
Cationic Residues of the HIV-1 Nucleocapsid Protein Enable DNA Condensation to Maintain Viral Core Particle Stability during Reverse Transcription

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Protein Purification and DNA Substrate
2.2. Optical Tweezers
2.3. AFM Imaging
3. Results
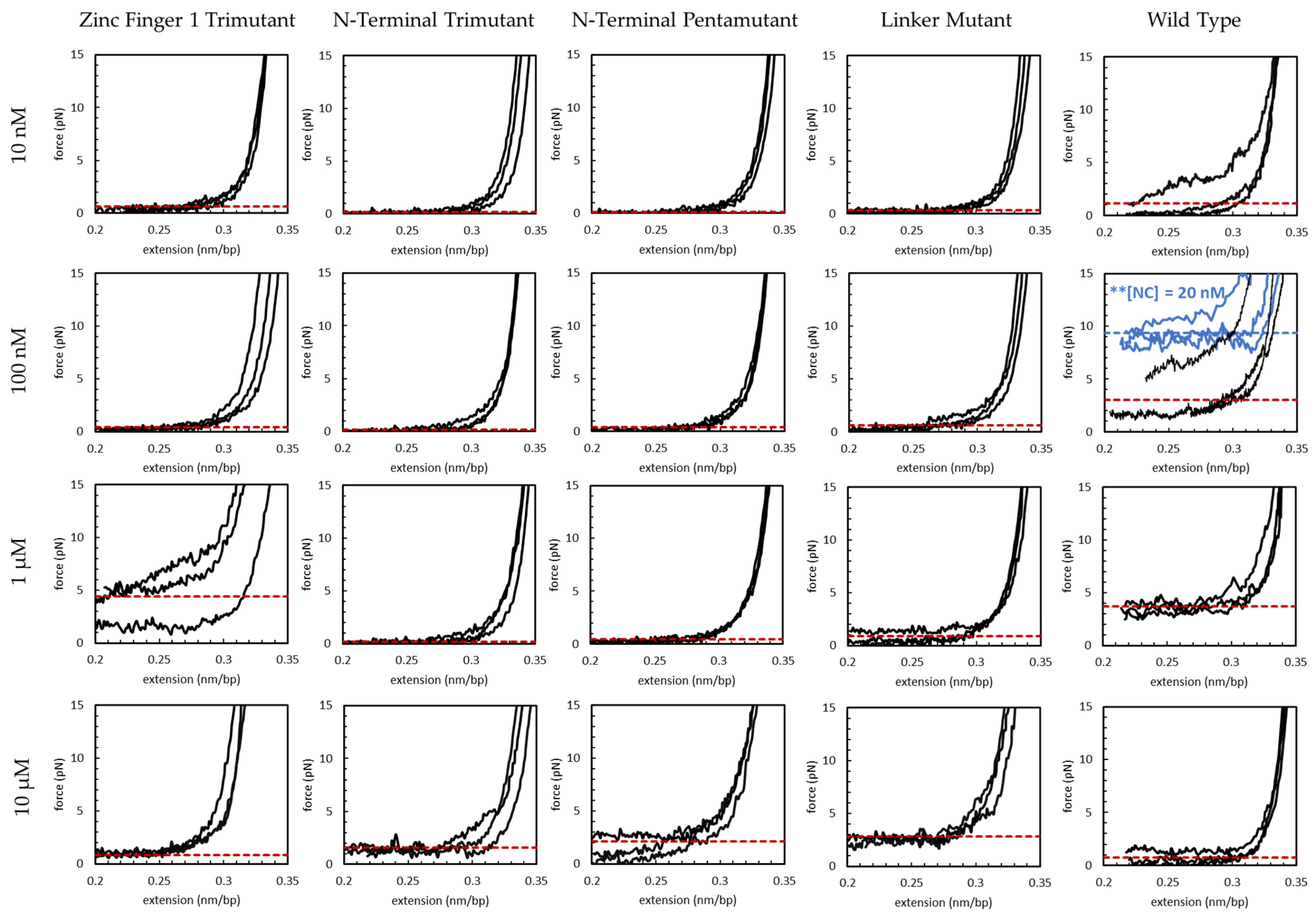
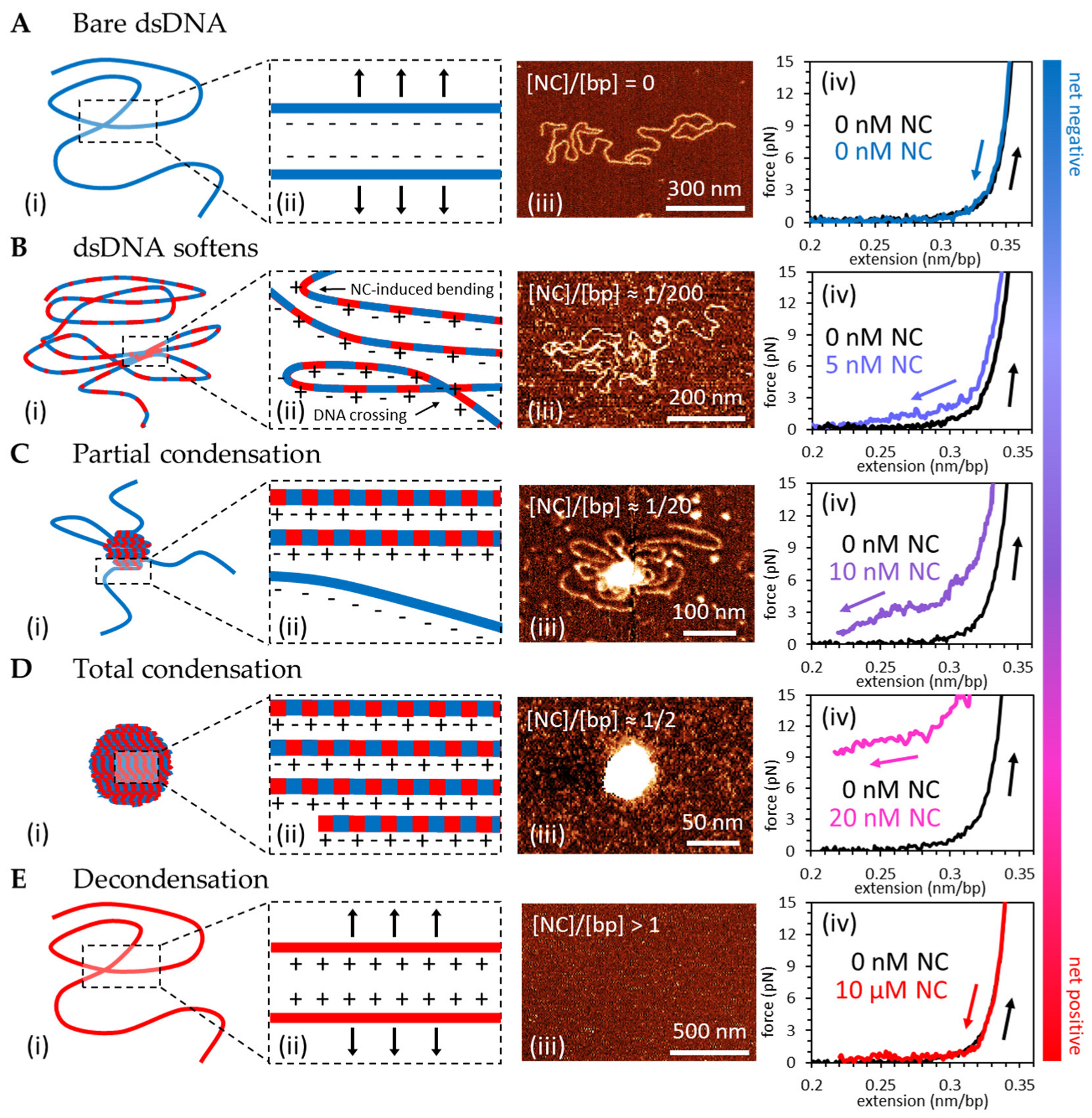
3.1. Force-Extension Measurements and AFM Imaging Show NC Softens, Condenses, Overcharges and Re-Solubilizes dsDNA
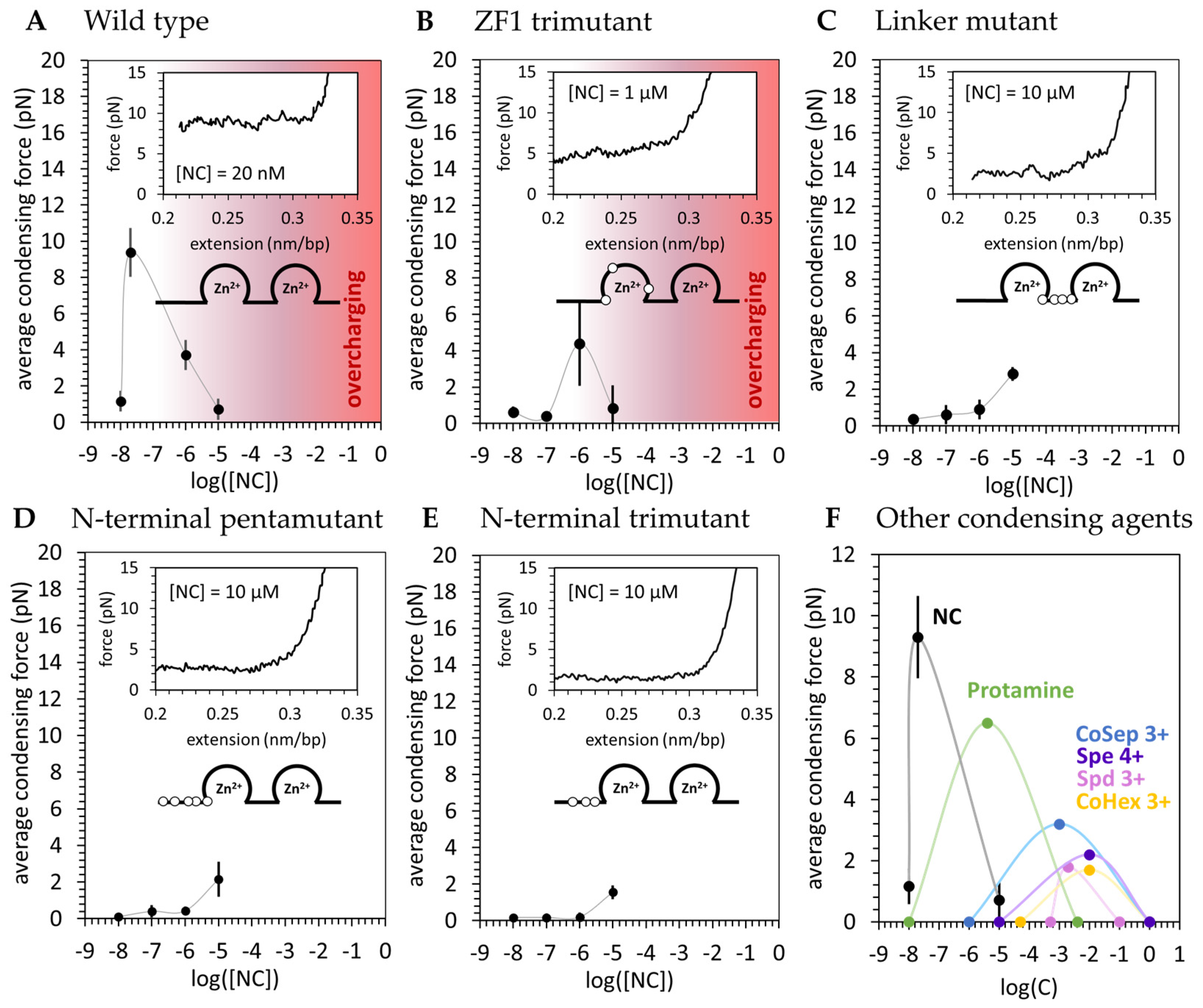
3.2. Basic Residue HIV-1 NC Mutants Are Defective in dsDNA Binding and Condensation
4. Discussion
4.1. NC Strongly Condenses DNA through a Counterion Correlation Mechanism That Is Sensitive to Cationic Mutations
4.2. DNA Condensation by NC Is Unusual in Its Ability to Phase Separate on DNA and Induce a Spherical Globule Instead of a Toroid
4.3. NC-Induced DNA Condensation Is Consistent with a Possible Function to Prevent Premature Capsid Uncoating during Reverse Transcription Leading to Defects in Viral Replication
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ligand or Protein | Cmin (M) | Cmid (M) | Cmax (M) | Fc,max (pN) | Z | Zeff | Buffer |
|---|---|---|---|---|---|---|---|
| Spd 3+ [51] | 5.0 × 10−4 | 2.0 × 10−3 | 1.0 × 10−1 | 1.80 | 3 | 3 | 10 mM Tris, pH 7 |
| CoHex 3+ [50] | 5.0 × 10−5 | 1.0 × 10−2 | 1.0 × 100 | 1.70 | 3 | 3 | 10 mM Tris, pH 7 |
| CoSep 3+ [50] | 1.0 × 10−6 | 1.0 × 10−3 | 1.0 × 100 | 3.20 | 3 | 3 | 10 mM Tris, 50 mM NaCl |
| Protamine [50] | 1.0 × 10−8 | 4.0 × 10−6 | 4.0 × 10−3 | 6.50 | 50 | 10 mM Tris, 50 mM NaCl | |
| Spe 4+ [50] | 1.0 × 10−5 | 1.0 × 10−2 | 1.0 × 100 | 2.20 | 4 | 4 | 10 mM Tris, 50 mM NaCl |
| NC 3.5+ | 1.0 × 10−8 | 2.0 × 10−8 | 1.0 × 10−5 | 9 +/− 1 | 11 | 3.5 | 10 mM Hepes, 45 mM NaCl, 5 mM NaOH, pH 7.5 |
References
- Aiken, C.; Rousso, I. The HIV-1 capsid and reverse transcription. Retrovirology 2021, 18, 29. [Google Scholar] [CrossRef]
- Briggs, J.A.G.; Simon, M.N.; Gross, I.; Kräusslich, H.-G.; Fuller, S.D.; Vogt, V.M.; Johnson, M.C. The stoichiometry of Gag protein in HIV-1. Nat. Struct. Mol. Biol. 2004, 11, 672–675. [Google Scholar] [CrossRef]
- Li, S.; Hill, C.P.; Sundquist, W.I.; Finch, J.T. Image reconstructions of helical assemblies of the HIV-1 CA protein. Nature 2000, 407, 409–413. [Google Scholar] [CrossRef] [PubMed]
- Ganser, B.K.; Li, S.; Klishko, V.Y.; Finch, J.T.; Sundquist, W.I. Assembly and Analysis of Conical Models for the HIV-1 Core. Science 1999, 283, 80–83. [Google Scholar] [CrossRef] [PubMed]
