
T-Cell Responses to COVID-19 Vaccines and Breakthrough Infection in People Living with HIV Receiving Antiretroviral Therapy
 , , and add
Show full author list
, , and add
Show full author list
Abstract
:1. Introduction
2. Methods
3. Results
3.1. Participant Details
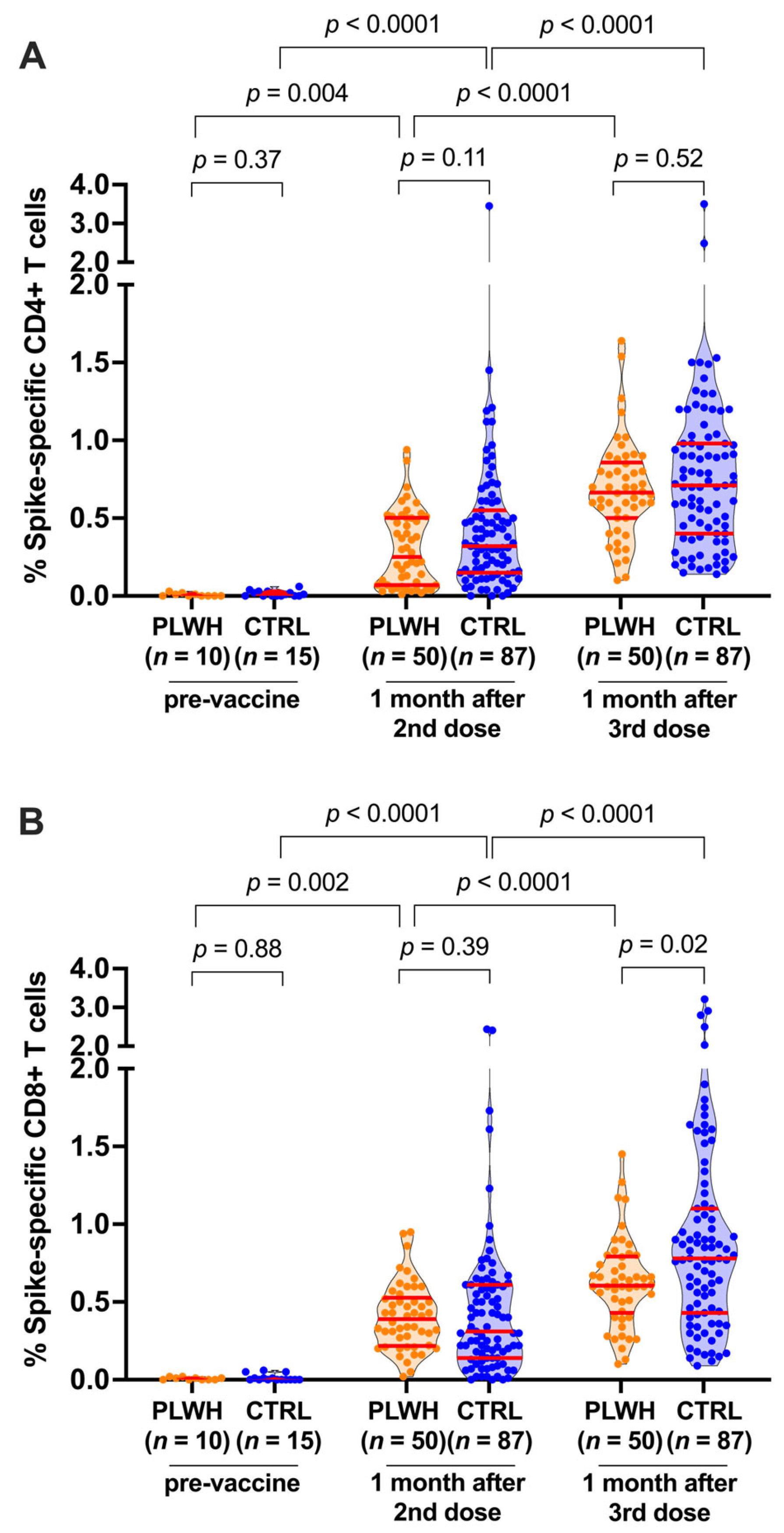
3.2. T-Cell Responses following Two and Three COVID-19 Vaccine Doses
3.3. Correlations between Vaccine-Induced T-Cell Responses and Other Immune Measures
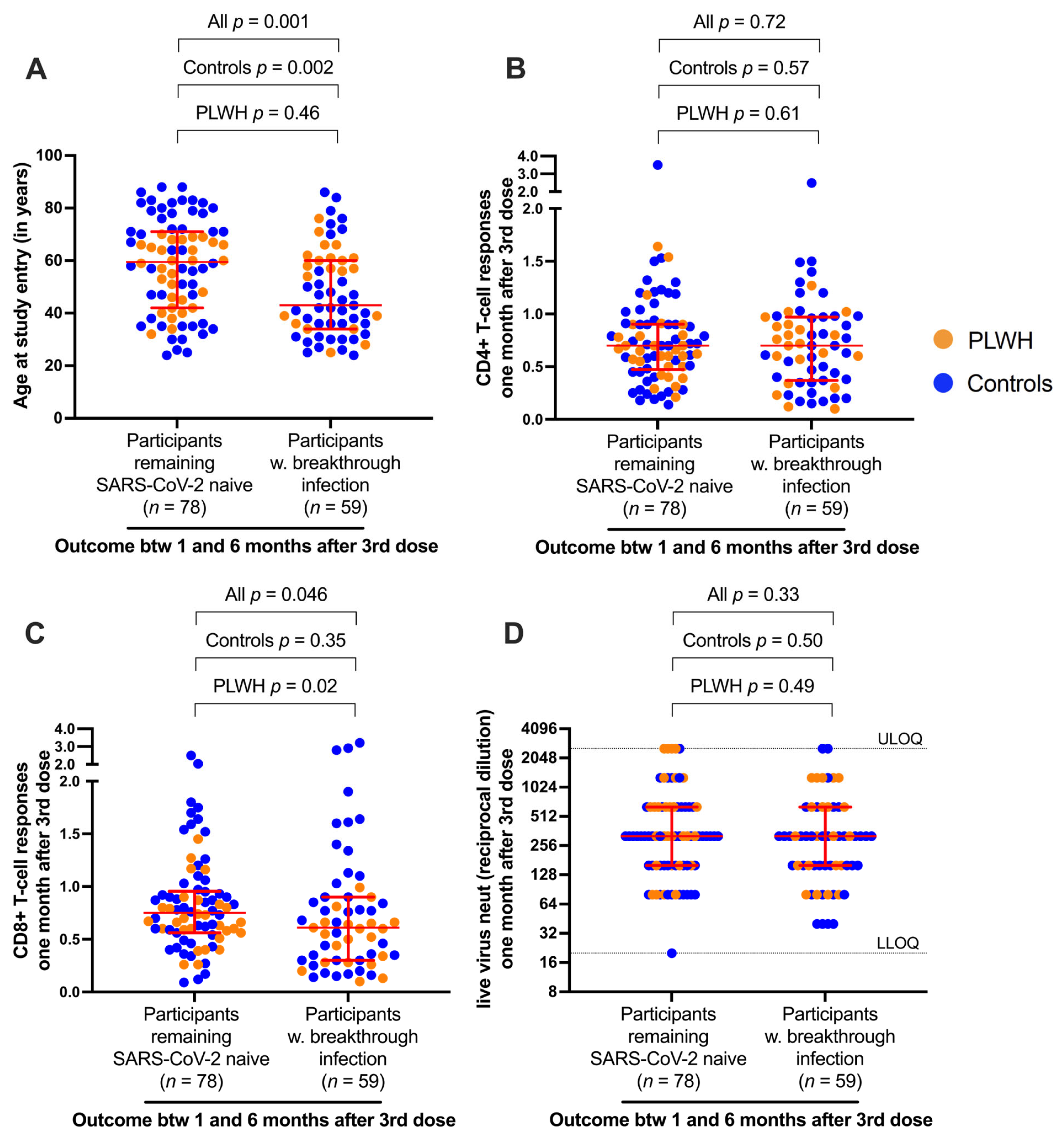
3.4. Breakthrough Infection Further Boosts CD4+ and CD8+ T-Cell Responses
3.5. Correlates of Protection against Breakthrough Infection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Western Cape Department of Health in collaboration with the National Institute for Communicable Diseases, South Africa. Risk Factors for Coronavirus Disease 2019 (COVID-19) Death in a Population Cohort Study from the Western Cape Province, South Africa. Clin. Infect. Dis. 2021, 73, e2005–e2015. [Google Scholar] [CrossRef] [PubMed]
- Geretti, A.M.; Stockdale, A.J.; Kelly, S.H.; Cevik, M.; Collins, S.; Waters, L.; Villa, G.; Docherty, A.; Harrison, E.M.; Turtle, L.; et al. Outcomes of COVID-19 related hospitalization among people with HIV in the ISARIC WHO Clinical Characterization Protocol (UK): A prospective observational study. Clin. Infect. Dis. 2021, 73, e2095–e2106. [Google Scholar] [CrossRef] [PubMed]
- Bhaskaran, K.; Rentsch, C.T.; MacKenna, B.; Schultze, A.; Mehrkar, A.; Bates, C.J.; Eggo, R.M.; Morton, C.E.; Bacon, S.C.J.; Inglesby, P.; et al. HIV infection and COVID-19 death: A population-based cohort analysis of UK primary care data and linked national death registrations within the OpenSAFELY platform. Lancet HIV 2021, 8, e24–e32. [Google Scholar] [CrossRef]
- Tesoriero, J.M.; Swain, C.E.; Pierce, J.L.; Zamboni, L.; Wu, M.; Holtgrave, D.R.; Gonzalez, C.J.; Udo, T.; Morne, J.E.; Hart-Malloy, R.; et al. COVID-19 Outcomes Among Persons Living with or without Diagnosed HIV Infection in New York State. JAMA Netw. Open 2021, 4, e2037069. [Google Scholar] [CrossRef] [PubMed]
