
Innate Immune Gene Polymorphisms and COVID-19 Prognosis
 ,
,  , , ,
, , ,  , ,
, ,  and
and
Abstract
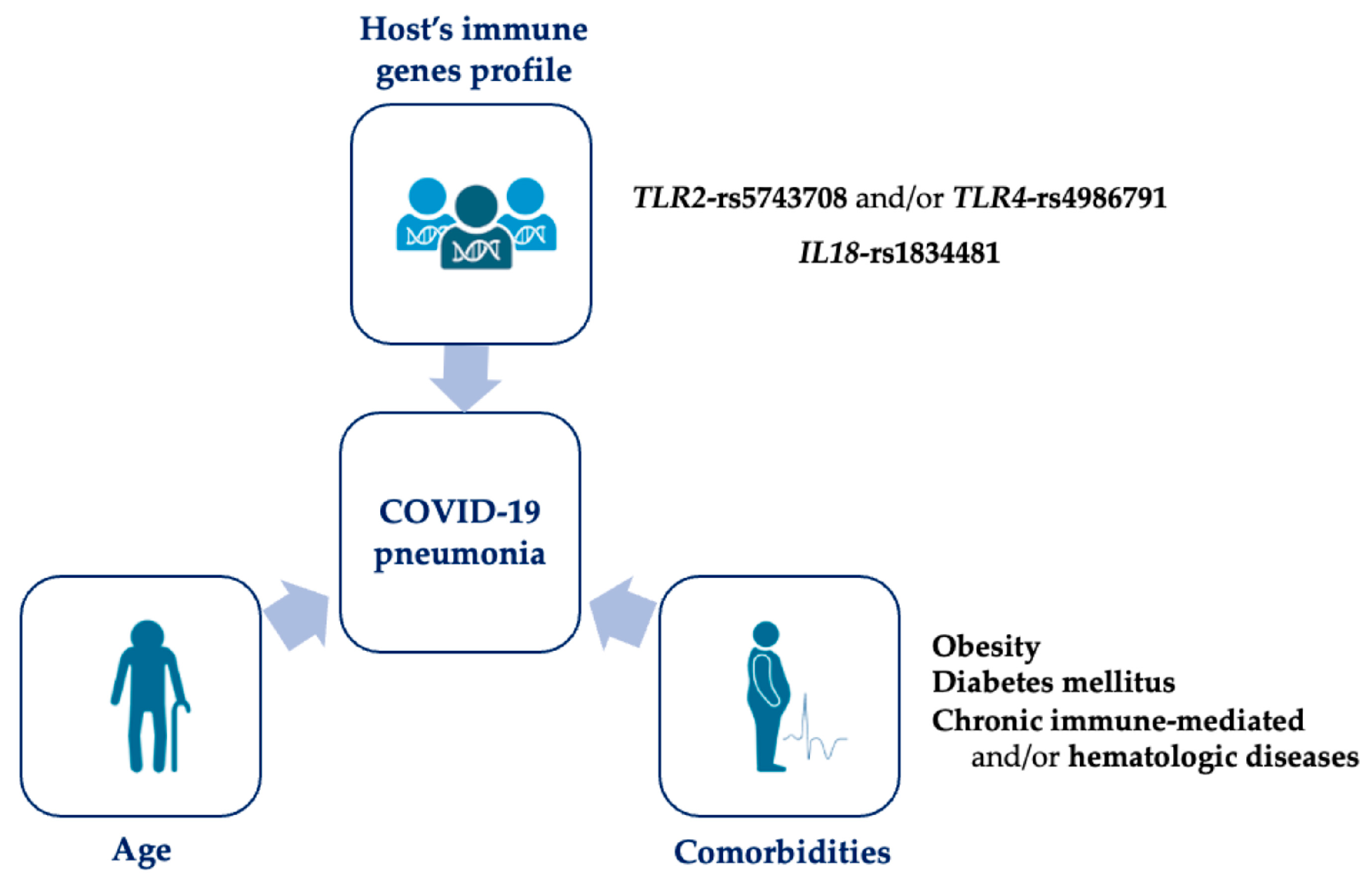
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Molecular Analyses
2.3. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jin, Y.; Yang, H.; Ji, W.; Wu, W.; Chen, S.; Zhang, W.; Duan, G. Virology, Epidemiology, Pathogenesis, and Control of COVID-19. Viruses 2020, 12, 372. [Google Scholar] [CrossRef] [PubMed]
- Kompaniyets, L.; Pennington, A.F.; Goodman, A.B.; Rosenblum, H.G.; Belay, B.; Ko, J.Y.; Chevinsky, J.R.; Schieber, L.Z.; Summers, A.D.; Lavery, A.M.; et al. Underlying Medical Conditions and Severe Illness Among 540,667 Adults Hospitalized with COVID-19, March 2020–March 2021. Prev. Chronic. Dis. 2021, 18, E66. [Google Scholar] [CrossRef] [PubMed]
- Speletas, M.; Dadouli, K.; Syrakouli, A.; Gatselis, N.; Germanidis, G.; Mouchtouri, V.A.; Koulas, I.; Samakidou, A.; Nikolaidou, A.; Stefos, A.; et al. MBL Deficiency-Causing B Allele (Rs1800450) as a Risk Factor for Severe COVID-19. Immunobiology 2021, 226, 152136. [Google Scholar] [CrossRef] [PubMed]
- Dalekos, G.N.; Stefos, A.; Georgiadou, S.; Lygoura, V.; Michail, A.; Ntaios, G.; Samakidou, A.; Giannoulis, G.; Gabeta, S.; Vlychou, M.; et al. Lessons from Pathophysiology: Use of Individualized Combination Treatments with Immune Interventional Agents to Tackle Severe Respiratory Failure in Patients with COVID-19. Eur. J. Intern. Med. 2021, 88, 52–62. [Google Scholar] [CrossRef] [PubMed]
- Moon, A.M.; Webb, G.J.; García-Juárez, I.; Kulkarni, A.V.; Adali, G.; Wong, D.K.; Lusina, B.; Dalekos, G.N.; Masson, S.; Shore, B.M.; et al. SARS-CoV-2 Infections Among Patients With Liver Disease and Liver Transplantation Who Received COVID-19 Vaccination. Hepatol. Commun. 2021, 6, 889–897. [Google Scholar] [CrossRef] [PubMed]
- Gatselis, N.K.; Lygoura, V.; Lyberopoulou, A.; Giannoulis, G.; Samakidou, A.; Vaiou, A.; Vatidis, G.; Antoniou, K.; Stefos, A.; Georgiadou, S.; et al. Soluble IL-2R Levels at Baseline Predict the Development of Severe Respiratory Failure and Mortality in COVID-19 Patients. Viruses 2022, 14, 787. [Google Scholar] [CrossRef]
- Ovsyannikova, I.G.; Haralambieva, I.H.; Crooke, S.N.; Poland, G.A.; Kennedy, R.B. The Role of Host Genetics in the Immune Response to SARS-CoV-2 and COVID-19 Sus-ceptibility and Severity. Immunol. Rev. 2020, 296, 205–219. [Google Scholar] [CrossRef]
