
Viral Dynamics in the Tropical Pacific Ocean: A Comparison between Within and Outside a Warm Eddy
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Sampling
2.2. Viral Production
2.3. Flow Cytometric Analyses
2.4. Statistical Analysis
3. Results
3.1. Station Characteristics
3.2. Bacterial and Viral Abundance
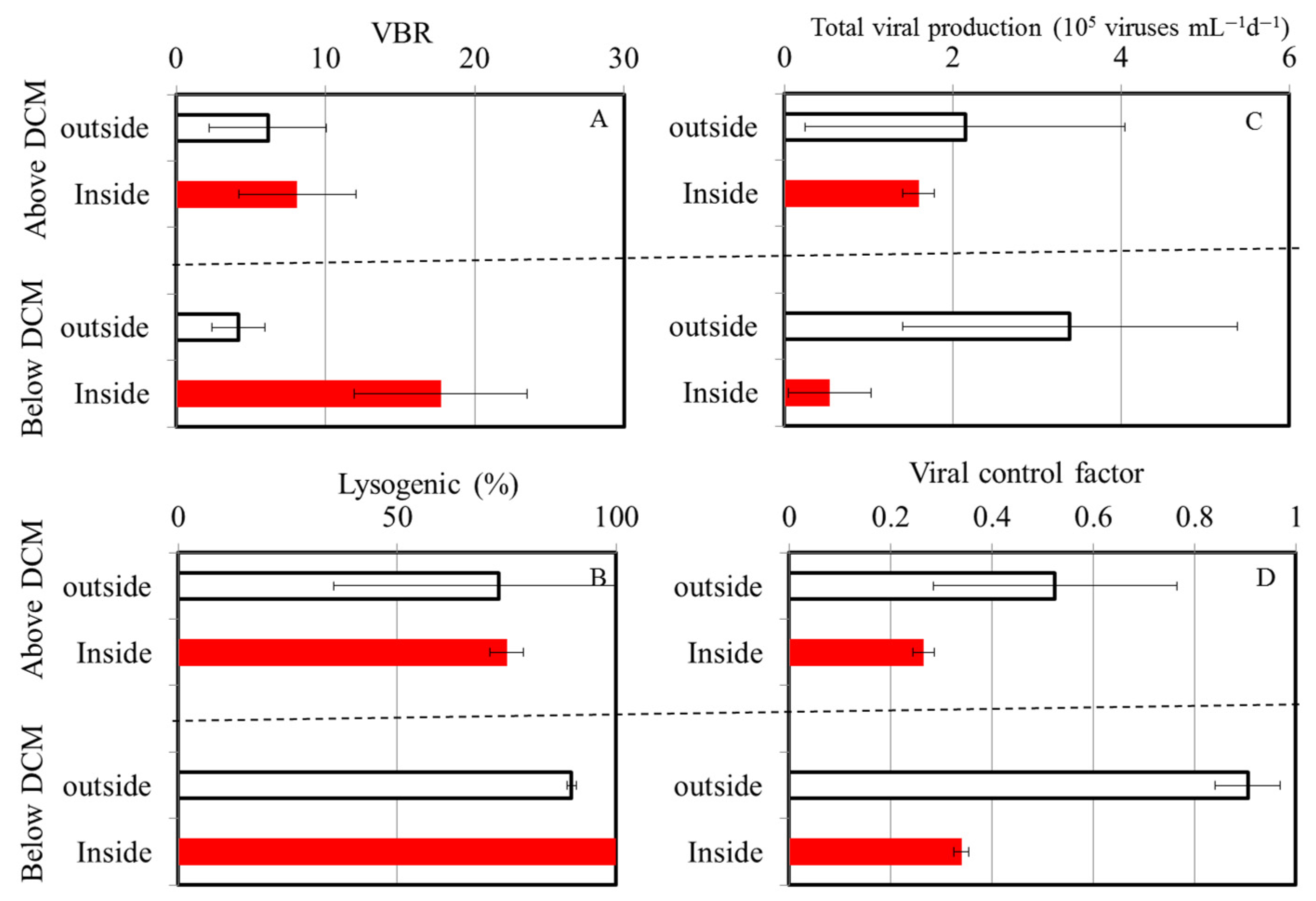
3.3. Viral Production
3.4. Virus-Mediated Bacterial Mortality
4. Discussion
4.1. Vertical Patterns of Bacterial Abundances
4.2. Percentage of Lysogenic Viral Production
4.3. Virus-to-Bacteria Ratios
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chelton, D.B.; Gaube, P.; Schlax, M.G.; Early, J.J.; Samelson, R.M. The influence of nonlinear mesoscale eddies on near—Surface oceanic chlorophyll. Science 2011, 334, 328–332. [Google Scholar] [CrossRef]
- Chang, Y.L.; Miyazawa, Y.; Oey, L.Y.; Kodaira, T.; Huang, S. The formation processes of phytoplankton growth and decline in mesoscale eddies in the western north pacific ocean. J. Geophys. Res. Oce. 2017, 122, 4444–4455. [Google Scholar] [CrossRef]
- An, L.; Liu, X.; Xu, F.; Fan, X.; Wang, P.; Yin, W.; Huang, B. Different responses of plankton community to mesoscale eddies in the western equatorial Pacific Ocean. Deep Sea Res. I 2024, 203, 104219. [Google Scholar] [CrossRef]
- McGillicuddy, D.J., Jr. Mechanisms of physical-biological-biogeochemical interaction at the oceanic mesoscale. Ann. Rev. Mar. Sci. 2016, 8, 125–159. [Google Scholar] [CrossRef] [PubMed]
- Zohary, T.; Robarts, R.D. Bacterial numbers, bacterial production, and heterotrophic nanoplankton abundance in a warm core eddy in the Eastern Mediterranean. Mar. Ecol. Prog. Ser. 1992, 84, 133–137. [Google Scholar] [CrossRef]
- Robarts, R.D.; Zohary, T.; Waiser, M.J.; Yacobi, Y.Z. Bacterial abundance, biomass, and production in relation to phytoplankton biomass in the Levantine Basin of the southeastern Mediterranean Sea. Mar. Ecol. Prog. Ser. 1996, 137, 273–281. [Google Scholar] [CrossRef]
- Li, J.; Jiang, X.; Li, G.; Jing, Z.; Zhou, L.; Ke, Z.; Tan, Y. Distribution of picoplankton in the northeastern South China Sea with special reference to the effects of the Kuroshio intrusion and the associated mesoscale eddies. Sci. Tot. Environ. 2017, 589, 1–10. [Google Scholar] [CrossRef]
- Hyun, J.H.; Kim, D.; Shin, C.W.; Noh, J.H.; Yang, E.J.; Mok, J.S.; Kim, S.H.; Kim, H.C.; Yoo, S. Enhanced phytoplankton and bacterioplankton production coupled to coastal upwelling and an anticyclonic eddy in the Ulleung Basin, East Sea. Aquat. Microb. Ecol. 2009, 54, 45–54. [Google Scholar] [CrossRef]
- Ewart, C.S.; Meyers, M.K.; Wallner, E.R.; McGillicuddy, D.J., Jr.; Carlson, C.A. Microbial dynamics in cyclonic and anticyclonic mode-water eddies in the northwestern Sargasso Sea. Deep Sea Res. Part II Top. Stud. Oceanogra. 2008, 55, 1334–1347. [Google Scholar] [CrossRef]
