
Genetic and Biological Characteristics of Duck-Origin H4N6 Avian Influenza Virus Isolated in China in 2022
 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells
2.2. Virus Isolation
2.3. Genome Sequencing and Phylogenetic Analysis
2.4. Classification of Genotype
2.5. Receptor Binding Analysis
2.6. Virus Replication Kinetics In Vitro
2.7. Chicken Infection Studies
2.8. Chicken Transmission Experiments
2.9. Mouse Infection Studies
2.10. Statistical Analysis
3. Results
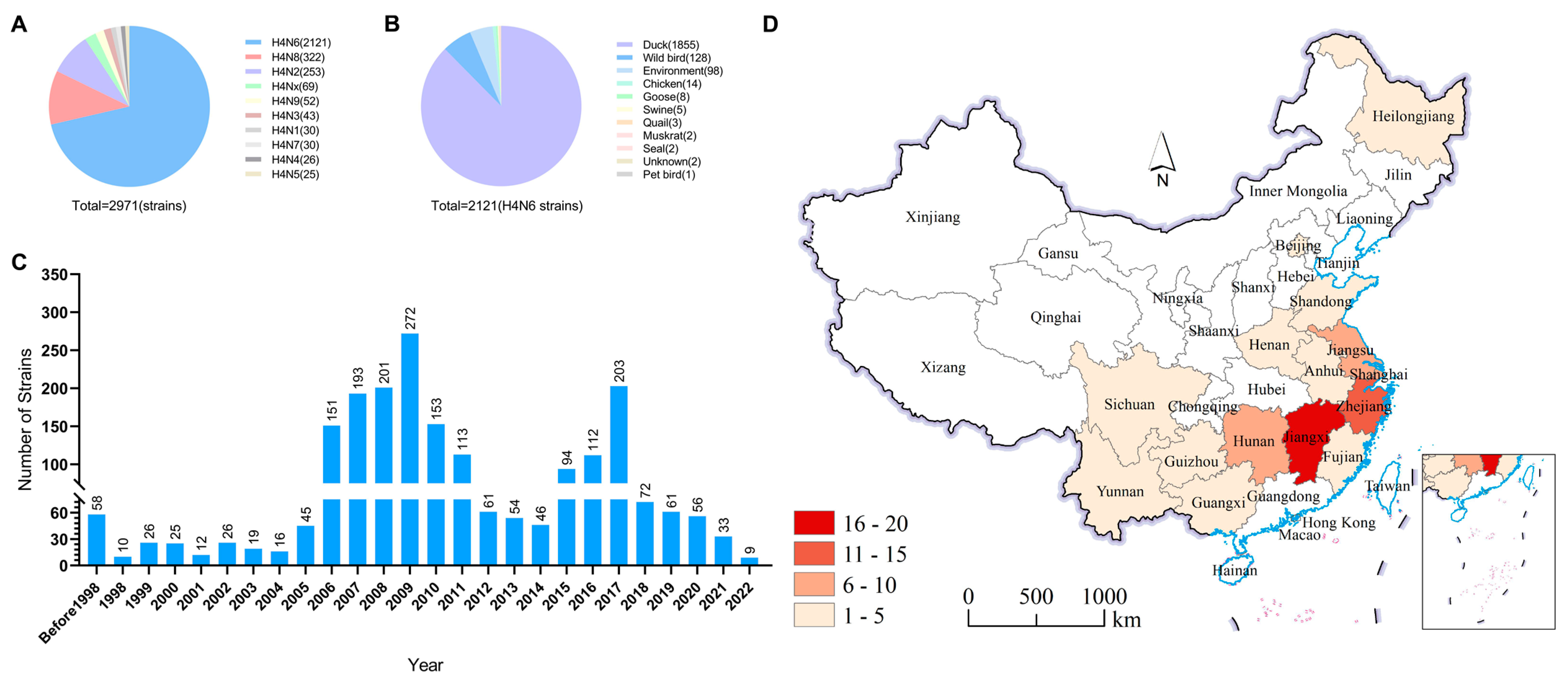
3.1. Prevalence of H4N6 AIVs in Wild Birds and Waterfowl
3.2. Isolation of H4N6 AIVs from Duck Farms around Poyang Lake
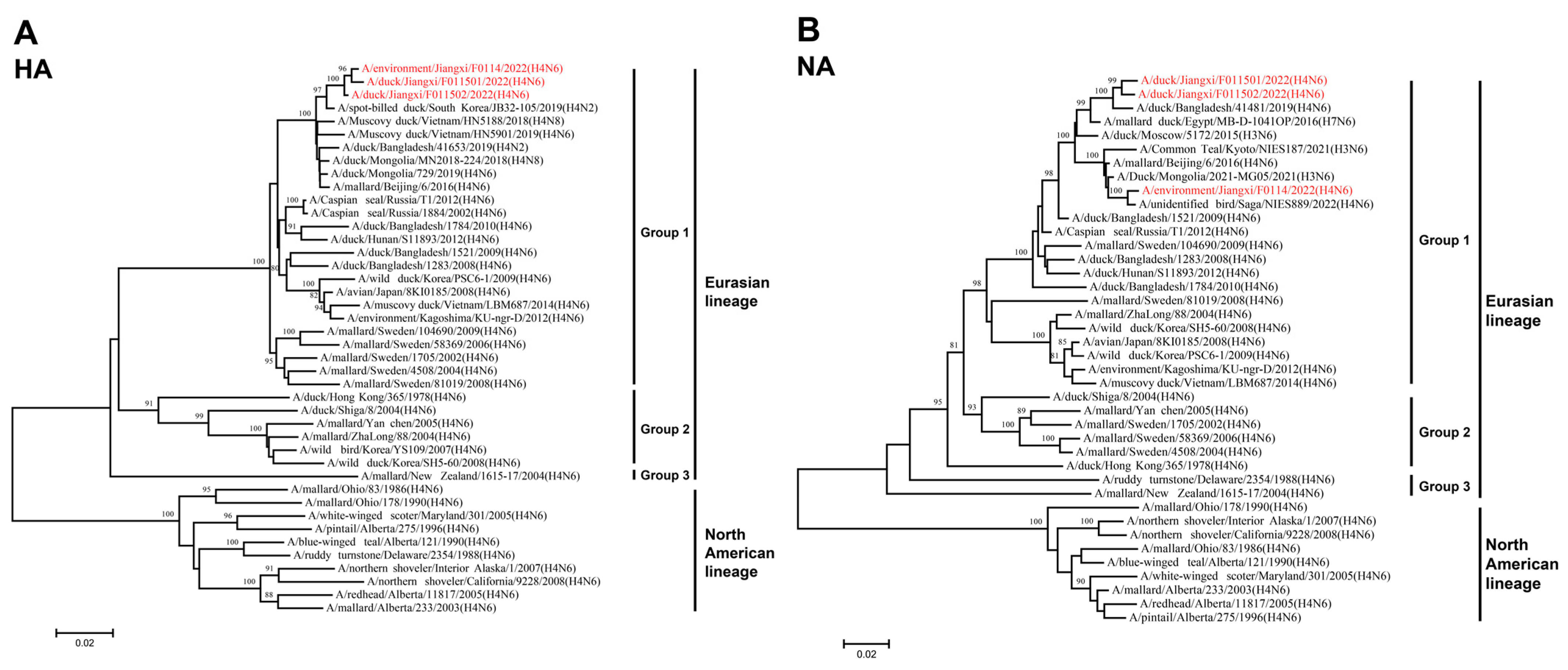
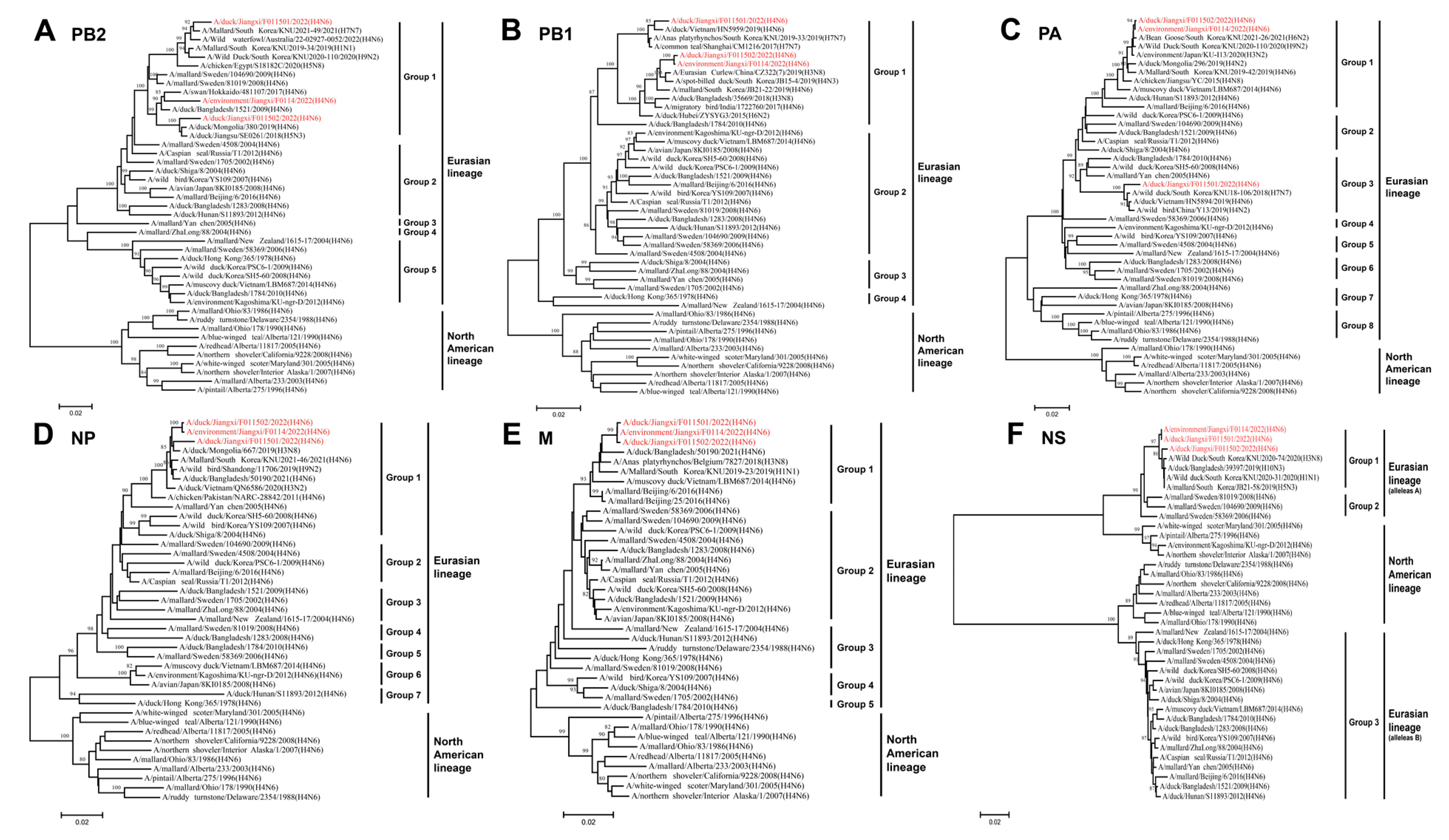
3.3. Phylogenetic Analysis of H4N6 AIVs
3.4. Molecular Characteristics of the Isolated H4N6 AIVs
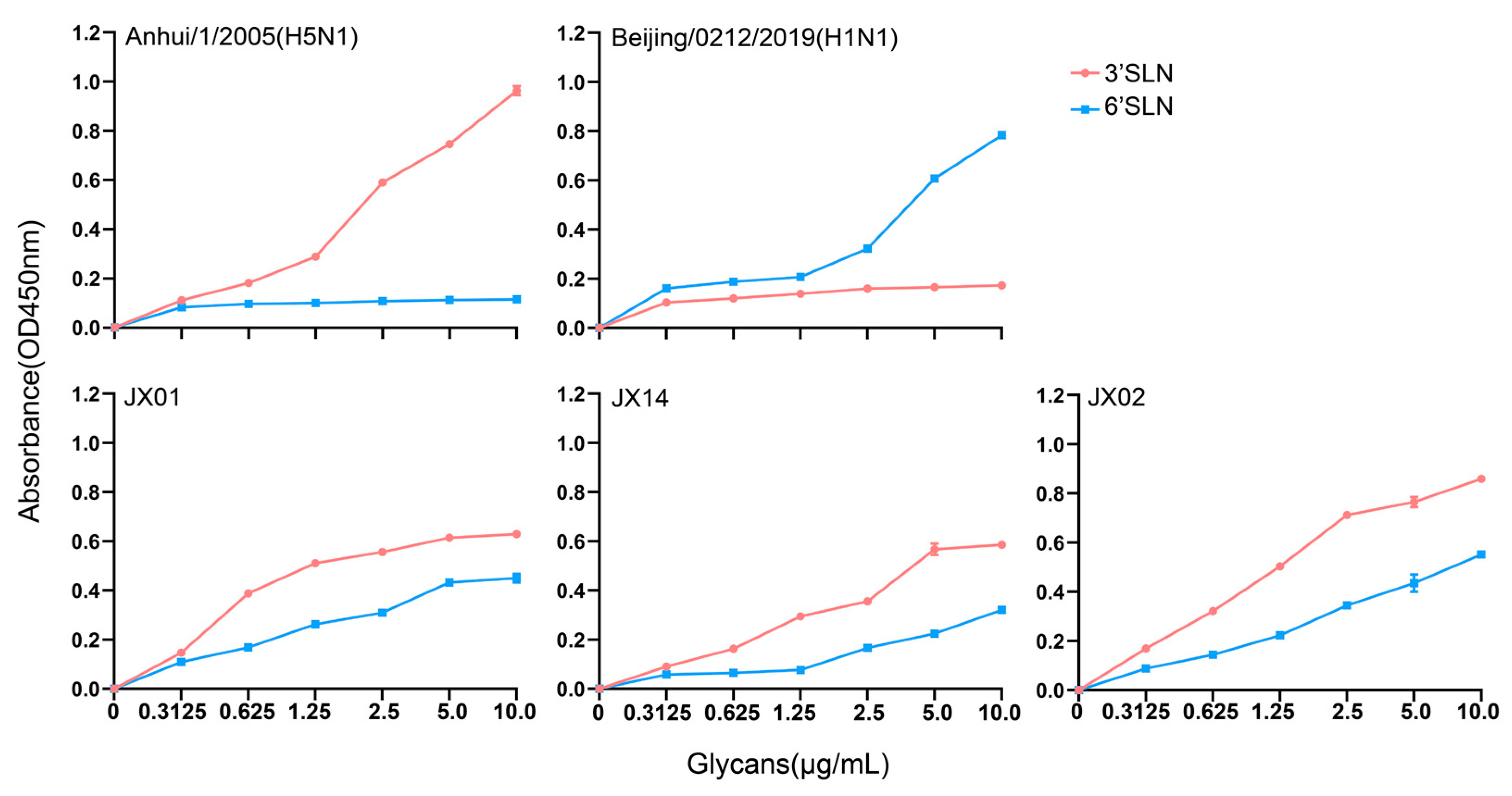
3.5. Receptor Binding Properties of the Isolated H4N6 AIVs
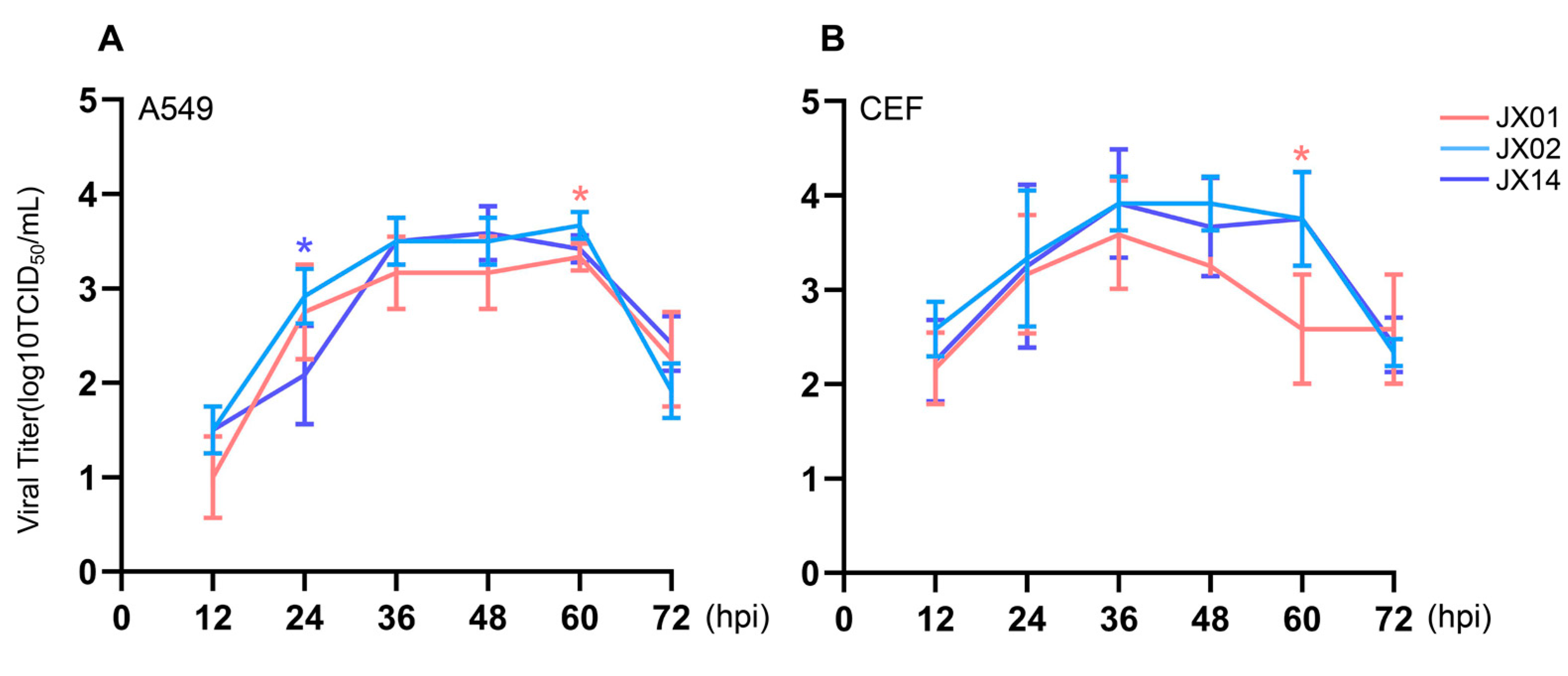
3.6. Replication of H4N6 AIVs in Mammalian Cells and Avian Cells
3.7. Replication and Pathogenicity of H4N6 AIV in Chickens
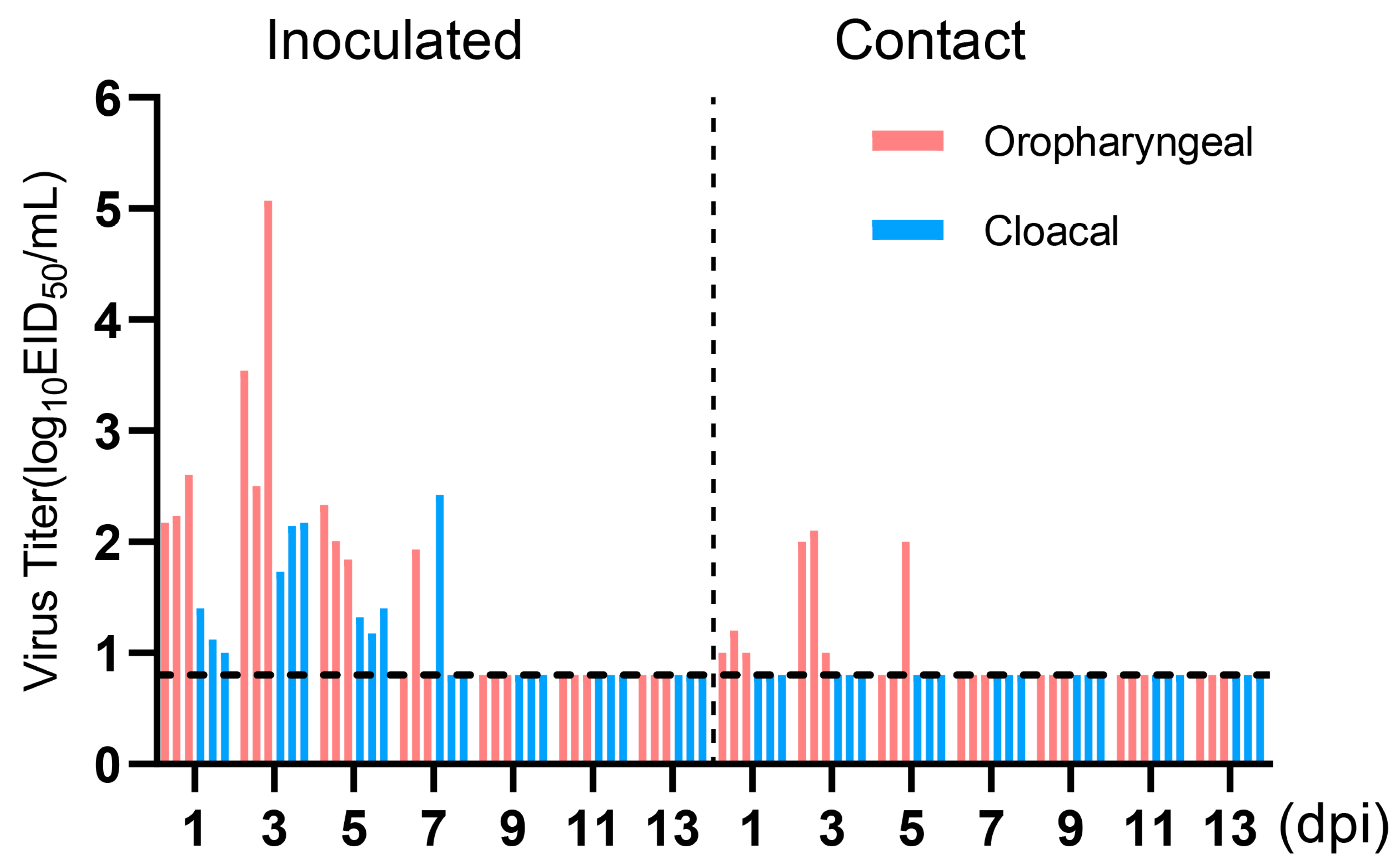
3.8. Transmission of H4N6 AIV in Chickens
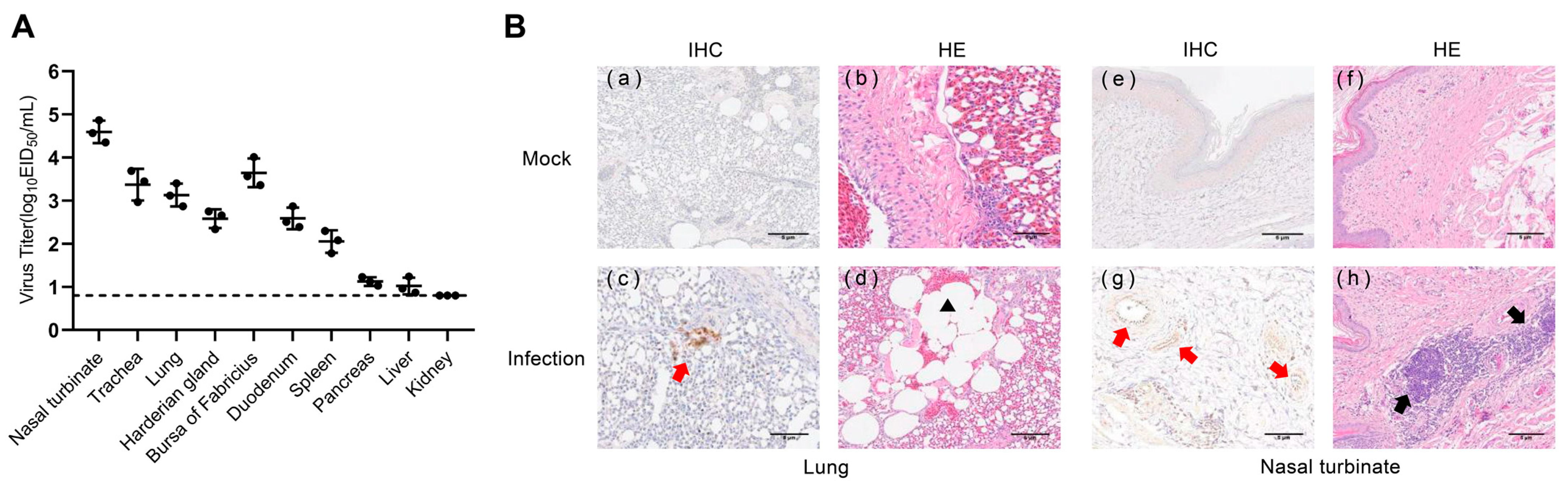
3.9. Replication and Pathogenicity of H4N6 AIV in Mice
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Olsen, B.; Munster, V.J.; Wallensten, A.; Waldenstrom, J.; Osterhaus, A.D.; Fouchier, R.A. Global patterns of influenza a virus in wild birds. Science 2006, 312, 384–388. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, M.; Zhang, H.; Zhao, C.; Zhang, Y.; Shen, J.; Sun, X.; Xu, H.; Xie, Y.; Gao, X.; et al. Prevalence, evolution, replication and transmission of H3N8 avian influenza viruses isolated from migratory birds in eastern China from 2017 to 2021. Emerg. Microbes Infect. 2023, 12, 2184178. [Google Scholar] [CrossRef]
