Abstract
EBV and KSHV are both gamma-herpesviruses which express multiple viral microRNAs. Various methods have been used to investigate the functions of these microRNAs, largely through identification of microRNA target genes. Surprisingly, these related viruses do not share significant sequence homology in their microRNAs. A number of reports have described functions of EBV and KSHV microRNA targets, however only three experimentally validated target genes have been shown to be targeted by microRNAs from both viruses. More sensitive methods to identify microRNA targets have predicted approximately 60% of host targets could be shared by EBV and KSHV microRNAs, but by targeting different sequences in the host targets. In this review, we explore the similarities of microRNA functions and targets of these related viruses.
1. Introduction
Approximately twenty percent of human cancers are associated with various infectious agents [1]. Two such agents are Epstein-Barr virus (EBV/human herpesvirus 4) and Kaposi’s sarcoma-associated herpesvirus (KSHV/human herpesvirus 8). Other viruses directly linked to human cancers include hepatitis B virus (HBV), hepatitis C virus (HCV), human papillomavirus (HPV), human T-cell lymphotropic virus (HTLV-1), and Merkel cell polyomavirus. EBV and KSHV are members of the herpesvirus family that can infect lymphocytes and are associated with various proliferative disorders. EBV associated disorders include Burkitt’s lymphoma (BL), Hodgkin’s lymphoma (HL), NK/T-cell lymphoma, diffuse large B-cell lymphoma (DLBCL) post-transplant lymphoproliferative disorder, and nasopharyngeal carcinoma (NPC) (reviewed in [2]). KSHV infections are associated with Kaposi’s sarcoma, primary effusion lymphoma (PEL) and multicentric Castleman’s disease (MCD). Both gamma-herpesviruses, EBV and KSHV have evolved to maintain life-long latent infections in their human hosts. Both viruses can infect B-lymphocytes, while in vivo evidence shows that EBV can also infect epithelial cells and KSHV also infects endothelial cells.
The complex steps involved in viral oncogenesis have largely been characterized by studying the interactions between human and viral proteins. A new class of molecules- microRNAs, have begun to be appreciated in virus-host interactions starting with the discovery of virally-encoded microRNAs in EBV [3] and in other herpesviruses, including KSHV [4]. It is believed that EBV and KSHV microRNAs are processed in the same fashion as human microRNAs (though viral microRNA processing can follow a unique pathway in some viruses [4,5]). MicroRNAs have been reviewed extensively elsewhere [6], but briefly, they are short (~22 nt) single-stranded RNA molecules that arise as RNA polymerase II primary transcripts after a number of processing steps. Once the mature microRNAs are generated, they can be incorporated into the RNA-induced silencing complex (RISC), where they can target mRNAs through imperfect complementarity to suppress gene expression. Repression of target mRNAs can trigger destabilization of the mRNA followed by degradation and/or inhibit protein translation. In the context of latent viral infection, these are attractive molecules to modulate host gene expression since they require little viral genome space, avoid generation of additional potentially antigenic viral latent proteins, and are unlikely to be selectively targeted by the host, since humans may lack the ability to distinguish the viral miRNA sequences from human miRNA sequences. Viruses may utilize miRNAs to exploit a faster evolution rate given that a change of few nucleotides can dramatically change the repertoire of targeted mRNAs. Finally, recent studies with mutant strains of viruses, which lack certain miRNAs imply that viral miRNAs play important roles in maintaining the latent/lytic switch [7,8].
Multiple insightful reviews have covered the cellular and viral targets of viral miRNAs [9,10,11,12,13]. Here we focus on the cellular targets of EBV and KSHV miRNAs and look for similarities in the targets and functions of miRNAs from these related viruses. As a result of multiple deep-sequencing efforts, we currently feel we have discovered all of the EBV and KSHV microRNAs. Although the miRNAs from both these viruses lack sequence similarity, one similarity is in their genomic organization. Both viruses show clustering of the miRNA genes and most miRNAs come from the same starting primary miRNA transcript. In KSHV, the 12 microRNA genes are largely in one cluster in the latency locus while the 25 microRNAs of EBV, are largely in two clusters- BART miRNAs and three BHRF1 miRNAs [14].
A variety of approaches have been employed to identify the targets of miRNAs. Bioinformatic methods can be used to find targets through identification of mRNA sequences that are complementary to the seed region of miRNAs (reviewed in [15]). Frequently, miRNA targets are identified by scanning the 3’ untranslated regions (UTRs) of mRNAs for sequences complementarity. Additionally, gene expression studies can be used to find gene products that are altered when viral miRNAs are introduced or inhibited [16,17,18]. Recent methods utilize microarrays or the sensitivity of deep sequencing to find mRNA sequences that are biochemically purified with the RNA-induced silencing complex (RISC) [19,20,21,22]. In crosslinking and immunoprecipitation (CLIP) assay, miRNAs and their targets are UV crosslinked to the RISC proteins. The ternary Ago-miRNA-mRNA complexes are then purified by immunoprecipitation of the Ago protein. Immunoprecipitated mRNAs are converted to cDNA and analyzed by high throughput sequencing (HITS-CLIP). HITS-CLIP generates a genome wide-map of miRNA binding sites, but the direct position of interaction between mRNA and protein is not precisely identified. To enhance the poor crosslink efficiency between RNAs and proteins provided by UV irradiation, neo-synthesized RNAs could be labeled with photoactivatable ribonucleoside enhanced crosslinking and immunoprecipitation (PAR-CLIP) such as 4-thiouridine (4-SU). Furthermore, crosslinked 4-SU induces a thymidine to cytidine transition during the process and consequently marks the position of successfully crosslinked sequences. Comparing CLIP reads between non-infected and infected cells allows the determination of specific targets of viral miRNAs. However, to identify the precise miRNA binding sites a bio-informatic analysis based on seed sequence homology is often required. Each method has inherent advantages and limitations, but the combination of these approaches have been successful in identifying many miRNA targets (Table 1). After identifying miRNA targets using bioinformatic or cell culture-based screening methods, a variety of assays are used to validate predicted miRNA targets. The most common way to validate a miRNA binding site is to clone this binding site in the 3’UTR of a luciferase reporter and to express this reporter in the presence of the miRNA. An effective binding of the miRNA to a target mRNA then represses luciferase activity of the reporter. These assays are essentially identical to validation studies performed with cellular miRNAs, with the advantage that the uninfected cells are ideal control cells for introducing viral miRNAs. Additionally, the numbers of different viral miRNAs expressed in an infected cell is experimentally more manageable than the hundreds of cellular miRNAs expressed in a given cell type.
Here, we explore the similarities and differences in the functions of miRNA targets from the limited number of experimentally validated miRNA targets and the larger number of targets predicted by recent reports using high-throughput sequencing of RNA isolated by crosslinking immunoprecipitation (HITS-CLIP and PAR-CLIP) [20,21,22] (Table 2). The exact significance of these targets needs to be further investigated to understand their potential roles in infection and viral-host interactions. Nevertheless, some main cellular functions emerged from the miRNA targets of EBV and KSHV miRNAs.

Table 1.
List of experimentally validated cellular targets of KSHV- and EBV- encoded miRNAs. The normal cellular functions of these proteins and the effect of their knockdown during viral infection are also listed. The targets that are underlined are repressed by both KSHV- and EBV-encoded miRNAs. ND- No data.