- Spinelli, M.A.; Jones, B.L.H.; Gandhi, M. COVID-19 Outcomes and Risk Factors Among People Living with HIV. Curr. HIV/AIDS Rep. 2022, 19, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Chambers, C.; Samji, H.; Cooper, C.L.; Costiniuk, C.T.; Janjua, N.Z.; Kroch, A.E.; Arbess, G.; Benoit, A.C.; Buchan, S.A.; Chung, H.; et al. Coronavirus disease 2019 vaccine effectiveness among a population-based cohort of people living with HIV. AIDS 2022, 36, F17–F26. [Google Scholar] [CrossRef] [PubMed]
- Fowokan, A.; Samji, H.; Puyat, J.H.; Janjua, N.Z.; Wilton, J.; Wong, J.; Grennan, T.; Chambers, C.; Kroch, A.; Costiniuk, C.T.; et al. Effectiveness of COVID-19 vaccines in people living with HIV in British Columbia and comparisons with a matched HIV-negative cohort: A test-negative design. Int. J. Infect. Dis. 2023, 127, 162–170. [Google Scholar] [CrossRef]
- Madhi, S.A.; Koen, A.L.; Izu, A.; Fairlie, L.; Cutland, C.L.; Baillie, V.; Padayachee, S.D.; Dheda, K.; Barnabas, S.L.; Bhorat, Q.E.; et al. Safety and immunogenicity of the ChAdOx1 nCoV-19 (AZD1222) vaccine against SARS-CoV-2 in people living with and without HIV in South Africa: An interim analysis of a randomised, double-blind, placebo-controlled, phase 1B/2A trial. Lancet HIV 2021, 8, e568–e580. [Google Scholar] [CrossRef] [PubMed]
- Brumme, Z.L.; Mwimanzi, F.; Lapointe, H.R.; Cheung, P.K.; Sang, Y.; Duncan, M.C.; Yaseen, F.; Agafitei, O.; Ennis, S.; Ng, K.; et al. Humoral immune responses to COVID-19 vaccination in people living with HIV receiving suppressive antiretroviral therapy. NPJ Vaccines 2022, 7, 28. [Google Scholar] [CrossRef]
- Jedicke, N.; Stankov, M.V.; Cossmann, A.; Dopfer-Jablonka, A.; Knuth, C.; Ahrenstorf, G.; Ramos, G.M. Behrens GMN: Humoral immune response following prime and boost BNT162b2 vaccination in people living with HIV on antiretroviral therapy. HIV Med. 2022, 23, 558–563. [Google Scholar] [CrossRef]
- Lapointe, H.R.; Mwimanzi, F.; Cheung, P.K.; Sang, Y.; Yaseen, F.; Speckmaier, S.; Barad, E.; Moran-Garcia, N.; Datwani, S.; Duncan, M.C.; et al. Antibody response durability following three-dose COVID-19 vaccination in people with HIV receiving suppressive ART. AIDS 2023, 37, 709–721. [Google Scholar] [CrossRef] [PubMed]