- Gao, R.; Cao, B.; Hu, Y.; Feng, Z.; Wang, D.; Hu, W.; Chen, J.; Jie, Z.; Qiu, H.; Xu, K.; et al. Human infection with a novel avian-origin influenza A (H7N9) virus. N. Engl. J. Med. 2013, 368, 1888–1897. [Google Scholar] [CrossRef] [PubMed]
- Ke, C.; Mok, C.K.P.; Zhu, W.; Zhou, H.; He, J.; Guan, W.; Wu, J.; Song, W.; Wang, D.; Liu, J.; et al. Human Infection with Highly Pathogenic Avian Influenza A(H7N9) Virus, China. Emerg. Infect. Dis. 2017, 23, 1332–1340. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Sun, H.; Gao, F.; Luo, K.; Huang, Z.; Tong, Q.; Song, H.; Han, Q.; Liu, J.; Lan, Y.; et al. Human infection of avian influenza A H3N8 virus and the viral origins: A descriptive study. Lancet Microbe 2022, 3, e824–e834. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.J.; Wu, Y.; Bi, Y.; Shi, W.; Wang, D.; Shi, Y.; Gao, G.F. Emerging HxNy Influenza A Viruses. Cold Spring Harb. Perspect. Med. 2022, 12, a038406. [Google Scholar] [CrossRef]
- Kang, H.M.; Choi, J.G.; Kim, K.I.; Park, H.Y.; Park, C.K.; Lee, Y.J. Genetic and antigenic characteristics of H4 subtype avian influenza viruses in Korea and their pathogenicity in quails, domestic ducks and mice. J. Gen. Virol. 2013, 94, 30–39. [Google Scholar] [CrossRef]
- Liang, L.; Deng, G.; Shi, J.; Wang, S.; Zhang, Q.; Kong, H.; Gu, C.; Guan, Y.; Suzuki, Y.; Li, Y.; et al. Genetics, Receptor Binding, Replication, and Mammalian Transmission of H4 Avian Influenza Viruses Isolated from Live Poultry Markets in China. J. Virol. 2016, 90, 1455–1469. [Google Scholar] [CrossRef]
- Reid, S.M.; Brookes, S.M.; Núñez, A.; Banks, J.; Parker, C.D.; Ceeraz, V.; Russell, C.; Seekings, A.; Thomas, S.S.; Puranik, A.; et al. Detection of non-notifiable H4N6 avian influenza virus in poultry in Great Britain. Vet. Microbiol. 2018, 224, 107–115. [Google Scholar] [CrossRef]