- Perilla, J.R.; Schulten, K. Physical properties of the HIV-1 capsid from all-atom molecular dynamics simulations. Nat. Commun. 2017, 8, 15959. [Google Scholar] [CrossRef]
- Campbell, E.M.; Hope, T.J. HIV-1 capsid: The multifaceted key player in HIV-1 infection. Nat. Rev. Microbiol. 2015, 13, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Bukrinsky, M. A hard way to the nucleus. Mol. Med. 2004, 10, 1–5. [Google Scholar] [CrossRef]
- Francis, A.C.; Marin, M.; Shi, J.; Aiken, C.; Melikyan, G.B. Time-Resolved Imaging of Single HIV-1 Uncoating In Vitro and in Living Cells. PLoS. Pathog. 2016, 12, e1005709. [Google Scholar] [CrossRef]
- Hulme, A.E.; Perez, O.; Hope, T.J. Complementary assays reveal a relationship between HIV-1 uncoating and reverse transcription. Proc. Natl. Acad. Sci. USA 2011, 108, 9975–9980. [Google Scholar] [CrossRef]
- Grewe, C.; Beck, A.; Gelderblom, H.R. HIV: Early virus-cell interactions. J. Acquir. Immune. Defic. Syndr. 1990, 3, 965–974. [Google Scholar]
- Arhel, N.J.; Souquere-Besse, S.; Munier, S.; Souque, P.; Guadagnini, S.; Rutherford, S.; Prevost, M.C.; Allen, T.D.; Charneau, P. HIV-1 DNA Flap formation promotes uncoating of the pre-integration complex at the nuclear pore. EMBO J. 2007, 26, 3025–3037. [Google Scholar] [CrossRef] [PubMed]
- Arhel, N. Revisiting HIV-1 uncoating. Retrovirology 2010, 7, 96. [Google Scholar] [CrossRef]
- Doitsh, G.; Galloway, N.L.K.; Geng, X.; Yang, Z.; Monroe, K.M.; Zepeda, O.; Hunt, P.W.; Hatano, H.; Sowinski, S.; Muñoz-Arias, I.; et al. Cell death by pyroptosis drives CD4 T-cell depletion in HIV-1 infection. Nature 2014, 505, 509–514. [Google Scholar] [CrossRef] [PubMed]
- Sumner, R.P.; Harrison, L.; Touizer, E.; Peacock, T.P.; Spencer, M.; Zuliani-Alvarez, L.; Towers, G.J. Disrupting HIV-1 capsid formation causes cGAS sensing of viral DNA. EMBO J. 2020, 39, e103958. [Google Scholar] [CrossRef]
- Deshpande, A.; Bryer, A.J.; Andino, J.; Shi, J.; Hong, J.; Torres, C.; Harel, S.; Francis, A.C.; Perilla, J.R.; Aiken, C.; et al. Elasticity of the HIV-1 Core Facilitates Nuclear Entry and Infection. bioRxiv 2023, 2023.09.29.560083. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.; Engelman, A.N. Capsid-host interactions for HIV-1 ingress. Microbiol. Mol. Biol. Rev. 2023, 87, e0004822. [Google Scholar] [CrossRef] [PubMed]
- Xue, G.; Yu, H.J.; Buffone, C.; Huang, S.-W.; Lee, K.; Goh, S.L.; Gres, A.T.; Guney, M.H.; Sarafianos, S.G.; Luban, J.; et al. The HIV-1 capsid core is an opportunistic nuclear import receptor. Nat. Commun. 2023, 14, 3782. [Google Scholar] [CrossRef] [PubMed]