- Lapointe, H.R.; Mwimanzi, F.; Cheung, P.K.; Sang, Y.; Yaseen, F.; Umviligihozo, G.; Kalikawe, R.; Speckmaier, S.; Moran-Garcia, N.; Datwani, S.; et al. People With Human Immunodeficiency Virus Receiving Suppressive Antiretroviral Therapy Show Typical Antibody Durability After Dual Coronavirus Disease 2019 Vaccination and Strong Third Dose Responses. J. Infect. Dis. 2023, 227, 838–849. [Google Scholar] [CrossRef] [PubMed]
- Nault, L.; Marchitto, L.; Goyette, G.; Tremblay-Sher, D.; Fortin, C.; Martel-Laferrière, V.; Trottier, B.; Richard, J.; Durand, M.; Kaufmann, D.; et al. COVID-19 vaccine immunogenicity in people living with HIV-1. Vaccine 2022, 40, 3633–3637. [Google Scholar] [CrossRef] [PubMed]
- Vergori, A.; Cozzi Lepri, A.; Cicalini, S.; Matusali, G.; Bordoni, V.; Lanini, S.; Meschi, S.; Iannazzo, R.; Mazzotta, V.; Colavita, F.; et al. Immunogenicity to COVID-19 mRNA vaccine third dose in people living with HIV. Nat. Commun. 2022, 13, 4922. [Google Scholar] [CrossRef] [PubMed]
- Woldemeskel, B.A.; Karaba, A.H.; Garliss, C.C.; Beck, E.J.; Wang, K.H.; Laeyendecker, O.; Cox, A.L. Blankson JN: The BNT162b2 mRNA Vaccine Elicits Robust Humoral and Cellular Immune Responses in People Living with Human Immunodeficiency Virus (HIV). Clin. Infect. Dis. 2022, 74, 1268–1270. [Google Scholar] [CrossRef] [PubMed]
- Frater, J.; Ewer, K.J.; Ogbe, A.; Pace, M.; Adele, S.; Adland, E.; Alagaratnam, J.; Aley, P.K.; Ali, M.; Ansari, M.A.; et al. Safety and immunogenicity of the ChAdOx1 nCoV-19 (AZD1222) vaccine against SARS-CoV-2 in HIV infection: A single-arm substudy of a phase 2/3 clinical trial. Lancet HIV 2021, 8, e474–e485. [Google Scholar] [CrossRef] [PubMed]
- Fidler, S.; Fox, J.; Tipoe, T.; Longet, S.; Tipton, T.; Abeywickrema, M.; Adele, S.; Alagaratnam, J.; Ali, M.; Aley, P.K.; et al. Booster vaccination against SARS-CoV-2 induces potent immune responses in people with HIV. Clin. Infect. Dis. 2023, 76, 201–209. [Google Scholar] [CrossRef]