- Abente, E.J.; Gauger, P.C.; Walia, R.R.; Rajao, D.S.; Zhang, J.; Harmon, K.M.; Killian, M.L.; Vincent, A.L. Detection and characterization of an H4N6 avian-lineage influenza A virus in pigs in the Midwestern United States. Virology 2017, 511, 56–65. [Google Scholar] [CrossRef]
- Karasin, A.I.; Brown, I.H.; Carman, S.; Olsen, C.W. Isolation and characterization of H4N6 avian influenza viruses from pigs with pneumonia in Canada. J. Virol. 2000, 74, 9322–9327. [Google Scholar] [CrossRef]
- Gulyaeva, M.; Sobolev, I.; Sharshov, K.; Kurskaya, O.; Alekseev, A.; Shestopalova, L.; Kovner, A.; Bi, Y.; Shi, W.; Shchelkanov, M.; et al. Characterization of Avian-like Influenza A (H4N6) Virus Isolated from Caspian Seal in 2012. Virol. Sin. 2018, 33, 449–452. [Google Scholar] [CrossRef]
- Kayali, G.; Barbour, E.; Dbaibo, G.; Tabet, C.; Saade, M.; Shaib, H.A.; Debeauchamp, J.; Webby, R.J. Evidence of infection with H4 and H11 avian influenza viruses among Lebanese chicken growers. PLoS ONE 2011, 6, e26818. [Google Scholar] [CrossRef]
- Veits, J.; Weber, S.; Stech, O.; Breithaupt, A.; Gräber, M.; Gohrbandt, S.; Bogs, J.; Hundt, J.; Teifke, J.P.; Mettenleiter, T.C.; et al. Avian influenza virus hemagglutinins H2, H4, H8, and H14 support a highly pathogenic phenotype. Proc. Natl. Acad. Sci. USA 2012, 109, 2579–2584. [Google Scholar] [CrossRef]
- Li, X.; Liu, B.; Ma, S.; Cui, P.; Liu, W.; Li, Y.; Guo, J.; Chen, H. High frequency of reassortment after co-infection of chickens with the H4N6 and H9N2 influenza A viruses and the biological characteristics of the reassortants. Vet. Microbiol. 2018, 222, 11–17. [Google Scholar] [CrossRef]