- Shen, Q.; Kumari, S.; Xu, C.; Jang, S.; Shi, J.; Burdick, R.C.; Levintov, L.; Xiong, Q.; Wu, C.; Devarkar, S.C.; et al. The capsid lattice engages a bipartite NUP153 motif to mediate nuclear entry of HIV-1 cores. Proc. Natl. Acad. Sci. USA 2023, 120, e2202815120. [Google Scholar] [CrossRef]
- Shen, Q.; Feng, Q.; Wu, C.; Xiong, Q.; Tian, T.; Yuan, S.; Shi, J.; Bedwell, G.J.; Yang, R.; Aiken, C.; et al. Modeling HIV-1 nuclear entry with nucleoporin-gated DNA-origami channels. Nat. Struct. Mol. Biol. 2023, 30, 425–435. [Google Scholar] [CrossRef]
- Burdick, R.C.; Li, C.; Munshi, M.; Rawson, J.M.O.; Nagashima, K.; Hu, W.S.; Pathak, V.K. HIV-1 uncoats in the nucleus near sites of integration. Proc. Natl. Acad. Sci. USA 2020, 117, 5486–5493. [Google Scholar] [CrossRef]
- Dharan, A.; Bachmann, N.; Talley, S.; Zwikelmaier, V.; Campbell, E.M. Nuclear pore blockade reveals that HIV-1 completes reverse transcription and uncoating in the nucleus. Nat. Microbiol. 2020, 5, 1088–1095. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Burdick, R.C.; Nagashima, K.; Hu, W.S.; Pathak, V.K. HIV-1 cores retain their integrity until minutes before uncoating in the nucleus. Proc. Natl. Acad. Sci. USA 2021, 118, e2019467118. [Google Scholar] [CrossRef] [PubMed]
- Zila, V.; Margiotta, E.; Turonova, B.; Muller, T.G.; Zimmerli, C.E.; Mattei, S.; Allegretti, M.; Borner, K.; Rada, J.; Muller, B.; et al. Cone-shaped HIV-1 capsids are transported through intact nuclear pores. Cell 2021, 184, 1032–1046. [Google Scholar] [CrossRef] [PubMed]
- Francis, A.C.; Marin, M.; Singh, P.K.; Achuthan, V.; Prellberg, M.J.; Palermino-Rowland, K.; Lan, S.; Tedbury, P.R.; Sarafianos, S.G.; Engelman, A.N.; et al. HIV-1 replication complexes accumulate in nuclear speckles and integrate into speckle-associated genomic domains. Nat. Commun. 2020, 11, 3505. [Google Scholar] [CrossRef] [PubMed]
- Selyutina, A.; Persaud, M.; Lee, K.; KewalRamani, V.; Diaz-Griffero, F. Nuclear Import of the HIV-1 Core Precedes Reverse Transcription and Uncoating. Cell Rep. 2020, 32, 108201. [Google Scholar] [CrossRef] [PubMed]
- Rankovic, S.; Varadarajan, J.; Ramalho, R.; Aiken, C.; Rousso, I. Reverse Transcription Mechanically Initiates HIV-1 Capsid Disassembly. J. Virol. 2017, 91, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Christensen, D.E.; Ganser-Pornillos, B.K.; Johnson, J.S.; Pornillos, O.; Sundquist, W.I. Reconstitution and visualization of HIV-1 capsid-dependent replication and integration in vitro. Science 2020, 370, eabc8420. [Google Scholar] [CrossRef] [PubMed]