- Adli, A.; Rahimi, M.; Khodaie, R.; Hashemzaei, N.; Hosseini, S.M. Role of Genetic Variants and Host Polymorphisms on COVID-19: From Viral Entrance Mechanisms to Immunological Reactions. J. Med. Virol. 2022, 94, 1846–1865. [Google Scholar] [CrossRef]
- Zepeda-Cervantes, J.; Martínez-Flores, D.; Ramírez-Jarquín, J.O.; Tecalco-Cruz, Á.C.; Alavez-Pérez, N.S.; Vaca, L.; Sarmiento-Silva, R.E. Implications of the Immune Polymorphisms of the Host and the Genetic Variability of SARS-CoV-2 in the Development of COVID-19. Viruses 2022, 14, 94. [Google Scholar] [CrossRef]
- Thair, S.A.; He, Y.D.; Hasin-Brumshtein, Y.; Sakaram, S.; Pandya, R.; Toh, J.; Rawling, D.; Remmel, M.; Coyle, S.; Dalekos, G.N.; et al. Transcriptomic Similarities and Differences in Host Response between SARS-CoV-2 and Other Viral Infections. iScience 2020, 24, 101947. [Google Scholar] [CrossRef]
- El-Hefnawy, S.M.; Eid, H.A.; Mostafa, R.G.; Soliman, S.S.; Omar, T.A.; Azmy, R.M. COVID-19 Susceptibility, Severity, Clinical Outcome and Toll-like Receptor (7) MRNA Expression Driven by TLR7 Gene Polymorphism (Rs3853839) in Middle-Aged Individuals without Previous Comorbidities. Gene Rep. 2022, 27, 101612. [Google Scholar] [CrossRef]
- Pojero, F.; Candore, G.; Caruso, C.; Di Bona, D.; Groneberg, D.A.; Ligotti, M.E.; Accardi, G.; Aiello, A. The Role of Immunogenetics in COVID-19. Int. J. Mol. Sci. 2021, 22, 2636. [Google Scholar] [CrossRef] [PubMed]
- Diamond, M.S.; Kanneganti, T.-D. Innate Immunity: The First Line of Defense against SARS-CoV-2. Nat. Immunol. 2022, 23, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Paludan, S.R.; Mogensen, T.H. Innate immunological pathways in COVID-19 pathogenesis. Sci. Immunol. 2022, 7, eabm5505. [Google Scholar] [CrossRef]
- Hatzianastasiou, S.; Mouchtouri, V.A.; Pavli, A.; Tseroni, M.; Sapounas, S.; Vasileiou, C.; Dadouli, K.; Kyritsi, M.; Koureas, M.; Prezerakos, P.; et al. COVID-19 Outbreak on a Passenger Ship and Assessment of Response Measures, Greece, 2020. Emerg. Infect. Dis. 2021, 27, 1927–1930. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Clinical Management of COVID-19: Living Guideline, 15 September 2022. World Health Organization. 2022. Available online: https://apps.who.int/iris/handle/10665/362783 (accessed on 21 July 2022).