- Song, H.; Qi, J.; Xiao, H.; Bi, Y.; Zhang, W.; Xu, Y.; Wang, F.; Shi, Y.; Gao, G.F. Avian-to-Human Receptor-Binding Adaptation by Influenza A Virus Hemagglutinin H4. Cell Rep. 2017, 20, 1201–1214. [Google Scholar] [CrossRef]
- Zhang, T.; Fan, K.; Zhang, X.; Xu, Y.; Xu, J.; Xu, B.; Li, R. Diversity of avian influenza A(H5N6) viruses in wild birds in southern China. J. Gen. Virol. 2020, 101, 902–909. [Google Scholar] [CrossRef] [PubMed]
- Killian, M.L. Hemagglutination Assay for Influenza Virus. Methods Mol. Biol. 2020, 2123, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.N.; Zhang, H.J.; Li, D.; Zhou, J.N.; Chen, Y.Y.; Sun, Y.H.; Adeola, A.C.; Fu, X.Q.; Shao, Y.; Zhang, M.L. Whole-genome sequencing reveals origin and evolution of influenza A(H1N1)pdm09 viruses in Lincang, China, from 2014 to 2018. PLoS ONE 2020, 15, e0234869. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.M.; Li, K.S.; Smith, G.J.; Li, J.W.; Tai, H.; Zhang, J.X.; Webster, R.G.; Peiris, J.S.; Chen, H.; Guan, Y. Evolution and molecular epidemiology of H9N2 influenza A viruses from quail in southern China, 2000 to 2005. J. Virol. 2007, 81, 2635–2645. [Google Scholar] [CrossRef]
- Duan, L.; Bahl, J.; Smith, G.J.D.; Wang, J.; Vijaykrishna, D.; Zhang, L.J.; Zhang, J.X.; Li, K.S.; Fan, X.H.; Cheung, C.L.; et al. The development and genetic diversity of H5N1 influenza virus in China, 1996–2006. Virology 2008, 380, 243–254. [Google Scholar] [CrossRef]
- Sun, H.; Li, H.; Tong, Q.; Han, Q.; Liu, J.; Yu, H.; Song, H.; Qi, J.; Li, J.; Yang, J.; et al. Airborne transmission of human-isolated avian H3N8 influenza virus between ferrets. Cell 2023, 186, 4074–4084.e11. [Google Scholar] [CrossRef]
- Watanabe, Y.; Ibrahim, M.S.; Ellakany, H.F.; Kawashita, N.; Mizuike, R.; Hiramatsu, H.; Sriwilaijaroen, N.; Takagi, T.; Suzuki, Y.; Ikuta, K. Acquisition of human-type receptor binding specificity by new H5N1 influenza virus sublineages during their emergence in birds in Egypt. PLoS Pathog. 2011, 7, e1002068. [Google Scholar] [CrossRef]
