
DNA Vaccine Co-Expressing Hemagglutinin and IFN-γ Provides Partial Protection to Ferrets against Lethal Challenge with Canine Distemper Virus
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Viruses, Cells, and Ferrets
2.2. Preparation of DNA Vaccines
2.3. Identification of DNA Vaccine Plasmids via Indirect Immunofluorescence Assay (IFA)
2.4. Administration of DNA Vaccines and Sample Collection
2.5. Assessment of Clinical Symptoms
2.6. Virus Neutralization and Cytokine Detection
2.7. Real-Time RT-PCR Detected CDV RNA
2.8. Histology and Immunohistochemistry
2.9. Statistical Analysis
3. Results
3.1. Codon Optimization Enhances the Expression of H and N Proteins
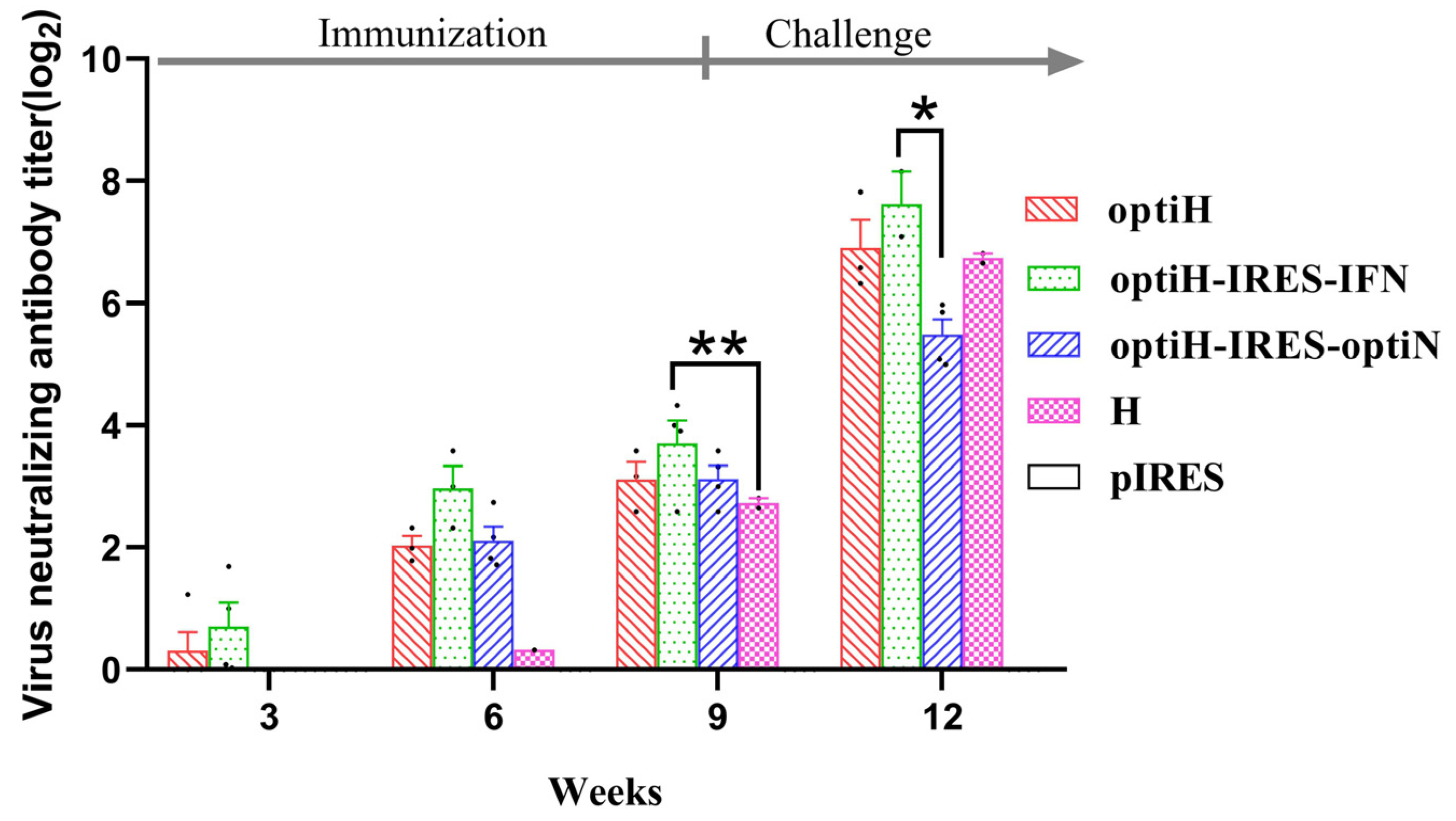
3.2. Virus-Neutralizing Antibody Responses after DNA Vaccine Immunization
3.3. Cytokine Responses in Sera after Immunization with DNA Vaccines
3.4. Vaccine Immunization Alleviates Clinical Symptoms in Infected Ferrets
3.5. Vaccine Immunization Reduces CDV RNA Loads in Infected Ferrets
3.6. Pathological Changes and Immunohistochemistry of Ferrets after CDV Challenge
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward/Reverse Primer/Probe |
|---|---|
| SD(14)7-H | 5′-CGGCTAGCgccgccaccATGCTCTCCTAC -3′ |
| 5′-CCGCTCGAGTCAAGGTTTTGAACGGTTAC-3′ | |
| SD(14)7-N | 5′- GCGTCGACgccgccaccATGGCTAGCCTT-3′ |
| 5′-ATTTTGCGGCCGCTTAATTGAGTAGCTCT-3′ | |
| Ferret IFN-γ | 5′-GCGTCGACgccgccaccATGAATTATACA-3′ |
| 5′-ATTTGCGGCCGCTTATTTCGATGCTCTGCG-3′ |
References
- Kapil, S.; Yeary, T.J. Canine Distemper Spillover in Domestic Dogs from Urban Wildlife. Vet. Clin. N. Am. Small Anim. Pract. 2011, 41, 1069–1086. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Wang, S.; Li, P.; Yue, F.; Zhang, Y.; Pan, B.; Liu, X. Apoptotic investigation of brain tissue cells in dogs naturally infected by canine distemper virus. Virol. J. 2021, 18, 165. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Li, A.; Ye, H.; Shi, Y.; Hu, Z.; Zeng, L. Natural infection with canine distemper virus in hand-feeding Rhesus monkeys in China. Vet. Microbiol. 2010, 141, 374–378. [Google Scholar] [CrossRef] [PubMed]
- Almberg, E.S.; Cross, P.C.; Smith, D.W. Persistence of canine distemper virus in the Greater Yellowstone ecosystem’s carnivore community. Ecol. Appl. 2010, 20, 2058–2074. [Google Scholar] [CrossRef]
- Riley, M.C.; Wilkes, R.P. Sequencing of emerging canine distemper virus strain reveals new distinct genetic lineage in the United States associated with disease in wildlife and domestic canine populations. Virol. J. 2015, 12, 219. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.C.; Chen, C.A.; Chen, C.M.; Yen, C.H.; Lee, M.H.; Chuang, C.K.; Tu, C.F.; Su, B.L. Application of xenogeneic anti-canine distemper virus antibodies in treatment of canine distemper puppies. J. Small Anim. Pract. 2016, 57, 626–630. [Google Scholar] [CrossRef]