- Lorenz, E.; Frees, K.L.; Schwartz, D.A. Determination of the TLR4 Genotype Using Allele-Specific PCR. BioTechniques 2001, 31, 22–24. [Google Scholar] [CrossRef]
- Liadaki, K.; Petinaki, E.; Skoulakis, C.; Tsirevelou, P.; Klapsa, D.; Germenis, A.E.; Speletas, M. Toll-like receptor 4 gene (TLR4), but not TLR2, polymorphisms modify the risk of tonsillar disease due to Streptococcus pyogenes and Haemophilus influenzae. Clin. Vaccine Immunol. 2011, 18, 217–222. [Google Scholar] [CrossRef]
- Speletas, M.; Bakaros, E.; Peristeri, A.-M.; Voulgaridi, I.; Sarrou, S.; Paliatsa, V.; Nasika, A.; Tseroni, M.; Anagnostopoulos, L.; Theodoridou, K.; et al. The Rs1883832 Polymorphism (CD40-1C>T) Affects the Intensity of IgA Responses after BNT162b2 Vaccination. Int J Mol Sci 2022, 23, 14056. [Google Scholar] [CrossRef]
- Mukherjee, S.; Karmakar, S.; Babu, S.P.S. TLR2 and TLR4 Mediated Host Immune Responses in Major Infectious Diseases: A Review. Br. J. Infect. Dis. 2016, 20, 193–204. [Google Scholar] [CrossRef]
- Rodrigues, T.S.; de Sá, K.S.G.; Ishimoto, A.Y.; Becerra, A.; Oliveira, S.; Almeida, L.; Gonçalves, A.V.; Perucello, D.B.; Andrade, W.A.; Castro, R.; et al. Inflammasomes Are Activated in Response to SARS-CoV-2 Infection and Are Associated with COVID-19 Severity in Patients. J. Exp. Med. 2020, 218, e20201707. [Google Scholar] [CrossRef]
- Satış, H.; Özger, H.S.; Aysert Yıldız, P.; Hızel, K.; Gulbahar, Ö.; Erbaş, G.; Aygencel, G.; Guzel Tunccan, O.; Öztürk, M.A.; Dizbay, M.; et al. Prognostic Value of Interleukin-18 and Its Association with Other Inflammatory Markers and Disease Severity in COVID-19. Cytokine 2021, 137, 155302. [Google Scholar] [CrossRef]
- Nolan, K.F.; Greaves, D.R.; Waldmann, H. The Human Interleukin 18 GeneIL18Maps to 11q22.2–Q22.3, Closely Linked to the DRD2 Gene Locus and Distinct from Mapped IDDM Loci. Genomics 1998, 51, 161–163. [Google Scholar] [CrossRef]
- He, M.; Cornelis, M.C.; Kraft, P.; van Dam, R.M.; Sun, Q.; Laurie, C.C.; Mirel, D.B.; Chasman, D.I.; Ridker, P.M.; Hunter, D.J.; et al. Genome-Wide Association Study Identifies Variants at IL18-BCO2 Locus Associated with Interleukin-18 Levels. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 885–890. [Google Scholar] [CrossRef] [PubMed]
- Vergara, C.; Thio, C.; Latanich, R.; Cox, A.L.; Kirk, G.D.; Mehta, S.H.; Busch, M.; Murphy, E.L.; Villacres, M.C.; Peters, M.G.; et al. Genetic Basis for Variation in Plasma IL-18 Levels in Persons with Chronic Hepatitis C Virus and Human Immunodeficiency Virus-1 Infections. Genes Immun. 2017, 18, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Koch, W.; Wolferstetter, H.; Schatke, A.; Schömig, A.; Kastrati, A. Interleukin 18 Gene Variation and Risk of Acute Myocardial Infarction. Cytokine 2011, 56, 786–791. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Dang, S.; Zhang, G.; He, H.; Wen, X. Genetic Polymorphisms of IL-10, IL-18 and IL12B Are Associated with Risk of Non-Small Cell Lung Cancer in a Chinese Han Population. Int. Immunopharmacol. 2019, 77, 105938. [Google Scholar] [CrossRef]