- Liu, M.; Huang, L.Z.X.; Smits, A.A.; Bull, C.; Narimatsu, Y.; van Kuppeveld, F.J.M.; Clausen, H.; de Haan, C.A.M.; de Vries, E. Human-type sialic acid receptors contribute to avian influenza A virus binding and entry by hetero-multivalent interactions. Nat. Commun. 2022, 13, 4054. [Google Scholar] [CrossRef] [PubMed]
- Llompart, C.M.; Nieto, A.; Rodriguez-Frandsen, A. Specific Residues of PB2 and PA Influenza Virus Polymerase Subunits Confer the Ability for RNA Polymerase II Degradation and Virus Pathogenicity in Mice. J. Virol. 2014, 88, 3455–3463. [Google Scholar] [CrossRef]
- Tian, J.M.; Li, M.H.; Bai, X.L.; Li, Y.L.; Wang, X.F.; Wang, F.H.; Shi, J.Z.; Zeng, X.Y.; Tian, G.B.; Li, Y.B. H5 low pathogenic avian influenza viruses maintained in wild birds in China. Vet. Microbiol. 2021, 263, 109268. [Google Scholar] [CrossRef]
- Dong, X.; Soong, L. Emerging and Re-emerging zoonoses are major and global challenges for public health. Zoonoses 2021, 1. [Google Scholar] [CrossRef]
- Qi, X.; Qiu, H.; Hao, S.; Zhu, F.; Huang, Y.; Xu, K.; Yu, H.; Wang, D.; Zhou, L.; Dai, Q.; et al. Human Infection with an Avian-Origin Influenza A (H10N3) Virus. N. Engl. J. Med. 2022, 386, 1087–1088. [Google Scholar] [CrossRef]
- Zhang, Y.; Shi, J.; Cui, P.; Zhang, Y.; Chen, Y.; Hou, Y.; Liu, L.; Jiang, Y.; Guan, Y.; Chen, H.; et al. Genetic analysis and biological characterization of H10N3 influenza A viruses isolated in China from 2014 to 2021. J. Med. Virol. 2023, 95, e28476. [Google Scholar] [CrossRef] [PubMed]
- Pasick, J.; Pedersen, J.; Hernandez, M.S. Avian Influenza in North America, 2009–2011. Avian Dis. 2012, 56, 845–848. [Google Scholar] [CrossRef] [PubMed]
