

Begomovirus Transmission to Tomato Plants Is Not Hampered by Plant Defenses Induced by Dicyphus hesperus Knight
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plants, Virus, and Insects
2.2. Plant Defenses Induced by Mirids in Plants without TYLCV
2.3. Virus Transmission by B. tabaci in Plants Exposed to Mirids
2.4. Virus Acquisition by Whiteflies from TYLCV-Infected Plants Exposed to Mirids
2.5. Data Analysis
3. Results
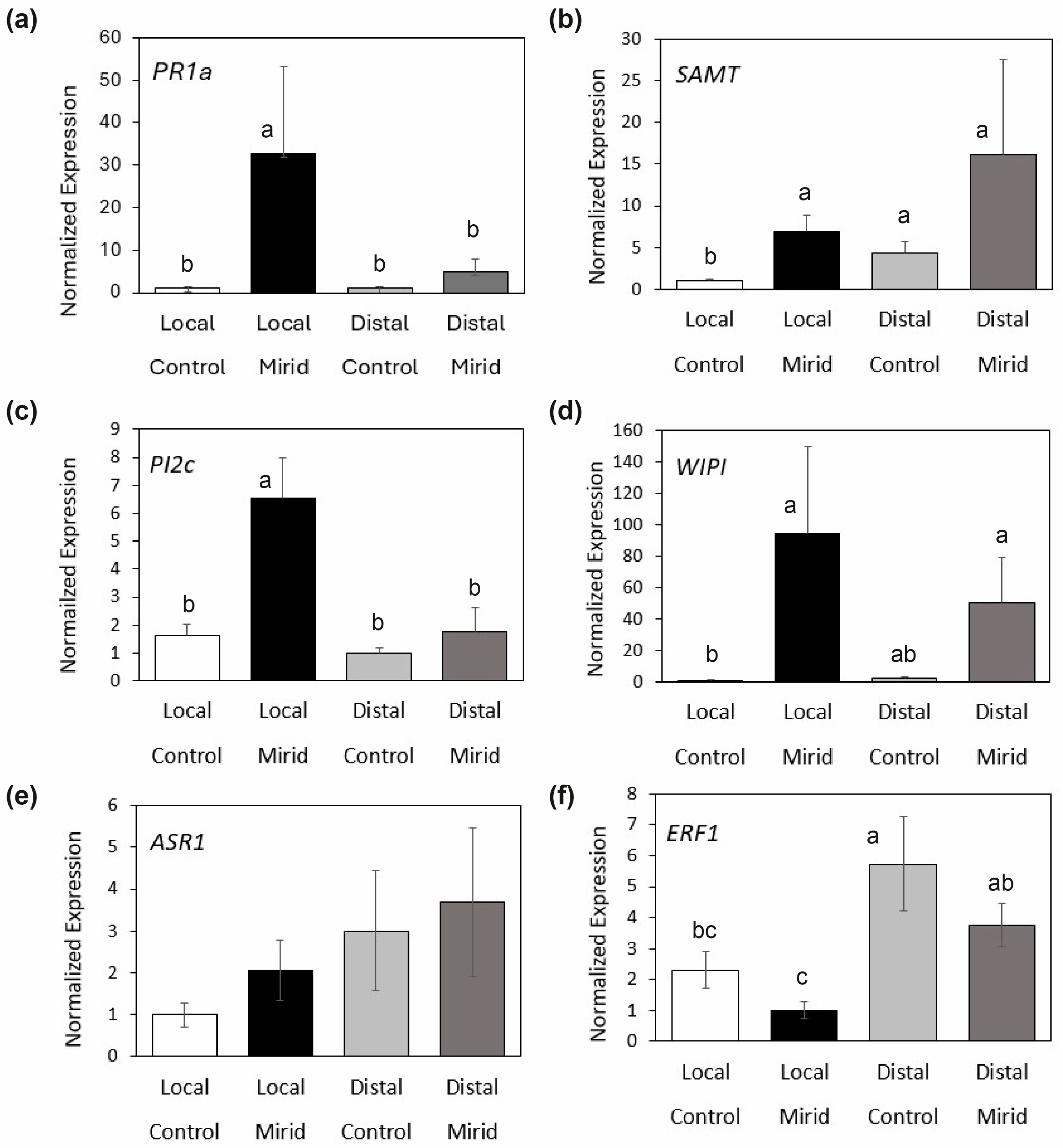
3.1. Tomato Plant Defenses Induced by D. hesperus
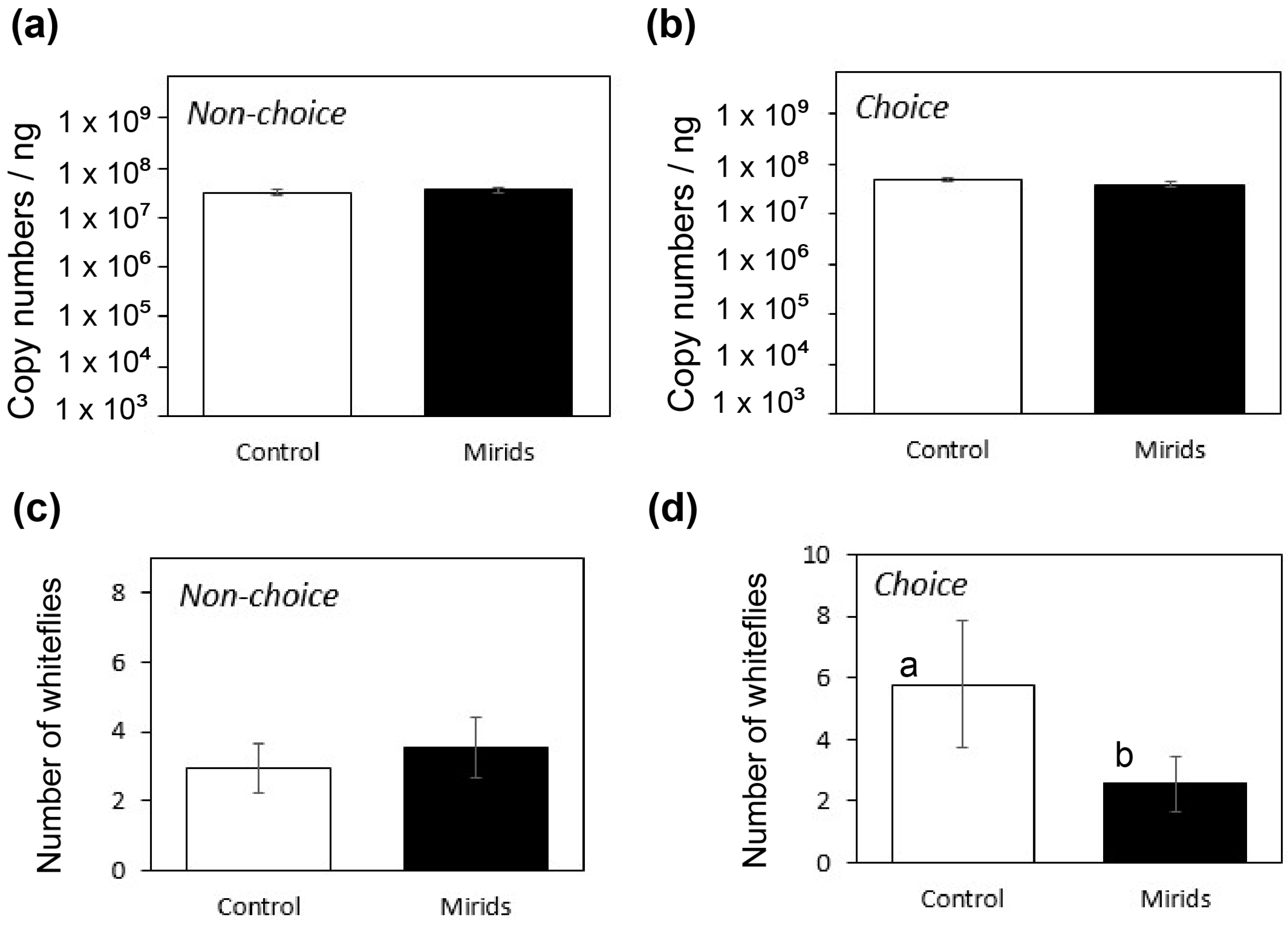
3.2. Virus Transmission in Plants Exposed to D. hesperus
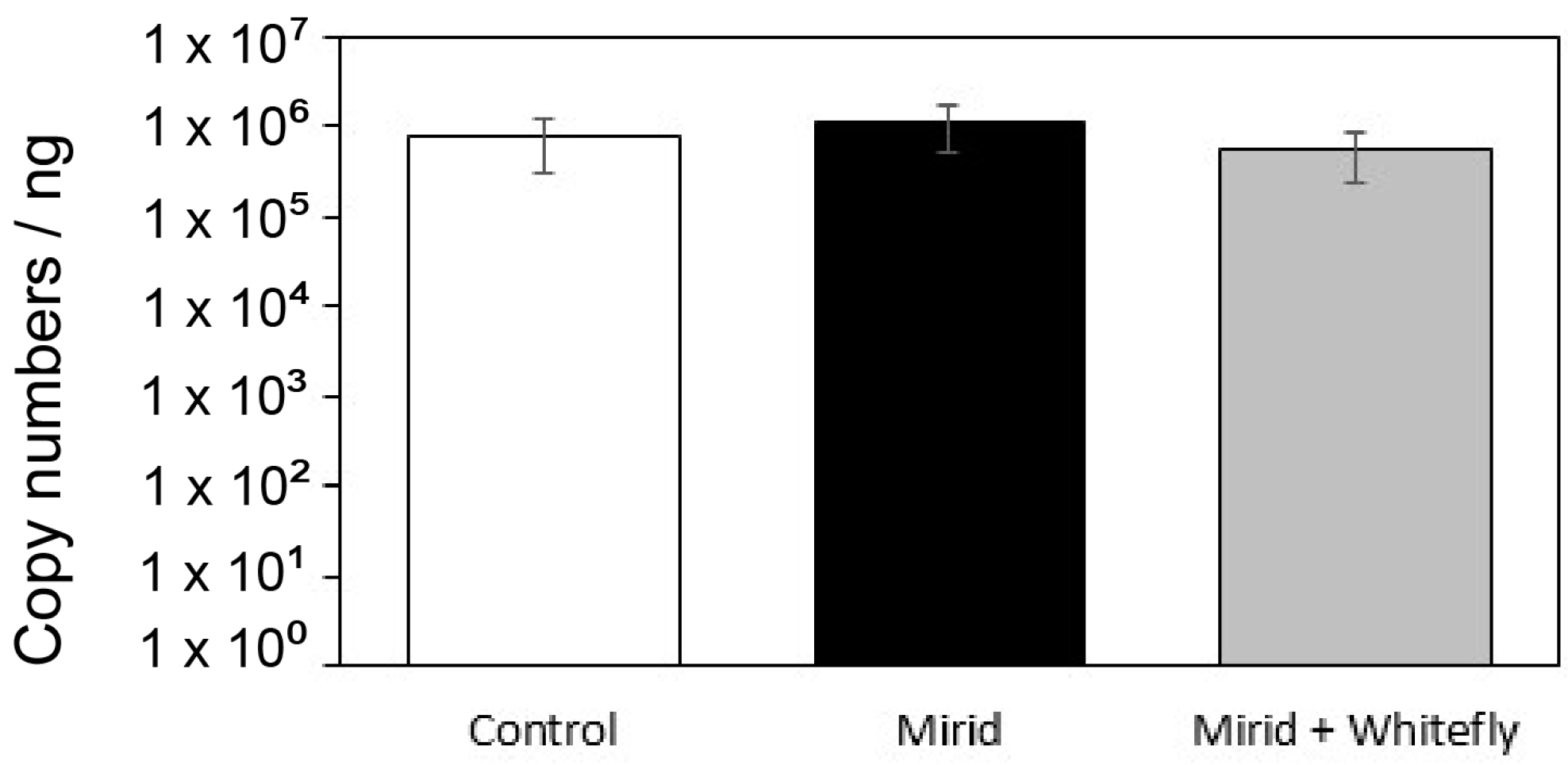
3.3. Acquisition by Whiteflies of TYLCV from Plants Exposed to D. hesperus
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Walling, L.L. The myriad plant responses to herbivores. J. Plant Growth Regul. 2000, 19, 195–216. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; Van Der Does, D.; Zamioudis, C.; Leon-Reyes, A.; Van Wees, S.C.M. Hormonal modulation of plant immunity. Annu. Rev. Cell Dev. Biol. 2012, 28, 489–521. [Google Scholar] [CrossRef]
- Pan, L.L.; Miao, H.; Wang, Q.; Walling, L.L.; Liu, S.S. Virus-induced phytohormone dynamics and their effects on plant–insect interactions. New Phytol. 2021, 230, 1305–1320. [Google Scholar] [CrossRef]
- Howe, G.A.; Jander, G. Plant immunity to insect herbivores. Annu. Rev. Plant Biol. 2008, 59, 41–66. [Google Scholar] [CrossRef]
- Zhu-Salzman, K.; Luthe, D.S.; Felton, G.W. Arthropod-inducible proteins: Broad spectrum defenses against multiple herbivores. Plant Physiol. 2008, 146, 852–858. [Google Scholar] [CrossRef]
- Chen, H.; Wilkerson, C.G.; Kuchar, J.A.; Phinney, B.S.; Howe, G.A. Jasmonate-inducible plant enzymes degrade essential amino acids in the herbivore midgut. Proc. Natl. Acad. Sci. USA 2005, 102, 19237–19242. [Google Scholar] [CrossRef]
- Park, S.W.; Kaimoyo, E.; Kumar, D.; Mosher, S.; Klessig, D.F. Methyl salicylate is a critical mobile signal for plant systemic acquired resistance. Science 2007, 318, 113–116. [Google Scholar] [CrossRef]
