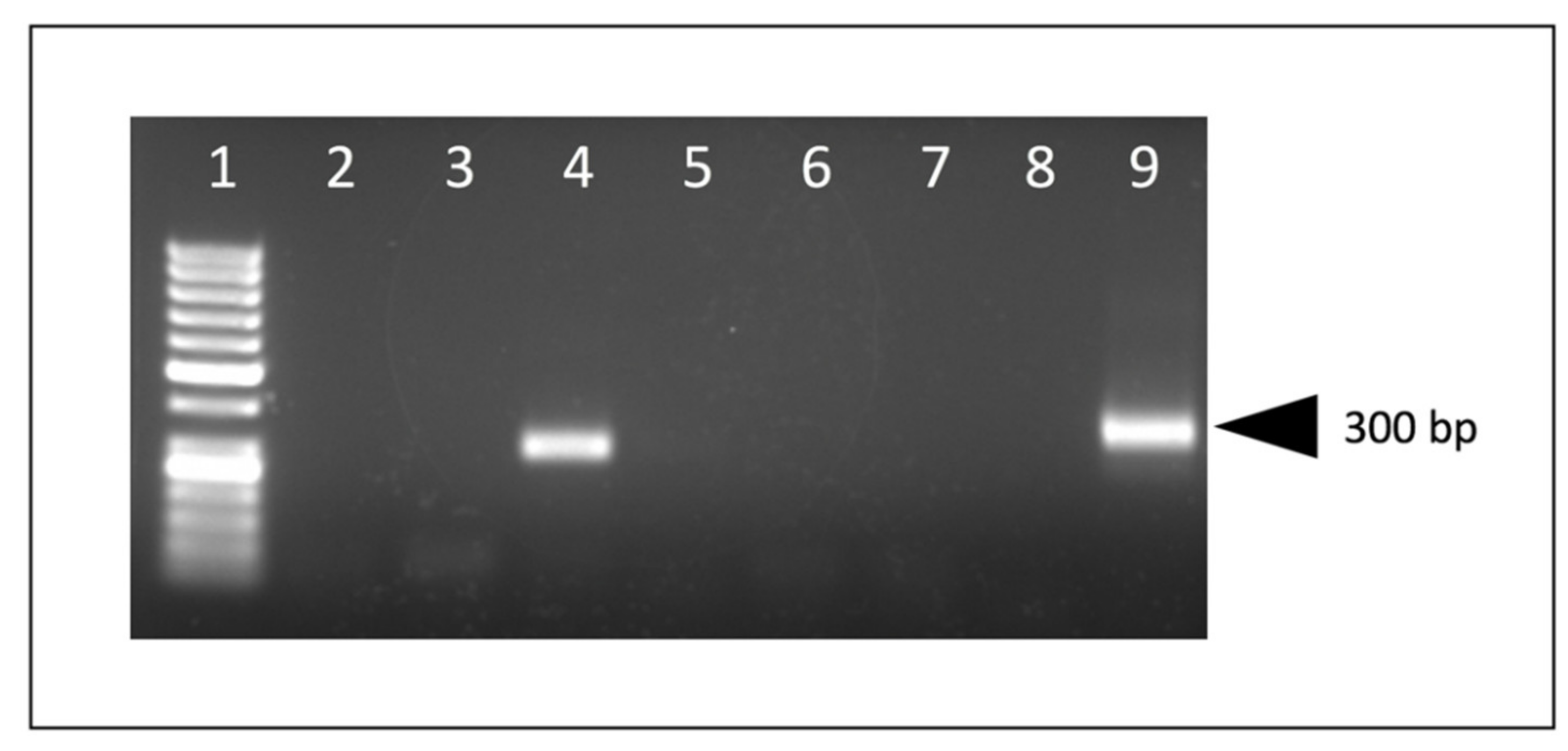
Genomic DNA (proviral DNA) was isolated from either formalin-fixed paraffin-embedded (FFPE) scrolls of caprine tissue using QIAGEN’s QIAamp DNA FFPE Tissue Kit or from fresh-frozen tissue using QIAamp DNA Mini or Blood Mini Kit following the manufacturer’s protocol. The entire 3′ CAEV U3 region was amplified using a nested PCR strategy with recombinant Taq DNA Polymerase (Invitrogen, Waltham, MA, USA) following the manufacturer’s instructions for a 50 μL volume reaction. Primers that flank the 3′ CAEV U3 region, Revfor (5′-CTG ACG ATG GGA ATC TGG ATA AAT GG), and R region of the long terminal repeat, R2rev (5′-CTC GGT ACC TCC TCG GAG AGG AGA G), were used for a first round of amplification (456 nucleotide amplicon) under the following cycling conditions: 95 °C for 2 min; 25 cycles of 95 °C for 15 s, 56 °C for 30 s and 72 °C for 30 s; and a final extension step at 72 °C for 5 min. A second round of amplification was performed under the same cycling conditions with the substitution of 50 cycles instead of 25, using 5 μL of the first PCR reaction along with the primers CAEVnested LTRfor (5′-TAA ATG GAM RGC KTG GAG AAC ACC WC) and CAEVnested LTRrev (5′-CTA GGA GSR MST CTC CYA GAA CTC) resulting in an amplicon of ~359 nucleotides that included the entire 3′ CAEV U3 region. All of the primer binding sites were located outside of the U3 region. Resulting amplicons from both the primary and nested PCR were screened on an agarose gel for size. A water template negative control was run in parallel with each PCR reaction set.