- Wille, M.; Tolf, C.; Latorre-Margalef, N.; Fouchier, R.A.M.; Halpin, R.A.; Wentworth, D.E.; Ragwani, J.; Pybus, O.G.; Olsen, B.; Waldenstrom, J. Evolutionary features of a prolific subtype of avian influenza A virus in European waterfowl. Virus Evol. 2022, 8, veac074. [Google Scholar] [CrossRef]
- Shi, Y.; Cui, H.; Wang, J.; Chi, Q.; Li, X.; Teng, Q.; Chen, H.; Yang, J.; Liu, Q.; Li, Z. Characterizations of H4 avian influenza viruses isolated from ducks in live poultry markets and farm in Shanghai. Sci. Rep. 2016, 6, 37843. [Google Scholar] [CrossRef] [PubMed]
- Quan, C.S.; Huang, T.; Chen, X.W.; Zhang, J.; Wang, Q.L.; Zhang, C.; Zhang, T.Y.; Zhou, L.J.; Shu, L.M.; Long, C.R.; et al. Genomic characterizations of H4 subtype avian influenza viruses from live poultry markets in Sichuan province of China, 2014–2015. Sci. China-Life Sci. 2018, 61, 1123–1126. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
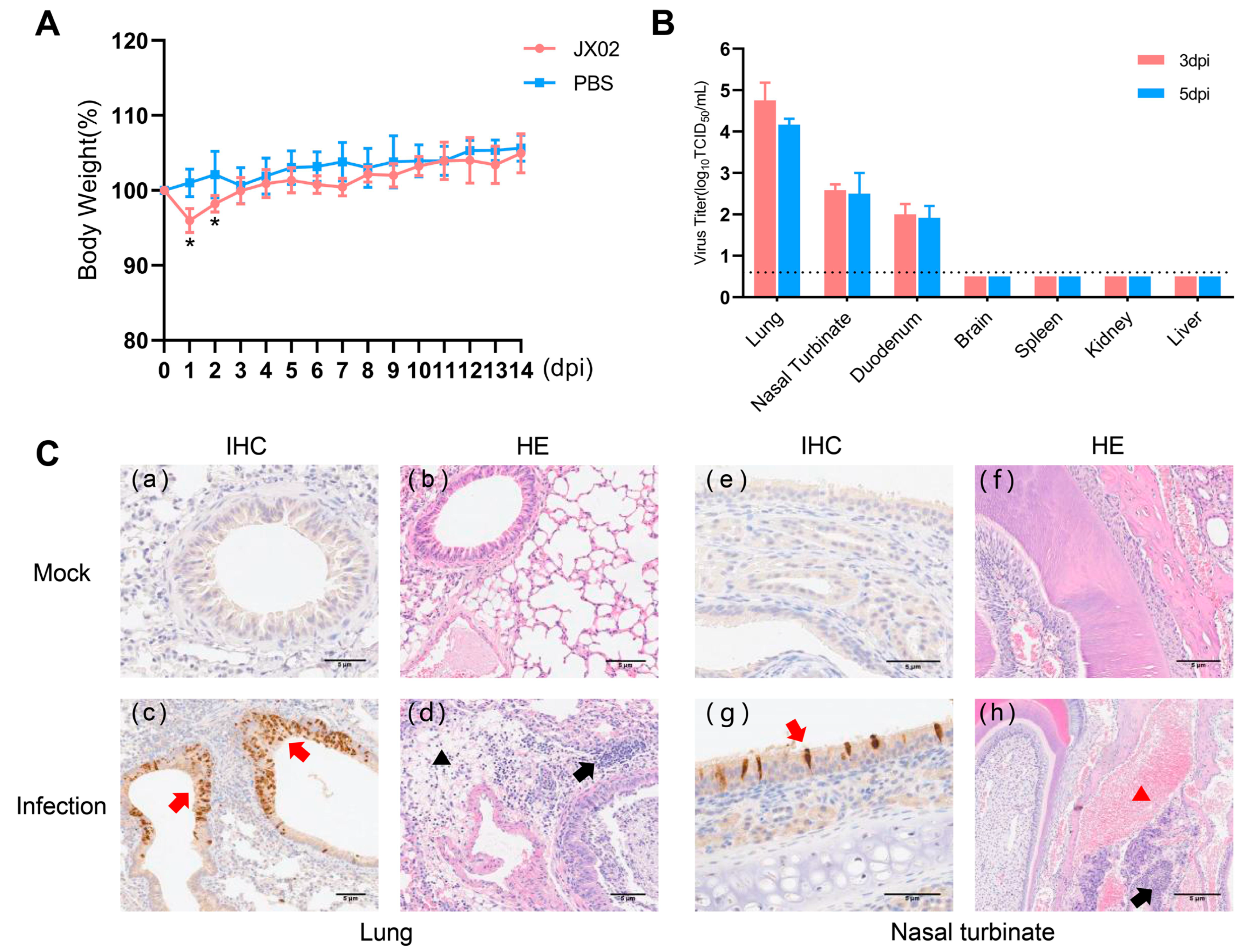{kind=link}
| Virus | Group of Each Gene Segment in the Phylogenetic Tree | Genotype | |||||||
|---|---|---|---|---|---|---|---|---|---|
| HA | NA | PB2 | PB1 | PA | NP | M | NS | ||
| A/duck/Hong Kong/365/1978 | 2 | 2 | 5 | 4 | 7 | 7 | 3 | 3 | G1 |
| A/mallard/Sweden/1705/2002 | 1 | 2 | 2 | 3 | 6 | 3 | 4 | 3 | G2 |
| A/mallard/Sweden/4508/2004 | 1 | 2 | 2 | 2 | 5 | 2 | 2 | 3 | G3 |
| A/mallard/New_Zealand/1615-17/2004 | 3 | 3 | 5 | 4 | 5 | 3 | 3 | 3 | G4 |
| A/duck/Shiga/8/2004 | 2 | 2 | 2 | 3 | 2 | 1 | 4 | 3 | G5 |
| A/mallard/ZhaLong/88/2004 | 2 | 1 | 4 | 3 | 7 | 3 | 2 | 3 | G6 |
| A/mallard/Yan chen/2005 | 2 | 2 | 3 | 3 | 3 | 1 | 2 | 3 | G7 |
| A/mallard/Sweden/58369/2006 | 1 | 2 | 5 | 2 | 4 | 5 | 2 | 2 | G8 |
| A/wild_bird/Korea/YS109/2007 | 2 | 1 | 2 | 2 | 5 | 1 | 4 | 3 | G9 |
| A/duck/Bangladesh/1283/2008 | 1 | 1 | 2 | 2 | 6 | 4 | 2 | 3 | G10 |
