
Mesenchymal Stem Cell-Derived Exosomes Attenuate Murine Cytomegalovirus-Infected Pneumonia via NF-κB/NLRP3 Signaling Pathway

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Cell Culture
2.2.1. Identification of Exosomes from 3T3 and mMSCs
2.2.2. Viral Plaque Assay and Viral Infections
2.2.3. Bronchoalveolar Lavage (BAL)
2.2.4. FACS Analysis
2.2.5. Histological Analysis and Immunofluorescence Staining
2.2.6. Enzyme-Linked Immunosorbent Assay (ELISA)
2.2.7. Western Blot Analysis
2.2.8. Statistical Analysis
3. Results
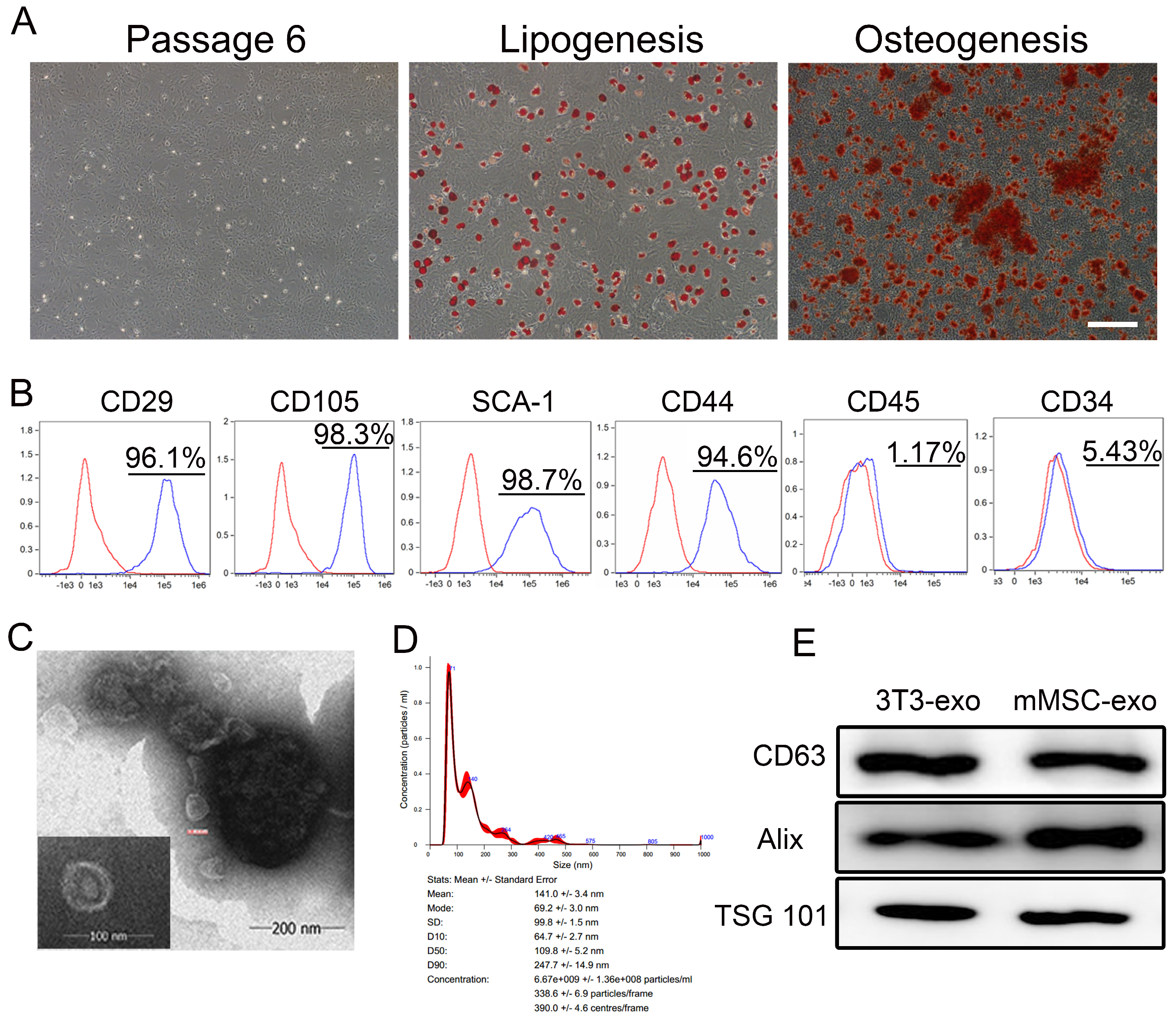
3.1. Characterization of mMSC-exos
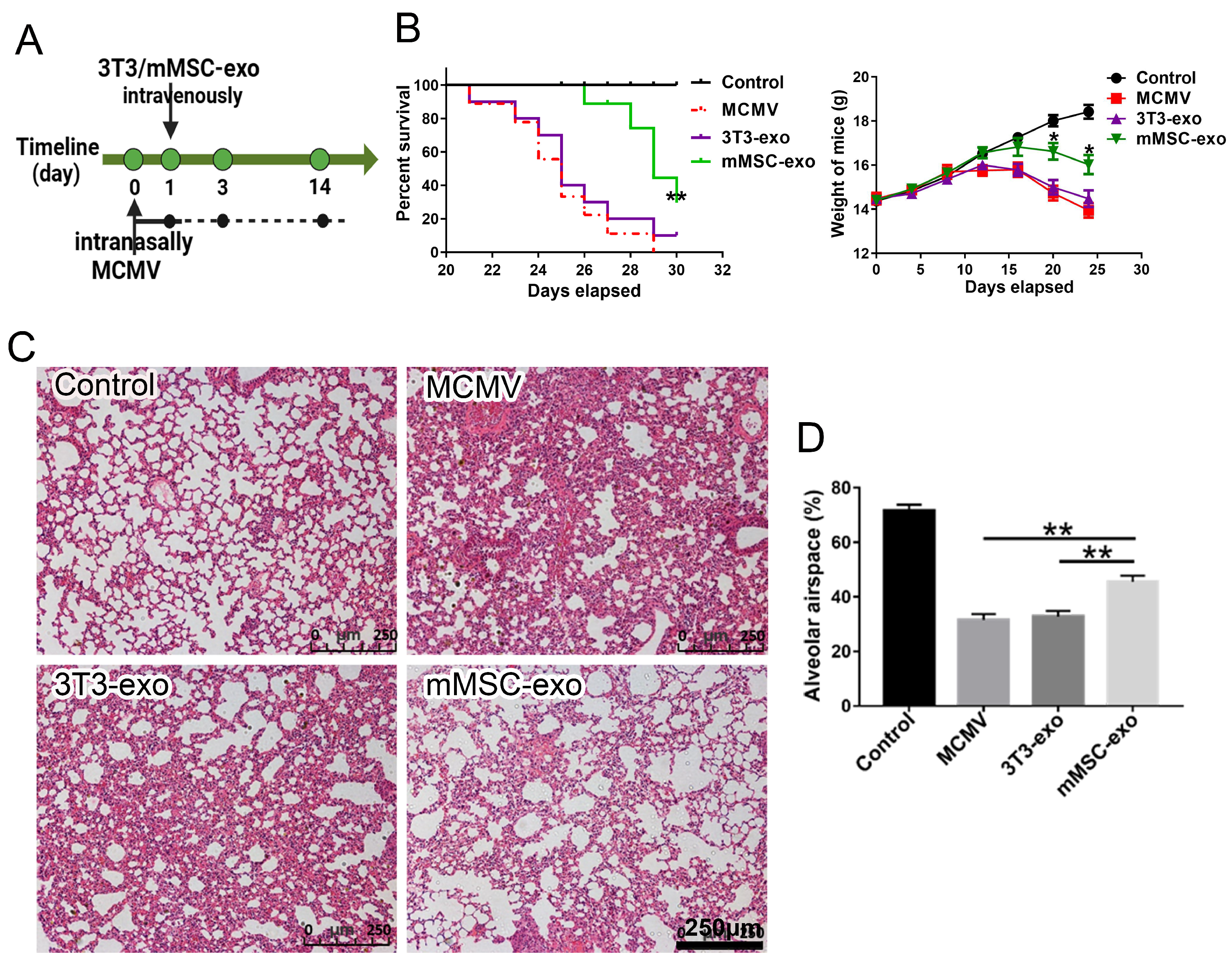
3.2. mMSC-exos Improved Survival and Reduced Lung Injury in MCMV-Infected Mice
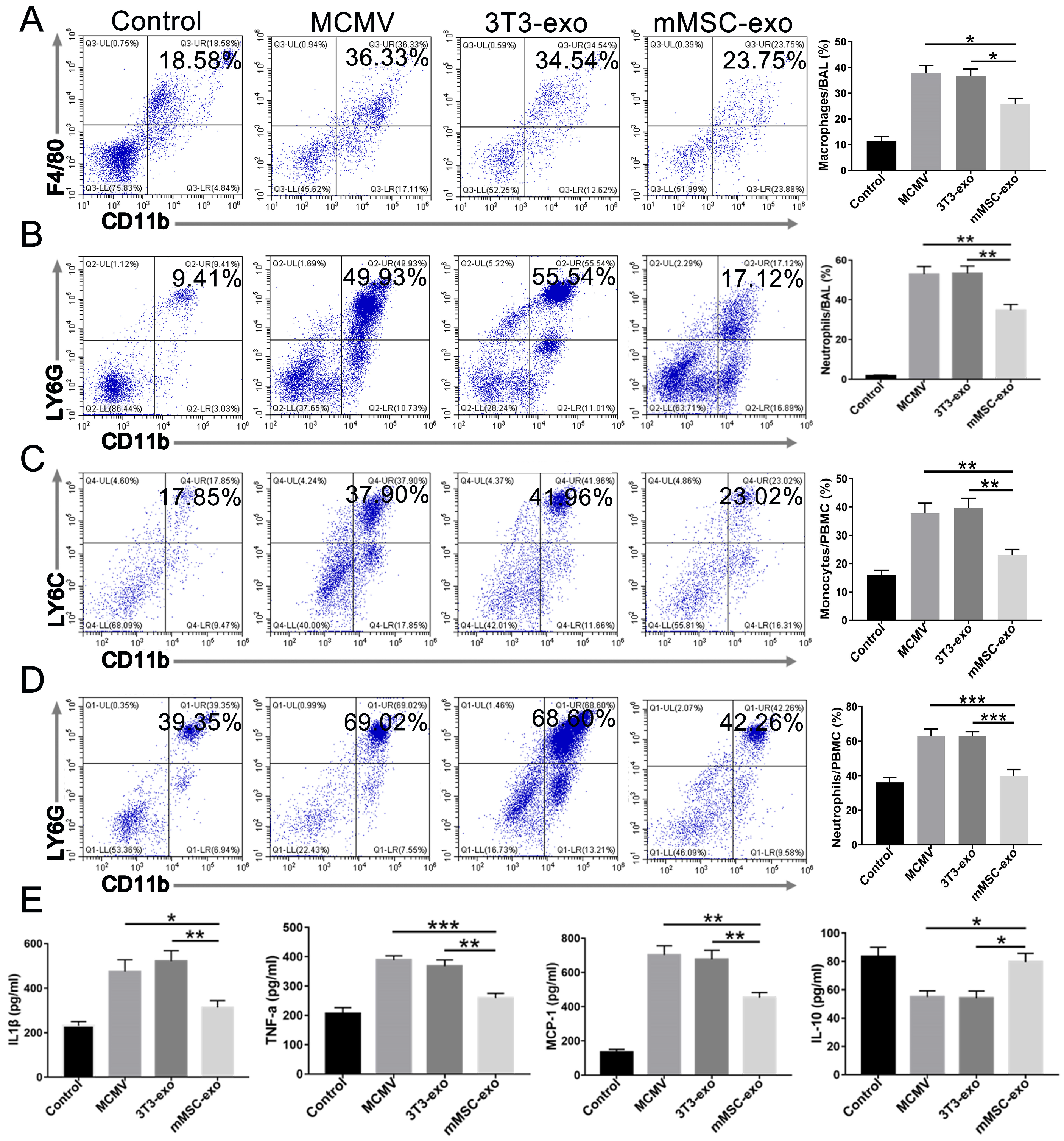
3.3. mMSC-exos Suppressed the Inflammatory Response in MCMV-Infected Mice
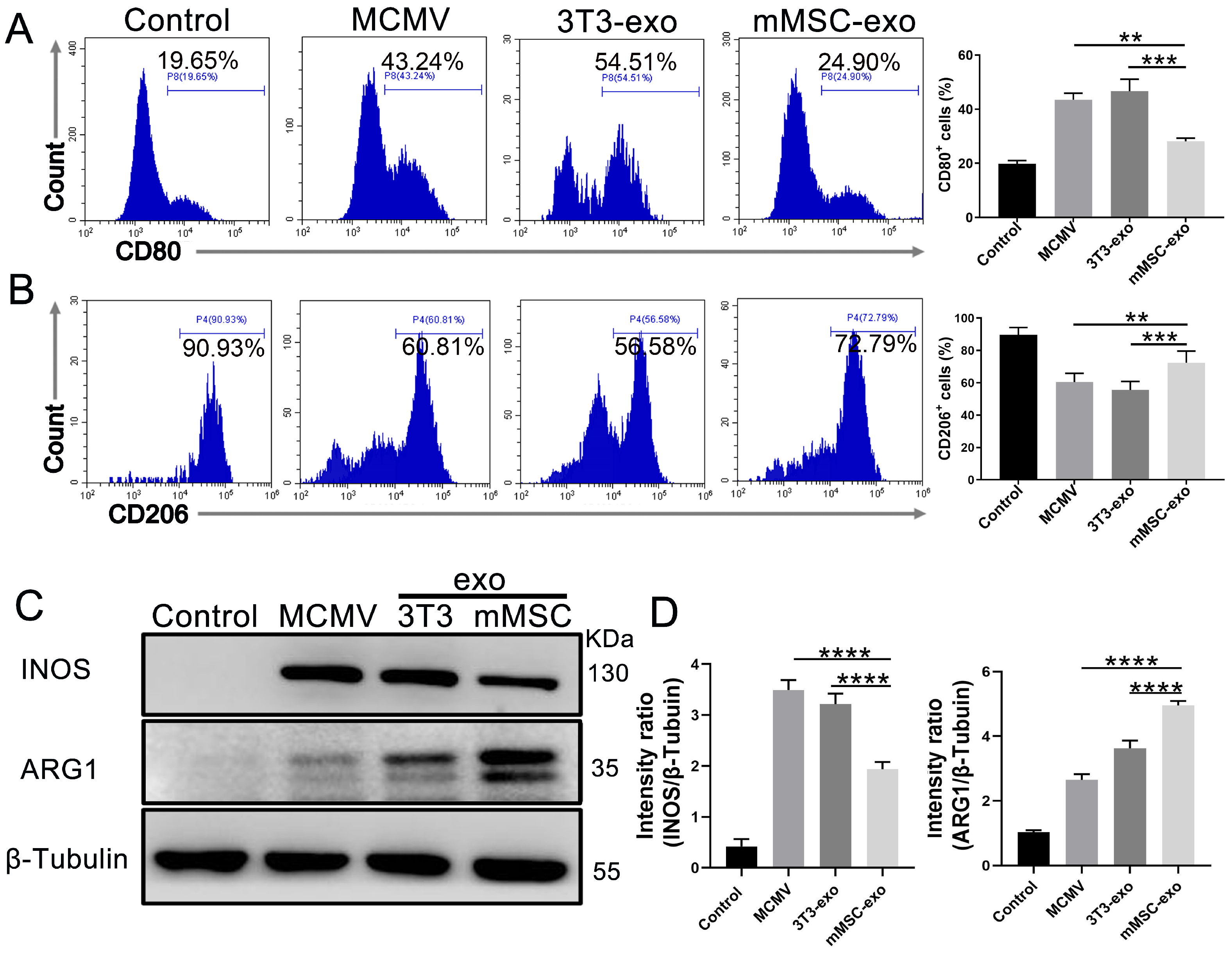
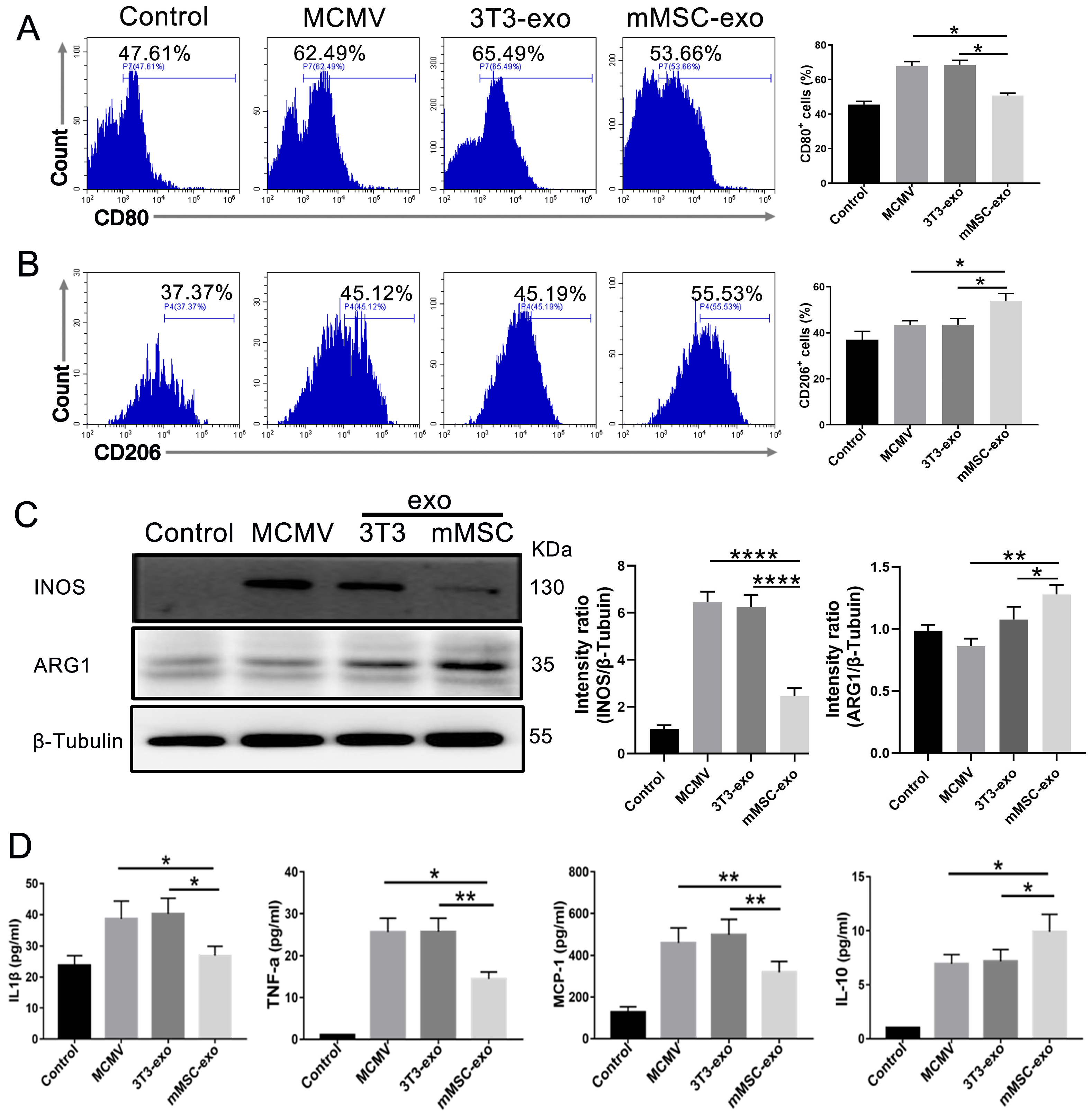
3.4. mMSC-exos Modulate Macrophage Polarization
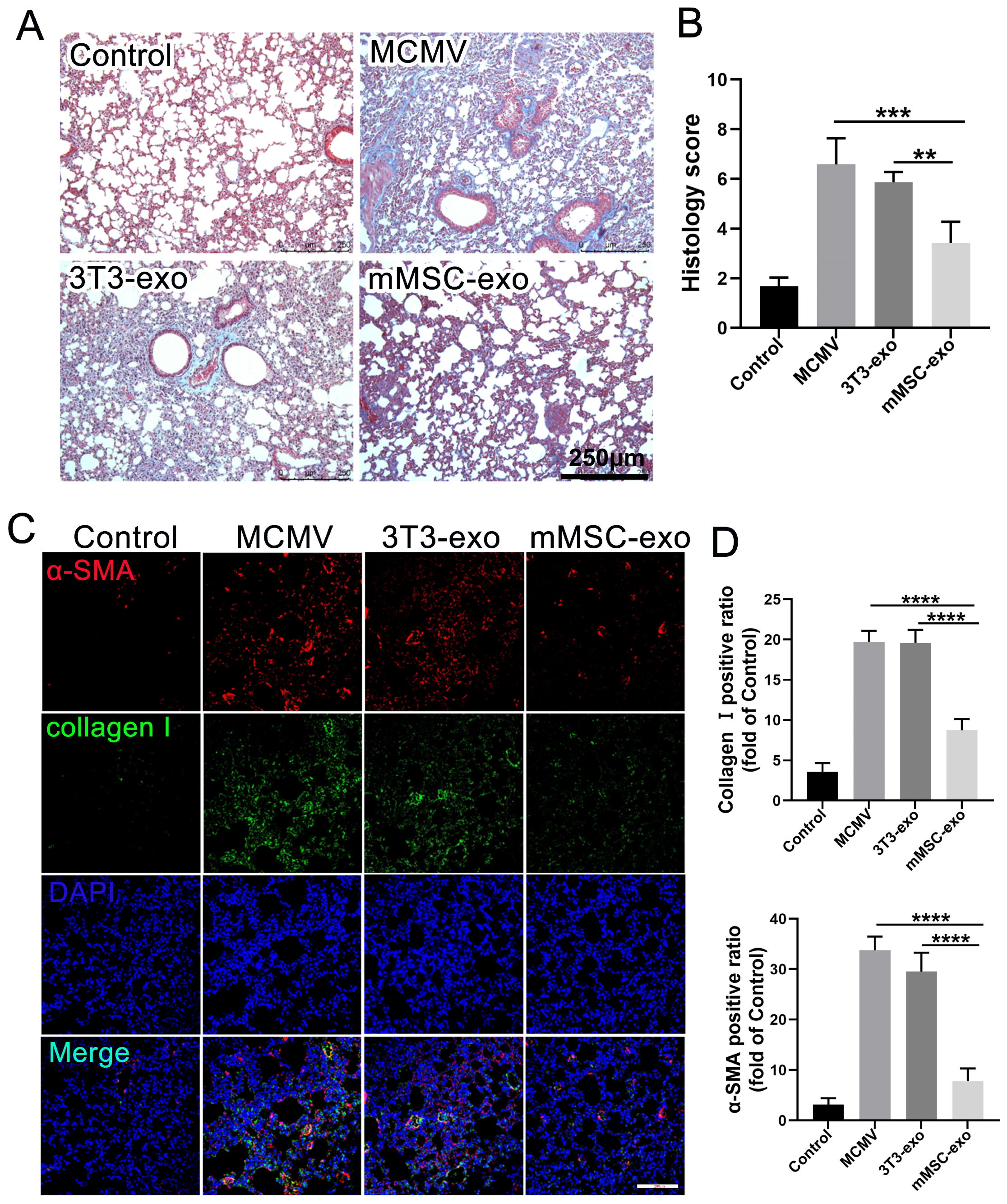
3.5. mMSC-exos Reduce Pulmonary Fibrosis in MCMV-Infected Mice
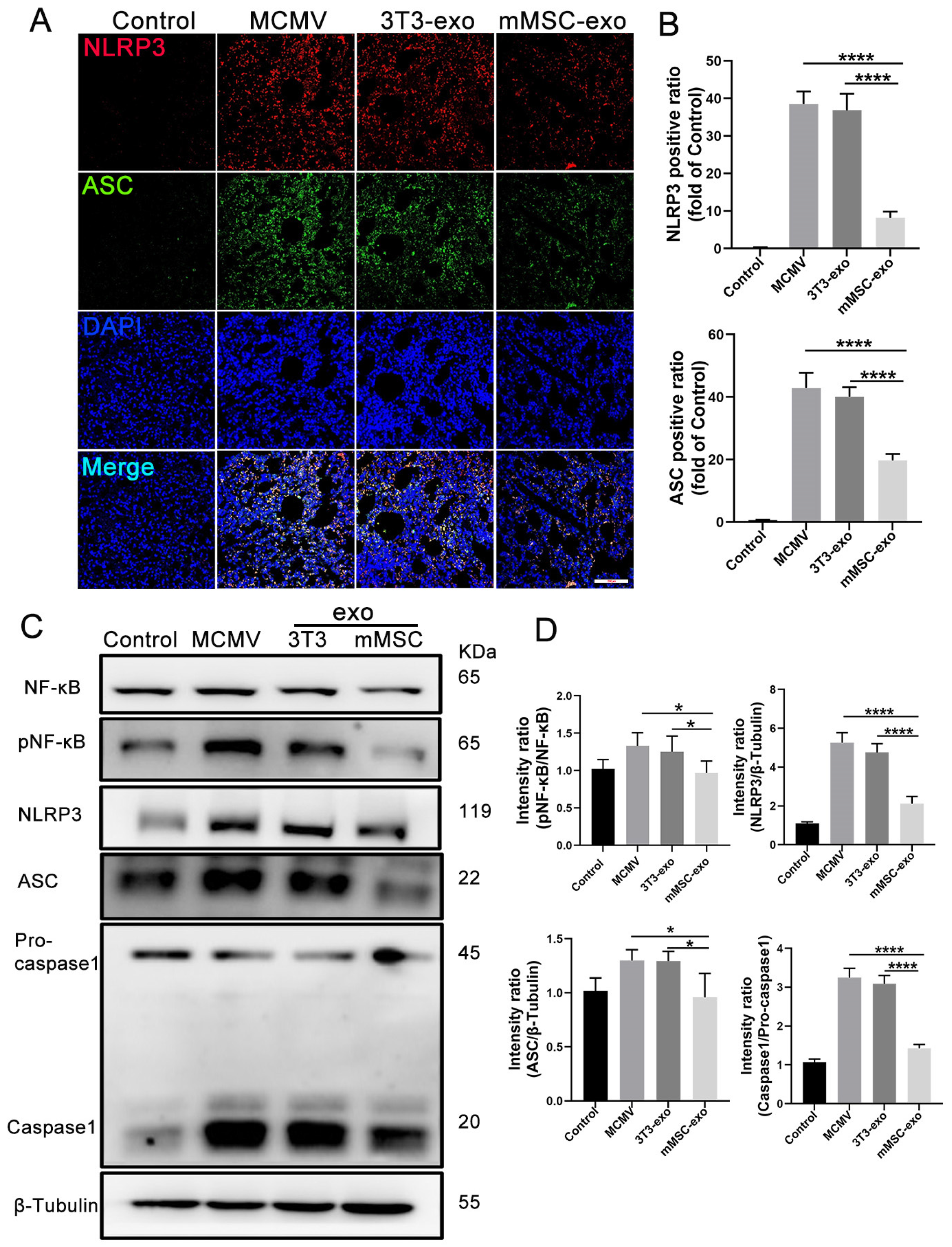
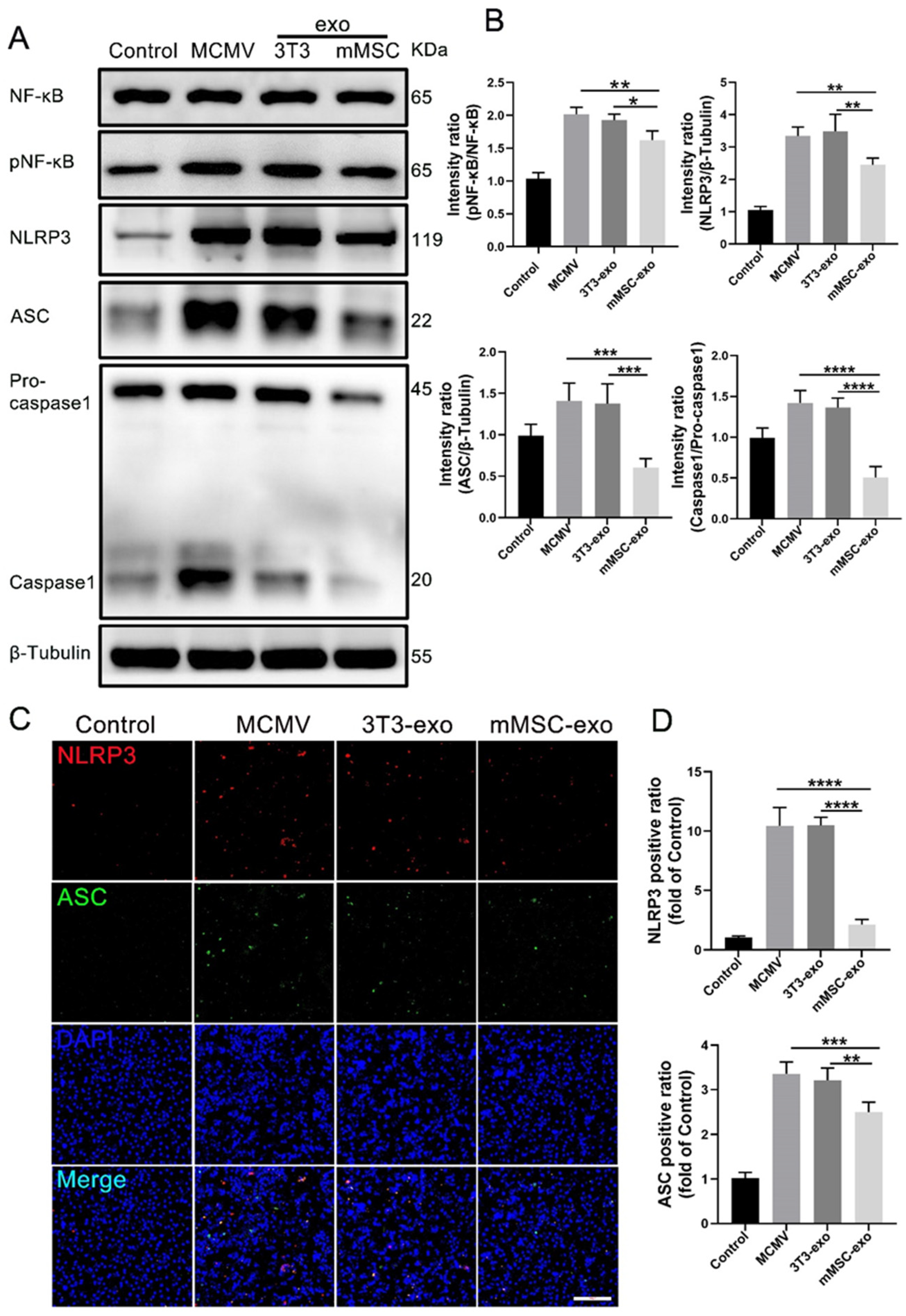
3.6. mMSC-exos Inhibit NF-κB and NLRP3 Signaling Pathways
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pittenger, M.F.; Discher, D.E.; Péault, B.M.; Phinney, D.G.; Hare, J.M.; Caplan, A.I. Mesenchymal stem cell perspective: Cell biology to clinical progress. npj Regen. Med. 2019, 4, 22. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, G.M.; Horak, D.A.; Niland, J.C.; Duncan, S.R.; Forman, S.J.; Zaia, J.A. A Randomized, Controlled Trial of Prophylactic Ganciclovir for Cytomegalovirus Pulmonary Infection in Recipients of Allogeneic Bone Marrow Transplants. N. Engl. J. Med. 1991, 324, 1005–1011. [Google Scholar] [CrossRef] [PubMed]