- Hensley, K.S.; Jongkees, M.J.; Geers, D.; GeurtsvanKessel, C.H.; Mueller, Y.M.; Dalm, V.; Papageorgiou, G.; Steggink, H.; Gorska, A.; Bogers, S.; et al. Immunogenicity and reactogenicity of SARS-CoV-2 vaccines in people living with HIV in the Netherlands: A nationwide prospective cohort study. PLoS Med. 2022, 19, e1003979. [Google Scholar] [CrossRef]
- Hung, C.Y.; Hsiao, S.H.; Huang, C.G.; Chang, C.S.; Chen, G.Y.; Huang, Y.L.; Dutta, A.; Huang, C.T. Relatively preserved functional immune capacity with standard COVID-19 vaccine regimen in people living with HIV. Front. Immunol. 2023, 14, 1204314. [Google Scholar] [CrossRef]
- Sop, J.; Traut, C.C.; Dykema, A.G.; Hunt, J.H.; Beckey, T.P.; Basseth, C.R.; Antar, A.A.R.; Laeyendecker, O.; Smith, K.N.; Blankson, J.N. Bivalent mRNA COVID vaccines elicit predominantly cross-reactive CD4+ T cell clonotypes. Cell Rep. Med. 2024, 5, 101442. [Google Scholar] [CrossRef]
- Lopez-Cortes, L.F.; Saborido-Alconchel, A.; Trujillo-Rodriguez, M.; Serna-Gallego, A.; Llaves-Flores, S.; Munoz-Muela, E.; Perez-Santos, M.J.; Lozano, C.; Mejias-Trueba, M.; Roca, C.; et al. Humoral and cellular immunity to SARS-CoV-2 after vaccination with mRNA vaccines in PLWH with discordant immune response. Influence of the vaccine administered. Front. Immunol. 2023, 14, 1129753. [Google Scholar] [CrossRef] [PubMed]
- Malin, J.J.; Suarez, I.; Biehl, L.M.; Schommers, P.; Knops, E.; Di Cristanziano, V.; Heger, E.; Pflieger, E.; Wyen, C.; Bettin, D.; et al. Immune response to mRNA-based COVID-19 booster vaccination in people living with HIV. HIV Med. 2023, 24, 785–793. [Google Scholar] [CrossRef]
- Alexandrova, Y.; Yero, A.; Mboumba Bouassa, R.S.; Comeau, E.; Samarani, S.; Brumme, Z.L.; Hull, M.; Crawley, A.M.; Langlois, M.A.; Angel, J.B.; et al. SARS-CoV-2 Vaccine-Induced T-Cell Response after Three Doses in People Living with HIV on Antiretroviral Therapy Compared to Seronegative Controls (CTN 328 COVAXHIV Study). Viruses 2023, 15, 575. [Google Scholar] [CrossRef]