- Pontillo, A.; Bricher, P.; Leal, V.N.C.; Lima, S.; Souza, P.R.E.; Crovella, S. Role of Inflammasome Genetics in Susceptibility to HPV Infection and Cervical Cancer Development. J. Med. Virol. 2016, 88, 1646–1651. [Google Scholar] [CrossRef]
- Cambui, R.A.G.; do Espírito Santo, G.F.; Fernandes, F.P.; Leal, V.N.C.; Galera, B.B.; Fávaro, E.G.P.; Rizzo, L.A.; Elias, R.M.; Pontillo, A. Double-Edged Sword of Inflammasome Genetics in Colorectal Cancer Prognosis. Clin. Immunol. 2020, 213, 108373. [Google Scholar] [CrossRef]
- Park, S.; Hisham, Y.; Shin, H.M.; Yeom, S.C.; Kim, S. Interleukin-18 Binding Protein in Immune Regulation and Autoimmune Diseases. Biomedicines 2022, 10, 1750. [Google Scholar] [CrossRef]
- Yasuda, K.; Nakanishi, K.; Tsutsui, H. Interleukin-18 in Health and Disease. Int. J. Mol. Sci. 2019, 20, 649. [Google Scholar] [CrossRef]
- Leal, V.N.C.; Paulino, L.M.; Cambui, R.A.G.; Zupelli, T.G.; Yamada, S.M.; Oliveira, L.A.T.; Dutra, V.d.F.; Bub, C.B.; Sakashita, A.M.; Yokoyama, A.P.H.; et al. A Common Variant Close to the “Tripwire” Linker Region of NLRP1 Contributes to Severe COVID-19. Inflamm. Res. 2022; online ahead of print. [Google Scholar] [CrossRef]
- Schröder, N.W.; Schumann, R.R. Single Nucleotide Polymorphisms of Toll-like Receptors and Susceptibility to Infectious Disease. Lancet Infect. Dis. 2005, 5, 156–164. [Google Scholar] [CrossRef]
- Choudhury, A.; Mukherjee, S. In Silico Studies on the Comparative Characterization of the Interactions of SARS-CoV-2 Spike Glycoprotein with ACE-2 Receptor Homologs and Human TLRs. J. Med. Virol. 2020, 92, 2105–2113. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Kuang, M.; Li, J.; Zhu, L.; Jia, Z.; Guo, X.; Hu, Y.; Kong, J.; Yin, H.; Wang, X.; et al. Publisher Correction: SARS-CoV-2 Spike Protein Interacts with and Activates TLR4. Cell Res. 2021, 31, 825. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.; Karki, R.; Williams, E.P.; Yang, D.; Fitzpatrick, E.; Vogel, P.; Jonsson, C.B.; Kanneganti, T.-D. TLR2 Senses the SARS-CoV-2 Envelope Protein to Produce Inflammatory Cytokines. Nat. Immunol. 2021, 22, 829–838. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, S.; Oliviero, B.; Varchetta, S.; Renieri, A.; Mondelli, M.U. TLRs: Innate Immune Sentries Against SARS-CoV-2 Infection. Int. J. Mol. Sci. 2023, 24, 8065. [Google Scholar] [CrossRef]