- Tripathi, D.; Raikhy, G.; Kumar, D. Chemical elicitors of systemic acquired resistance—Salicylic acid and its functional analogs. Curr. Plant Biol. 2019, 17, 48–59. [Google Scholar] [CrossRef]
- Ament, K.; Krasikov, V.; Allmann, S.; Rep, M.; Takken, F.L.W.; Schuurink, R.C. Methyl salicylate production in tomato affects biotic interactions. Plant J. 2010, 62, 124–134. [Google Scholar] [CrossRef]
- Zhu, F.; Cao, M.Y.; Zhang, Q.P.; Mohan, R.; Schar, J.; Mitchell, M.; Chen, H.; Liu, F.; Wang, D.; Fu, Z.Q. Join the green team: Inducers of plant immunity in the plant disease sustainable control toolbox. J. Adv. Res. 2023, 57, 15–42. [Google Scholar] [CrossRef]
- Walters, D.R.; Ratsep, J.; Havis, N.D. Controlling crop diseases using induced resistance: Challenges for the future. J. Exp. Bot. 2013, 64, 1263–1280. [Google Scholar] [CrossRef]
- Kenney, J.R.; Grandmont, M.E.; Mauck, K.E. Priming melon defenses with acibenzolar-s-methyl attenuates infections by phylogenetically distinct viruses and diminishes vector preferences for infected hosts. Viruses 2020, 12, 257. [Google Scholar] [CrossRef]
- Doungous, O.; Khatabi, B.; Hanna, R.; Tchuanyo, M.; Kuate, A.F.; Fondong, V.N. Acibenzolar-s-methyl induces resistance against cassava mosaic geminiviruses in Nicotiana benthamiana and their vector Bemisia tabaci in cassava (Manihot esculenta). Crop Prot. 2021, 150, 105796. [Google Scholar] [CrossRef]
- Pappu, H.R.; Csinos, A.S.; McPherson, R.M.; Jones, D.C.; Stephenson, M.G. Effect of acibenzolar-s-methyl and imidacloprid on suppression of tomato spotted wilt tospovirus in flue-cured tobacco. Crop Prot. 2000, 19, 349–354. [Google Scholar] [CrossRef]
- Su, Q.; Yang, F.; Zhang, Q.; Tong, H.; Hu, Y.; Zhang, X.; Xie, W.; Wang, S.; Wu, Q.; Zhang, Y. Defence priming in tomato by the green leaf volatile (Z)-3-hexenol reduces whitefly transmission of a plant virus. Plant Cell Environ. 2020, 43, 2797–2811. [Google Scholar] [CrossRef]