- Antinori, A.; Cicalini, S.; Meschi, S.; Bordoni, V.; Lorenzini, P.; Vergori, A.; Lanini, S.; De Pascale, L.; Matusali, G.; Mariotti, D.; et al. Humoral and Cellular Immune Response Elicited by mRNA Vaccination Against Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) in People Living with Human Immunodeficiency Virus Receiving Antiretroviral Therapy Based on Current CD4 T-Lymphocyte Count. Clin. Infect. Dis. 2022, 75, e552–e563. [Google Scholar] [PubMed]
- Matsumoto, Y.; Murata, M.; Ohta, A.; Yamasaki, S.; Ikezaki, H.; Toyoda, K.; Shimono, N. The humoral and cellular immune responses following booster vaccination with SARS-CoV-2 mRNA in people living with human immunodeficiency virus. J. Infect. Chemother. 2024, 30, 417–422. [Google Scholar] [CrossRef] [PubMed]
- Benet, S.; Blanch-Lombarte, O.; Ainsua-Enrich, E.; Pedreno-Lopez, N.; Munoz-Basagoiti, J.; Raich-Regue, D.; Perez-Zsolt, D.; Pena, R.; Jimenez, E.; de la Concepcion, M.L.R.; et al. Limited Humoral and Specific T-Cell Responses After SARS-CoV-2 Vaccination in PWH With Poor Immune Reconstitution. J. Infect. Dis. 2022, 226, 1913–1923. [Google Scholar] [CrossRef] [PubMed]
- Vergori, A.; Cozzi-Lepri, A.; Matusali, G.; Cicalini, S.; Bordoni, V.; Meschi, S.; Mazzotta, V.; Colavita, F.; Fusto, M.; Cimini, E.; et al. Long Term Assessment of Anti-SARS-CoV-2 Immunogenicity after mRNA Vaccine in Persons Living with HIV. Vaccines 2023, 11, 1739. [Google Scholar] [CrossRef]
- Cheung, P.K.; Lapointe, H.R.; Sang, Y.; Ennis, S.; Mwimanzi, F.; Speckmaier, S.; Barad, E.; Dong, W.; Liang, R.; Simons, J.; et al. SARS-CoV-2 live virus neutralization after four COVID-19 vaccine doses in people with HIV receiving suppressive antiretroviral therapy. AIDS 2023, 37, F11–F18. [Google Scholar] [CrossRef] [PubMed]
- Public Health Agency of Canada. NACI Rapid Response: Extended Dose Intervals for COVID-19 Vaccines to Optimize Early Vaccine Rollout and Population Protection in Canada. 2021. Available online: https://www.canada.ca/en/public-health/services/immunization/national-advisory-committee-on-immunization-naci/rapid-response-extended-dose-intervals-covid-19-vaccines-early-rollout-population-protection.html (accessed on 9 January 2024).