- Cesaro, S.; Ljungman, P.; Tridello, G.; Mikulska, M.; Wendel, L.; Styczynski, J.; Averbuch, D.; de la Camara, R. New trends in the management of cytomegalovirus infection after allogeneic hematopoietic cell transplantation: A survey of the Infectious Diseases Working Pary of EBMT. Bone Marrow Transplant. 2023, 58, 203–208. [Google Scholar] [CrossRef]
- Redeker, A.; Remmerswaal, E.B.M.; van der Gracht, E.T.I.; Welten, S.P.M.; Höllt, T.; Koning, F.; Cicin-Sain, L.; Nikolich-Žugich, J.; Berge, I.J.M.T.; van Lier, R.A.W.; et al. The Contribution of Cytomegalovirus Infection to Immune Senescence Is Set by the Infectious Dose. Front. Immunol. 2018, 8, 1953. [Google Scholar] [CrossRef] [PubMed]
- Fisher, M.A.; Lloyd, M.L. A Review of Murine Cytomegalovirus as a Model for Human Cytomegalovirus Disease—Do Mice Lie? Int. J. Mol. Sci. 2021, 22, 214. [Google Scholar] [CrossRef] [PubMed]
- Restrepo-Gualteros, S.M.; Gutierrez, M.J.; Villamil-Osorio, M.; Arroyo, M.A.; Nino, G. Challenges and Clinical Implications of the Diagnosis of Cytomegalovirus Lung Infection in Children. Curr. Infect. Dis. Reports 2019, 21, 24. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Cai, J.; Wang, X.; Lu, Y.; Guo, B.; Lai, M.; Lan, L.; Peng, Y.; Zheng, X. Human cytomegalovirus infection activates NLRP3 inflammasome by releasing mtDNA into the cytosol in human THP-1 cells. Microbiol. Immunol. 2023, 67, 303–313. [Google Scholar] [CrossRef]
- DeMeritt, I.B.; Milford, L.E.; Yurochko, A.D. Activation of the NF-κB Pathway in Human Cytomegalovirus-Infected Cells Is Necessary for Efficient Transactivation of the Major Immediate-Early Promoter. J. Virol. 2004, 78, 4498–4507. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Jin, X.; Yu, D.; Liu, G. Suppression of NLRP3 and NF-κB signaling pathways by α-Cyperone via activating SIRT1 contributes to attenuation of LPS-induced acute lung injury in mice. Int. Immunopharmacol. 2019, 76, 105886. [Google Scholar] [CrossRef]