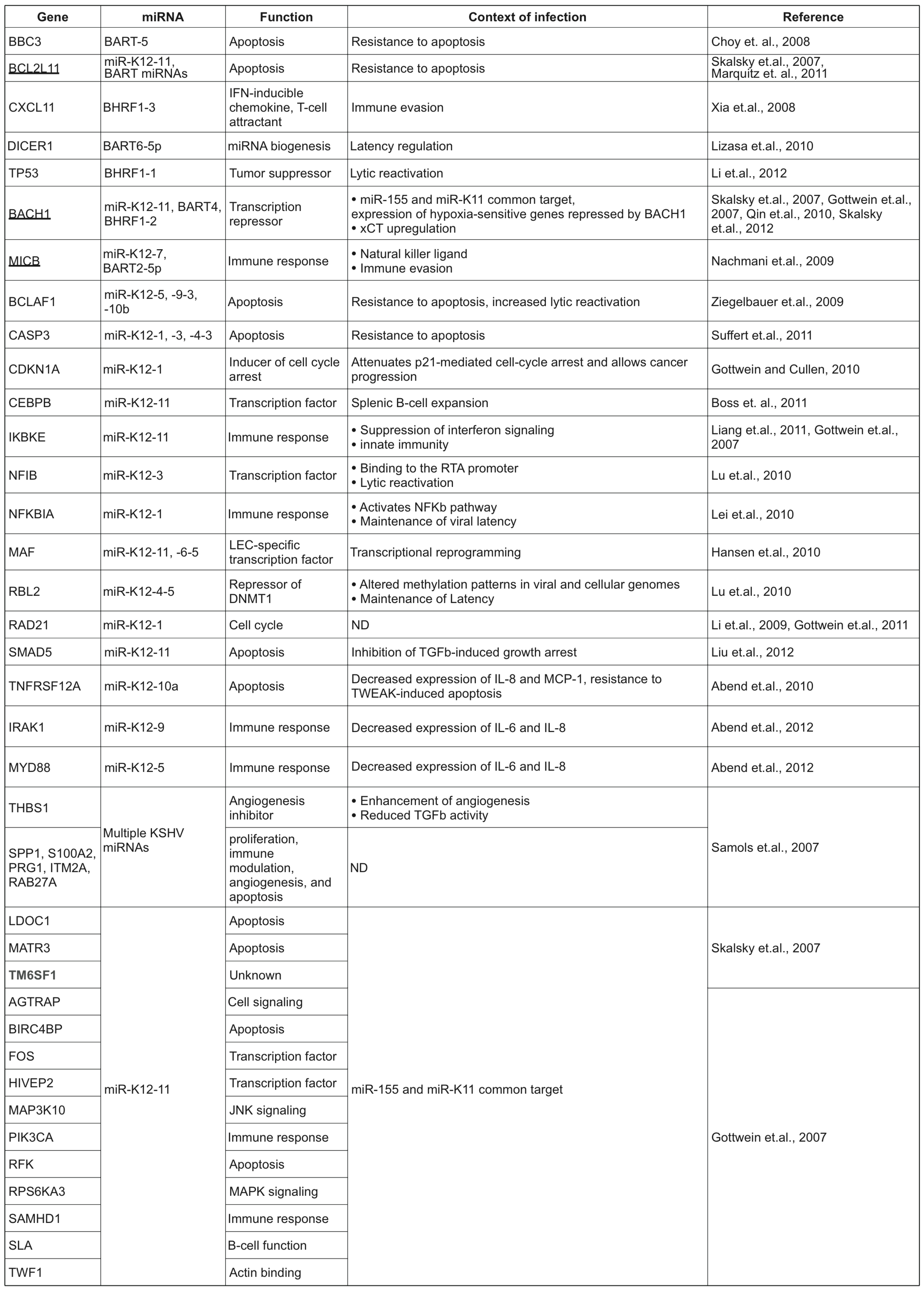 |
Table 2.
Validated targets from high-throughput studies. Shown are cellular targets identified by photoactivatable ribonucleoside enhanced crosslinking and immunoprecipitation (PAR-CLIP), High-throughput sequencing of RNA isolated by crosslinking immunoprecipitation (HITS-CLIP) or RNA immunoprecipitation and microarray (RIP-Chip) techniques. Validation of these targets was done using luciferase reporter assays.
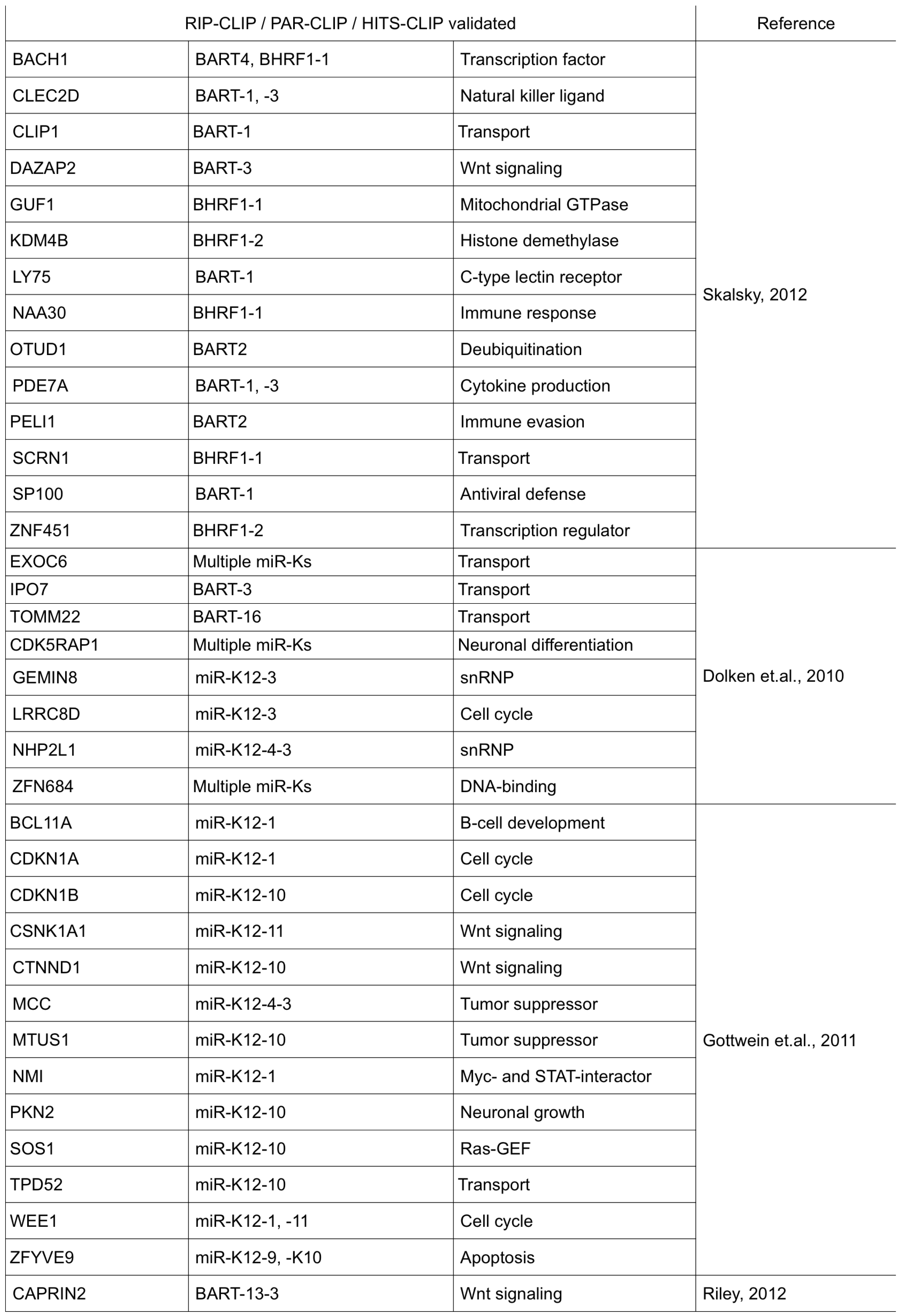 |
3. Regulation of apoptosis by KSHV and EBV miRNAs
Resistance to apoptosis is a common mechanism evolved by many viruses to persist in the host and many cellular targets of KSHV and EBV miRNAs are involved in apoptotic pathways. Using a microarray-based approach to identify mRNAs whose levels were repressed in the presence of KSHV miRNAs, BCLAF1 (Bcl-2-associated factor), a pro-apoptotic protein, was identified as a target of miR-K12-5, -9, -3 and -10b. The repression of BCLAF1 by KSHV miRNAs enabled the cells to overcome etoposide-induced caspase activation [18]. Virally encoded miRNAs can also attenuate cellular responses to extracellular signals as demonstrated in the case of TNFRSF12A (TWEAKR), which is the cellular receptor for the TNF-like weak inducer of apoptosis (TWEAK). Knockdown of TWEAKR levels by miR-K12-10a protected cells from TWEAK-induced apoptosis and also reduced the levels of the cytokines- IL-8 and MCP-1, secreted in response to TWEAK-induction [37]. In another study, caspase 3 was itself identified as a target of multiple KSHV miRNAs, which conferred resistance to apoptosis [38]. In addition, many proteins involved in apoptosis regulation like the tumor suppressors- p53 (for EBV- BHRF1-1) [39], LDOC1 (leucine zipper down-regulated in cancer 1; KSHV miR-K12-11) [40] and MTUS1 (microtubule associated tumor suppressor 1) [20] have been validated. KSHV miR-K12-1 down-regulated the levels of the cyclin-dependent kinase inhibitor, p21, thereby allowing the infected cells to overcome p21-mediated cell cycle arrest [41]. Further, HITS-CLIP and PAR-CLIP studies have identified numerous proteins involved in Wnt signaling (CAPRIN2, CTNND1, DAZAP2, CSNK1A1) as targets of KSHV and EBV-encoded miRNAs [20,21,22].