- Rankovic, S.; Deshpande, A.; Harel, S.; Aiken, C.; Rousso, I. HIV-1 uncoating occurs via a series of rapid biomechanical changes in the core related to individual stages of reverse transcription. J. Virol. 2021, 95, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Burdick, R.C.; Morse, M.; Rouzina, I.; Williams, M.C.; Hu, W.-S.; Pathak, V.K. HIV-1 uncoating requires long double-stranded reverse transcription products. Sci. Adv. 2024, 10, eadn7033. [Google Scholar] [CrossRef]
- Gien, H.; Morse, M.; McCauley, M.J.; Kitzrow, J.P.; Musier-Forsyth, K.; Gorelick, R.J.; Rouzina, I.; Williams, M.C. HIV-1 Nucleocapsid Protein Binds Double-Stranded DNA in Multiple Modes to Regulate Compaction and Capsid Uncoating. Viruses 2022, 14, 235. [Google Scholar] [CrossRef]
- Wilson, R.W.; Bloomfield, V.A. Counterion-induced condensation of deoxyribonucleic acid. A light-scattering study. Biochemistry 1979, 18, 2192–2196. [Google Scholar] [CrossRef]
- Pelta, J.; Livolant, F.; Sikorav, J.-L. DNA Aggregation Induced by Polyamines and Cobalthexamine (*). J. Biol. Chem. 1996, 271, 5656–5662. [Google Scholar] [CrossRef]
- Bloomfield, V.A. Condensation of DNA by multivalent cations: Considerations on mechanism. Biopolymers 1991, 31, 1471–1481. [Google Scholar] [CrossRef] [PubMed]
- Hud, N.V.; Vilfan, I.D. Toroidal DNA condensates: Unraveling the fine structure and the role of nucleation in determining size. Annu. Rev. Biophys. Biomol. Struct. 2005, 34, 295–318. [Google Scholar] [CrossRef]
- Webb, J.A.; Jones, C.P.; Parent, L.J.; Rouzina, I.; Musier-Forsyth, K. Distinct binding interactions of HIV-1 Gag to Psi and non-Psi RNAs: Implications for viral genomic RNA packaging. RNA 2013, 19, 1078–1088. [Google Scholar] [CrossRef] [PubMed]
- Vo, M.-N.; Barany, G.; Rouzina, I.; Musier-Forsyth, K. Effect of Mg2+ and Na+ on the Nucleic Acid Chaperone Activity of HIV-1 Nucleocapsid Protein: Implications for Reverse Transcription. J. Mol. Biol. 2009, 386, 773–788. [Google Scholar] [CrossRef] [PubMed]
- Rouzina, I.; Bloomfield, V.A. Influence of Ligand Spatial Organization on Competitive Electrostatic Binding to DNA. J. Phys. Chem. 1996, 100, 4305–4313. [Google Scholar] [CrossRef]
- Levin, J.G.; Mitra, M.; Mascarenhas, A.; Musier-Forsyth, K. Role of HIV-1 nucleocapsid protein in HIV-1 reverse transcription. RNA Biol. 2010, 7, 754–774. [Google Scholar] [CrossRef]
- Williams, M.C.; Rouzina, I.; Wenner, J.R.; Gorelick, R.J.; Musier-Forsyth, K.; Bloomfield, V.A. Mechanism for nucleic acid chaperone activity of HIV-1 nucleocapsid protein revealed by single molecule stretching. Proc. Natl. Acad. Sci. USA 2001, 98, 6121–6126. [Google Scholar] [CrossRef]
- Williams, M.C.; Gorelick, R.J.; Musier-Forsyth, K. Specific zinc-finger architecture required for HIV-1 nucleocapsid protein’s nucleic acid chaperone function. Proc. Natl. Acad. Sci. USA 2002, 99, 8614–8619. [Google Scholar] [CrossRef]