The nested PCR products of goats A, D and E were directly sequenced from both the 5′ and 3′ ends using the amplifying primers, CAEV
nested LTR
for and CAEV
nested LTR
rev. Prior to direct sequencing, PCR amplicons were concentrated and purified using Zymo Research’s DNA Clean and Concentrator Kit (goat A) or with Amicon Ultra-0.5 Centrifugal Filter Unit (goats D and E). The consensus sequence was based upon both the forward and reverse sequences. The PCR amplicons of goats B, C and F were ligated into the pCR 2.1 plasmid using Invitrogen’s TA Cloning Kit and transformed into One Shot TOP10 Chemically Competent
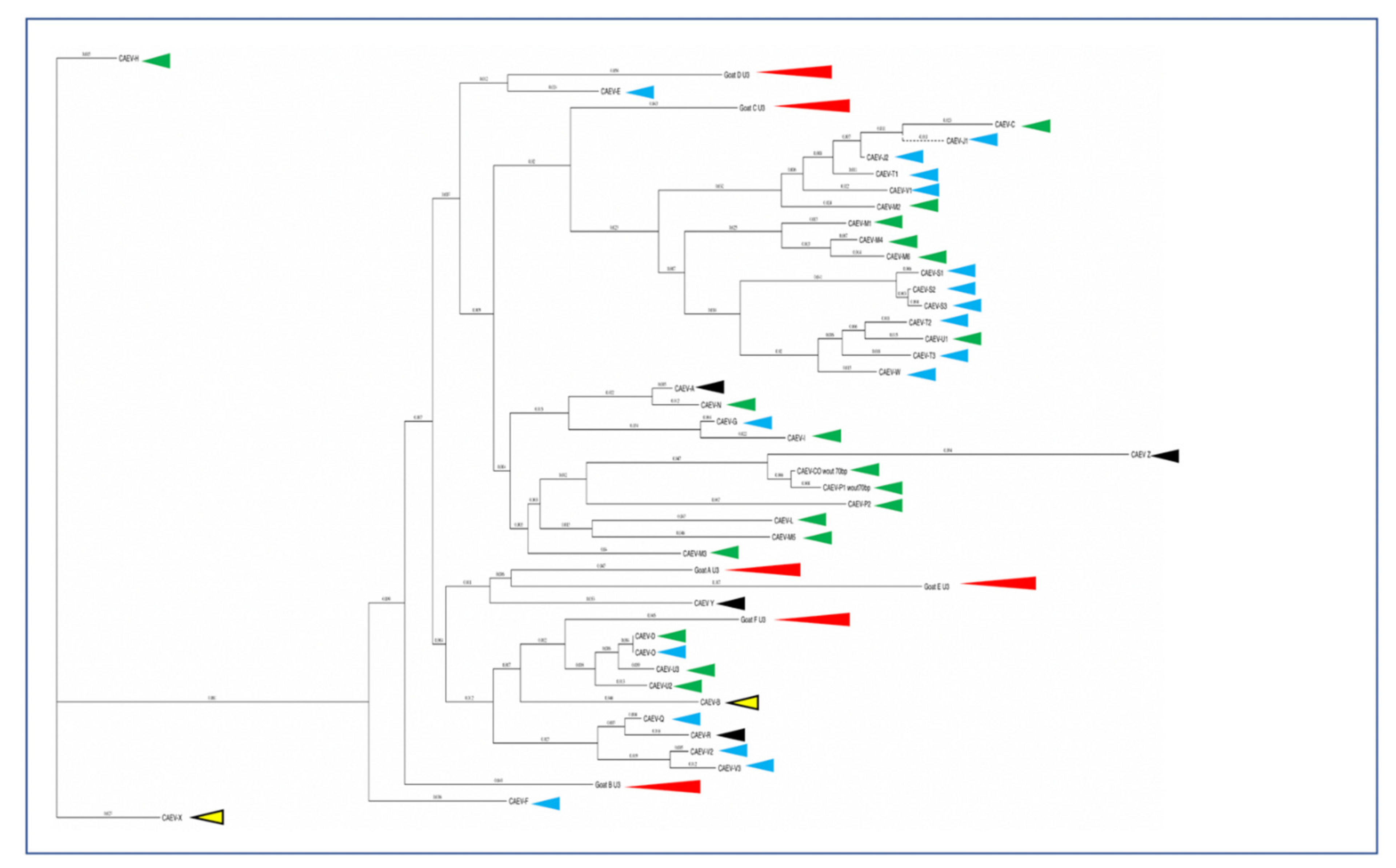
E. coli (ThermoFisher Scientific, Waltham, MA, USA). Three white bacterial colonies from each transformation reaction were amplified overnight in 5 mL of LB broth containing 100 μg/mL carbenicillin. Bacterial plasmids were purified using ZymoPURE Plasmid Miniprep Kit (Zymo Research, Irvine, CA) and were submitted for sequencing at the UC Davis College of Biological Sciences UCDNA Sequencing Facility. Amplicon sequences were digitally trimmed to include the entire U3 region and were aligned using MacVector software. A phylogenetic dendrogram of CAEV U3 sequences amplified from various tissues (mammary gland, lung, brain/spinal cord, joint synovium and kidney) of CAEV-infected goats in North America [
14] was created using 48 sequences and MacVector software (nucleic acid alignment, MacVector 14.0, Apex, North Carolina) [
14,
15,
16].