- Zhang, N.X.; Stephan, J.G.; Björkman, C.; Puentes, A. Global change calls for novel plant protection: Reviewing the potential of omnivorous plant-inhabiting arthropods as predators and plant defence inducers. Curr. Opin. Insect Sci. 2021, 47, 103–110. [Google Scholar] [CrossRef]
- Roda, A.; Castillo, J.; Allen, C.; Urbaneja, A.; Pérez-Hedo, M.; Weihman, S.; Stansly, P.A. Biological control potential and drawbacks of three zoophytophagous mirid predators against Bemisia tabaci in the United States. Insects 2020, 11, 670. [Google Scholar] [CrossRef]
- Smith, H.A.; Krey, K.L. Three release rates of Dicyphus hesperus (Hemiptera: Miridae) for management of Bemisia tabaci (Hemiptera: Aleyrodidae) on greenhouse tomato. Insects 2019, 10, 213. [Google Scholar] [CrossRef]
- Pappas, M.L.; Steppuhn, A.; Geuss, D.; Topalidou, N.; Zografou, A.; Sabelis, M.W.; Broufas, G.D. Beyond predation: The zoophytophagous predator Macrolophus pygmaeus induces tomato resistance against spider mites. PLoS ONE 2015, 10, e0127251. [Google Scholar] [CrossRef]
- Zhang, N.X.; Messelink, G.J.; Alba, J.M.; Schuurink, R.C.; Kant, M.R.; Janssen, A. Phytophagy of omnivorous predator Macrolophus pygmaeus affects performance of herbivores through induced plant defences. Oecologia 2018, 186, 101–113. [Google Scholar] [CrossRef]
- Bouagga, S.; Urbaneja, A.; Depalo, L.; Rubio, L.; Pérez-Hedo, M. Zoophytophagous predator-induced defences restrict accumulation of the tomato spotted wilt virus. Pest Manag. Sci. 2020, 76, 561–567. [Google Scholar] [CrossRef]
- Pérez-Hedo, M.; Bouagga, S.; Jaques, J.A.; Flors, V.; Urbaneja, A. Tomato plant responses to feeding behavior of three zoophytophagous predators (Hemiptera: Miridae). Biol. Control 2015, 86, 46–51. [Google Scholar] [CrossRef]
- Pérez-Hedo, M.; Bouagga, S.; Zhang, N.X.; Moerkens, R.; Messelink, G.; Jaques, J.A.; Flors, V.; Broufas, G.; Urbaneja, A.; Pappas, M.L. Induction of plant defenses: The added value of zoophytophagous predators. J. Pest Sci. 2022, 95, 1501–1517. [Google Scholar] [CrossRef]
