

Antibodies Induced by Smallpox Vaccination after at Least 45 Years Cross-React with and In Vitro Neutralize Mpox Virus: A Role for Polyclonal B Cell Activation?
 , , , , , , ,
, , , , , , ,  , , , ,
, , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Viruses
2.3. ELISA
2.4. Neutralization Assay Plaque Reduction Neutralization Test (PRNT) for MPXV
2.5. SDS-PAGE and Western Blot Analysis
2.6. Monocytes Isolation, Dendritic Cells Generation and In Vitro Cell Infection
2.7. Antigen Presentation Assays
2.8. FACS Analysis
2.9. Statistical Analysis
3. Results
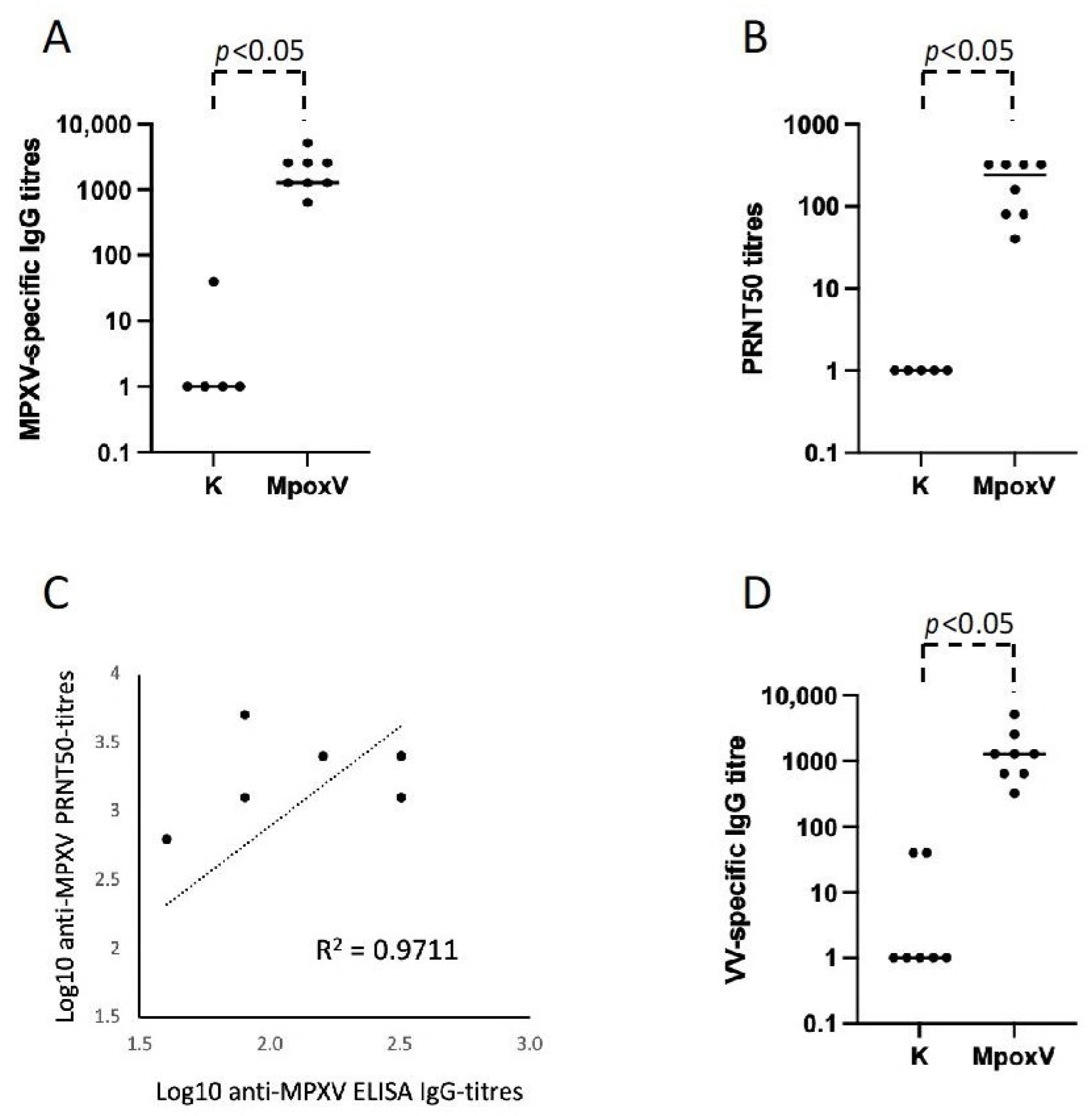
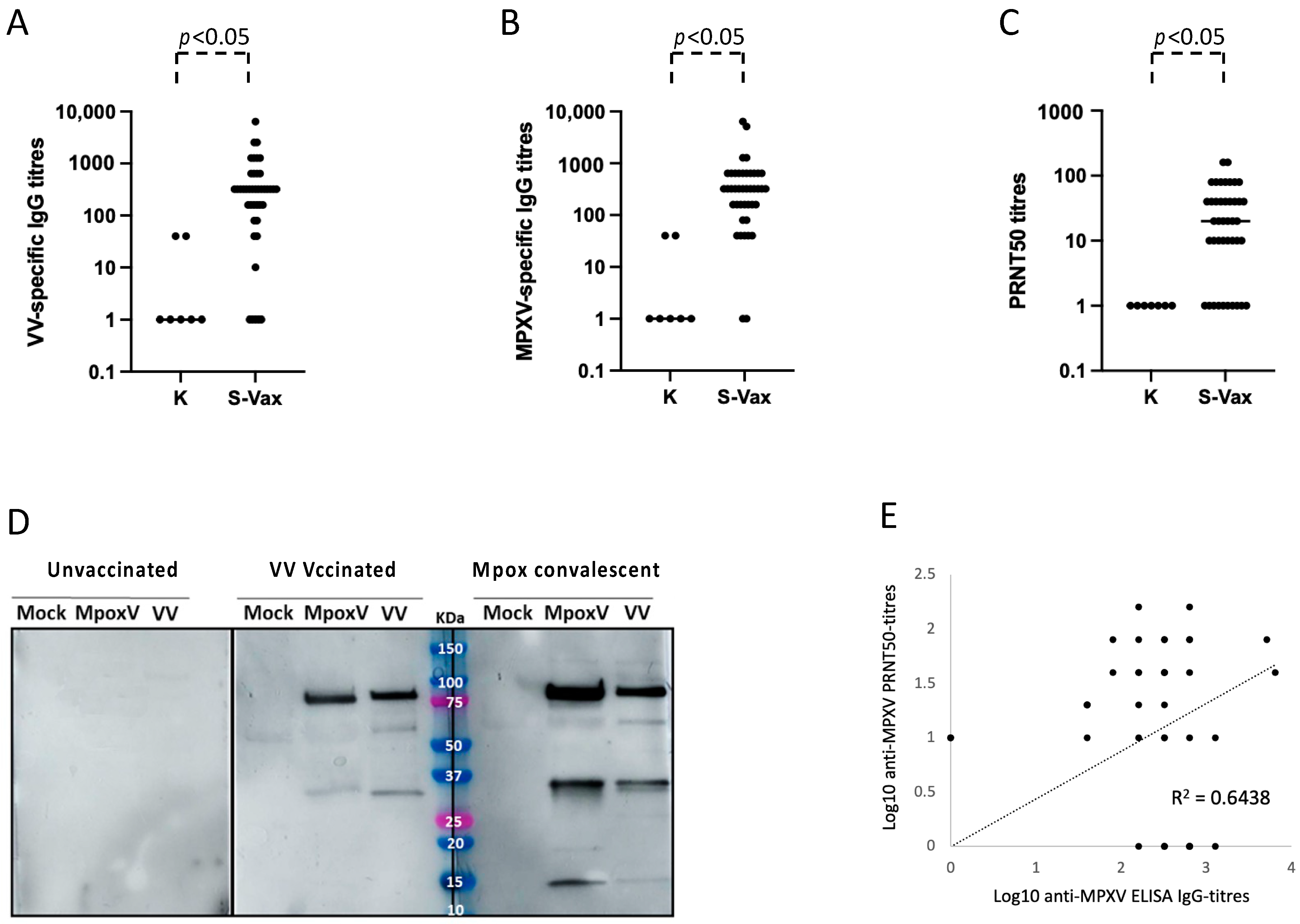