4. Discussion
The clinical diagnosis of SRLV infections in small ruminants is typically based upon the identification of characteristic clinical signs of caprine arthritis, interstitial pneumonia, mastitis or disease of the central nervous system and then confirmed using serology. Renal and vascular diseases are not considered to be a typical clinical feature of CAEV infection, and renal lesions in goats infected with CAEV are rare. Here we describe renal interstitial nephritis lesions associated with amplifiable CAEV provirus in the kidneys of five dairy goats (Oberhasli, Nubian, Nigerian Dwarf and Alpine) and one non-dairy goat doe (Boer) with various multisystemic CAEV infections. Five of the six goats were serologically positive for CAEV antibodies; serum from goat F was not available for testing.
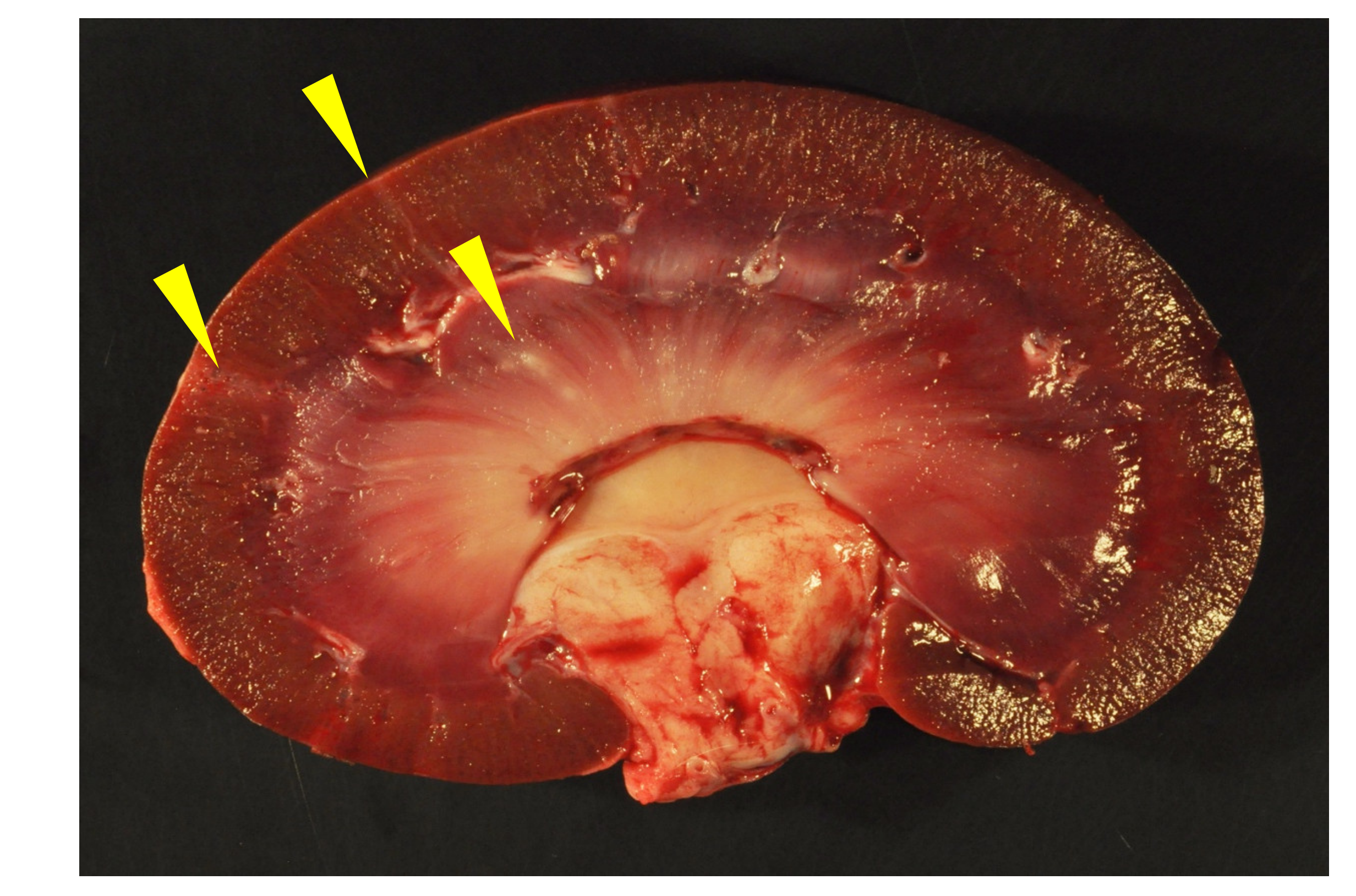
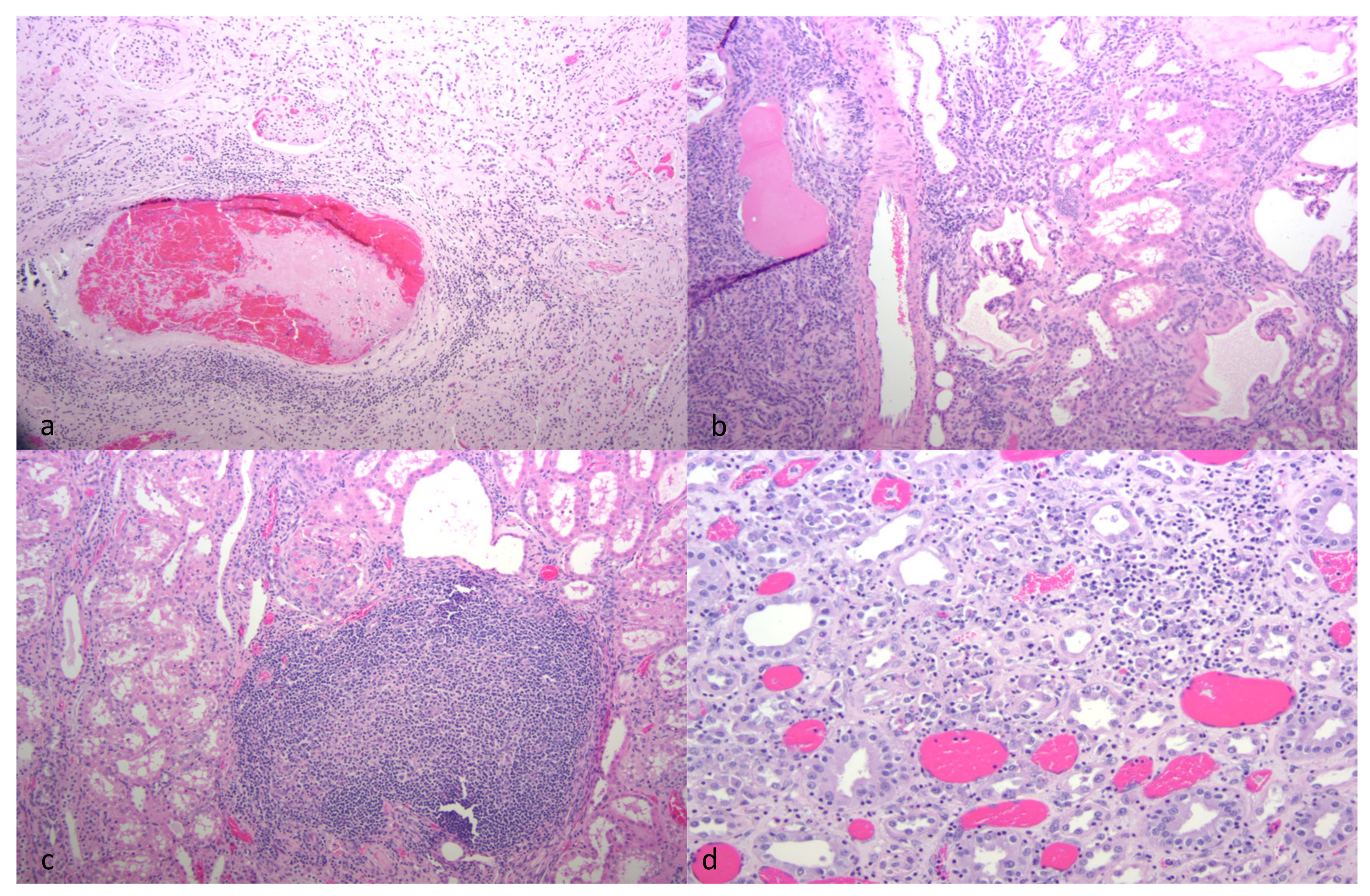
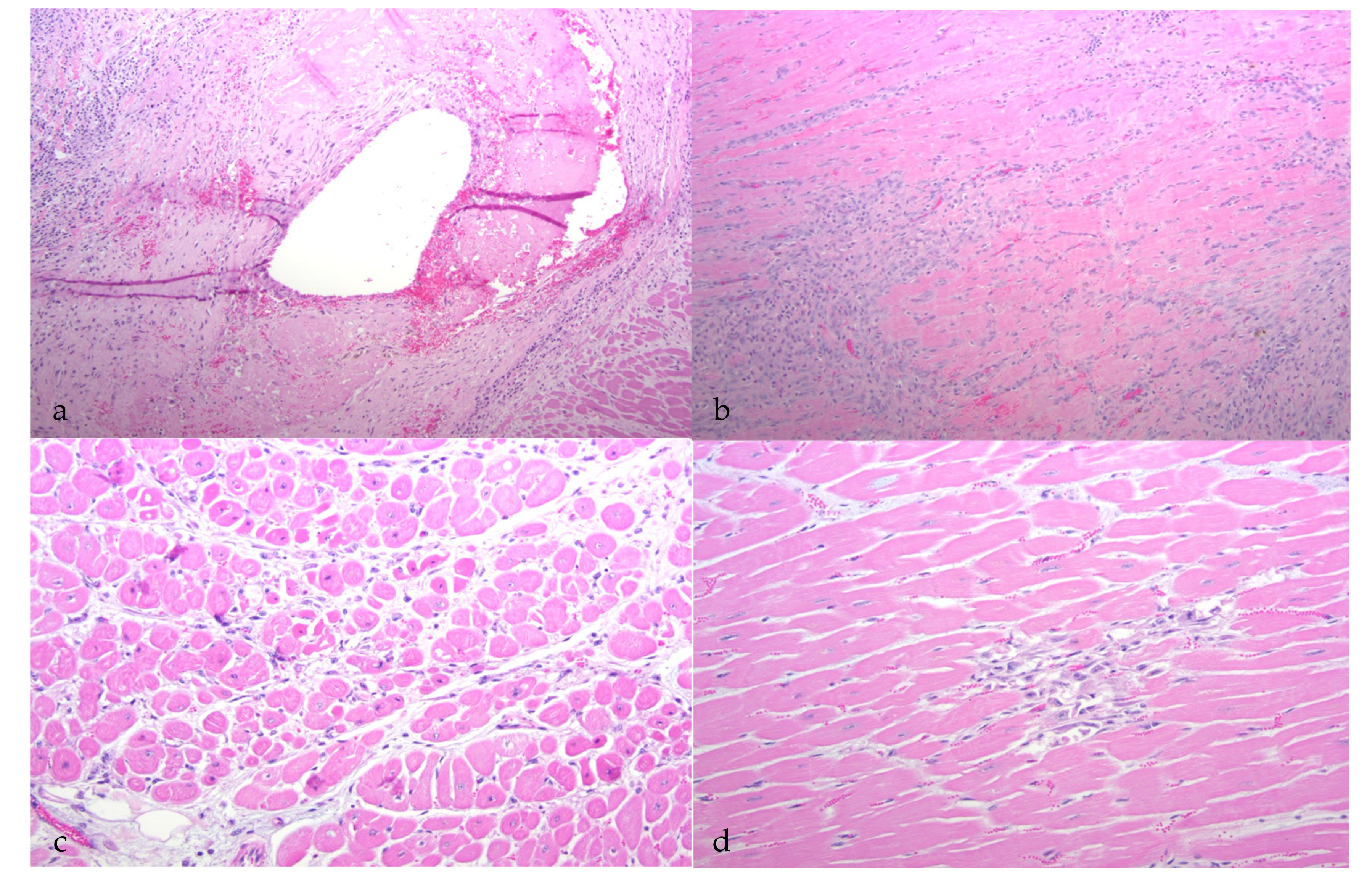
Although all of the animals had histological evidence of renal inflammation (interstitial nephritis), gross lesions in the kidneys were identified and reported in only three of the five goats. Gross renal lesions were identified as cortical infarcts (goat A) or multifocal pinpoint pale foci in either the cortex or medulla (goats B and D). Although the severity of the interstitial nephritis lesions varied markedly from animal to animal, all of the interstitial lesions featured a predominance of lymphocytes, with the lesions of some animals also demonstrating small numbers of plasma cells, neutrophils and/or the deposition of interstitial fibrous tissue. Lesion severity likely explains why the gross renal lesions were variably detected. One animal (goat B) had lymphoid follicle-like structures within the renal interstitium, a common histological feature in the lung and joint tissue of CAEV-infected animals. Inflammatory lesions in goat B were associated with tubular necrosis. In addition to the interstitial nephritis lesions identified in all of the goats, glomerular lesions were found in three animals (goats B, E and F). Mural arteritis lesions were identified in both the kidney and the myocardium of goat A associated with vascular thrombosis and tissue infarction. To our knowledge, this CAEV-associated lesion has not been reported previously in the peer-reviewed literature.