- Heng, X.; Kharytonchyk, S.; Garcia, E.L.; Lu, K.; Divakaruni, S.S.; LaCotti, C.; Edme, K.; Telesnitsky, A.; Summers, M.F. Identification of a minimal region of the HIV-1 5′-leader required for RNA dimerization, NC binding, and packaging. J. Mol. Biol. 2012, 417, 224–239. [Google Scholar] [CrossRef]
- Beltz, H.; Azoulay, J.; Bernacchi, S.; Clamme, J.P.; Ficheux, D.; Roques, B.; Darlix, J.L.; Mély, Y. Impact of the terminal bulges of HIV-1 cTAR DNA on its stability and the destabilizing activity of the nucleocapsid protein NCp7. J. Mol. Biol. 2003, 328, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Kafaie, J.; Song, R.; Abrahamyan, L.; Mouland, A.J.; Laughrea, M. Mapping of nucleocapsid residues important for HIV-1 genomic RNA dimerization and packaging. Virology 2008, 375, 592–610. [Google Scholar] [CrossRef]
- Mouhand, A.; Pasi, M.; Catala, M.; Zargarian, L.; Belfetmi, A.; Barraud, P.; Mauffret, O.; Tisné, C. Overview of the Nucleic-Acid Binding Properties of the HIV-1 Nucleocapsid Protein in Its Different Maturation States. Viruses 2020, 12, 1109. [Google Scholar] [CrossRef]
- Belfetmi, A.; Zargarian, L.; Tisné, C.; Sleiman, D.; Morellet, N.; Lescop, E.; Maskri, O.; René, B.; Mély, Y.; Fossé, P.; et al. Insights into the mechanisms of RNA secondary structure destabilization by the HIV-1 nucleocapsid protein. RNA 2016, 22, 506–517. [Google Scholar] [CrossRef]
- Rouzina, I.; Bloomfield, V.A. Macroion Attraction Due to Electrostatic Correlation between Screening Counterions. 1. Mobile Surface-Adsorbed Ions and Diffuse Ion Cloud. J. Phys. Chem. 1996, 100, 9977–9989. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Rouzina, I.; Shklovskii, B.I. Reentrant condensation of DNA induced by multivalent counterions. J. Chem. Phys. 2000, 112, 2562–2568. [Google Scholar] [CrossRef]
- Shklovskii, B.I. Screening of a macroion by multivalent ions: Correlation-induced inversion of charge. Phys. Rev. E Stat. Phys. Plasmas Fluids Relat. Interdiscip. Topics 1999, 60, 5802–5811. [Google Scholar] [CrossRef]
- Grosberg, A.Y.; Nguyen, T.T.; Shklovskii, B.I. Colloquium: The physics of charge inversion in chemical and biological systems. Rev. Mod. Phys. 2002, 74, 329–345. [Google Scholar] [CrossRef]
- Besteman, K.; Van Eijk, K.; Lemay, S.G. Charge inversion accompanies DNA condensation by multivalent ions. Nat. Phys. 2007, 3, 641–644. [Google Scholar] [CrossRef]
- Murayama, Y.; Sakamaki, Y.; Sano, M. Elastic Response of Single DNA Molecules Exhibits a Reentrant Collapsing Transition. Phys. Rev. Lett. 2003, 90, 018102. [Google Scholar] [CrossRef] [PubMed]