- Hunninghake, G.W.; Monick, M.M.; Geist, L.J. Cytomegalovirus Infection. Am. J. Respir. Cell Mol. Biol. 1999, 21, 150–152. [Google Scholar] [CrossRef]
- Monsel, A.; Zhu, Y.G.; Gennai, S.; Hao, Q.; Hu, S.; Rouby, J.J.; Rosenzwajg, M.; Matthay, M.A.; Lee, J.W. Therapeutic Effects of Human Mesenchymal Stem Cell-derived Microvesicles in Severe Pneumonia in Mice. Am. J. Respir. Crit. Care Med. 2015, 192, 324–336. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Kim, S.; Lim, H.; Liu, A.; Hu, S.; Lee, J.; Zhuo, H.; Hao, Q.; Matthay, M.A.; Lee, J.W. Therapeutic effects of human mesenchymal stem cell microvesicles in an ex vivo perfused human lung injured with severe E. coli pneumonia. Thorax 2019, 74, 43–50. [Google Scholar] [CrossRef]
- Chu, M.; Wang, H.; Bian, L.; Huang, J.; Wu, D.; Zhang, R.; Fei, F.; Chen, Y.; Xia, J. Nebulization Therapy with Umbilical Cord Mesenchymal Stem Cell-Derived Exosomes for COVID-19 Pneumonia. Stem Cell Rev. Rep. 2022, 18, 2152–2163. [Google Scholar] [CrossRef]
- Hao, Q.; Gudapati, V.; Monsel, A.; Park, J.H.; Hu, S.; Kato, H.; Lee, J.H.; Zhou, L.; He, H. Mesenchymal Stem Cell-Derived Extracellular Vesicles Decrease Lung Injury in Mice. J. Immunol. 2019, 203, 1961–1972. [Google Scholar] [CrossRef]
- Zhang, X.; Cheng, Z.; Zeng, M.; He, Z. The efficacy of extracellular vesicles for acute lung injury in preclinical animal models: A meta-analysis. BMC Pulm. Med. 2023, 23, 40. [Google Scholar] [CrossRef] [PubMed]
- Abraham, A.; Krasnodembskaya, A. Mesenchymal stem cell-derived extracellular vesicles for the treatment of acute respiratory distress syndrome. Stem Cells Transl. Med. 2020, 9, 28–38. [Google Scholar] [CrossRef]
- Ridzuan, N.; Zakaria, N.; Widera, D.; Sheard, J.; Morimoto, M.; Kiyokawa, H.; Isa, S.A.M.; Singh, G.K.C.; Then, K.-Y.; Ooi, G.-C.; et al. Human umbilical cord mesenchymal stem cell-derived extracellular vesicles ameliorate airway inflammation in a rat model of chronic obstructive pulmonary disease (COPD). Stem Cell Res. Ther. 2021, 12, 54. [Google Scholar] [CrossRef]