- Martella, V.; Blixenkrone-Møller, M.; Elia, G.; Lucente, M.S.; Cirone, F.; Decaro, N.; Nielsen, L.; Bányai, K.; Carmichael, L.E.; Buonavoglia, C. Lights and shades on an historical vaccine canine distemper virus, the Rockborn strain. Vaccine 2011, 29, 1222–1227. [Google Scholar] [CrossRef]
- Cherpillod, P.; Tipold, A.; Griot-Wenk, M.; Cardozo, C.; Schmid, I.; Fatzer, R.; Schobesberger, M.; Zurbriggen, R.; Bruckner, L.; Roch, F.; et al. DNA vaccine encoding nucleocapsid and surface proteins of wild type canine distemper virus protects its natural host against distemper. Vaccine 2000, 18, 2927–2936. [Google Scholar] [CrossRef]
- Jensen, T.H.; Nielsen, L.; Aasted, B.; Blixenkrone-Møller, M. Early life DNA vaccination with the H gene of Canine distemper virus induces robust protection against distemper. Vaccine 2009, 27, 5178–5183. [Google Scholar] [CrossRef]
- Jensen, T.H.; Nielsen, L.; Aasted, B.; Pertoldi, C.; Blixenkrone-Møller, M. Canine distemper virus DNA vaccination of mink can overcome interference by maternal antibodies. Vaccine 2015, 33, 1375–1381. [Google Scholar] [CrossRef]
- Martínez, O.; Bravo Cruz, A.; Santos, S.; Ramírez, M.; Miranda, E.; Shisler, J.; Otero, M. Vaccination with a codon-optimized A27L-containing plasmid decreases virus replication and dissemination after vaccinia virus challenge. Vaccine 2017, 35, 6007–6014. [Google Scholar] [PubMed]
- Grote, A.; Hiller, K.; Scheer, M.; Münch, R.; Nörtemann, B.; Hempel, D.C.; Jahn, D. JCat: A novel tool to adapt codon usage of a target gene to its potential expression host. Nucleic Acids Res. 2005, 33, 526–531. [Google Scholar] [CrossRef] [PubMed]
- Osman, R.; Gonzalez-Cano, P.; Brownlie, R.; Griebel, P.J. Induction of interferon and interferon-induced antiviral effector genes following a primary bovine herpesvirus-1 (BHV-1) respiratory infection. J. Gen. Virol. 2017, 98, 1831–1842. [Google Scholar] [PubMed]
- Toporovski, R.; Morrow, M.P.; Weiner, D.B. Interferons as potential adjuvants in prophylactic vaccines. Expert. Opin. Biol. Ther. 2010, 10, 1489–1500. [Google Scholar]
- von Messling, V.; Springfeld, C.; Devaux, P.; Cattaneo, R. A ferret model of canine distemper virus virulence and immunosuppression. J. Virol. 2003, 77, 12579–12591. [Google Scholar]
- von Messling, V.; Milosevic, D.; Cattaneo, R. Tropism illuminated: Lymphocyte-based pathways blazed by lethal morbillivirus through the host immune system. Proc. Natl. Acad. Sci. USA 2004, 101, 14216–14221. [Google Scholar]
- Pan, H.; Liu, M.; Tao, R.; Li, X.; Gong, C.; Chen, J.; Zhao, J. Pathogenicity of canine distemper virus SD(14)7 strain on beagle dogs. Vet. Sci. China 2020, 50, 321–329. [Google Scholar]