- Mengarelli, V.; Auvray, L.; Pastré, D.; Zeghal, M. Charge inversion, condensation and decondensation of DNA and polystyrene sulfonate by polyethylenimine. Eur. Phys. J. E Soft Matter 2011, 34, 127. [Google Scholar] [CrossRef] [PubMed]
- Raspaud, E.; Pelta, J.; de Frutos, M.; Livolant, F. Solubility and Charge Inversion of Complexes of DNA and Basic Proteins. Phys. Rev. Lett. 2006, 97, 068103. [Google Scholar] [CrossRef] [PubMed]
- Jang, Y.H.; Raspaud, E.; Lansac, Y. DNA-protamine condensates under low salt conditions: Molecular dynamics simulation with a simple coarse-grained model focusing on electrostatic interactions. Nanoscale Adv. 2023, 5, 4798–4808. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, A.; de Izarra, A.; Degrouard, J.; Olive, E.; Maiti, P.K.; Jang, Y.H.; Lansac, Y. Protamine-Controlled Reversible DNA Packaging: A Molecular Glue. ACS Nano 2021, 15, 13094–13104. [Google Scholar] [CrossRef] [PubMed]
- Balhorn, R.; Brewer, L.; Corzett, M. DNA condensation by protamine and arginine-rich peptides: Analysis of toroid stability using single DNA molecules. Mol. Reprod. Dev. 2000, 56, 230–234. [Google Scholar] [CrossRef]
- Wu, H.; Mitra, M.; Naufer, M.N.; McCauley, M.J.; Gorelick, R.J.; Rouzina, I.; Musier-Forsyth, K.; Williams, M.C. Differential contribution of basic residues to HIV-1 nucleocapsid protein’s nucleic acid chaperone function and retroviral replication. Nucleic Acids Res. 2014, 42, 2525–2537. [Google Scholar] [CrossRef] [PubMed]
- Carteau, S.; Gorelick, R.J.; Bushman, F.D. Coupled Integration of Human Immunodeficiency Virus Type 1 cDNA Ends by Purified Integrase In Vitro: Stimulation by the Viral Nucleocapsid Protein. J. Virol. 1999, 73, 6670–6679. [Google Scholar] [CrossRef] [PubMed]
- Cruceanu, M.; Urbaneja, M.A.; Hixson, C.V.; Johnson, D.G.; Datta, S.A.; Fivash, M.J.; Stephen, A.G.; Fisher, R.J.; Gorelick, R.J.; Casas-Finet, J.R.; et al. Nucleic acid binding and chaperone properties of HIV-1 Gag and nucleocapsid proteins. Nucleic Acids Res. 2006, 34, 593–605. [Google Scholar] [CrossRef]
- Morse, M.; Sefcikova, J.; Rouzina, I.; Beuning, P.J.; Williams, M.C. Structural domains of SARS-CoV-2 nucleocapsid protein coordinate to compact long nucleic acid substrates. Nucleic Acids Res. 2022, 51, 290–303. [Google Scholar] [CrossRef]
- Naufer, M.N.; Morse, M.; Möller, G.B.; McIsaac, J.; Rouzina, I.; Beuning, P.J.; Williams, M.C. Multiprotein E. coli SSB–ssDNA complex shows both stable binding and rapid dissociation due to interprotein interactions. Nucleic Acids Res. 2021, 49, 1532–1549. [Google Scholar] [CrossRef] [PubMed]