- Zhang, N.X.; van Wieringen, D.; Messelink, G.J.; Janssen, A. Herbivores avoid host plants previously exposed to their omnivorous predator Macrolophus pygmaeus. J. Pest Sci. 2019, 92, 737–745. [Google Scholar] [CrossRef]
- Pérez-Hedo, M.; Alonso-Valiente, M.; Vacas, S.; Gallego, C.; Rambla, J.L.; Navarro-Llopis, V.; Granell, A.; Urbaneja, A. Eliciting tomato plant defenses by exposure to herbivore induced plant volatiles. Entomol. Gen. 2021, 41, 209–218. [Google Scholar] [CrossRef]
- Yan, Z.; Wolters, A.M.A.; Navas-castillo, J.; Bai, Y. The global dimension of tomato yellow leaf curl disease: Current status and breeding perspectives. Microorganisms 2021, 9, 740. [Google Scholar] [CrossRef]
- Srinivasan, R.; Riley, D.; Diffie, S.; Sparks, A.; Adkins, S. Whitefly population dynamics and evaluation of whitefly-transmitted tomato yellow leaf curl virus (TYLCV)-resistant tomato genotypes as whitefly and TYLCV reservoirs. J. Econ. Entomol. 2012, 105, 1447–1456. [Google Scholar] [CrossRef]
- Su, Q.; Mescher, M.C.; Wang, S.; Chen, G.; Xie, W.; Wu, Q.; Wang, W.; Zhang, Y. Tomato yellow leaf curl virus differentially influences plant defence responses to a vector and a non-vector herbivore. Plant Cell Environ. 2016, 39, 597–607. [Google Scholar] [CrossRef]
- Alba, J.M.; Schimmel, B.C.J.; Glas, J.J.; Ataide, L.M.S.; Pappas, M.L.; Villarroel, C.A.; Schuurink, R.C.; Sabelis, M.W.; Kant, M.R. Spider mites suppress tomato defenses downstream of jasmonate and salicylate independently of hormonal crosstalk. New Phytol. 2015, 205, 828–840. [Google Scholar] [CrossRef]
- van Kan, J.A.L.; Joosten, M.H.A.J.; Wagemakers, C.A.M.; van den Berg-Velthuis, G.C.M.; de Wit, P.J.G.M. Differential accumulation of mRNAs encoding extracellular and intracellular PR proteins in tomato induced by virulent and avirulent races of Cladosporium fulvum. Plant Mol. Biol. 1992, 20, 513–527. [Google Scholar] [CrossRef]