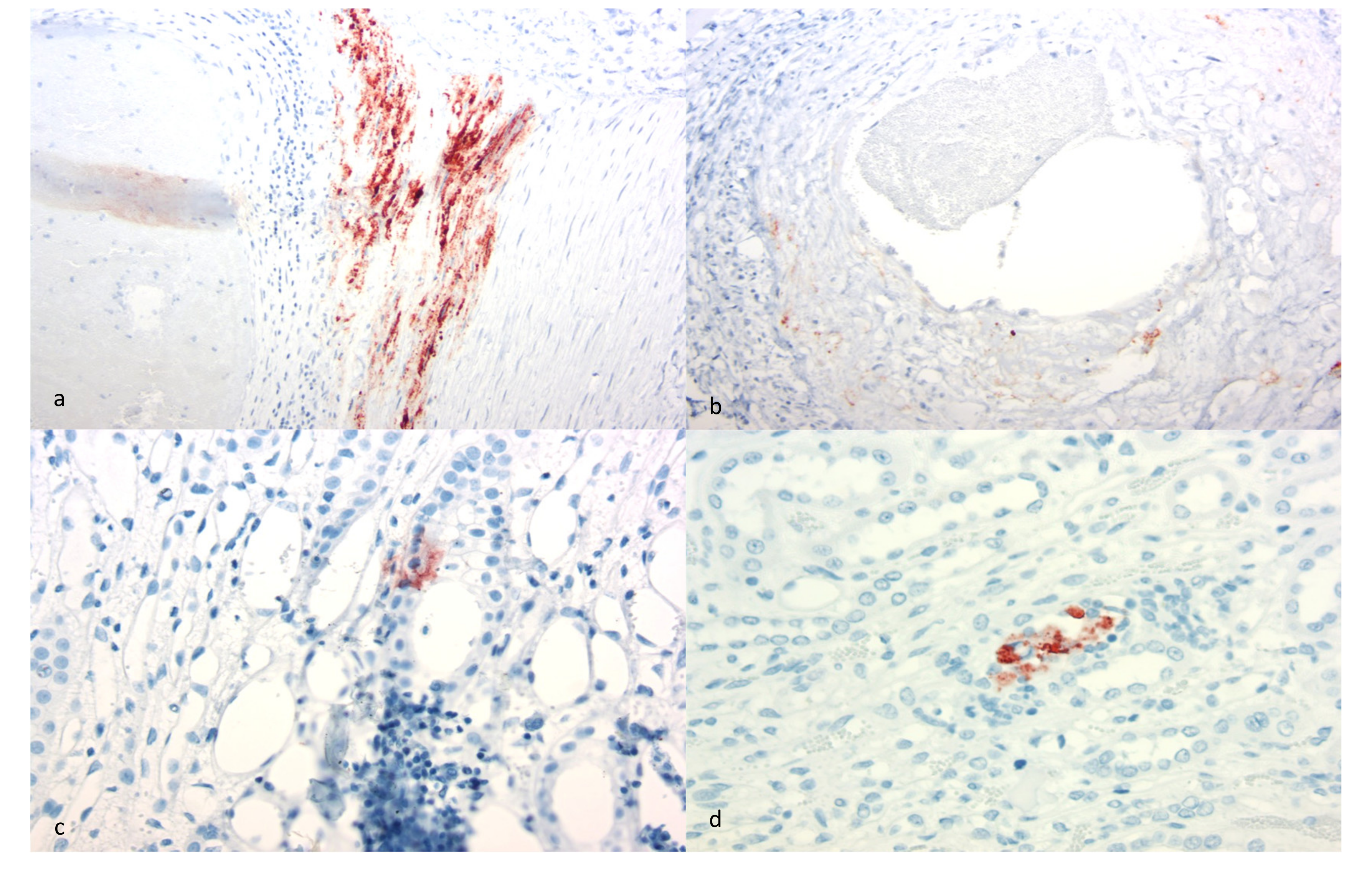
Although interstitial nephritis was identified in all of the goats, deposition of CAEV antigen was identified in only three animals by IHC (goats A, C and E). Viral antigen was also identified within the mural arteritis lesions of both the kidney and heart in goat A and within scattered medullary tubules (goats C and E) and endothelial cells (goat C). Whether the immunopositive material identified within the arteritis lesions represents smooth muscle cells or infiltrating macrophages was not determined. Although CAE viral RNA has been previously identified in the renal tubular epithelium [
12], CAE viral antigen has not. The immunohistochemical detection of CAEV antigen remains a problematic aspect of CAEV diagnosis. Although the antibody utilized for this IHC assay is one of the antibodies used in the competitive ELISA serology test (VMRD), the sensitivity and specificity of the SRLV IHC assay have been questioned. Florid arthritis, mastitis and pneumonia lesions in CAEV-infected goats rarely demonstrate the presence of abundant CAEV antigen using IHC-based detection methods (authors’ personal experience). This may reflect limited viral antigen expression within the lesion or may be the result of assay insensitivity. The specificity of the IHC assay has also been questioned as pulmonary nematodes, intra-alveolar
Pneumocystis organisms and caprine epithelial cells can stain as false positives with the CAEV IHC assay (authors’ personal experience). For these reasons, we confirmed the etiology using both serology and the tissue-specific presence of CAEV provirus using a nested PCR amplification strategy followed by sequencing.