The pro-apoptotic proteins, BIRC4BP/XAF1 (XIAP associated factor), PUMA (p53-upregulated modulator of apoptosis) and BCL2L11 (or Bim) were also down-regulated by KSHV or EBV miRNAs [17,40,42]. PUMA and Bim are BH3-only proteins of the Bcl-2 family that inhibit the anti-apoptotic functions of Bcl-2 and knockdown of these proteins by EBV-BART-miRNAs conferred cell survival [42,43,44]. Further, both PUMA and Bim are activated by two transcription factors- p53 and C/EBPβ, that are themselves regulated by viral/ virally-induced miRNAs (see below). Interestingly, Bim was repressed by both KSHV and EBV miRNAs, demonstrating the significance of Bim knockdown in the biology of these viruses [40,44].
The TGF-β pathway confers a strong anti-proliferative phenotype to many epithelial and endothelial cells and hence, it is presumable that KSHV has developed mechanisms to escape from this inhibition. SMAD5, the downstream effector of TGF-β is repressed by KSHV- miR-K12-11, and this allowed the cells to overcome the cytostatic effects mediated by TGF-β and enhanced cell proliferation [45]. In addition, thrombospondin 1 (THBS1) (KSHV; see below) and DAZAP2 (EBV; BART3) are proteins of the TGF-β pathway that are repressed by viral miRNAs [16,21]. The TGF-β pathway is also down-regulated via a different mechanism by KSHV. The LANA protein was shown to inhibit TGFβ-type II receptor expression by epigenetic silencing in PEL cells [46]. This is an example of synergy between latently expressed miRNAs and proteins to modulate the same cellular pathway. This redundancy will likely be observed with other miRNA targets as viruses routinely use redundant mechanisms. An additional method of promoting growth is by EBV modulating expression of the human miRNA, miR-34a [82].
7. Similarities between miR-155 and miR-K12-11 in the context of KSHV/ EBV infection
One interesting discovery in the study of virally encoded miRNAs was the identification of the KSHV miRNA, miR-K12-11, as the viral ortholog of the cellular miRNA, hsa-miR-155 (miR-155) [17,40]. Two independent groups showed that miR-K12-11 and miR-155 have the exact ‘seed’ sequence and therefore, could regulate a similar set of genes. The common set of genes that were down-regulated by both miRNAs included BACH1 (BTB and CNC homology 1), a transcription repressor involved in the regulation of genes regulating cell cycle and oxidative stress [59]. In addition, by microarray profiling and luciferase assays, the authors validated LDOC1 (leucine zipper down-regulated in cancer 1), MATR3 (Matrin 3), TM6SF1 (transmembrane 6 superfamily member 1), PIK3CA (phosphoinositide-3-kinase, catalytic α subunit), XAF1, HIVEP2 (HIV-1 enhancer binding protein 2) and Fos as common targets of both miRNAs [17,40].
In order to identify miR-K12-11 as a true functional ortholog of miR-155, Boss et.al. stably expressed either miR-K12-11 or miR-155 in CD34+ human cord blood progenitor cells and studied immune reconstitution using the NOD/LtSz-scid IL2Rγnull mouse model [60]. These mice have neither circulating complement nor functional NK, B, T, APC cells, and they are deficient in cytokine signaling. Therefore, they can be humanized by transplantation of human cord blood derived CD34+ progenitors and use as an in vivo model for KSHV infection. It was found that the expression of miR-K12-11 or miR-155 repressed the levels of C/EBPβ, a transcription factor that negatively regulates IL-6 and contributes to the development of many B-cell lymphomas. Further, this repression of C/EBPβ by either miR-K12-11 or miR-155 resulted in an increased proliferation of B-cells in the spleen, contributing to lymphomagenesis [60]. More recently, it was shown that SMAD5 is a direct target of both miR-K12-11 [45] and miR-155 [61]. The repression of SMAD5 by both miRNAs was demonstrated to overcome TGFβ -induced growth arrest, thereby, leading to increased cell division and tumor development [45,61].
miR-155 is an oncomiR found to be upregulated in many B-cell lymphomas [62,63]. Interestingly, EBV, which does not encode an ortholog of miR-155, upregulates miR-155 upon infection and this is important for B-cell immortalization [64]. Further, the α-herpesvirus, Marek’s Disease Virus (MDV) expresses MDV-miR-M4, an ortholog of miR-155. The two miRNAs were shown to regulate a common set of genes including C/EBPβ and HIVEP2, that are also validated targets of KSHV-miR-K12-11 [65]. Together, these studies suggest that the viral orthologs of hsa-miR-155 may perform overlapping functions and the deregulation of the target genes of these miRNAs might contribute to the development of B-cell lymphomas.
8. Other targets of KSHV and EBV encoded miRNAs
In addition to repressing factors involved in apoptosis, immune evasion and cell cycle arrest, several proteins involved in splicing, miRNA biogenesis and transcription factors have been identified as targets of KSHV and EBV miRNAs. In RIP-CHIP analysis of KSHV infected cells, two proteins of the small nuclear ribonucleoprotein (snRNP) family- NHP2L1 (non-histone chromosome protein 2-like 1) and GEMIN8 (gem (nuclear organelle) associated protein 8), were repressed by KSHV miRNAs, miR-K12-4-3 and -3, respectively. Interestingly, the binding sites for these two miRNAs were found to be in the coding regions of these two genes. In the same study, the authors also identified the CDK5 inhibitor, CDK5RAP1 (CDK5 regulatory subunit associated protein 1), which is predominantly involved in neuronal differentiation and neuroskeletal differentiation, as a target of KSHV miRNA [19]. As mentioned earlier, Dicer, the key enzyme involved in miRNA biogenesis, was also identified as the target of EBV-BART6-5 [47].
In addition, several transcription factors were identified as targets of KSHV miRNAs, among which, ZFN684 [19], BACH1 [17,40], HIVEP2 and Fos [17] have been validated. Transcription factors were among the most enriched family of proteins in PAR-CLIP analyses of both KSHV and EBV infected cells [20]. KSHV infection of lymphatic endothelial cells (LECs) causes transcriptional reprogramming and induces the expression of markers specific for blood vessel endothelial cells (BECs) [66]. Down-regulation of the LEC-specific transcription repressor, MAF, by KSHV miRNAs- miR-K12-6 and miR-K12-11, was shown to induce the expression of several BEC-specific markers like CXCR4, thereby contributing to transcriptional reprogramming [67].