- Chaurasiya, K.R.; Paramanathan, T.; McCauley, M.J.; Williams, M.C. Biophysical characterization of DNA binding from single molecule force measurements. Phys. Life Rev. 2010, 7, 299–341. [Google Scholar] [CrossRef] [PubMed]
- Morse, M.; Naufer, M.N.; Feng, Y.; Chelico, L.; Rouzina, I.; Williams, M.C. HIV restriction factor APOBEC3G binds in multiple steps and conformations to search and deaminate single-stranded DNA. eLife 2019, 8, e52649. [Google Scholar] [CrossRef]
- Smith, S.B.; Cui, Y.; Bustamante, C. Overstretching B-DNA: The Elastic Response of Individual Double-Stranded and Single-Stranded DNA Molecules. Science 1996, 271, 795–799. [Google Scholar] [CrossRef] [PubMed]
- Hamon, L.; Pastré, D.; Dupaigne, P.; Breton, C.L.; Cam, E.L.; Piétrement, O. High-resolution AFM imaging of single-stranded DNA-binding (SSB) protein—DNA complexes. Nucleic Acids Res. 2007, 35, e58. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Musier-Forsyth, K.; Falk, C.; Barbara, P.F. Single-Molecule Spectroscopic Study of Dynamic Nanoscale DNA Bending Behavior of HIV-1 Nucleocapsid Protein. J. Phys. Chem. B 2013, 117, 4183–4196. [Google Scholar] [CrossRef] [PubMed]
- Baumann, C.G.; Bloomfield, V.A.; Smith, S.B.; Bustamante, C.; Wang, M.D.; Block, S.M. Stretching of Single Collapsed DNA Molecules. Biophys. J. 2000, 78, 1965–1978. [Google Scholar] [CrossRef] [PubMed]
- Rouzina, I.; Bloomfield, V.A. DNA bending by small, mobile multivalent cations. Biophys. J. 1998, 74, 3152–3164. [Google Scholar] [CrossRef] [PubMed]
- Jiang, K.; Humbert, N.; Sriram, K.K.; Rouzina, I.; Mely, Y.; Westerlund, F. The HIV-1 nucleocapsid chaperone protein forms locally compacted globules on long double-stranded DNA. Nucleic Acids Res. 2021, 49, 4550–4563. [Google Scholar] [CrossRef]
- McMillan, R.B.; Bediako, H.; Devenica, L.M.; Velasquez, A.; Hardy, I.P.; Ma, Y.E.; Roscoe, D.M.; Carter, A.R. Protamine folds DNA into flowers and loop stacks. Biophys. J. 2023, 122, 4288–4302. [Google Scholar] [CrossRef]
- DeRouchey, J.; Hoover, B.; Rau, D.C. A comparison of DNA compaction by arginine and lysine peptides: A physical basis for arginine rich protamines. Biochemistry 2013, 52, 3000–3009. [Google Scholar] [CrossRef] [PubMed]
- van den Broek, B.; Noom, M.C.; van Mameren, J.; Battle, C.; Mackintosh, F.C.; Wuite, G.J. Visualizing the formation and collapse of DNA toroids. Biophys. J. 2010, 98, 1902–1910. [Google Scholar] [CrossRef] [PubMed]
- Deserno, M.; Arnold, A.; Holm, C. Attraction and Ionic Correlations between Charged Stiff Polyelectrolytes. Macromolecules 2003, 36, 249–259. [Google Scholar] [CrossRef]