| A/avian/Japan/8KI0185/2008 | 1 | 1 | 2 | 2 | 7 | 6 | 2 | 3 | G11 |
| A/mallard/Sweden/81019/2008 | 1 | 1 | 1 | 2 | 6 | 4 | 3 | 2 | G12 |
| A/wild_duck/Korea/SH5-60/2008 | 1 | 1 | 5 | 2 | 3 | 1 | 2 | 3 | G13 |
| A/duck/Bangladesh/1521/2009 | 1 | 1 | 1 | 2 | 2 | 3 | 2 | 3 | G14 |
| A/wild_duck/Korea/PSC6-1/2009 | 1 | 1 | 5 | 2 | 2 | 2 | 2 | 3 | G15 |
| A/mallard/Sweden/104690/2009 | 1 | 1 | 1 | 2 | 2 | 2 | 2 | 2 | G16 |
| A/duck/Bangladesh/1784/2010 | 1 | 1 | 5 | 1 | 3 | 5 | 5 | 3 | G17 |
| A/environment/Kagoshima/KU-ngr-D/2012 | 1 | 1 | 5 | 2 | 4 | 6 | 2 | 3 | G18 |
| A/duck/Hunan/S11893/2012 | 1 | 1 | 2 | 2 | 1 | 7 | 3 | 3 | G19 |
| A/Caspian_seal/Russia/T1/2012 | 1 | 1 | 2 | 2 | 2 | 2 | 2 | 3 | G20 |
| A/muscovy_duck/Vietnam/LBM687/2014 | 1 | 1 | 5 | 2 | 1 | 6 | 1 | 3 | G21 |
| A/mallard/Beijing/6/2016 | 1 | 1 | 2 | 2 | 1 | 2 | 1 | 3 | G22 |
| A/environment/Jiangxi/F0114/2022 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | G23 |
| A/duck/Jiangxi/F011501/2022 | 1 | 1 | 1 | 1 | 3 | 1 | 1 | 1 | G24 |
| A/duck/Jiangxi/F011502/2022 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | G23 |
| Gene | Site | Function | JX14 | JX01 | JX02 |
|---|---|---|---|---|---|
| HA (H3 numbering) | Cleavage site | 324PEKASR↓GLF | 324PEKASR↓GLF | 324PEKASR↓GLF | |
| T160A | RBS positions altered receptor specificity | A | A | A | |
| E190D | E | E | E | ||
| G225D | G | G | G | ||
| Q226L | Q | Q | Q | ||
| G228S | G | G | G | ||
| Glycosylation sites | 6NYT8, 22NGT24, 165NLT167, 296NIS298, 483NGT485 | 6NYT8, 22NGT24, 165NLT167, 296NIS298, 483NGT485 | 6NYT8, 22NGT24, 165NLT167, 296NIS298, 483NGT485 | ||
| NA (N2 numbering) | H274Y | Antiviral resistance | H | H | H |