9. Overlap of targets from KSHV and EBV high-throughput miRNA studies
More recently miRNA targets have been predicted using more sensitive approaches of high-throughput sequencing of RISC-associated mRNAs in KSHV and/or EBV infected cell lines [20,68,69]. These methods yield short mRNA sequences that are associated with RISC and through searching these sequences for seed-matching sequences, an association between a specific mRNA and miRNA can be predicted. All of these reports have shown the ability to use this technology to identify validated miRNA targets using 3’UTR luciferase assays and Western blot assays. Here we elucidate the similarities among the genes predicted to be targets from these studies. However, we share the limitations of this analysis. First, the definition of a predicted target differs across the various reports. Second, the completeness of predicted targets made publicly available differed between the reports. Third, the mRNAs predicted to be viral targets are dependent on the sequencing reads possessing a perfect seed-matching site. This raises the possibility that certain mRNA targets being targeted by viral miRNAs using non-canonical seed-matching are not included in the list of predicted hits. Additionally, if a human miRNA having the same seed-match site as a viral miRNA targets an mRNA, then the mRNA would be erroneously labeled as a target of both miRNAs. This analysis does not distinguish miRNA sequences beyond the seed region. Finally, long 3’UTRs, certain secondary structures, and base composition may influence transient, but non-biologically relevant cross-linking events resulting in a predicted miRNA target site. Nevertheless, it is a worthwhile exercise to investigate the similarities and differences of the predicted targets using an additional filter for finding viral miRNA targets, to reveal similar viral-host interactions, and to possibly identify differences in the virus-host interactions between KSHV and EBV. While the specific target sites are unlikely to be conserved between these viruses, recent data suggests a significant overlap of miRNA targets of these viruses. The specific genes targeted and the classes of targets may improve our understanding of viral oncogenesis and immune evasion.
The overlap of targets of EBV and KSHV miRNAs has been approached using biochemical purification of RISC followed by microarrays to measure mRNAs associated with RISC in EBV and KSHV infected cell lines [70]. This analysis predicted 114 targets for KSHV miRNAs and 44 targets for EBV miRNAs. Analysis of these predicted targets revealed only three genes were predicted as miRNA targets for both KSHV and EBV miRNAs (PDCD1LG2, JARID1B, and SOCS3). These genes have been shown to be involved with immune response [71], protein demethylation [72], and cytokine responses [73]. KSHV miRNA targets showed an enrichment of gene ontology terms for genes involved in splicing, gene expression and protein import into the nucleus [70].
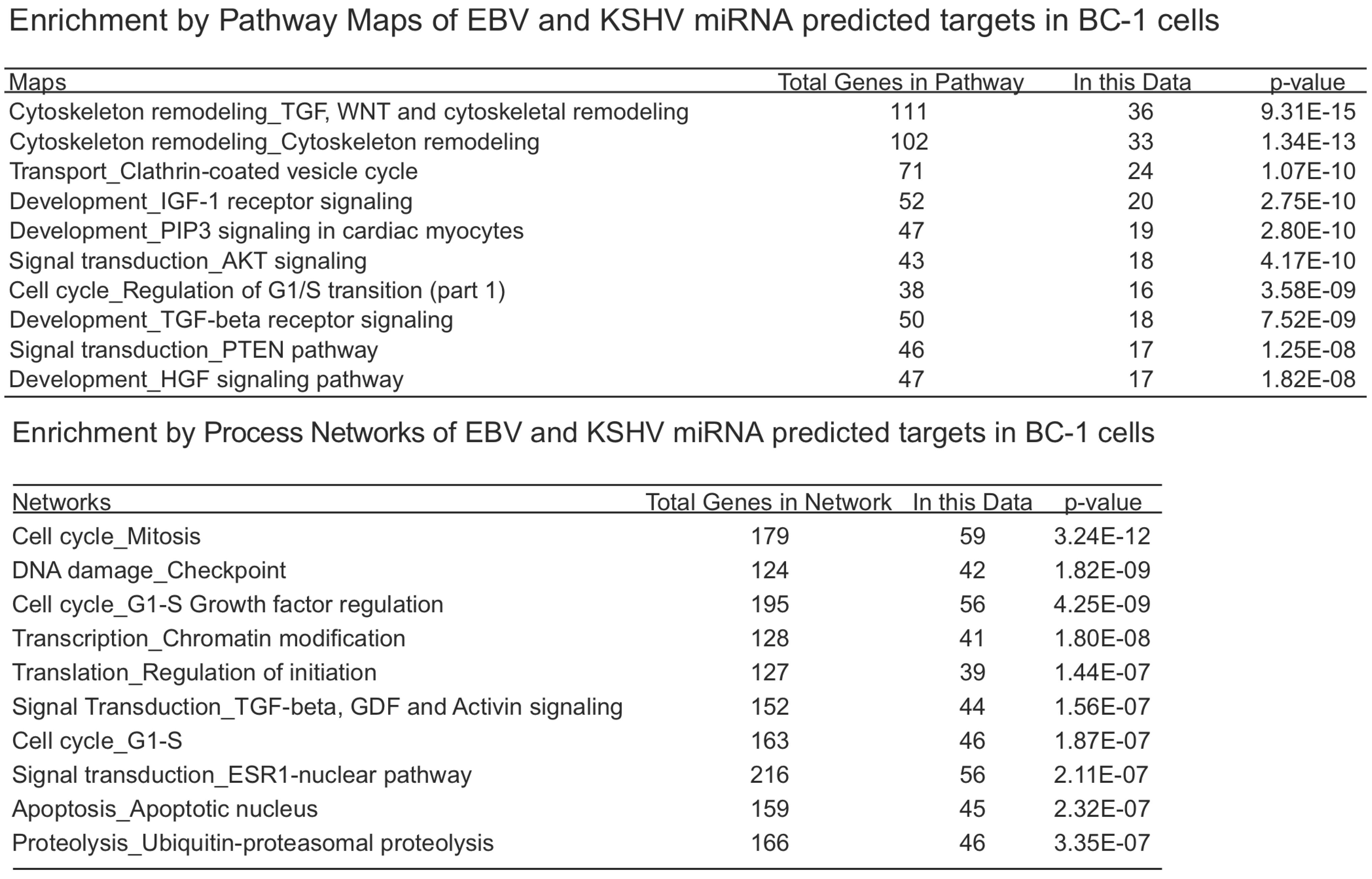
We investigated the overlapping genes across the various lists of predicted targets from PAR-CLIP [20,68] and HITS-CLIP [69] assays. For the first analysis, we considered the KSHV and EBV miRNAs that are all naturally expressed in the primary effusion cell line, BC-1 (which do not express the BHRF1 EBV miRNAs). In the original report using only 3’UTR predicted targets, it was observed that fifty-eight percent of KSHV targets were also predicted to be targets of EBV miRNAs. In the analysis here, we included all coding sequence (CDS) and 3’UTR predicted targets. Of the KSHV miRNA predicted targets (3796 genes), sixty-four percent were also EBV predicted targets (Figure 1). Likewise, of the EBV miRNA predicted targets (4328 genes), fifty-six percent are also KSHV predicted targets. Initially, this high degree of correlation may seem surprising since there are no KSHV and EBV miRNAs which share the same seed region. However, this suggests KSHV and EBV miRNAs are both targeting a majority of similar genes through different target regions within the same transcripts and these viral miRNAs are playing similar roles in modulating gene expression. Using the 2423 overlapping genes predicted to be targets of EBV and KSHV miRNAs in the co-infected BC-1 cell line, we analyzed this list for enrichment of certain pathways or process networks (manually curated gene sets based on pathways and gene ontology terms; similar to gene set enrichment analysis) in a process similar to measuring enrichment of gene ontology terms (using MetaCore from Thomson Reuters). As shown in Table 3, there was a significant enrichment of pathways involved in cytoskeleton remodeling. Endothelial cells undergo morphological changes to form spindle cells upon KSHV infection [74,75]). Additionally, KSHV utilizes clathrin-mediated endocytosis and actin remodeling an important role in KSHV entry into cells [76]. Another pathway that is enriched in the EBV and KSHV targets is the PTEN pathway. The predicted targets in this pathway include p21 [41], p53, PTEN, caspase-9, FOXO3, and others. A KSHV protein, LANA, has been shown to inhibit p53 [77], suggesting that KSHV latent proteins and miRNAs can cooperate to inhibit p53. FOXO3 was found to be poorly expressed in nasopharyngeal carcinoma tissues and may represent an important marker for this EBV-related cancer [78]. In the enriched process networks in Table 3, the networks related to cell cycle control and apoptosis are expected based on earlier work on p21 [41], caspase 3 [38] and others [18]. Perhaps more surprising, was the network of genes involved in translation initiation. While translation inhibition upon viral infection has been investigated since 1964 [79], it has been generally described during lytic infection (unlike the latent infection in BC-1 cells). Inspection of the overlapping genes revealed twelve genes from the eukaryotic translation initiation family. Together, this analysis reveals some common themes of predicted KSHV and EBV miRNA targets in BC-1 cells.