3.1. Development of ELISA Test and Neutralization Assay for the Serological Diagnosis of Mpox
3.2. Role of the Residual Immunological Smallpox IgG Memory Response in the Serological Diagnosis of Mpox
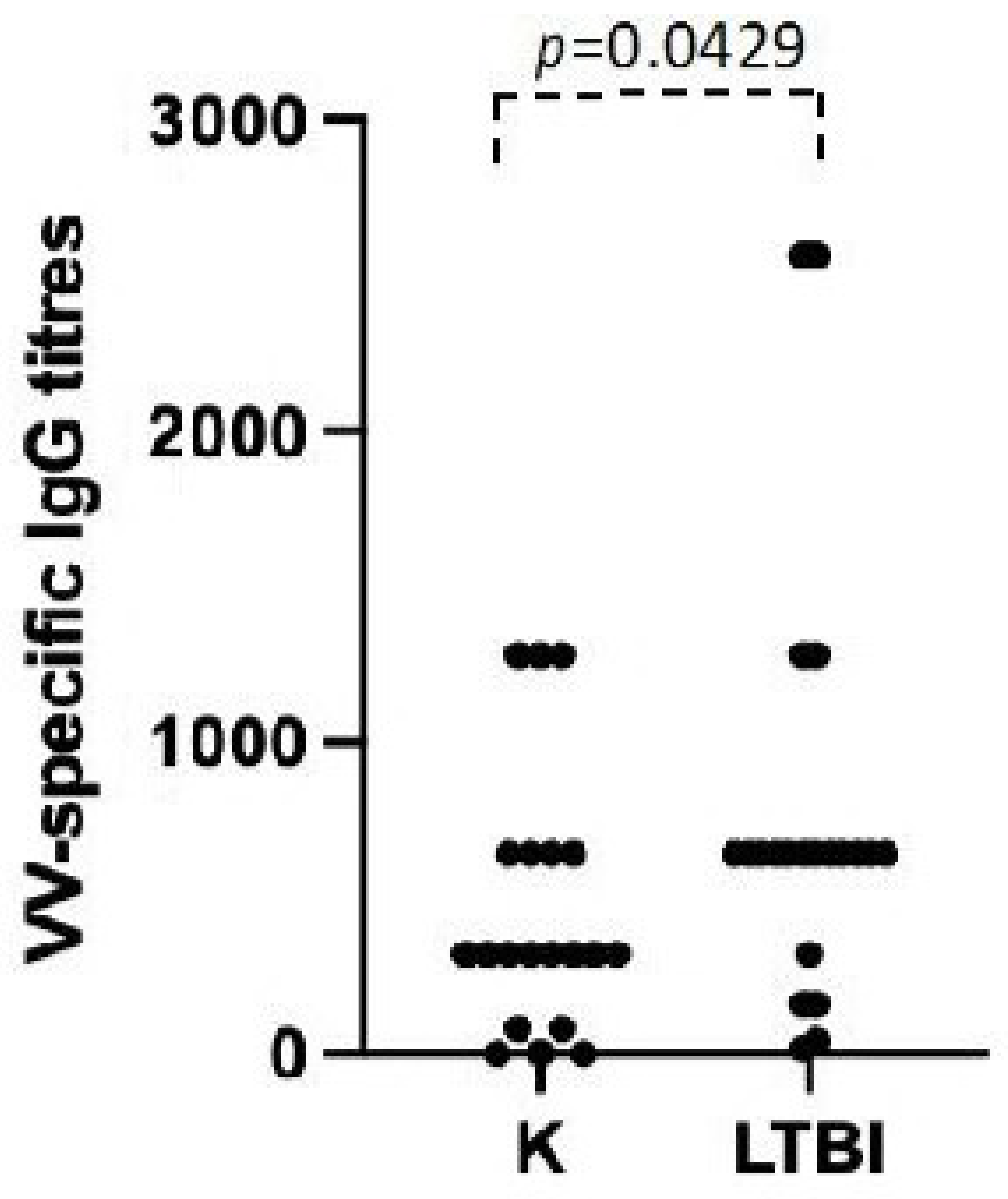
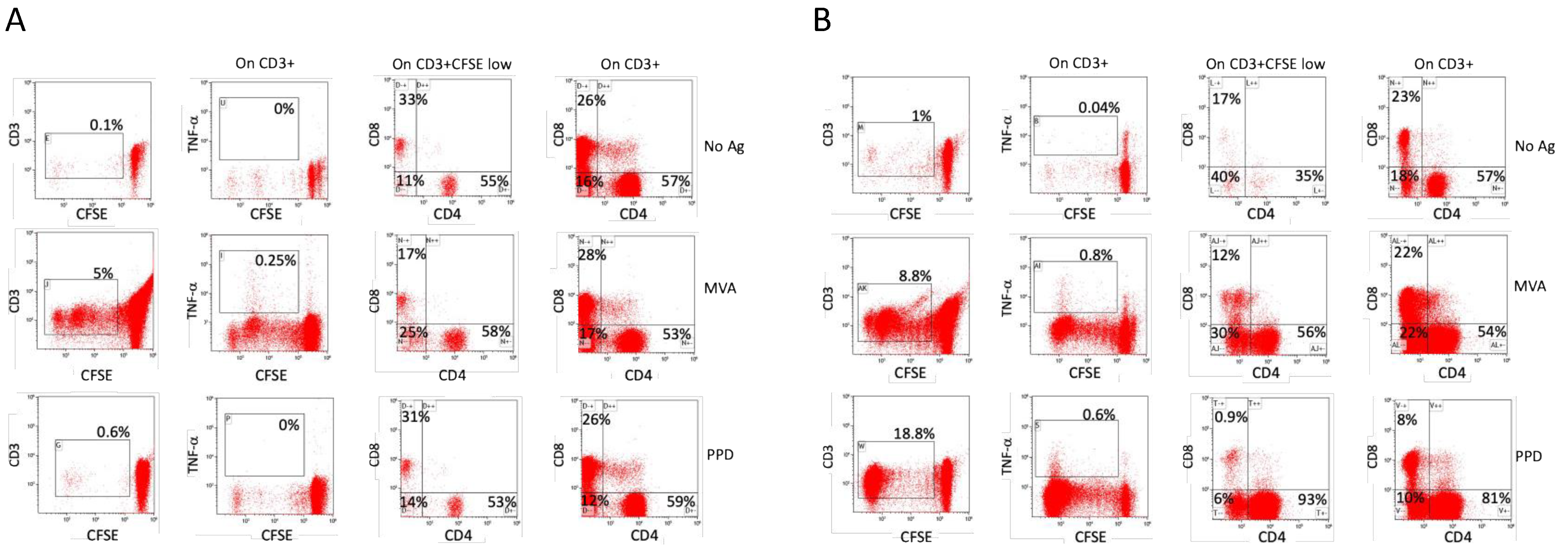
3.3. Possible Role of Polyclonal Non-Specific Activation in the Maintenance of Immunological Memory
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gessain, A.; Nakoune, E.; Yazdanpanah, Y. Monkeypox. N. Engl. J. Med. 2022, 387, 1783–1793. [Google Scholar] [CrossRef] [PubMed]
- Dubois, M.E.; Slifka, M.K. Retrospective Analysis of Monkeypox Infection. Emerg. Infect. Dis. 2008, 14, 592–599. [Google Scholar] [CrossRef] [PubMed]