- Zhang, H.; Zhao, J.; Bai, X.; Zhang, L.; Fan, S.; Hu, B.; Liu, H.; Zhang, D.; Xu, S.; Yan, X. Cloning and expression of mink (Neovison vison) interferon-γ gene and development of an antiviral assay. Res. Vet. Sci. 2015, 101, 93–98. [Google Scholar]
- Zhang, H.; Zhang, S.; Fan, S.; Zhang, L.; Hu, B.; Bai, X.; Zhang, D.; Lu, R.; Zhao, J.; Lian, S.; et al. Identification and primary application of hybridomas cell secreting monoclonal antibodies against mink (Neovison vison) interferon-gamma. Cytokine 2022, 150, 155777. [Google Scholar]
- da Fontoura Budaszewski, R.; Hudacek, A.; Sawatsky, B.; Krämer, B.; Yin, X.; Schnell, M.J.; von Messling, V. Inactivated Recombinant Rabies Viruses Displaying Canine Distemper Virus Glycoproteins Induce Protective Immunity against Both Pathogens. J. Virol. 2017, 91, e02077-16. [Google Scholar]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoint. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar]
- Sakai, K.; Nagata, N.; Ami, Y.; Seki, F.; Suzaki, Y.; Iwata-Yoshikawa, N.; Suzuki, T.; Fukushi, S.; Mizutani, T.; Yoshikawa, T.; et al. Lethal canine distemper virus outbreak in cynomolgus monkeys in Japan in 2008. J. Virol. 2013, 87, 1105–1114. [Google Scholar]
- Beineke, A.; Puff, C.; Seehusen, F.; Baumgärtner, W. Pathogenesis and immunopathology of systemic and nervous canine distemper. Vet Immunol. Immunopathol. 2009, 127, 1–18. [Google Scholar]
- Mizuguchi, H.; Xu, Z.; Ishii-Watabe, A.; Uchida, E.; Hayakawa, T. IRES-dependent second gene expression is significantly lower than cap-dependent first gene expression in a bicistronic vector. Mol. Ther. 2000, 1, 376–382. [Google Scholar] [PubMed]
- Li, W.; Li, T.; Liu, Y.; Gao, Y.; Yang, S.; Feng, N.; Sun, H.; Wang, S.; Wang, L.; Bu, Z.; et al. Genetic characterization of an isolate of canine distemper virus from a Tibetan Mastiff in China. Virus Genes 2014, 49, 45–57. [Google Scholar] [PubMed]
- Iwatsuki, K.; Miyashita, N.; Yoshida, E.; Gemma, T.; Shin, Y.S.; Mori, T.; Hirayama, N.; Kai, C.; Mikami, T. Molecular and phylo-genetic analyses of the haemagglutinin (H) proteins of field isolates of canine distemper virus from naturally infected dogs. J. Gen. Virol. 1997, 78, 373–380. [Google Scholar] [PubMed]
- Nielsen, L.; Søgaard, M.; Karlskov-Mortensen, P.; Jensen, T.H.; Jensen, T.D.; Aasted, B.; Blixenkrone-Møller, M. Humoral and cell-mediated immune responses in DNA immunized mink challenged with wild-type canine distemper virus. Vaccine 2009, 27, 4791–4797. [Google Scholar]
- Shirai, S.; Shibuya, M.; Kawai, A.; Tamiya, S.; Munakata, L.; Omata, D.; Suzuki, R.; Aoshi, T.; Yoshioka, Y. Lipid Nanoparticles Potentiate CpG-Oligodeoxynucleotide-Based Vaccine for Influenza Virus. Front. Immunol. 2019, 10, 3018. [Google Scholar]