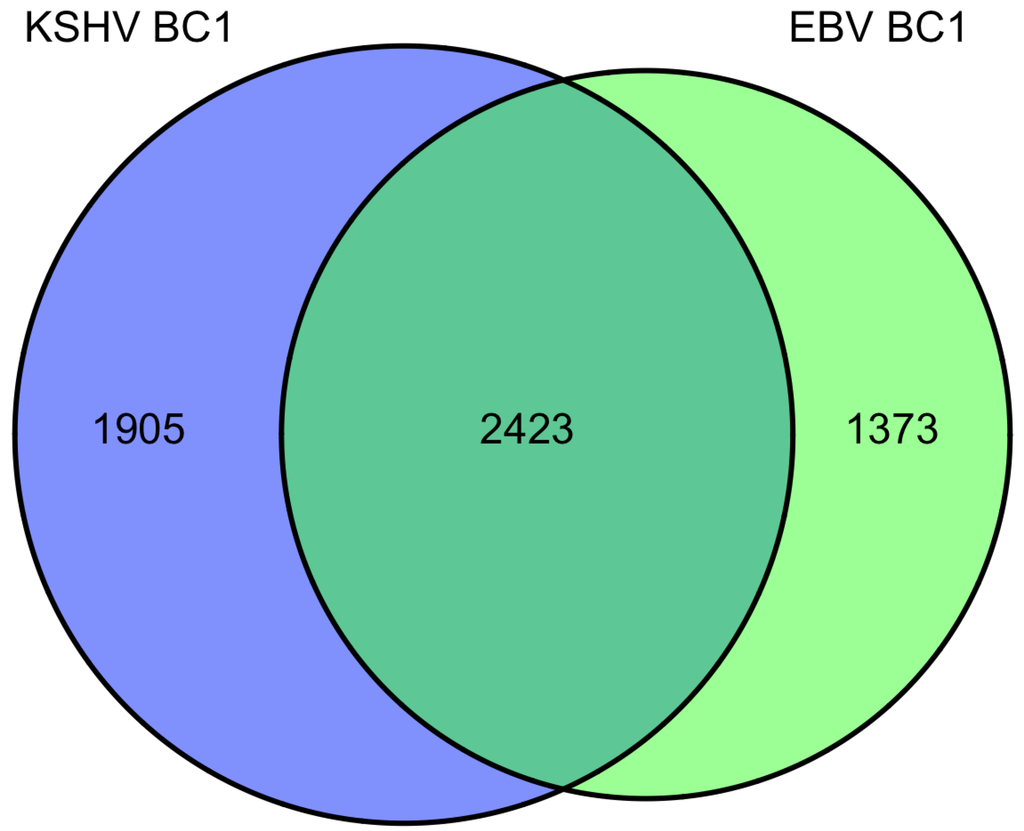
Figure 1.
Venn diagram of BC1 predicted miRNA targets from Gottwein et al. 2011.
Figure 1.
Venn diagram of BC1 predicted miRNA targets from Gottwein et al. 2011.
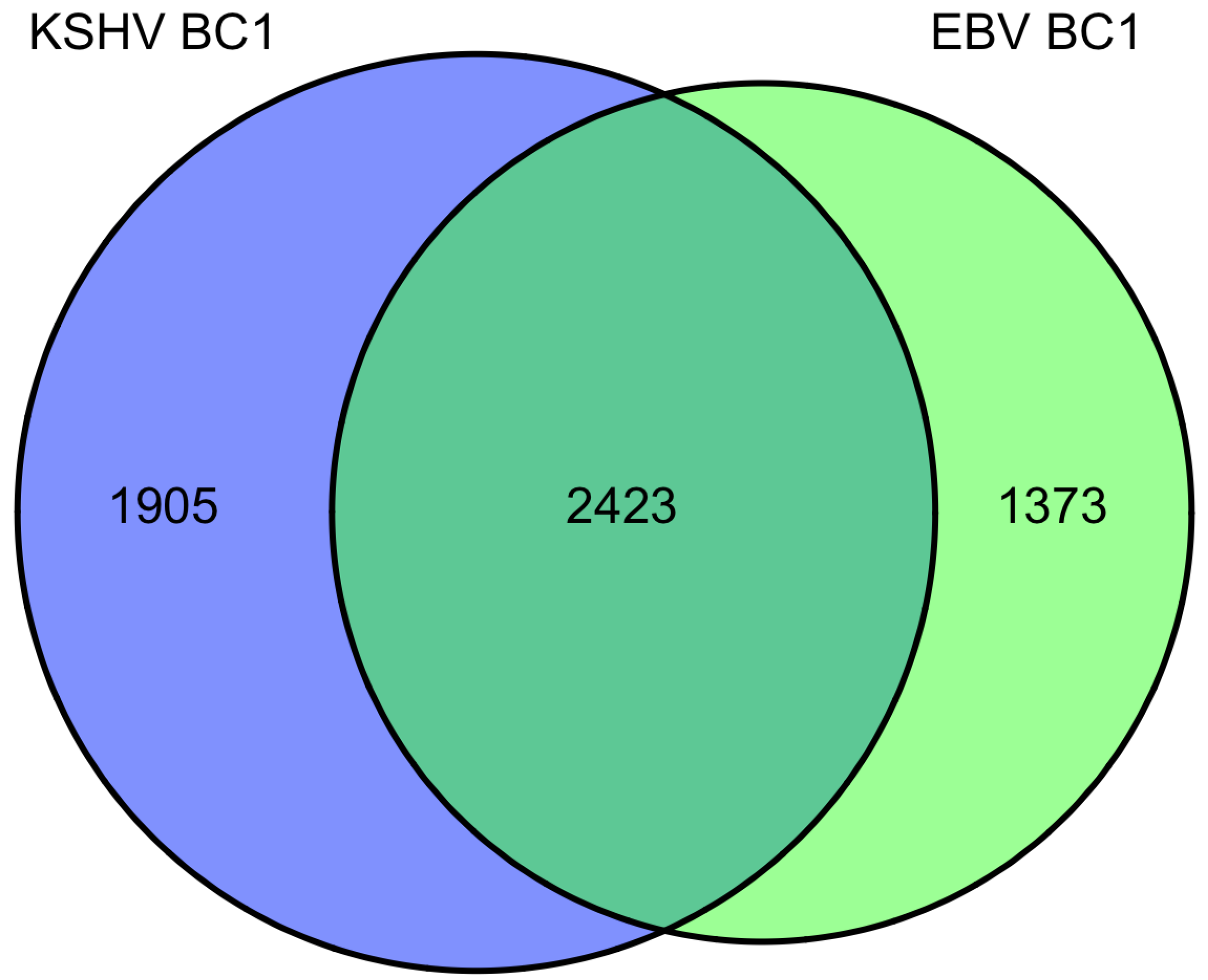
Table 3.
BC1 overlapping miRNA target associations.
 |
As mentioned above, there are four datasets from PAR-CLIP or HITS-CLIP assays looking at EBV and KSHV miRNA targets. As shown in Figure 2, there were 31 genes that were found in all four datasets. Given, the differences between the methods used, arbitrary cutoffs, miRNA expression differences among different cell lines, we sought to investigate a broader group of genes. We selected 478 genes (472 are predicted KSHV miRNA targets) that were found in at least three of the four datasets. Using a similar analysis as above, we analyzed this set for functional themes. Again, we observed enrichment of EBV and KSHV predicted miRNAs involved in cell cycle regulation, apoptosis, cytoskeleton remodeling (Table 4). Additionally, we observed a theme of predicted targets involved in immune responses, specifically genes in the B-cell receptor pathway, interleukin regulation, and the interferon pathway. This analysis highlighted interferon receptor 1 and 2 as predicted miRNA targets in all of the datasets, except the EBV Jijoye dataset [22]. But, it should be noted that all of the predicted targets from this dataset were not publicly available, raising the possibility that this was predicted in this dataset as well. Closer analysis shows the PAR-CLIP analysis yielded RISC-associated clusters that are distinct for each viral miRNA, yet in a same region (10% of the 3’UTR sequence length) near the 5’end of the 3’UTR (Figure 3). Could this reflect a region of the mRNA that is less bound by obstructing RNA binding proteins and/or mRNA structures that are more accessible to RISC? Answering these questions could improve bioinformatic methods to identify miRNA targets.