- Elsayed, S.; Bondy, L.; Hanage, W.P. Monkeypox Virus Infections in Humans. Clin. Microbiol. Rev. 2022, 35, e0009222. [Google Scholar] [CrossRef] [PubMed]
- Doty, J.B.; Malekani, J.M.; Kalemba, L.N.; Stanley, W.T.; Monroe, B.P.; Nakazawa, Y.U.; Mauldin, M.R.; Bakambana, T.L.; Liyandja Dja Liyandja, T.; Braden, Z.H.; et al. Assessing Monkeypox Virus Prevalence in Small Mammals at the Human-Animal Interface in the Democratic Republic of the Congo. Viruses 2017, 9, 283. [Google Scholar] [CrossRef]
- Silva, S.; Kohl, A.; Pena, L.; Pardee, K. Clinical and Laboratory Diagnosis of Monkeypox (mpox): Current Status and Future Directions. iScience 2023, 26, 106759. [Google Scholar] [CrossRef] [PubMed]
- Thornhill, J.P.; Barkati, S.; Walmsley, S.; Rockstroh, J.; Antinori, A.; Harrison, L.B.; Palich, R.; Nori, A.; Reeves, I.; Habibi, M.S.; et al. Monkeypox Virus Infection in Humans across 16 Countries—April–June 2022. N. Engl. J. Med. 2022, 387, 679–691. [Google Scholar] [CrossRef] [PubMed]
- WHO. WHO Director-General Declares the Ongoing Monkeypox Outbreak a Public Health Emergency of International Concern. 2022. Available online: https://www.who.int/europe/news/item/23-07-2022-who-director-general-declares-the-ongoing-monkeypox-outbreak-a-public-health-event-of-international-concern (accessed on 10 January 2024).