- Sun, F.; Xia, X.; Simon, M.; Wang, Y.; Zhao, H.; Sun, C.; Cheng, H.; Wang, Y.; Hu, S.; Fei, J.; et al. Anticyclonic eddy driving significant changes in prokaryotic and eukaryotic communities in the south China Sea. Front. Mar. Sci. 2022, 9, 773548. [Google Scholar] [CrossRef]
- Weinbauer, M.G. Ecology of prokaryotic viruses. FEMS Microbiol. Rev. 2004, 28, 127–181. [Google Scholar] [CrossRef] [PubMed]
- Suttle, C.A. Viruses in the sea. Nature 2005, 437, 356–361. [Google Scholar] [CrossRef] [PubMed]
- Wommack, K.E.; Colwell, R.R. Virioplankton: Viruses in aquatic ecosystems. Microbiol. Mol. Biol. Rev. 2000, 64, 69–114. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Wei, W.; Cai, L.L. The fate and biogeochemical cycling of viral elements. Nat. Rev. Microbiol. 2014, 12, 850–851. [Google Scholar] [CrossRef] [PubMed]
- Auguet, J.C.; Montanie, H.; Delmas, D.; Hartmann, H.J.; Huet, V. Dynamic of virioplankton abundance and its environmental control in the Charente estuary (France). Microb. Ecol. 2005, 50, 337–349. [Google Scholar] [CrossRef] [PubMed]
- Thomas, R.; Berdjeb, L.; Sime-Ngando, T.; Jacquet, S. Viral abundance, production, decay rates and life strategies (lysogeny versus lysis) in Lake Bourget (France). Environ. Microbiol. 2011, 13, 616–630. [Google Scholar] [CrossRef] [PubMed]
- Junger, P.C.; Amado, A.M.; Paranhos, R.; Cabral, A.S.; Jacques, S.M.; Farjalla, V.F. Salinity drives the virioplankton abundance but not production in tropical coastal lagoons. Microb. Ecol. 2018, 75, 52–63. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; He, M.; Shi, Z.; Xu, J. Nutritional Status Regulates Bacteria-Virus Interactions in the Northern South China Sea. J. Geophy. Res. 2023, 128, e2023JG007469. [Google Scholar] [CrossRef]
- Butina, T.V.; Bukin, Y.S.; Krasnopeev, A.S.; Belykh, O.I.; Tupikin, A.E.; Kabilov, M.R.; Sakirko, M.V.; Belikov, S.I. Estimate of the diversity of viral and bacterial assemblage in the coastal water of Lake Baikal. FEMS Microbiol. Let. 2019, 366, fnz094. [Google Scholar] [CrossRef]
- Wigington, C.H.; Sonderegger, D.; Brussaard, C.P.D.; Buchan, A.; Finke, J.F.; Fuhrman, J.A.; Lennon, J.T.; Middelboe, M.; Suttle, C.A.; Stock, C.; et al. Re-examination of the relationship between marine virus and microbial cell abundances. Nat. Microbiol. 2016, 1, 1–9. [Google Scholar] [CrossRef]
- Parikka, K.J.; Le Romancer, M.; Wauters, N.; Jacquet, S. Deciphering the virus-to-prokaryote ratio (VPR): Insights into virus–host relationships in a variety of ecosystems. Biol. Rev. 2016, 92, 1081–1100. [Google Scholar] [CrossRef]
- Taylor, G.T.; Hein, C.; Iabichella, M. Temporal variations in viral distributions in the anoxic Cariaco Basin. Aquat. Microb. Ecol. 2003, 30, 103–116. [Google Scholar] [CrossRef]
- De Corte, D.; Sintes, E.; Winter, C.; Yokokawa, T.; Reinthaler, T.; Herndl, G.J. Links between viral and prokaryotic communities throughout the water column in the (sub) tropical Atlantic Ocean. ISME J. 2010, 4, 1431–1442. [Google Scholar] [CrossRef]