- Tieman, D.; Zeigler, M.; Schmelz, E.; Taylor, M.G.; Rushing, S.; Jones, J.B.; Klee, H.J. Functional analysis of a tomato salicylic acid methyl transferase and its role in synthesis of the flavor volatile methyl salicylate. Plant J. 2010, 62, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Gadea, J.; Mayda, M.E.; Conejero, V.; Vera, P. Characterization of defense-related genes ectopically expressed in viroid-infected tomato plants. Mol. Plant-Microbe Interact. 1996, 9, 409–415. [Google Scholar] [CrossRef] [PubMed]
- Graham, J.S.; Pearce, G.; Merryweather, J.; Titani, K.; Ericsson, L.H.; Ryan, C.A. Wound-induced proteinase inhibitors from tomato leaves. II. The cDNA-deduced primary structure of pre-inhibitor II. J. Biol. Chem. 1985, 260, 6561–6564. [Google Scholar] [CrossRef]
- Nambeesan, S.; AbuQamar, S.; Laluk, K.; Mattoo, A.K.; Mickelbart, M.V.; Ferruzzi, M.G.; Mengiste, T.; Handa, A.K. Polyamines attenuate ethylene-mediated defense responses to abrogate resistance to Botrytis cinerea in tomato. Plant Physiol. 2012, 158, 1034–1045. [Google Scholar] [CrossRef] [PubMed]
- Naselli, M.; Urbaneja, A.; Siscaro, G.; Jaques, J.A.; Zappalà, L.; Flors, V.; Pérez-Hedo, M. Stage-related defense response induction in tomato plants by Nesidiocoris tenuis. Int. J. Mol. Sci. 2016, 17, 1210. [Google Scholar] [CrossRef]
- The Tomato Genome Consortium. The tomato genome sequence provides insights into fleshy fruit evolution. Nature 2012, 485, 635–641. [Google Scholar] [CrossRef]
- Legarrea, S.; Barman, A.; Marchant, W.; Diffie, S.; Srinivasan, R. Temporal effects of a begomovirus infection and host plant resistance on the preference and development of an insect vector, Bemisia tabaci, and implications for epidemics. PLoS ONE 2015, 10, e0142114. [Google Scholar] [CrossRef]
- Pappas, M.L.; Steppuhn, A.; Broufas, G.D. The role of phytophagy by predators in shaping plant interactions with their pests. Commun. Integr. Biol. 2016, 9, e0127251. [Google Scholar] [CrossRef]
- Bouagga, S.; Urbaneja, A.; Rambla, J.L.; Flors, V.; Granell, A.; Jaques, J.A.; Pérez-Hedo, M. Zoophytophagous mirids provide pest control by inducing direct defences, antixenosis and attraction to parasitoids in sweet pepper plants. Pest Manag. Sci. 2018, 74, 1286–1296. [Google Scholar] [CrossRef]