- Mo, Y.; Kim, Y.; Bang, J.-Y.; Jung, J.; Lee, C.-G.; Elias, J.A.; Kang, H.-R. Mesenchymal Stem Cells Attenuate Asthmatic Inflammation and Airway Remodeling by Modulating Macrophages/Monocytes in the IL-13-Overexpressing Mouse Model. Immune Netw. 2022, 22, e40. [Google Scholar] [CrossRef] [PubMed]
- Kletukhina, S.; Mutallapova, G.; Titova, A.; Gomzikova, M. Role of Mesenchymal Stem Cells and Extracellular Vesicles in Idiopathic Pulmonary Fibrosis. Int. J. Mol. Sci. 2022, 23, 11212. [Google Scholar] [CrossRef]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef]
- Di Bella, M.A. Overview and Update on Extracellular Vesicles: Considerations on Exosomes and Their Application in Modern Medicine. Biology 2022, 11, 804. [Google Scholar] [CrossRef]
- Phinney, D.G.; Pittenger, M.F. Concise Review: MSC-Derived Exosomes for Cell-Free Therapy. Stem Cells 2017, 35, 851–858. [Google Scholar] [CrossRef] [PubMed]
- Ashcroft, T.; Simpson, J.M.; Timbrell, V. Simple method of estimating severity of pulmonary fibrosis on a numerical scale. J. Clin. Pathol. 1988, 41, 467. [Google Scholar] [CrossRef] [PubMed]
- Gilda, J.E.; Gomes, A.V. Stain-Free total protein staining is a superior loading control to β-actin for Western blots. Anal. Biochem. 2013, 440, 186–188. [Google Scholar] [CrossRef]
- Grundy, J.E.; Lawson, K.M.; MacCormac, L.P.; Fletcher, J.M.; Yong, K.L. Cytomegalovirus-Infected Endothelial Cells Recruit Neutrophils by the Secretion of C-X-C Chemokines and Transmit Virus by Direct Neutrophil-Endothelial Cell Contact and during Neutrophil Transendothelial Migration. J. Infect. Dis. 1998, 177, 1465–1474. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, M.; Shnayder, M.; Nachshon, A.; Arazi, T.; Kitsberg, Y.; Samia, R.L.; Lavi, M.; Kuint, R.; Tsabari, R.; Stern-Ginossar, N. Molecular characterization of human cytomegalovirus infection with single-cell transcriptomics. Nat. Microbiol. 2023, 8, 455–468. [Google Scholar] [CrossRef] [PubMed]