- Alhabibi, A.M.; Hassan, A.S.; Abd Elbaky, N.M.; Eid, H.A.; Khalifa, M.A.A.A.; Wahab, M.A.; Althoqapy, A.A.; Abdou, A.E.; Zakaria, D.M.; Nassef, E.M.; et al. Impact of Toll-Like Receptor 2 and 9 Gene Polymorphisms on COVID-19: Susceptibility, Severity, and Thrombosis. J. Inflamm. Res. 2023, 16, 665–675. [Google Scholar] [CrossRef]
- Taha, S.I.; Shata, A.K.; Baioumy, S.A.; Fouad, S.H.; Anis, S.G.; Mossad, I.M.; Moustafa, N.M.; Abdou, D.M.; Youssef, M.K. Toll-Like Receptor 4 Polymorphisms (896A/G and 1196C/T) as an Indicator of COVID-19 Severity in a Convenience Sample of Egyptian Patients. J. Inflamm. Res. 2021, 14, 6293–6303. [Google Scholar] [CrossRef]
- Razmara, M.; Srinivasula, S.M.; Wang, L.; Poyet, J.-L.; Geddes, B.J.; DiStefano, P.S.; Bertin, J.; Alnemri, E.S. CARD-8 Protein, a New CARD Family Member That Regulates Caspase-1 Activation and Apoptosis. J. Biol. Chem. 2002, 277, 13952–13958. [Google Scholar] [CrossRef]
- de Sá, N.B.R.; Neira-Goulart, M.; Ribeiro-Alves, M.; Perazzo, H.; Geraldo, K.M.; Ribeiro, M.P.D.; Cardoso, S.W.; Grinsztejn, B.; Veloso, V.G.; Capão, A.; et al. Inflammasome Genetic Variants Are Associated with Protection to Clinical Severity of COVID-19 among Patients from Rio de Janeiro, Brazil. Biomed. Res. Int. 2022, 2022, 9082455. [Google Scholar] [CrossRef]
- Di Gioia, M.; Zanoni, I. Toll-like Receptor Co-Receptors as Master Regulators of the Immune Response. Mol. Immunol. 2015, 63, 143–152. [Google Scholar] [CrossRef]
- Pati, A.; Padhi, S.; Panda, D.; Suvankar, S.; Panda, A.K. A Cluster of Differentiation 14 (CD14) Polymorphism (C-159T Rs2569190) Is Associated with SARS-CoV-2 Infection and Mortality in the European Population. J. Infect. Dis. 2021, 224, 921–922. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Wang, X.; Saredy, J.; Yuan, Z.; Yang, X.; Wang, H. Innate-Adaptive Immunity Interplay and Redox Regulation in Immune Response. Redox Biol. 2020, 37, 101759. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.-L.; Hu, J.; Xiao, X.-F.; Peng, Y.; Zhao, S.-P.; Xiao, X.-Z.; Yang, M.-S. The CD40 Rs1883832 Polymorphism Affects Sepsis Susceptibility and SCD40L Levels. Biom. Res. Int. 2018, 2018, 7497314. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Total | Asymptomatic Disease | Mild Disease | Moderate Disease | Severe Disease | Critical Disease | p Value # | |
|---|---|---|---|---|---|---|---|
| No | 249 | 58 | 88 | 26 | 34 | 43 | - |
| Sex (male/female) | 172/77 | 40/18 | 60/28 | 16/10 | 27/7 | 29/14 | 0.648 (C) |
| Age | 42.8 ± 18.5 | 35.3 ± 14.3 | 31.3 ± 11.6 | 48.2 ± 17.3 | 58.1 ± 14.8 | 60.8 ± 14.6 | <0.001 (K−W) |
| Comorbidities (n, %) | 89, 35.7 | 6, 10.3 | 9, 10.2 | 15, 57.7 | 23, 67.6 | 36, 83.7 | <0.001 (C) |
| Obesity (n, %) | 28, 11.2 | 0, 0.0 | 2, 2.3 | 4, 15.4 | 4, 11.8 | 18, 41.9 | <0.001 (C) |