- Silva, D.B.; Hanel, A.; Franco, F.P.; de Castro Silva-Filho, M.; Bento, J.M.S. Two in one: The neotropical mirid predator Macrolophus basicornis increases pest control by feeding on plants. Pest Manag. Sci. 2022, 78, 3314–3323. [Google Scholar] [CrossRef]
- Tripathi, D.; Jiang, Y.L.; Kumar, D. SABP2, a methyl salicylate esterase is required for the systemic acquired resistance induced by acibenzolar-s-methyl in plants. FEBS Lett. 2010, 584, 3458–3463. [Google Scholar] [CrossRef] [PubMed]
- Takeshita, M.; Okuda, M.; Okuda, S.; Hyodo, A.; Hamano, K.; Furuya, N.; Tsuchiya, K. Induction of antiviral responses by acibenzolar-s-methyl against cucurbit chlorotic yellows virus in melon. Phytopathology 2013, 103, 960–965. [Google Scholar] [CrossRef] [PubMed]
- Johnston, N.; Martini, X. The influence of visual and olfactory cues in host selection for Bemisia tabaci biotype b in the presence or absence of tomato yellow leaf curl virus. Insects 2020, 11, 115. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.X.; Andringa, J.; Brouwer, J.; Alba, J.M.; Kortbeek, R.W.J.; Messelink, G.J.; Janssen, A. The omnivorous predator Macrolophus pygmaeus induces production of plant volatiles that attract a specialist predator. J. Pest Sci. 2022, 95, 1343–1355. [Google Scholar] [CrossRef]
- Bouagga, S.; Urbaneja, A.; Rambla, J.L.; Granell, A.; Pérez-Hedo, M. Orius laevigatus strengthens its role as a biological control agent by inducing plant defenses. J. Pest Sci. 2018, 91, 55–64. [Google Scholar] [CrossRef]
- Cruz-Miralles, J.; Cabedo-López, M.; Pérez-Hedo, M.; Flors, V.; Jaques, J.A. Zoophytophagous mites can trigger plant-genotype specific defensive responses affecting potential prey beyond predation: The case of Euseius stipulatus and Tetranychus urticae in citrus. Pest Manag. Sci. 2019, 75, 1962–1970. [Google Scholar] [CrossRef] [PubMed]