- Hazra, A.; Zucker, J.; Bell, E.; Flores, J.; Gordon, L.; Mitja, O.; Suner, C.; Lemaignen, A.; Jamard, S.; Nozza, S.; et al. Mpox in People with Past Infection or a Complete Vaccination Course: A Global Case Series. Lancet Infect. Dis. 2024, 24, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.F.; Sohail, M.S.; Quadeer, A.A.; McKay, M.R. Vaccinia-Virus-Based Vaccines Are Expected to Elicit Highly Cross-Reactive Immunity to the 2022 Monkeypox Virus. Viruses 2022, 14, 1960. [Google Scholar] [CrossRef]
- Poland, G.A.; Kennedy, R.B.; Tosh, P.K. Prevention of Monkeypox with Vaccines: A Rapid Review. Lancet Infect. Dis. 2022, 22, e349–e358. [Google Scholar] [CrossRef] [PubMed]
- Riccardo, V.; Pablo, G.C. Neutralization Determinants on Poxviruses. Viruses 2023, 15, 2396. [Google Scholar] [CrossRef] [PubMed]
- Shafaati, M.; Zandi, M. Human Monkeypox (hMPXV) Re-Emergence: Host Immunity Status and Current Vaccines Landscape. J. Med. Virol. 2023, 95, e28251. [Google Scholar] [CrossRef] [PubMed]
- Bernasconi, N.L.; Traggiai, E.; Lanzavecchia, A. Maintenance of Serological Memory by Polyclonal Activation of Human Memory B Cells. Science 2002, 298, 2199–2202. [Google Scholar] [CrossRef] [PubMed]
- Mazurek, G.H.; Jereb, J.; Vernon, A.; LoBue, P.; Goldberg, S.; Castro, K.; Igra Expert Committee; Control Centers for Disease, and Prevention. Updated Guidelines for Using Interferon Gamma Release Assays to Detect Mycobacterium tuberculosis Infection—United States, 2010. MMWR Recomm. Rep. 2010, 59, 1–25. [Google Scholar] [PubMed]
- Accardi, L.; Prehaud, C.; Di Bonito, P.; Mochi, S.; Bouloy, M.; Giorgi, C. Activity of Toscana and Rift Valley Fever Virus Transcription Complexes on Heterologous Templates. J. Gen. Virol. 2001, 82, 781–785. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ferlito, C.; Biselli, R.; Cattaruzza, M.S.; Teloni, R.; Mariotti, S.; Tomao, E.; Salerno, G.; Peragallo, M.S.; Lulli, P.; Caporuscio, S.; et al. Immunogenicity of Meningococcal Polysaccharide ACWY Vaccine in Primary Immunized or Revaccinated Adults. Clin. Exp. Immunol. 2018, 194, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Titanji, B.K.; Eick-Cost, A.; Partan, E.S.; Epstein, L.; Wells, N.; Stahlman, S.L.; Devineni, P.; Munyoki, B.; Pyarajan, S.; Balajee, A.; et al. Effectiveness of Smallpox Vaccination to Prevent Mpox in Military Personnel. N. Engl. J. Med. 2023, 389, 1147–1148. [Google Scholar] [CrossRef] [PubMed]
- Krieg, A.M. CpG Motifs in Bacterial DNA and Their Immune Effects. Annu. Rev. Immunol. 2002, 20, 709–760. [Google Scholar] [CrossRef] [PubMed]
- Poltorak, A.; He, X.; Smirnova, I.; Liu, M.Y.; Van Huffel, C.; Du, X.; Birdwell, D.; Alejos, E.; Silva, M.; Galanos, C.; et al. Defective LPS Signaling in C3H/HeJ and C57BL/10ScCr Mice: Mutations in Tlr4 Gene. Science 1998, 282, 2085–2088. [Google Scholar] [CrossRef] [PubMed]