| Hypertension (n, %) | 45, 18.1 | 1, 1.7 | 2, 2.3 | 5, 19.2 | 12, 35.3 | 25, 58.1 | <0.001 (C) |
| Dyslipidemia (n, %) | 17, 6.8 | 1, 1.7 | 0, 0.0 | 5, 19.2 | 4, 11.8 | 7, 16.3 | <0.001 (C) |
| Diabetes mellitus (n, %) | 20, 8.0 | 0, 0.0 | 2, 2.3 | 1, 3.8 | 8, 23.5 | 9, 20.9 | <0.001 (C) |
| Thyroid disease (n, %) | 10, 4.0 | 0, 0.0 | 3, 3.4 | 3, 11.5 | 1, 2.9 | 3, 7.0 | 0.116 (C) |
| CHD (n, %) | 11, 4.4 | 0, 0.0 | 0, 0.0 | 1, 3.8 | 4, 11.8 | 6, 14.0 | <0.001 (C) |
| CVD (n, %) | 8, 3.2 | 0, 0.0 | 0, 0.0 | 1, 3.8 | 0, 0.0 | 7, 16.3 | <0.001 (C) |
| CRD (n, %) | 18, 7.2 | 1, 1.7 | 1, 1.1 | 0, 0.0 | 6, 17.6 | 10, 23.3 | <0.001 (C) |
| Chronic liver and/or renal disease (n, %) | 6, 2.4 | 1, 1.7 | 0, 0.0 | 2, 7.7 | 1, 2.9 | 2, 4.7 | 0.176 (C) |
| Malignancies (n, %) | 6, 2.4 | 0, 0.0 | 2, 2.3 | 0, 0.0 | 1, 2.9 | 3, 7.0 | 0.204 (C) |
| Chronic immune and/or hematologic diseases (n, %) ^ | 6, 2.4 | 1, 1.7 | 0, 0.0 | 0, 0.0 | 3, 8.8 | 2, 4.7 | 0.044 (C) |
| Comorbidity-others (n, %) * | 15, 6.0 | 3, 5.2 | 0, 0.0 | 1, 3.8 | 5, 14.7 | 6, 14.0 | 0.004 (C) |
| Gene | SNP | Primer Sequences | PCR Conditions | PCR Product (bp) | Digestion Enzyme | Expected RFLP Fragments (bp) * |
|---|---|---|---|---|---|---|
| TLR2 | rs5743708 | F: 5′-TATGGTCCAGGAGCTGGAGA-3′ | 94 °C 2 min, 38 cycles (94 °C 30 s, 64 °C 30 s, 72 °C 45 s), 72 °C 5 min | 430 | SfcI | 430 (wt/GG), 430 + 307 + 123 (heter/GA), 307 + 123 (homo/AA) |
| R: 5′-TGACATAAAGATCCCAACTAGACAA-3′ | ||||||
| TLR4 | rs4986790 | F: 5′-GATTAGCATACTTAGACTACTACCTCCATG-3′ | 94 °C 2 min, 32 cycles (94 °C 30 s, 57 °C 30 s, 72 °C 60 s), 72 °C 5 min | 249 | NcoI | 249 (wt/AA), 249 + 218 + 31 (heter/AG), 218 + 31 (homo/GG) |
| R: 5′-GATCAACTTCTGAAAAAGCATTCCCAC-3′ | ||||||
| rs4986791 | F: 5′-GGTTGCTGTTCTCAAAGTGATTTTGGGAGAA-3′ | 94 °C 2 min, 32 cycles (94 °C 30 s, 57 °C 30 s, 72 °C 60 s), 72 °C 5 min | 404 | HinfI | 404 (wt/CC), 404 + 375 + 29 (heter/CT), 375 + 29 (homo/TT) | |
| R: 5′-ACCTGAAGACTGGAGAGTGAGTTAAATGC-3′ | ||||||
| CD14 | rs2569190 | F: 5′-GCCAACAGATGAGGTTCACA-3′ | 94 °C 2 min, 30 cycles (94 °C 30 s, 60 °C 30 s, 72 °C 30 s), 72 °C 1 min | 641 | HaeIII | 308 + 174 (wt/CC), 522 + 308 + 174 (heter/CG), 522 (homo/GG) |
| R: 5′-GTTCGACCCCAAGACCCTAC-3′ | ||||||
| CARD8 | rs2043211 | F: 5′-AAGAGCAGCATATAGTGGGAAA-3′ | 94 °C 2 min, 30 cycles (94 °C 30 s, 57 °C 30 s, 72 °C 30 s), 72 °C 1 min | 475 | MboII | 151 + 324 (wt/AA), 475 + 324 + 151 (heter/AT), 475 (homo/TT) |
| R: 5′-TGAGTTCGATGAAAAACACCC-3′ | ||||||
| IL18 | rs1834481 | F: 5′-TTCTGTTGTAGGGGGAGCGT-3′ | 94 °C 2 min, 30 cycles (94 °C 30 s, 60 °C 30 s, 72 °C 30 s), 72 °C 1 min | 522 | HaeIII | 308 + 174 (wt/CC), 522 + 308 + 174 (heter/CG), 522 (homo/GG) |
| R: 5′-CATCAGGTTGCTTCCAGCCA-3′ | ||||||