- Rosen, R.; Kanakala, S.; Kliot, A.; Cathrin Pakkianathan, B.; Farich, B.A.; Santana-Magal, N.; Elimelech, M.; Kontsedalov, S.; Lebedev, G.; Cilia, M.; et al. Persistent, circulative transmission of begomoviruses by whitefly vectors. Curr. Opin. Virol. 2015, 15, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Czosnek, H.; Ghanim, M.; Ghanim, M. The circulative pathway of begomoviruses in the whitefly vector Bemisia tabaci—Insights from studies with tomato yellow leaf curl virus. Ann. Appl. Biol. 2002, 140, 215–231. [Google Scholar] [CrossRef]
- Naalden, D.; van Kleeff, P.J.M.; Dangol, S.; Mastop, M.; Corkill, R.; Hogenhout, S.A.; Kant, M.R.; Schuurink, R.C. Spotlight on the roles of whitefly effectors in insect–plant interactions. Front. Plant Sci. 2021, 12, 661141. [Google Scholar] [CrossRef]
- Shi, X.; Pan, H.; Zhang, H.; Jiao, X.; Xie, W.; Wu, Q.; Wang, S.; Fang, Y.; Chen, G.; Zhou, X.; et al. Bemisia tabaci Q carrying tomato yellow leaf curl virus strongly suppresses host plant defenses. Sci. Rep. 2014, 4, 5230. [Google Scholar] [CrossRef]
- Nomikou, M.; Janssen, A.; Sabelis, M.W. Herbivore host plant selection: Whitefly learns to avoid host plants that harbour predators of her offspring. Oecologia 2003, 136, 484–488. [Google Scholar] [CrossRef] [PubMed]
- Momol, M.T.; Olson, S.M.; Funderburk, J.E.; Stavisky, J.; Marois, J.J. Integrated management of tomato spotted wilt on field-grown tomatoes. Plant Dis. 2004, 88, 882–890. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, D.; Pappu, H.R. Evaluation of acibenzolar-s-methyl-induced resistance against iris yellow spot tospovirus. Eur. J. Plant Pathol. 2015, 142, 855–864. [Google Scholar] [CrossRef]
- Anfoka, G.H. Benzo-(1,2,3)-thiadiazole-7-carbothioic acid S-methyl ester induces systemic resistance in tomato (Lycopersicon esculentum. Mill cv. Vollendung) to cucumber mosaic virus. Crop Prot. 2000, 19, 401–405. [Google Scholar] [CrossRef]