Although CAEV is classically thought to be a monocyte/macrophage-tropic lentivirus, some studies have provided evidence that the viral host cell range may actually be broader. CAE viral transcripts have been identified in a wide variety of tissue types including the renal tubular epithelial cells, brain, spinal cord, lung, joint synovium, mammary gland, liver, spleen, lymph node, thyroid follicles and intestinal enterocytes [
12]. Notably absent from this list of tissues are the smooth muscle cells of the arterial tunica media. It is important to acknowledge that this study does not allow the determination of whether or not these inflammatory lesions in the kidney and heart were actually caused by CAEV.
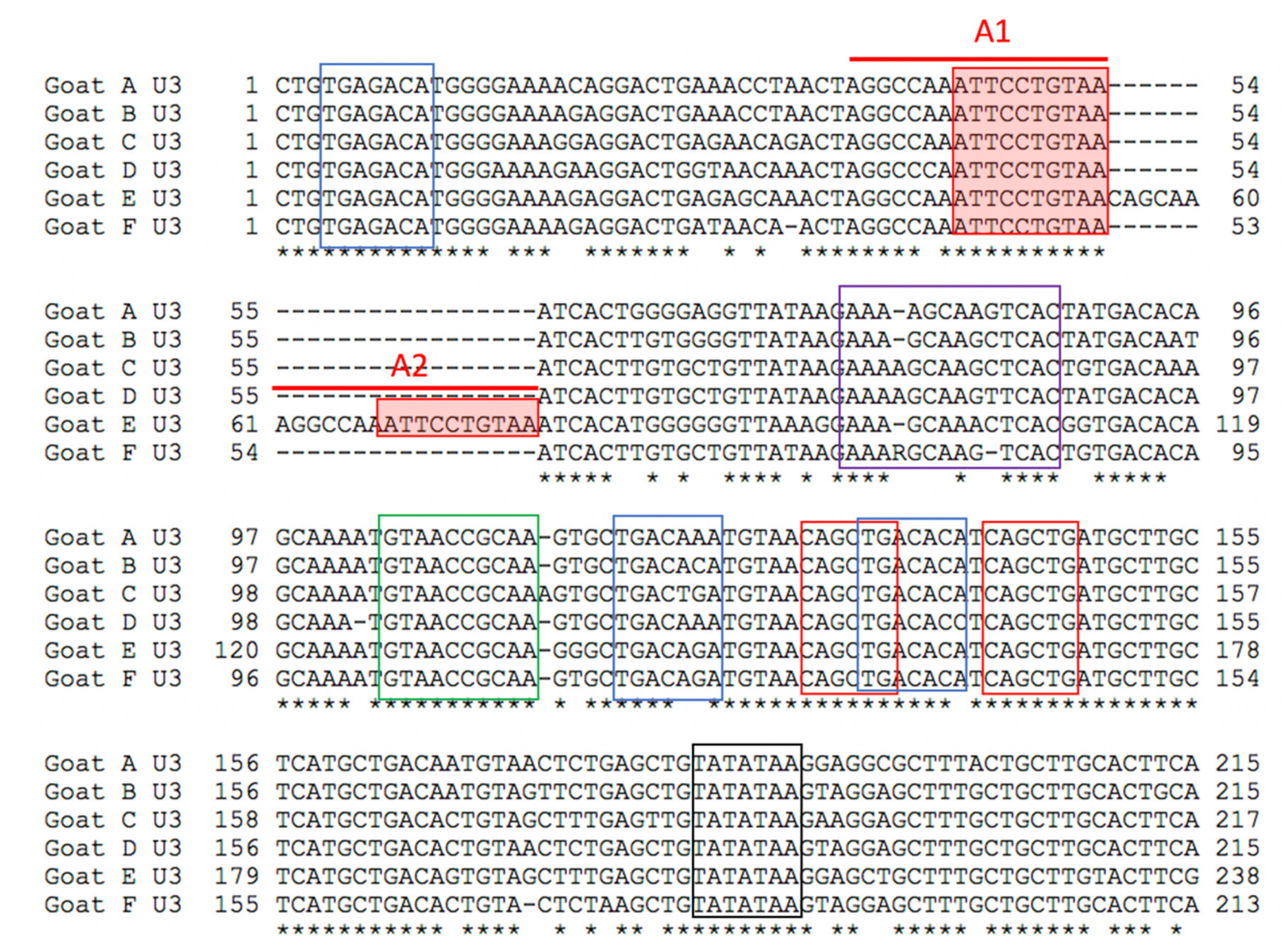
In these six CAEV U3 sequences, several of the transcription factor binding sites (GAS, AML, AP4 and TATA) are conserved, while other sites (AP1 and TAS) demonstrate some degree of sequence polymorphism. This pattern has been described previously for CAEV U3 sequences isolated from naturally infected goats [
14]. The biological relevance of several of these sites has been previously explored [
17,
18,
19]. The interferon gamma-mediated transcriptional activation of CAE viral transcription is mediated through the STAT1 pathway and requires the 10 nucleotide GAS within the CAE promoter [