- Austermann, J.; Roth, J.; Barczyk-Kahlert, K. The Good and the Bad: Monocytes’ and Macrophages’ Diverse Functions in Inflammation. Cells 2022, 11, 1979. [Google Scholar] [CrossRef]
- Ti, D.; Hao, H.; Tong, C.; Liu, J.; Dong, L.; Zheng, J.; Zhao, Y.; Liu, H.; Fu, X.; Han, W. LPS-preconditioned mesenchymal stromal cells modify macrophage polarization for resolution of chronic inflammation via exosome-shuttled let-7b. J. Transl. Med. 2015, 13, 308. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, R.; Chen, Y.; Zhang, J.; Zheng, Y.; Xu, M.; Liang, J.; Li, J.; Huang, Y.; Xu, Y.; et al. Association between active cytomegalovirus infection and lung fibroproliferation in adult patients with acute respiratory distress syndrome: A retrospective study. BMC Infect. Dis. 2022, 22, 788. [Google Scholar] [CrossRef] [PubMed]
- Pocock, J.M.; Storisteanu, D.M.; Reeves, M.B.; Juss, J.K.; Wills, M.R.; Cowburn, A.S.; Chilvers, E.R. Human Cytomegalovirus Delays Neutrophil Apoptosis and Stimulates the Release of a Prosurvival Secretome. Front. Immunol. 2017, 8, 1185. [Google Scholar] [CrossRef]
- Fonseca Brito, L.; Brune, W.; Stahl, F.R. Cytomegalovirus (CMV) Pneumonitis: Cell Tropism, Inflammation, and Immunity. Int. J. Mol. Sciences. 2019, 20, 3865. [Google Scholar] [CrossRef] [PubMed]
- Marandu, T.F.; Dombek, M.; Gutknecht, M.; Griessl, M.; Riça, I.G.; Vlková, B.; Macáková, K.; Panagioti, E.; Griffith, A.; Lederer, J.; et al. Cytomegalovirus durably primes neutrophil oxidative burst. J. Leukoc. Biol. 2023, 114, 459–474. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Sun, P.; Qin, Y.; Wang, X.J.; Wang, M.; Lin, Y.; Zhou, R.; Hu, W.; Liu, Q.; Yu, X.; et al. Disulfiram attenuates MCMV-Induced pneumonia by inhibition of NF-κB/NLRP3 signaling pathway in immunocompromised mice. Int. Immunopharmacol. 