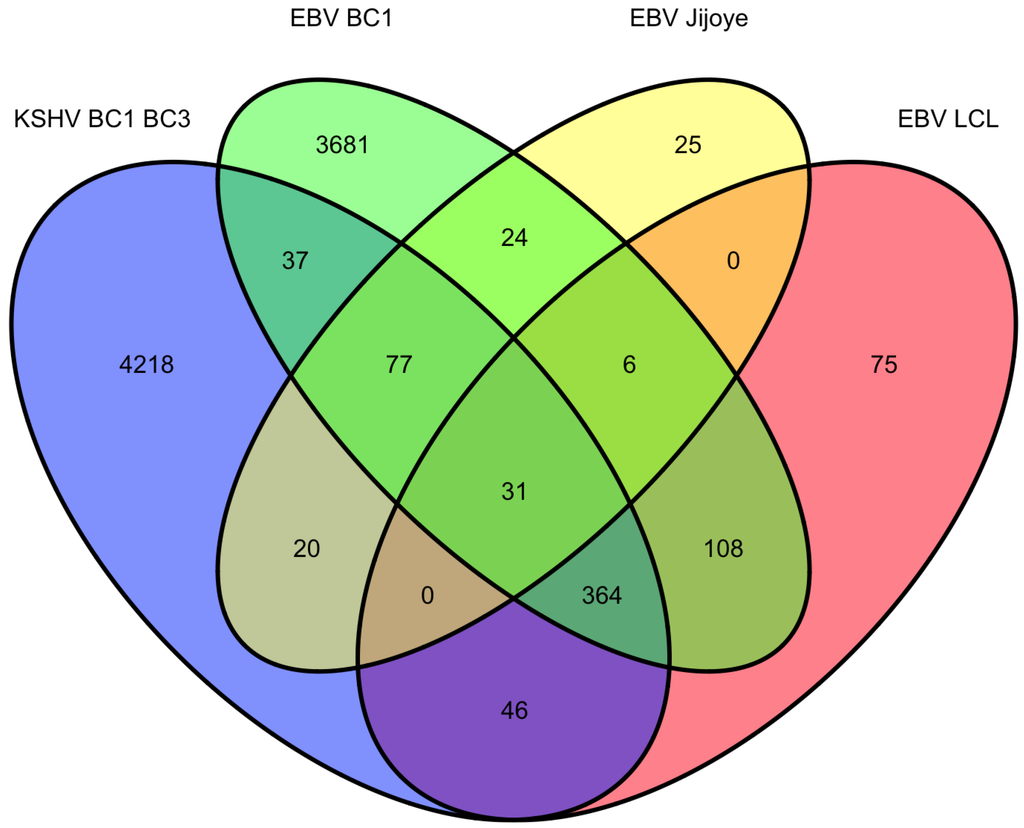
Figure 2.
Venn diagram of BC1 predicted miRNA targets from Gottwein et al. 2011.
Figure 2.
Venn diagram of BC1 predicted miRNA targets from Gottwein et al. 2011.
Table 4.
Three of four overlapping miRNA target associations. Shown are the enriched associations of miRNA targets identified in at least three of the four CLIP datasets shown in Figure 2.
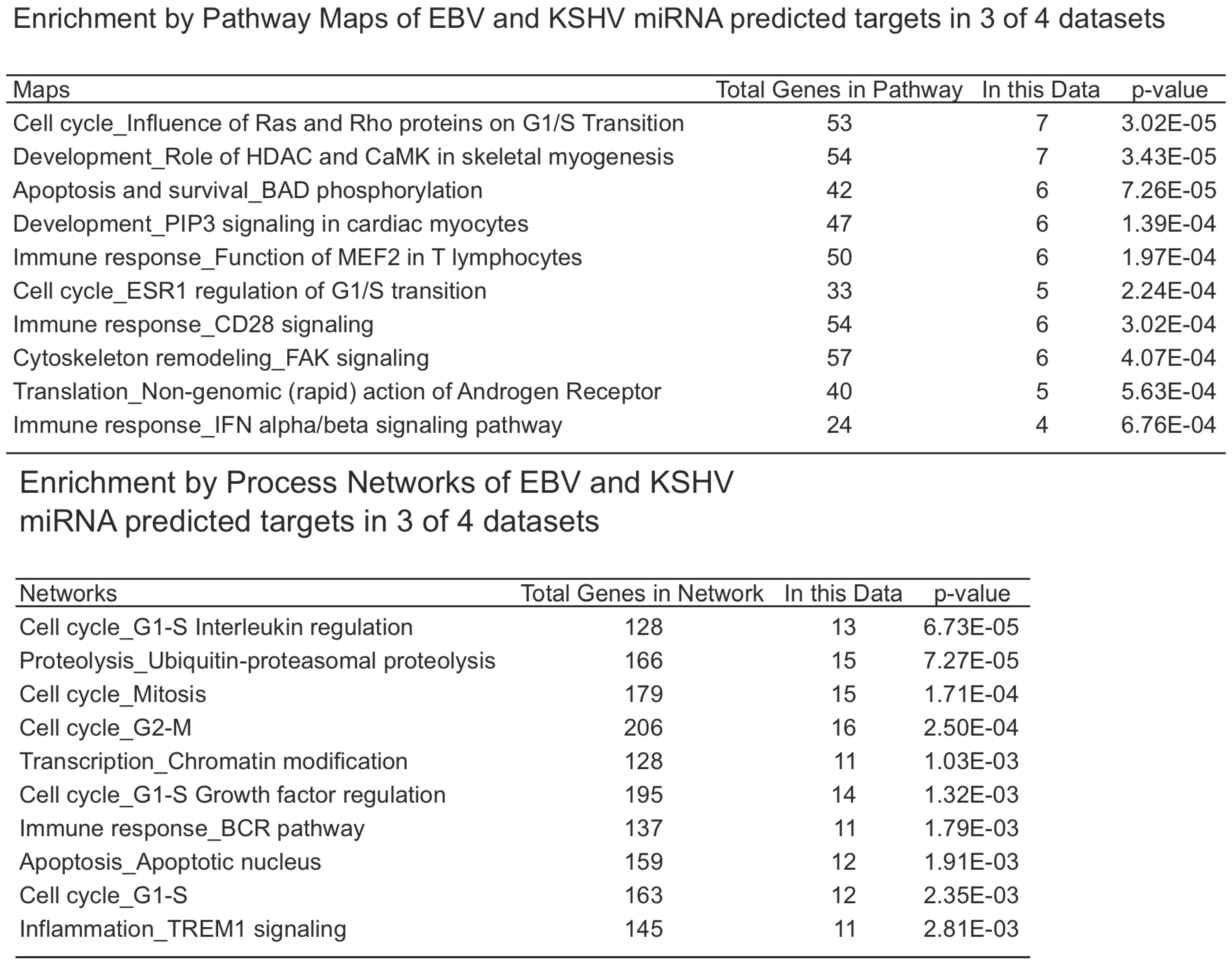 |
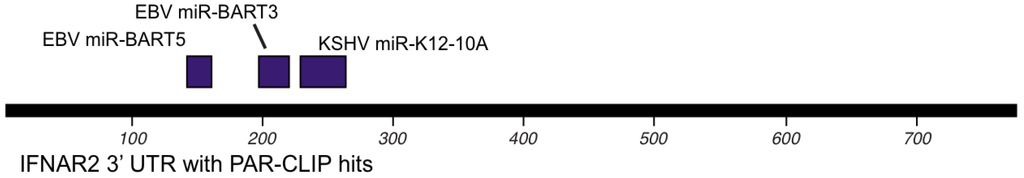
Figure 3.