20]. The CAEV transcriptional response to IFN gamma has been shown to be proportional to the number of copies of GAS in the CAEV promoter [
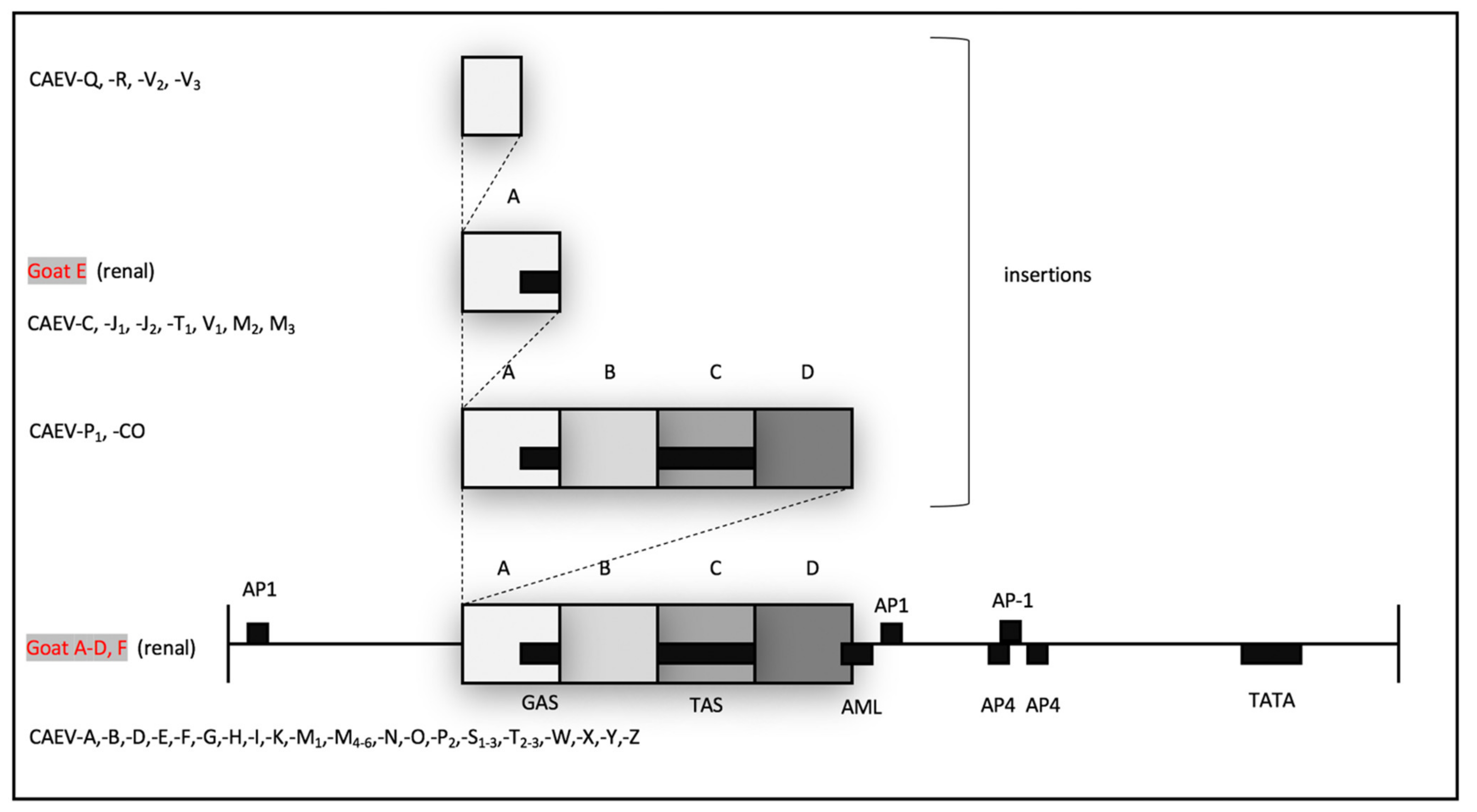
14]. The CAEV-Co isolate, first isolated and described in the 1970s and 1980s (Linda Cork and collaborators), has a conserved 70 base pair repeat (tandem direct repeat) containing the A block at the 5′ end. A naturally occurring CAE isolate encoding an identical 70 base pair repeat has only been described one other time, isolated from the central nervous system of a young Toggenburg goat from the University of Minnesota (CAEV-P1) [
14]. However, 14 different CAE viral isolates encoding A block (or sub A block) repeats have been sequenced from the tissues of goats naturally infected with CAEV. Whether two copies of the GAS confer a selective advantage for viral replication (transcription) remains speculative. Although two of these renal isolates (A and E) form a phylogenetic cluster, the other renal isolates do not. As a result, there does not appear to be an association between the tissue of origin and the sequence of the CAEV U3 promoter. This study describes renal and arterial lesions in goats infected with CAEV, expanding the typical quartet of presentations of CAEV infection; however, the complete significance and viral causation of these lesions need further investigation.


 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}