2022, 103, 108453. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Inoue, M.; Osawa, R.; Wagener, M.M.; Shinohara, M.L. Inflammasome expression and cytomegalovirus viremia in critically ill patients with sepsis. J. Clin. Virol. 2017, 93, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Xu, R.; Wang, S.; Xu, Z.; Zhang, C.; Li, Y.; Yang, T.; Shi, L.; Fu, J.; Jiang, T.; et al. Human umbilical cord-derived mesenchymal stem cell therapy in patients with COVID-19: A phase 1 clinical trial. Signal Transduct. Target. Ther. 2020, 5, 172. [Google Scholar]
- Atluri, S.; Manchikanti, L.; Hirsch, J.A. Expanded Umbilical Cord Mesenchymal Stem Cells (UC-MSCs) as a Therapeutic Strategy in Managing Critically Ill COVID-19 Patients: The Case for Compassionate Use. Pain. Physician 2020, 23, E71–E83. [Google Scholar] [PubMed]
- Kourembanas, S. Exosomes: Vehicles of Intercellular Signaling, Biomarkers, and Vectors of Cell Therapy. Annu. Rev. Physiol. 2015, 77, 13–27. [Google Scholar] [PubMed]
- Burrello, J.; Monticone, S.; Gai, C.; Gomez, Y.; Kholia, S.; Camussi, G. Stem Cell-Derived Extracellular Vesicles and Immune-Modulation. Front. Cell Dev. Biol. 2016, 4, 83. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, V.; Sengupta, S.; Lazo, A.; Woods, P.; Nolan, A.; Bremer, N. Exosomes Derived from Bone Marrow Mesenchymal Stem Cells as Treatment for Severe COVID-19. Stem Cells Dev. 2020, 29, 747–754. [Google Scholar] [CrossRef]
- Akbari, A.; Rezaie, J. Potential therapeutic application of mesenchymal stem cell-derived exosomes in SARS-CoV-2 pneumonia. Stem Cell Res. Ther. 2020, 11, 356. [Google Scholar] [CrossRef]
- Valiukevičius, P.; Mačiulaitis, J.; Pangonytė, D.; Siratavičiūtė, V.; Kluszczyńska, K.; Kuzaitytė, U.; Insodaitė, R.; Čiapienė, I.; Grigalevičiūtė, R.; Zigmantaitė, V.; et al. Human Placental Mesenchymal Stem Cells and Derived Extracellular Vesicles Ameliorate Lung Injury in Acute Respiratory Distress Syndrome Murine Model. Cells 2023, 12, 2729. [Google Scholar] [CrossRef] [PubMed]