- Kornyshev, A.A.; Leikin, S. Symmetry laws for interaction between helical macromolecules. Biophys. J. 1998, 75, 2513–2519. [Google Scholar] [CrossRef] [PubMed]
- Kornyshev, A.A.; Leikin, S. Helical Structure Determines Different Susceptibilities of dsDNA, dsRNA, and tsDNA to Counterion-Induced Condensation. Biophys. J. 2013, 104, 2031–2041. [Google Scholar] [CrossRef] [PubMed]
- Wynveen, A.; Lee, D.J.; Kornyshev, A.A.; Leikin, S. Helical coherence of DNA in crystals and solution. Nucleic Acids Res. 2008, 36, 5540–5551. [Google Scholar] [CrossRef] [PubMed]
- Rouzina, I.; Bruinsma, R. DNA confinement drives uncoating of the HIV Virus. Eur. Phys. J. Spec. Top. 2014, 223, 1745–1754. [Google Scholar] [CrossRef]
- Purohit, P.K.; Inamdar, M.M.; Grayson, P.D.; Squires, T.M.; Kondev, J.; Phillips, R. Forces during bacteriophage DNA packaging and ejection. Biophys J. 2005, 88, 851–866. [Google Scholar] [CrossRef] [PubMed]
- Rau, D.C.; Lee, B.; Parsegian, V.A. Measurement of the repulsive force between polyelectrolyte molecules in ionic solution: Hydration forces between parallel DNA double helices. Proc. Natl. Acad. Sci. USA 1984, 81, 2621–2625. [Google Scholar] [CrossRef]



| Variant | RNA Packaging | Single-Round Infectivity | Relative Multiple round H9 Infectivity |
|---|---|---|---|
| Wild-type | 100 | 100 | 1.0 |
| N-terminal trimutant | 51 ± 19 | 7.4 ±1.6 | (3.8 ± 3.3) × 10−3 |
| Zinc finger 1 trimutant | 31 ± 14 | 8.8 ± 2.4 | (5.0 ± 4.4) × 10−5 |
| Zinc finger linker mutant | 61 ± 11 | 3.2 ± 0.7 | (7.2 ± 4.5) × 10−5 |
| N-terminal pentamutant | 3.0 ± 0.3 | 0.016 ± 0.007 | ≤(7.1) × 10−6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gien, H.; Morse, M.; McCauley, M.J.; Rouzina, I.; Gorelick, R.J.; Williams, M.C. Cationic Residues of the HIV-1 Nucleocapsid Protein Enable DNA Condensation to Maintain Viral Core Particle Stability during Reverse Transcription. Viruses 2024, 16, 872. https://doi.org/10.3390/v16060872
Gien H, Morse M, McCauley MJ, Rouzina I, Gorelick RJ, Williams MC. Cationic Residues of the HIV-1 Nucleocapsid Protein Enable DNA Condensation to Maintain Viral Core Particle Stability during Reverse Transcription. Viruses. 2024; 16(6):872. https://doi.org/10.3390/v16060872
Chicago/Turabian StyleGien, Helena, Michael Morse, Micah J. McCauley, Ioulia Rouzina, Robert J. Gorelick, and Mark C. Williams. 2024. "Cationic Residues of the HIV-1 Nucleocapsid Protein Enable DNA Condensation to Maintain Viral Core Particle Stability during Reverse Transcription" Viruses 16, no. 6: 872. https://doi.org/10.3390/v16060872