- World Health Organization. Interim Recommendations for Use of the Moderna mRNA-1273 Vaccine against COVID-19; World Health Organization: Geneva, Switzerland, 2021. [Google Scholar]
- British Columbia Centre for Disease Control. Weekly Update on Variants of Concern. Available online: http://www.bccdc.ca/health-info/diseases-conditions/covid-19/data (accessed on 9 January 2024).
- Lv, Z.; Lv, S.; Li, Q.; Xia, Y.; Feng, Z.; Zhang, H.; Yang, H.; Wu, Z.; Zou, N.; Mo, Q.; et al. A third (booster) dose of the inactivated SARS-CoV-2 vaccine elicits immunogenicity and T follicular helper cell responses in people living with HIV. Front. Immunol. 2023, 14, 1264160. [Google Scholar] [CrossRef]
- Bertoletti, A.; Le Bert, N.; Tan, A.T. SARS-CoV-2-specific T cells in the changing landscape of the COVID-19 pandemic. Immunity 2022, 55, 1764–1778. [Google Scholar] [CrossRef]
- Lapuente, D.; Winkler, T.H.; Tenbusch, M. B-cell and antibody responses to SARS-CoV-2: Infection, vaccination, and hybrid immunity. Cell Mol. Immunol. 2024, 21, 144–158. [Google Scholar] [CrossRef] [PubMed]
- Murphy, T.J.; Swail, H.; Jain, J.; Anderson, M.; Awadalla, P.; Behl, L.; Brown, P.E.; Charlton, C.L.; Colwill, K.; Drews, S.J.; et al. The evolution of SARS-CoV-2 seroprevalence in Canada: A time-series study, 2020–2023. CMAJ 2023, 195, E1030–E1037. [Google Scholar] [CrossRef] [PubMed]
- Ahava, M.J.; Jarva, H.; Jaaskelainen, A.J.; Lappalainen, M.; Vapalahti, O.; Kurkela, S. Rapid increase in SARS-CoV-2 seroprevalence during the emergence of Omicron variant, Finland. Eur. J. Clin. Microbiol. Infect. Dis. 2022, 41, 997–999. [Google Scholar] [CrossRef] [PubMed]
- Clarke, K.E.N.; Jones, J.M.; Deng, Y.; Nycz, E.; Lee, A.; Iachan, R.; Gundlapalli, A.V.; Hall, A.J.; MacNeil, A. Seroprevalence of Infection-Induced SARS-CoV-2 Antibodies—United States, September 2021–February 2022. MMWR Morb. Mortal. Wkly. Rep. 2022, 71, 606–608. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | PLWH (n = 50) | Controls (n = 87) |
|---|---|---|