- Willis, G.R.; Fernandez-Gonzalez, A.; Anastas, J.; Vitali, S.H.; Liu, X.; Ericsson, M.; Kwong, A.; Mitsialis, S.A.; Kourembanas, S. Mesenchymal Stromal Cell Exosomes Ameliorate Experimental Bronchopulmonary Dysplasia and Restore Lung Function through Macrophage Immunomodulation. Am. J. Respir. Crit. Care Med. 2018, 197, 104–116. [Google Scholar] [CrossRef]
- Oh, J.Y.; Ko, J.H.; Lee, H.J.; Yu, J.M.; Choi, H.; Kim, M.K.; Wee, W.R.; Prockop, D.J. Mesenchymal Stem/Stromal Cells Inhibit the NLRP3 Inflammasome by Decreasing Mitochondrial Reactive Oxygen Species. Stem Cells. 2014, 32, 1553–1563. [Google Scholar] [CrossRef] [PubMed]
- Harrell, C.R.; Djonov, V.; Antonijevic, A.; Volarevic, V. NLRP3 Inflammasome as a Potentially New Therapeutic Target of Mesenchymal Stem Cells and Their Exosomes in the Treatment of Inflammatory Eye Diseases. Cells 2023, 12, 2327. [Google Scholar] [CrossRef] [PubMed]
- Sarsenova, M.; Kim, Y.; Raziyeva, K.; Kazybay, B.; Ogay, V.; Saparov, A. Recent advances to enhance the immunomodulatory potential of mesenchymal stem cells. Front. Immunol. 2022, 13, 1010399. [Google Scholar] [CrossRef]
- Wang, H.; Pan, J.Q.; Luo, L.; Ning, X.J.; Ye, Z.P.; Yu, Z.; Li, W.S. NF-κB induces miR-148a to sustain TGF-β/Smad signaling activation in glioblastoma. Mol. Cancer 2015, 14, 2. [Google Scholar] [CrossRef]
- Miteva, K.; Pappritz, K.; Sosnowski, M.; El-Shafeey, M.; Müller, I.; Dong, F.; Savvatis, K.; Ringe, J.; Tschöpe, C.; Van Linthout, S. Mesenchymal stromal cells inhibit NLRP3 inflammasome activation in a model of Coxsackievirus B3-induced inflammatory cardiomyopathy. Sci. Rep. 2018, 8, 2820. [Google Scholar] [CrossRef]