Map of interferon receptor 2 (IFNAR2) 3’UTR with miRNA target sites as determined by CLIP assays.
Figure 3.
Map of interferon receptor 2 (IFNAR2) 3’UTR with miRNA target sites as determined by CLIP assays.
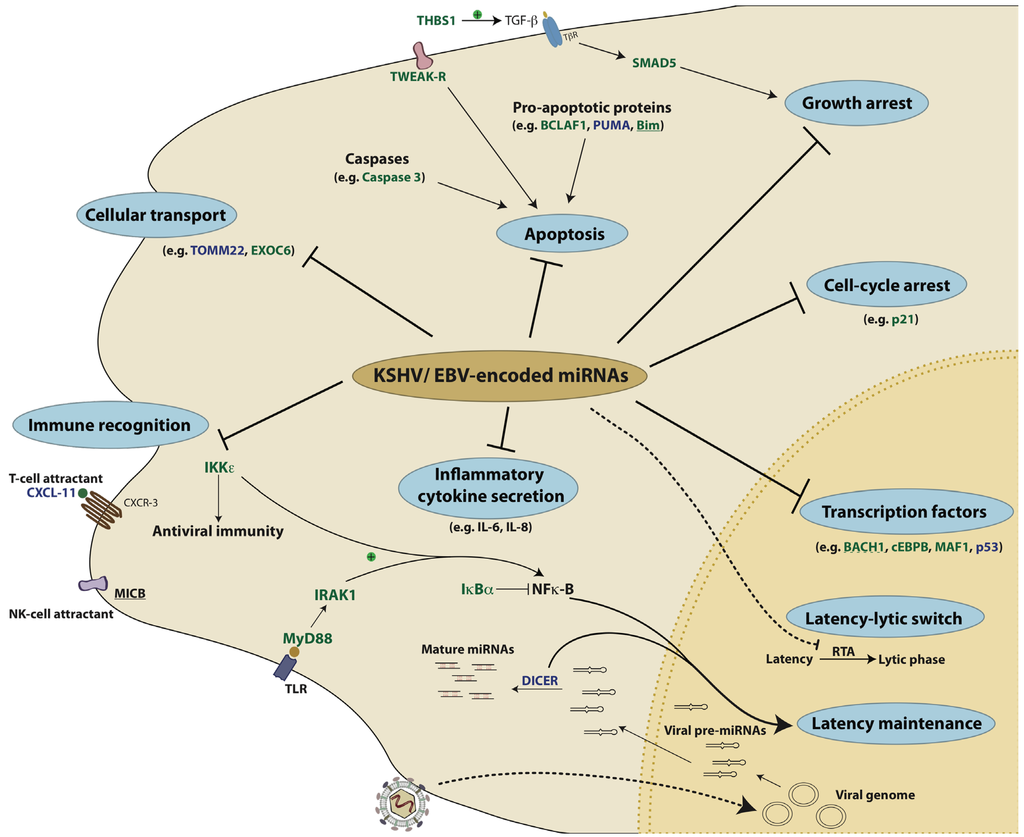
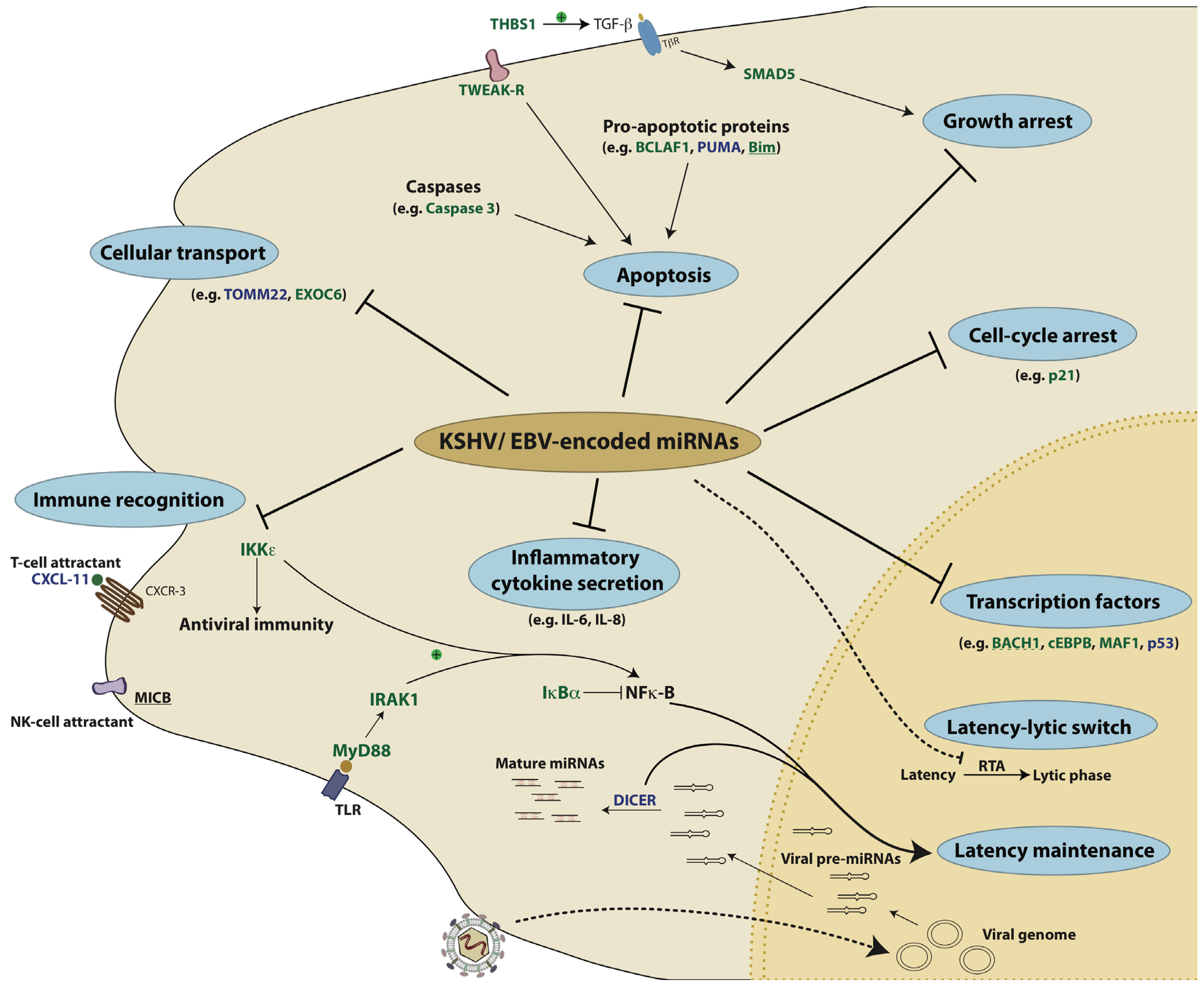
Figure 4.
KSHV- and EBV-encoded miRNAs target several key pathways to establish disease. 1) Viral miRNAs can inhibit pro-apoptotic proteins like Bim and BCLAF1. 2) Viral miRNAs can also enable the infected cells to evade recognition by the immune system by repressing either the T-cell attracting chemokine, CXCL-11 or the NK-cell attracting ligand, MICB. 3) miRNAs can also inhibit the production of interferons and inflammatory cytokines by repressing factors like IKKε, IRAK1 and MyD88. 4) By repressing factors like IκBα and Dicer, viral miRNAs can also regulate latency. In addition to cellular targets, the RTA protein of KSHV that acts as the master regulator of the latency-lytic switch, is itself a target of KSHV miRNAs. 5) Several transcription factors like BACH1, c/EBPβ and p53 are repressed by KSHV and EBV miRNAs. 6) KSHV miRNAs repress p21 and SMAD5 to overcome p21- and TGFβ- mediated growth arrest, respectively. In this illustration, the targets of KSHV miRNAs are in green, those of EBV miRNAs are in blue and targets that are repressed by both KSHV and EBV are underlined. The dotted line represents a viral target of KSHV-encoded miRNAs.
Figure 4.