| Sociodemographic and health variables a | ||
| Age in years, median (IQR) | 58 (42–65) | 50 (35–72) |
| Female sex at birth, n (%) | 6 (12%) | 58 (67%) |
| Non-white ethnicity, n (%) | 14 (28%) | 37 (43%) |
| Number of chronic health conditions, median (IQR) b | 1 (0–1) | 0 (0–1) |
| HIV plasma viral load (RNA copies per mL), median (IQR) | <50 (<50–<50) | N.A. |
| Recent CD4 cell count (per µL), median (IQR) | 695 (468–983) | N.A. |
| Nadir CD4 cell count (per µL), median (IQR) | 250 (140–490) | N.A. |
| Vaccine details | ||
| Initial regimen c | ||
| mRNA only, n (%) | 42 (84%) | 84 (97%) |
| ChAdOx1-containing, n (%) | 8 (16%) | 3 (3%) |
| Third dose | ||
| BNT162b2, n (%) | 17 (34%) | 34 (39%) |
| mRNA-1273, n (%) | 33 (66%) | 53 (61%) |
| Days between first and second doses, median (IQR) | 59 (53–68) | 89 (73–98) |
| Days between second and third doses, median (IQR) | 182 (141–191) | 197 (169–215) |
| Post-third-vaccine dose SARS-CoV-2 infections, n (%) d | 21 (42%) | 38 (44%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Datwani, S.; Kalikawe, R.; Waterworth, R.; Mwimanzi, F.M.; Liang, R.; Sang, Y.; Lapointe, H.R.; Cheung, P.K.; Omondi, F.H.; Duncan, M.C.; et al. T-Cell Responses to COVID-19 Vaccines and Breakthrough Infection in People Living with HIV Receiving Antiretroviral Therapy. Viruses 2024, 16, 661. https://doi.org/10.3390/v16050661
Datwani S, Kalikawe R, Waterworth R, Mwimanzi FM, Liang R, Sang Y, Lapointe HR, Cheung PK, Omondi FH, Duncan MC, et al. T-Cell Responses to COVID-19 Vaccines and Breakthrough Infection in People Living with HIV Receiving Antiretroviral Therapy. Viruses. 2024; 16(5):661. https://doi.org/10.3390/v16050661
Chicago/Turabian StyleDatwani, Sneha, Rebecca Kalikawe, Rachel Waterworth, Francis M. Mwimanzi, Richard Liang, Yurou Sang, Hope R. Lapointe, Peter K. Cheung, Fredrick Harrison Omondi, Maggie C. Duncan, and et al. 2024. "T-Cell Responses to COVID-19 Vaccines and Breakthrough Infection in People Living with HIV Receiving Antiretroviral Therapy" Viruses 16, no. 5: 661. https://doi.org/10.3390/v16050661