- Khatri, M.; Richardson, L.A.; Meulia, T. Mesenchymal stem cell-derived extracellular vesicles attenuate influenza virus-induced acute lung injury in a pig model. Stem Cell Res. Ther. 2018, 9, 17. [Google Scholar] [CrossRef]
- Tang, Y.; Zhou, Y.; Li, H.-J. Advances in mesenchymal stem cell exosomes: A review. Stem Cell Res. Ther. 2021, 12, 71. [Google Scholar] [CrossRef]
- Stacey, M.A.; Marsden, M.; Clare, S.; Dolton, G.; Stack, G.; Jones, E.; Klenerman, P.; Gallimore, A.M.; Taylor, P.R.; Snelgrove, R.J.; et al. Neutrophils Recruited by IL-22 in Peripheral Tissues Function as TRAIL-Dependent Antiviral Effectors against, MCMV. Cell Host Microbe 2014, 15, 471–483. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, F.; Chen, Z.; Wu, H.-T.; Chen, X.-X.; Zhan, P.; Wei, Z.-Y.; Ouyang, Z.; Jiang, X.; Shen, A.; Luo, M.-H.; et al. Mesenchymal Stem Cell-Derived Exosomes Attenuate Murine Cytomegalovirus-Infected Pneumonia via NF-κB/NLRP3 Signaling Pathway. Viruses 2024, 16, 619. https://doi.org/10.3390/v16040619
Chen F, Chen Z, Wu H-T, Chen X-X, Zhan P, Wei Z-Y, Ouyang Z, Jiang X, Shen A, Luo M-H, et al. Mesenchymal Stem Cell-Derived Exosomes Attenuate Murine Cytomegalovirus-Infected Pneumonia via NF-κB/NLRP3 Signaling Pathway. Viruses. 2024; 16(4):619. https://doi.org/10.3390/v16040619
Chicago/Turabian StyleChen, Fei, Zhida Chen, Hui-Ting Wu, Xin-Xiang Chen, Peiqi Zhan, Zheng-Yi Wei, Zizhang Ouyang, Xueyan Jiang, Ao Shen, Min-Hua Luo, and et al. 2024. "Mesenchymal Stem Cell-Derived Exosomes Attenuate Murine Cytomegalovirus-Infected Pneumonia via NF-κB/NLRP3 Signaling Pathway" Viruses 16, no. 4: 619. https://doi.org/10.3390/v16040619
APA StyleChen, F., Chen, Z., Wu, H.-T., Chen, X.-X., Zhan, P., Wei, Z.-Y., Ouyang, Z., Jiang, X., Shen, A., Luo, M.-H., Liu, Q., Zhou, Y.-P., & Qin, A. (2024). Mesenchymal Stem Cell-Derived Exosomes Attenuate Murine Cytomegalovirus-Infected Pneumonia via NF-κB/NLRP3 Signaling Pathway. Viruses, 16(4), 619. https://doi.org/10.3390/v16040619