- Latora, A.G.; Codod, C.B.; Legarrea, S.; Dutta, B.; Kemerait, R.C.; Adkins, S.; Turechek, W.; Coolong, T.; da Silva, A.L.B.R.; Srinivasan, R. Combining cultural tactics and insecticides for the management of the sweetpotato whitefly, Bemisia tabaci MEAM1, and viruses in yellow squash. Horticulturae 2022, 8, 341. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Gene Name | Marker Gene | Pathway | Sequence (5′->3′) | Reference |
|---|---|---|---|---|
| Pathogenesis-related protein 1a | PR1a | SA | Fw: TGGTGGTTCATTTCTTGCAACTAC Rv: ATCAATCCGATCCACTTATCATTTTA | [30] |
| Salicylic acid carboxyl methyltransferase | SAMT | SA | Fw: TCCCAGAAACATTATACATTGCTGAT Rv: AATGACCTTAACAAGTTCTGATACCACTAA | [31] |
| Proteinase Inhibitor IIc | PI2c | JA | Fw: CAGGATGTACGACGTGTTGC Rv: GAGTTTGCAACCCTCTCCTG | [32] |
| Wound-induced Proteinase Inhibitor II | WIPI | JA | Fw: GACAAGGTACTAGTAATCAATTATCC Rv: GGGCATATCCCGAACCCAAGA | [33] |
| Ethylene responsive factor | ERF | Ethylene | Fw: CGTCCGAGGAAGTGAAACTC Rv: CCGACTCGTAAGTTCCAAGC | [34] |
| Abscisic acid Stress Ripening protein 1 | ASR1 | ABA | Fw: ACACCACCACCACCACCTGT Rv: GTGTTTGTGTGCATGTTCTGGA | [35] |
| Actin | Actin | Reference * | Fw: TTAGCACCTTCCAGCAGATGT Rv: AACAGACAGGACACTCGCACT | [36] |
| Elongation Factor 1 | EF1 | Reference ** | Fw: GATTGGTGGTATTGGAACTGTC Rv: AGCTTCGTGGTGCATCTC | [35] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Legarrea, S.; LaTora, A.G.; Simmons, A.M.; Srinivasan, R. Begomovirus Transmission to Tomato Plants Is Not Hampered by Plant Defenses Induced by Dicyphus hesperus Knight. Viruses 2024, 16, 587. https://doi.org/10.3390/v16040587
Legarrea S, LaTora AG, Simmons AM, Srinivasan R. Begomovirus Transmission to Tomato Plants Is Not Hampered by Plant Defenses Induced by Dicyphus hesperus Knight. Viruses. 2024; 16(4):587. https://doi.org/10.3390/v16040587
Chicago/Turabian StyleLegarrea, Saioa, Angela Gabrielle LaTora, Alvin M. Simmons, and Rajagopalbabu Srinivasan. 2024. "Begomovirus Transmission to Tomato Plants Is Not Hampered by Plant Defenses Induced by Dicyphus hesperus Knight" Viruses 16, no. 4: 587. https://doi.org/10.3390/v16040587