- De Corte, D.; Sintes, E.; Yokokawa, T.; Lekunberri, I.; Herndl, G.J. Large-scale distribution of microbial and viral populations in the South Atlantic Ocean. Environ. Microbiol. Rep. 2016, 8, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Yokokawa, T.; Motegi, C.; Nagata, T. Large-scale distribution of viruses in deep waters of the Pacific and Southern Oceans. Aquat. Microb. Ecol. 2014, 71, 193–202. [Google Scholar] [CrossRef]
- Weinbauer, M.G.; Ingrid, B.; Manfred, G.H. Lysogeny and virus-induced mortality of bacterioplankton in surface, deep, and anoxic marine waters. Limnol. Oceanogr. 2003, 48, 1457–1465. [Google Scholar] [CrossRef]
- Wilhelm, S.W.; Brigden, S.M.; Suttle, C.A. A dilution technique for the direct measurement of viral production: A comparison in stratified and tidally mixed coastal waters. Microb. Ecol. 2002, 43, 168–173. [Google Scholar] [CrossRef]
- Weinbauer, M.G.; Hófle, M.G. Significance of viral lysis andflagellate grazing as factors controlling bacterioplankton pro-duction in a eutrophic lake. Appl. Environ. Microbiol. 1998, 64, 431–438. [Google Scholar]
- Hewson, I.; Fuhrman, J.A. Covariation of viral parameters with bacterial assemblage richness and diversity in the water column and sediments. Deep-Sea Res. I. 2007, 54, 811–830. [Google Scholar]
- Parada, V.; Herndl, G.J.; Weinbauer, M.G. Viral burst size of heterotrophic prokaryotes in aquatic systems. J. Mar. Biol. Assoc. 2006, 86, 613–621. [Google Scholar] [CrossRef]
- Brussaard, C.P.D. Optimization of procedures for counting viruses by flow cytometry. Appl. Environ. Microbiol. 2004, 70, 1506–1513. [Google Scholar] [CrossRef]
- Hammes, F.; Egli, T. Cytometric methods for measuring bacteria in water: Advantages, pitfalls and applications. Anal. Bioanal. Chem. 2010, 397, 1083–1095. [Google Scholar] [CrossRef] [PubMed]
- Calvo-Díaz, A.; Morán, X.A.G. Seasonal dynamics of picoplankton in shelf waters of the southern Bay of Biscay. Aquat. Microb. Ecol. 2006, 42, 159–174. [Google Scholar] [CrossRef]
- Yun, M.S.; Kim, Y.; Jeong, Y.; Joo, H.T.; Jo, Y.H.; Lee, C.H.; Bae, H.; Lee, D.; Bhavya, P.S.; Kim, D.; et al. Weak response of biological productivity and community structure of phytoplankton to mesoscale eddies in the oligotrophic Philippine Sea. J. Geophy. Res. Oce. 2020, 125, e2020JC016436. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, F.; He, X.; Wang, W.; Chang, L.; Kang, J. Latitudinal and meridional patterns of picophytoplankton variability are contrastingly associated with Ekman pumping and the warm pool in the tropical western Pacific. Ecol. Evolut. 2023, 13, e10589. [Google Scholar] [CrossRef] [PubMed]
- Kirchman, D.L.; Malmstrom, R.R.; Cottrell, M.T. Control of bacterial growth by temperature and organic matter in the Western Arctic. Deep Sea Res. II Top. Stud. Oceanogra. 2005, 52, 3386–3395. [Google Scholar] [CrossRef]