- Tian, M.; Zhou, Z.; Tan, S.; Fan, X.; Li, L.; Ullah, N. Formulation in DDA-MPLA-TDB Liposome Enhances the Immunogenicity and Protective Efficacy of a DNA Vaccine against Mycobacterium tuberculosis Infection. Front. Immunol. 2018, 9, 310. [Google Scholar]
- Waghule, T.; Singhvi, G.; Dubey, S.K.; Pandey, M.M.; Gupta, G.; Singh, M.; Dua, K. Microneedles: A smart approach and increasing potential for transdermal drug delivery system. Biomed. Pharmacother. 2019, 109, 1249–1258. [Google Scholar]
- Tezel, A.; Paliwal, S.; Shen, Z.; Mitragotri, S. Low-frequency ultrasound as a transcutaneous immunization adjuvant. Vaccine 2005, 23, 3800–3807. [Google Scholar] [PubMed]
- Babiuk, S.; Tsang, C.; van Drunen Littel-van den Hurk, S.; Babiuk, L.A.; Griebel, P.J. A single HBsAg DNA vaccination in combination with electroporation elicits long-term antibody responses in sheep. Bioelectrochemistry 2007, 70, 269–274. [Google Scholar] [PubMed]







| Figure Number | DNA Vaccine | Vaccine Dose (µg) | Route |
|---|---|---|---|
| #1, #2, #3, #4 | optiH | 800/800/500 | i.d. + i.m. |
| #5, #6, #7, #8 | optiH-IRES-IFN | 800/800/500 | i.d. + i.m. |
| #9, #10, #11, #12 | optiH-IRES-optiN | 800/800/500 | i.d. + i.m. |
| #13, #14, #15, #16 | H | 800/800/500 | i.d. + i.m. |
| #17, #18, #19, #20 | pIRES | 800/800/500 | i.d. + i.m. |
| Symptom | Clinical Score | ||
|---|---|---|---|
| 2 | 1 | 0 | |
| Conjunctivitis | Severe discharge | Moderate secretion | Mild discharge |
| Fever | ≥39.2 °C and ≥3 days | ≥39.2 °C and ≥2 days | 37.8–39.2 °C |
| Weight | Loss > 10% | 5% < loss < 10% | Loss ≤ 5% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, J.; Sun, Y.; Sui, P.; Pan, H.; Shi, Y.; Chen, J.; Zhang, H.; Wang, X.; Tao, R.; Liu, M.; et al. DNA Vaccine Co-Expressing Hemagglutinin and IFN-γ Provides Partial Protection to Ferrets against Lethal Challenge with Canine Distemper Virus. Viruses 2023, 15, 1873. https://doi.org/10.3390/v15091873
Zhao J, Sun Y, Sui P, Pan H, Shi Y, Chen J, Zhang H, Wang X, Tao R, Liu M, et al. DNA Vaccine Co-Expressing Hemagglutinin and IFN-γ Provides Partial Protection to Ferrets against Lethal Challenge with Canine Distemper Virus. Viruses. 2023; 15(9):1873. https://doi.org/10.3390/v15091873
Chicago/Turabian StyleZhao, Jianjun, Yiyang Sun, Ping Sui, Hongjun Pan, Yijun Shi, Jie Chen, Hailing Zhang, Xiaolong Wang, Rongshan Tao, Mengjia Liu, and et al. 2023. "DNA Vaccine Co-Expressing Hemagglutinin and IFN-γ Provides Partial Protection to Ferrets against Lethal Challenge with Canine Distemper Virus" Viruses 15, no. 9: 1873. https://doi.org/10.3390/v15091873
APA StyleZhao, J., Sun, Y., Sui, P., Pan, H., Shi, Y., Chen, J., Zhang, H., Wang, X., Tao, R., Liu, M., Sun, D., & Zheng, J. (2023). DNA Vaccine Co-Expressing Hemagglutinin and IFN-γ Provides Partial Protection to Ferrets against Lethal Challenge with Canine Distemper Virus. Viruses, 15(9), 1873. https://doi.org/10.3390/v15091873