| Glycosylation sites | 51NET53, 54 NST56, 62NNT64, 67NFT69, 70NIT72, 86NLT88, 146NGT148, 201NAS203, 402NWS404 | 51NET53, 54 NST56, 62NNT64, 67NFT69, 70NIT72, 86NLT88, 146NGT148, 201NAS203, 402NWS404 | 51NET53, 54 NST56, 62NNT64, 67NFT69, 70NIT72, 86NLT88, 146NGT148, 201NAS203, 402NWS404 | ||
| PB2 | I504V | Enhanced polymerase activity and increased virulence in mice | V | V | V |
| E627K/V | Mammalian adaptive mutations; enhanced polymerase activity and increased virulence in mice | E | E | E | |
| D701N | D | D | D | ||
| M1 | I43M | Increased virulence in mice | M | M | M |
| T215A | A | A | A | ||
| M2 | S31N | Antiviral resistance | S | S | S |
| NS1 | I106M | Increased virulence in mice | M | M | M |
| C138F | F | F | F |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, T.; Zhao, C.; Guo, Y.; Dong, J.; Du, F.; Zhou, Y.; Shu, S.; Liu, Y.; Cheng, Y.; Cao, Z.; et al. Genetic and Biological Characteristics of Duck-Origin H4N6 Avian Influenza Virus Isolated in China in 2022. Viruses 2024, 16, 207. https://doi.org/10.3390/v16020207
Li T, Zhao C, Guo Y, Dong J, Du F, Zhou Y, Shu S, Liu Y, Cheng Y, Cao Z, et al. Genetic and Biological Characteristics of Duck-Origin H4N6 Avian Influenza Virus Isolated in China in 2022. Viruses. 2024; 16(2):207. https://doi.org/10.3390/v16020207
Chicago/Turabian StyleLi, Tian, Chuankuo Zhao, Yuxin Guo, Jinze Dong, Fanshu Du, Yong Zhou, Sicheng Shu, Yang Liu, Yachang Cheng, Zhiyong Cao, and et al. 2024. "Genetic and Biological Characteristics of Duck-Origin H4N6 Avian Influenza Virus Isolated in China in 2022" Viruses 16, no. 2: 207. https://doi.org/10.3390/v16020207
APA StyleLi, T., Zhao, C., Guo, Y., Dong, J., Du, F., Zhou, Y., Shu, S., Liu, Y., Cheng, Y., Cao, Z., Cao, Q., Shi, S., Huang, Y., Pu, J., & Liu, L. (2024). Genetic and Biological Characteristics of Duck-Origin H4N6 Avian Influenza Virus Isolated in China in 2022. Viruses, 16(2), 207. https://doi.org/10.3390/v16020207