- Morán, X.A.G.; Baltar, F.; Carreira, C.; Lønborg, C. Responses of physiological groups of tropical heterotrophic bacteria to temperature and dissolved organic matter additions: Food matters more than warming. Environ. Microbiol. 2020, 22, 1930–1943. [Google Scholar] [CrossRef] [PubMed]
- Kirchman, D.; Rich, J. Regulation of Bacterial Growth Rates by Dissolved Organic Carbon and Temperature in the Equatorial Pacific Ocean. Microb. Ecol. 1997, 33, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Shiah, F.-K.; Ducklow, H.W. Temperature regulation of heterotrophic bacterioplankton abundance, production and specific growth rate in Chesapeake Bay, USA. Limnol. Oceanogr. 1994, 39, 1243–1258. [Google Scholar] [CrossRef]
- Chen, Z.; Sun, J.; Gu, T.; Zhang, G.; Wei, Y. Nutrient ratios driven by vertical stratification regulate phytoplankton community structure in the oligotrophic western Pacific Ocean. Oce. Sci. 2021, 17, 1775–1789. [Google Scholar] [CrossRef]
- Liu, H.; Yuan, X.; Xu, J.; Harrison, P.J.; He, L.; Yin, K. Effects of viruses on bacterial functions under contrasting nutritional conditions for four species of bacteria isolated from Hong Kong waters. Scient. Rep. 2015, 5, 14217. [Google Scholar] [CrossRef]
- Mine, A.H.; Coleman, M.L. Phosphorus release and regeneration following laboratory lysis of bacterial cells. Front. Microbiol. 2021, 12, 641700. [Google Scholar] [CrossRef] [PubMed]
- Tsai, A.-Y.; Gong, G.C.; Yu-Wen, H. Importance of the viral shunt in nitrogen cycling in Synechococcus spp. growth in subtropical Western Pacific coastal waters. Terr. Atmo. Oceanic Sci. 2014, 25, 839–846. [Google Scholar] [CrossRef]
- Boras, J.A.; Sala, M.M.; Baltar, F.; Arístegui, J.; Duarte, C.M.; Vaqué, D. Effect of viruses and protists on bacteria in eddies of the Canary Current region (subtropical northeast Atlantic). Limnol. Oceanogr. 2010, 55, 885–898. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, J.; Yu, J.; Wu, Q.; Sun, D. Anticyclonic mesoscale eddy induced mesopelagic biomass hotspot in the oligotrophic ocean. J. Mar. Syst. 2023, 237, 103831. [Google Scholar] [CrossRef]
- Weinbauer, M.G.; Rowe, J.M.; Wilhelm, S.W. Determining rates of virus production in aquatic systems by the virus reduction approach. Man. Aquat. Virus Ecol. ASLO 2010, 1–8. [Google Scholar] [CrossRef]
- Weinbauer, M.G.; Suttle, C.A. Lysogeny and prophage induction in coastal and offshore bacterial communities. Aquat. Microb. Ecol. 1999, 18, 217–225. [Google Scholar] [CrossRef]
- Williamson, S.J.; Houchin, L.A.; McDaniel, L.; Paul, J.H. Seasonal variation in lysogeny as depicted by prophage induction in Tampa Bay, Florida. Appl. Environ. Microbiol. 2002, 68, 4307–4314. [Google Scholar] [CrossRef] [PubMed]