- Banchereau, J.; Bazan, F.; Blanchard, D.; Briere, F.; Galizzi, J.P.; van Kooten, C.; Liu, Y.J.; Rousset, F.; Saeland, S. The CD40 Antigen and Its Ligand. Annu. Rev. Immunol. 1994, 12, 881–922. [Google Scholar] [CrossRef] [PubMed]
- Barry, C.E., 3rd; Boshoff, H.I.; Dartois, V.; Dick, T.; Ehrt, S.; Flynn, J.; Schnappinger, D.; Wilkinson, R.J.; Young, D. The Spectrum of Latent Tuberculosis: Rethinking the Biology and Intervention Strategies. Nat. Rev. Microbiol. 2009, 7, 845–855. [Google Scholar] [CrossRef] [PubMed]
- Veatch, A.V.; Kaushal, D. Opening Pandora’s Box: Mechanisms of Mycobacterium tuberculosis Resuscitation. Trends Microbiol. 2018, 26, 145–157. [Google Scholar] [CrossRef] [PubMed]
- Gengenbacher, M.; Kaufmann, S.H. Mycobacterium tuberculosis: Success through Dormancy. FEMS Microbiol. Rev. 2012, 36, 514–532. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PRNT50 Positives/Tested | PRNT50 Titers Mean ± SD (Median, Range) | PRNT80 Positives/Tested | PRNT80 Titers Mean ± SD (Median, Range) | |
|---|---|---|---|---|
| Vaccinated (N = 41) | 34/41 | 4.88 ± 1.28 (median 5.32, range 3.32–7.32) | 22/41 | 4.55 ± 0.75 (median 4.32, range 3.32–5.32) |
| Mpox convalescent patients (N = 8) | 8/8 | 7.32 ± 1.20 (median 7.82, range 5.32–8.32) | 8/8 | 5.20 ± 2.36 (median 3.82, range 3.32–8.32) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mariotti, S.; Venturi, G.; Chiantore, M.V.; Teloni, R.; De Santis, R.; Amendola, A.; Fortuna, C.; Marsili, G.; Grilli, G.; Lia, M.S.; et al. Antibodies Induced by Smallpox Vaccination after at Least 45 Years Cross-React with and In Vitro Neutralize Mpox Virus: A Role for Polyclonal B Cell Activation? Viruses 2024, 16, 620. https://doi.org/10.3390/v16040620
Mariotti S, Venturi G, Chiantore MV, Teloni R, De Santis R, Amendola A, Fortuna C, Marsili G, Grilli G, Lia MS, et al. Antibodies Induced by Smallpox Vaccination after at Least 45 Years Cross-React with and In Vitro Neutralize Mpox Virus: A Role for Polyclonal B Cell Activation? Viruses. 2024; 16(4):620. https://doi.org/10.3390/v16040620
Chicago/Turabian StyleMariotti, Sabrina, Giulietta Venturi, Maria Vincenza Chiantore, Raffaela Teloni, Riccardo De Santis, Antonello Amendola, Claudia Fortuna, Giulia Marsili, Giorgia Grilli, Maria Stella Lia, and et al. 2024. "Antibodies Induced by Smallpox Vaccination after at Least 45 Years Cross-React with and In Vitro Neutralize Mpox Virus: A Role for Polyclonal B Cell Activation?" Viruses 16, no. 4: 620. https://doi.org/10.3390/v16040620
APA StyleMariotti, S., Venturi, G., Chiantore, M. V., Teloni, R., De Santis, R., Amendola, A., Fortuna, C., Marsili, G., Grilli, G., Lia, M. S., Kiros, S. T., Lagi, F., Bartoloni, A., Iacobino, A., Cresta, R., Lastilla, M., Biselli, R., Di Bonito, P., Lista, F., & Nisini, R. (2024). Antibodies Induced by Smallpox Vaccination after at Least 45 Years Cross-React with and In Vitro Neutralize Mpox Virus: A Role for Polyclonal B Cell Activation? Viruses, 16(4), 620. https://doi.org/10.3390/v16040620