| CD40 | rs1883832 | F: 5′-ATAGGTGGACCGCGATTGGT-3′ | 95 °C 10 min, 32 cycles (94 °C 30 s, 58 °C 30 s, 72 °C 30 s), 72 °C 10 min | 216 | NcoI | 126 + 90 (wt/CC), 216 + 126 + 90 (heter/CT), 216 (homo/TT) |
| R: 5′-TCCCAACTCCCGTCTGGT-3′ |
| Gene | SNP | Genotype | Allele Frequency (n, %) | HWE | |||
|---|---|---|---|---|---|---|---|
| WT (n, %) | Heter (n, %) | Homo (n, %) | |||||
| TLR2 | rs5743708 | 239 (96.0) | 10 (4.0) | 0 (0.0) | G | A | 0.75 |
| (g.25877G>A) | (488, 98.0) | (10, 2.0) | |||||
| TLR4 | rs4986790 | 229 (92.0) | 19 (7.6) | 1 (0.4) | A | G | 0.38 |
| (g.13622A>G) | (477, 95.8) | (21, 4.2) | |||||
| rs4986791 | 230 (92.4) | 18 (7.2) | 1 (0.4) | C | T | 0.33 | |
| (g.13922C>T) | (478, 96.0) | (20, 4.0) | |||||
| CD14 | rs2569190 | 61 (24.5) | 123 (49.4) | 65 (26.1) | T | C | 0.85 |
| (g.5371T>C) | (245, 49.2) | (253, 50.8) | |||||
| IL18 | rs1834481 | 187 (75.1) | 54 (21.7) | 8 (31.2) | C | G | 0.11 |
| (g.16014C>G) | (428, 85.9) | (70, 14.1) | |||||
| CARD8 | rs2043211 | 99 (37.8) | 119 (47.8) | 31 (12.4) | A | T | 0.60 |
| (g.26821A>T) | (317, 63.7) | (181, 36.3) | |||||
| CD40 | rs1883832 | 108 (43.4) | 119 (47.8) | 22 (8.8) | C | T | 0.18 |
| (g.5077C>T) | (335, 67.3) | (163, 32.7) | |||||
| Gene | SNP | Genotype (N, %) ^ | Pneumonia (Yes vs. No) | Respiratory Distress (Yes vs. No) | Disease Severity (According to the WHO Classification) * | |||
|---|---|---|---|---|---|---|---|---|
| p | OR (95% CI) | p | OR (95% CI) | p | OR (95% CI) | |||
| TLR2 | rs5743708 | WT (239, 96.0) Heter + Homo (10, 4.0) | 0.061 | 3.73 (0.94–14.78) | 0.255 | 2.13 (0.58–7.81) | 0.103 | 2.57 (0.83–8.03) |
| TLR4 | rs4986790 | WT (229, 92.0) Heter + Homo (20, 8.0) | 0.329 | 1.75 (0.63–3.93) | 0.789 | 0.75 (0.24–2.35) | 0.305 | 1.53 (0.68–3.47) |
| rs4986791 | WT (230, 92.4) Heter + Homo (19, 7.6) | 0.093 | 2.22 (0.86–5.73) | 0.745 | 1.11 (0.38–3.22) | 0.058 | 2.14 (0.92–4.94) | |
| CD14 | rs2569190 | WT (65, 26.1) Heter + Homo (184, 73.9) | 0.804 | 1.08 (0.60–1.92) | 0.486 | 0.80 (0.42–1.51) | 0.416 | 0.81 (0.49–1.34) |
| IL18 | rs1834481 | WT (187, 75.1) Heter + Homo (62, 24.9) | 0.012 | 2.09 (1.17–3.73) | 0.101 | 1.70 (0.90–3.20) | 0.008 | 2.01 (1.20–3.38) |
| CARD8 | rs2043211 | WT (99, 39.8) Heter + Homo (150, 60.2) | 0.220 | 0.72 (0.43–1.21) | 0.084 | 0.72 (0.43–1.21) | 0.031 | 0.60 (0.38–0.96) |
| CD40 | rs1883832 | WT (108, 43.4) Heter + Homo (141, 56.6) | 0.665 | 1.14 (0.63–2.05) | 0.442 | 1.22 (0.73–2.05) | 0.368 | 1.23 (0.78–1.93) |
| TLR2 or/and TLR4 | rs5743708 or/and rs4986791 | WT (223, 89.6) Heter + Homo(26, 10.4 #) | 0.020 | 2.70 (1.17–6.22) | 0.432 | 1.43 (0.59–3.47) | 0.022 | 2.35 (1.13–4.88) |
| Parameter | Pneumonia | Respiratory Distress | Disease Severity (According to the WHO Classification) * | |||