- Howard-Varona, C.; Hargreaves, K.R.; Abedon, S.T.; Sullivan, M.B. Lysogeny in nature: Mechanisms, impact and ecology of temperate phages. ISME J. 2017, 11, 1511–1520. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Zhang, R.; Peng, L.; Liang, Y.; Jiao, N. Effects of temperature and photosynthetically active radiation on virioplankton decay in the western Pacific Ocean. Sci. Rep. 2018, 8, 1525. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Chen, X.; Weinbauer, M.G.; Jiao, N.; Zhang, R. Reduced bacterial mortality and enhanced viral productivity during sinking in the ocean. ISME J. 2022, 16, 1668–1675. [Google Scholar] [CrossRef]
- Weinbauer, M.G.; Bettarel, Y.; Cattaneo, R.; Luef, B.; Maier, C.; Motegi, C.; Peduzzi, P.; Mari, X. Viral ecology of organic and inorganic particles in aquatic systems: Avenues for further research. Aquat. Microb. Ecol. 2009, 57, 321–341. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Temperature | Salinity | Virusrs | Bacteria | VBR | Lytic Production | Lysogenic Production | Lysogenic (%) | TVP | VT | VMB # | LRB-V | Bacterial Growth Rate | Viral Control Factor | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (℃) | (psu) | (106 viruses mL−1) | (105 cells mL−1) | (105 viruses mL−1d−1) | (105 viruses mL−1d−1) | (105 viruses mL−1d−1) | (d−1) | (cells mL−1d−1) | (d−1) | (d−1) | |||||
| T1 | surface | 28.33 | 34.62 | 1± 0.26 | 1.12 ± 0.01 | 8.90 ± 2.43 | 0.00 | 3.84 * | 100.0 | 3.84 | 0.38 | 15,360 | 0.14 | 0.14 | 0.99 |
| DCM | 22.00 | 34.84 | 1.94 ± 0.45 | 5.95 ± 0.85 | 3.36 ± 1.27 | 0.34 | 0.29 | 46.3 | 0.63 | 0.03 | 2504 | 0.004 | 0.07 | 0.06 | |
| 200 m | 19.33 | 34.86 | 1.11 ± 0.02 | 4.44 ± 2.11 | 2.89 ± 1.54 | 0.53 | 4.27 * | 89.0 | 4.80 | 0.43 | 19,200 | 0.04 | 0.05 | 0.86 | |
| 500 m | 8.95 | 34.18 | 1.14 ± 0.11 | 2.1 ± 0.08 | 5.41 ± 0.22 | 0.19 | 1.80 * | 90.5 | 1.99 | 0.17 | 7960 | 0.04 | 0.04 | 0.95 | |
| T3 | surface | 29.19 | 34.63 | 1.31 ± 0.01 | 1.65 ± 1.20 | 10.87 ± 3.24 | 0.48 | 1.25 | 72.3 | 1.73 | 0.13 | 6920 | 0.04 | 0.17 | 0.25 |
| DCM | 25.00 | 34.93 | 0.86 ± 0.01 | 1.63 ± 0.06 | 5.32 ± 0.29 | 0.31 | 1.08 | 77.7 | 1.39 | 0.16 | 5560 | 0.03 | 0.12 | 0.28 | |
| 200 m | 21.59 | 34.90 | 1.16 ± 0.08 | 1.34 ± 1.09 | 13.61 ± 4.24 | 0.00 | 0.67 * | 100.0 | 0.67 | 0.06 | 2688 | 0.02 | 0.06 | 0.33 | |
| 500 m | 10.57 | 34.27 | 1 ± 0.03 | 0.48 ± 0.12 | 21.83 ± 5.11 | 0.00 | 0.17 * | 100.0 | 0.17 | 0.02 | 680 | 0.01 | 0.04 | 0.35 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, P.W.-Y.; Olivia, M.; Gong, G.-C.; Jan, S.; Tsai, A.-Y. Viral Dynamics in the Tropical Pacific Ocean: A Comparison between Within and Outside a Warm Eddy. Viruses 2024, 16, 937. https://doi.org/10.3390/v16060937
Chen PW-Y, Olivia M, Gong G-C, Jan S, Tsai A-Y. Viral Dynamics in the Tropical Pacific Ocean: A Comparison between Within and Outside a Warm Eddy. Viruses. 2024; 16(6):937. https://doi.org/10.3390/v16060937
Chicago/Turabian StyleChen, Patrichka Wei-Yi, Madeline Olivia, Gwo-Ching Gong, Sen Jan, and An-Yi Tsai. 2024. "Viral Dynamics in the Tropical Pacific Ocean: A Comparison between Within and Outside a Warm Eddy" Viruses 16, no. 6: 937. https://doi.org/10.3390/v16060937