|---|---|---|---|---|---|---|
| p | OR (95% CI) | p | OR (95% CI) | p | OR (95% CI) | |
| Age | <0.001 | 1.10 (1.06–1.13) | <0.001 | 1.06 (1.02–1.09) | <0.001 | 1.04 (1.02–1.06) |
| Sex (female/male) | 0.723 | 0.84 (0.31–2.24) | 0.118 | 0.42 (0.14–1.24) | 0.481 | 0.82 (0.47–1.43) |
| Obesity | 0.001 | 20.35 (3.46–119.68) | 0.002 | 7.38 (2.13–25.61) | <0.001 | 8.18 (3.08–21.73) |
| Hypertension | 0.909 | 0.91 (0.19–4.34) | 0.085 | 2.89 (0.86–9.65) | 0.021 | 3.05 (1.18–7.87) |
| Diabetes mellitus | 0.029 | 12.15 (1.30–113.81) | 0.013 | 6.59 (1.48–29.36) | 0.039 | 3.32 (1.06–10.38) |
| Dyslipidemia | 0.195 | 4.46 (0.46–42.86) | 0.813 | 1.21 (0.25–5.82) | 0.801 | 1.16 (0.37–3.66) |
| Thyroid disease | 0.323 | 2.85 (0.36–22.79) | 0.489 | 2.05 (0.27–15.70) | 0.220 | 2.29 (0.61–8.59) |
| CHD | 0.999 | - | 0.305 | 0.37 (0.05–2.49) | 0.801 | 0.82 (0.17–4.01) |
| CVD | 0.999 | - | 0.172 | 5.61 (0.47–66.54) | 0.055 | 9.26 (0.96–89.84) |
| CRD | 0.775 | 1.31 (0.20–8.56) | 0.307 | 2.05 (0.52–8.09) | 0.058 | 3.01 (0.96–9.39) |
| Chronic liver and/or renal disease | 0.754 | 1.54 (0.11–22.85) | 0.749 | 0.67 (0.06–7.58) | 0.917 | 0.91 (0.17–4.98) |
| Chronic immune and/or hematologic diseases | 0.002 | 57.89 (4.30–780.17) | 0.216 | 4.63 (0.41–52.35) | 0.004 | 11.92 (2.19–64.82) |
| Malignancies | 0.663 | 0.59 (0.06–6.28) | 0.199 | 4.72 (0.44–50.53) | 0.448 | 2.00 (0.33–12.05) |
| Comorbidity-others ^ | 0.897 | 1.13 (0.18–6.28) | 0.892 | 1.12 (0.23–5.53) | 0.466 | 1.55 (0.48–4.99) |
| TLR2-rs5743708 and/or TLR4-rs4986791 | 0.033 | 3.62 (1.11–11.83) | 0.897 | 1.09 (0.31–3.83) | 0.026 | 2.54 (1.12–5.79) |
| IL18-rs1834481 | 0.026 | 2.88 (1.13–7.32) | 0.607 | 1.28 (0.50–3.26) | 0.073 | 1.69 (0.95–2.99) |
| CARD8-rs2043211 | 0.751 | 1.15 (0.49–2.72) | 0.210 | 0.57 (0.24–1.37) | 0.204 | 0.72 (0.44–1.19) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bakaros, E.; Voulgaridi, I.; Paliatsa, V.; Gatselis, N.; Germanidis, G.; Asvestopoulou, E.; Alexiou, S.; Botsfari, E.; Lygoura, V.; Tsachouridou, O.; et al. Innate Immune Gene Polymorphisms and COVID-19 Prognosis. Viruses 2023, 15, 1784. https://doi.org/10.3390/v15091784
Bakaros E, Voulgaridi I, Paliatsa V, Gatselis N, Germanidis G, Asvestopoulou E, Alexiou S, Botsfari E, Lygoura V, Tsachouridou O, et al. Innate Immune Gene Polymorphisms and COVID-19 Prognosis. Viruses. 2023; 15(9):1784. https://doi.org/10.3390/v15091784
Chicago/Turabian StyleBakaros, Evangelos, Ioanna Voulgaridi, Vassiliki Paliatsa, Nikolaos Gatselis, Georgios Germanidis, Evangelia Asvestopoulou, Stamatia Alexiou, Elli Botsfari, Vasiliki Lygoura, Olga Tsachouridou, and et al. 2023. "Innate Immune Gene Polymorphisms and COVID-19 Prognosis" Viruses 15, no. 9: 1784. https://doi.org/10.3390/v15091784