KSHV- and EBV-encoded miRNAs target several key pathways to establish disease. 1) Viral miRNAs can inhibit pro-apoptotic proteins like Bim and BCLAF1. 2) Viral miRNAs can also enable the infected cells to evade recognition by the immune system by repressing either the T-cell attracting chemokine, CXCL-11 or the NK-cell attracting ligand, MICB. 3) miRNAs can also inhibit the production of interferons and inflammatory cytokines by repressing factors like IKKε, IRAK1 and MyD88. 4) By repressing factors like IκBα and Dicer, viral miRNAs can also regulate latency. In addition to cellular targets, the RTA protein of KSHV that acts as the master regulator of the latency-lytic switch, is itself a target of KSHV miRNAs. 5) Several transcription factors like BACH1, c/EBPβ and p53 are repressed by KSHV and EBV miRNAs. 6) KSHV miRNAs repress p21 and SMAD5 to overcome p21- and TGFβ- mediated growth arrest, respectively. In this illustration, the targets of KSHV miRNAs are in green, those of EBV miRNAs are in blue and targets that are repressed by both KSHV and EBV are underlined. The dotted line represents a viral target of KSHV-encoded miRNAs.
10. Conclusions
As time progresses, more discoveries will be shared about additional miRNA targets. Additionally, how these targets interact with each other and with viral proteins remains to be further studied. Future research will likely include investigating the roles of exosomes in manipulating the host outside of infected cells. Furthermore, understanding the roles of viral miRNAs during infection in vivo remains to be a challenge for the field and will likely be complicated by redundant functions of miRNAs and possibly viral proteins. One goal for the field is beyond identifying miRNA targets, but using viral miRNAs as tools to show us humans what the virus already knows is important for repression. A deeper understanding of the networks of miRNA targets (Figure 4) will improve our understanding of viral pathogenesis and yield new insights into the functions of host proteins and virus-host interactions.
Acknowledgments
This work was supported by the Intramural Research Program of the Center for Cancer Research, National Cancer Institute, National Institutes of Health.
Conflict of Interest
The authors declare no conflict of interest.
References and Notes
- Zur Hausen, H. The search for infectious causes of human cancers: where and why. Virology 2009, 392, 1–10. [Google Scholar] [CrossRef]
- Raab-Traub, N. EBV-induced oncogenesis. In Human Herpesviruses: Biology, Therapy, and Immunoprophylaxis; Arvin, A, Campadelli-Fiume, G, Mocarski, E., Eds.; Cambridge University Press: Cambridge, 2007; Chapter 55. [Google Scholar]
- Pfeffer, S.; Zavolan, M.; Grasser, F.A.; Chien, M.; Russo, J.J.; Ju, J.; John, B.; Enright, A.J.; Marks, D.; Sander, C.; Tuschl, T. Identification of virus-encoded microRNAs. Science 2004, 304, 734–736. [Google Scholar] [CrossRef]
- Pfeffer, S.; Sewer, A.; Lagos-Quintana, M.; Sheridan, R.; Sander, C.; Grasser, F.A.; van Dyk, L.F.; Ho, C.K.; Shuman, S.; Chien, M.; Russo, J.J.; et al. Identification of microRNAs of the herpesvirus family. Nat. Methods 2005, 2, 269–276. [Google Scholar] [CrossRef]
- Kincaid, R.P.; Burke, J.M.; Sullivan, C.S. RNA virus microRNA that mimics a B-cell oncomiR. Proc. Natl. Acad. Sci. USA 2012, 109, 3077–3082. [Google Scholar]
- Bartel, D.P. MicroRNAs: target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef]
- Lu, F.; Stedman, W.; Yousef, M.; Renne, R.; Lieberman, P.M. Epigenetic regulation of Kaposi's sarcoma-associated herpesvirus latency by virus-encoded microRNAs that target Rta and the cellular Rbl2-DNMT pathway. J. Virol. 2010, 84, 2697–2706. [Google Scholar] [CrossRef]
- Lei, X.; Bai, Z.; Ye, F.; Xie, J.; Kim, C.G.; Huang, Y.; Gao, S.J. Regulation of NF-kappaB inhibitor IkappaBalpha and viral replication by a KSHV microRNA. Nat. Cell Biol. 2010, 12, 193–199. [Google Scholar] [CrossRef]
- Marquitz, A.R.; Raab-Traub, N. The role of miRNAs and EBV BARTs in NPC. Semin. Cancer Biol. 2012, 22, 166–172. [Google Scholar] [CrossRef]
- Cullen, B.R. Viruses and microRNAs: RISCy interactions with serious consequences. Genes Dev. 2011, 25, 1881–1894. [Google Scholar] [CrossRef]
- Grundhoff, A.; Sullivan, C.S. Virus-encoded microRNAs. Virology 2011, 411, 325–343. [Google Scholar] [CrossRef]
- Plaisance-Bonstaff, K.; Renne, R. Viral miRNAs. Methods Mol. Biol. 2011, 721, 43–66. [Google Scholar] [CrossRef]
- Gottwein, E.; Cullen, B.R. Viral and cellular microRNAs as determinants of viral pathogenesis and immunity. Cell Host Microbe. 2008, 3, 375–387. [Google Scholar] [CrossRef]
- Walz, N.; Christalla, T.; Tessmer, U.; Grundhoff, A. A global analysis of evolutionary conservation among known and predicted gammaherpesvirus microRNAs. J. Virol. 2010, 84, 716–728. [Google Scholar]
- Sethupathy, P.; Megraw, M.; Hatzigeorgiou, A.G. A guide through present computational approaches for the identification of mammalian microRNA targets. Nat. Methods 2006, 3, 881–886. [Google Scholar] [CrossRef]
- Samols, M.A.; Skalsky, R.L.; Maldonado, A.M.; Riva, A.; Lopez, M.C.; Baker, H.V.; Renne, R. Identification of cellular genes targeted by KSHV-encoded microRNAs. PLoS. Pathog. 2007, 3, e65. [Google Scholar] [CrossRef]
- Gottwein, E.; Mukherjee, N.; Sachse, C.; Frenzel, C.; Majoros, W.H.; Chi, J.T.; Braich, R.; Manoharan, M.; Soutschek, J.; Ohler, U.; et al. A viral microRNA functions as an orthologue of cellular miR-155. Nature 2007, 450, 1096–1099. [Google Scholar]
- Ziegelbauer, J. M.; Sullivan, C.S.; Ganem, D. Tandem array-based expression screens identify host mRNA targets of virus-encoded microRNAs. Nat. Genet 2009, 41, 130–134. [Google Scholar]
- Dolken, L.; Malterer, G.; Erhard, F.; Kothe, S.; Friedel, C.C.; Suffert, G.; Marcinowski, L.; Motsch, N.; Barth, S.; Beitzinger, M.; et al. Systematic analysis of viral and cellular microRNA targets in cells latently infected with human gamma-herpesviruses by RISC immunoprecipitation assay. Cell Host Microbe 2010, 7, 324–334. [Google Scholar] [CrossRef]
- Gottwein, E.; Corcoran, D.L.; Mukherjee, N.; Skalsky, R.L.; Hafner, M.; Nusbaum, J.D.; Shamulailatpam, P.; Love, C.L.; Dave, S.S.; Tuschl, T.; et al. Viral MicroRNA Targetome of KSHV-Infected Primary Effusion Lymphoma Cell Lines. Cell Host Microbe 2011, 10, 515–526. [Google Scholar] [CrossRef]
- Skalsky, R.L.; Corcoran, D.L.; Gottwein, E.; Frank, C.L.; Kang, D.; Hafner, M.; Nusbaum, J.D.; Feederle, R.; Delecluse, H.J.; Luftig, M.A.; et al. The viral and cellular microRNA targetome in lymphoblastoid cell lines. PLoS.Pathog. 2012, 8, e1002484. [Google Scholar]
- Riley, K.J.; Rabinowitz, G.S.; Yario, T. A.; Luna, JM.; Darnell, R.B.; Steitz, J.A. EBV and human microRNAs co-target oncogenic and apoptotic viral and human genes during latency. EMBO. J. 2012, 31, 2207–2221. [Google Scholar] [CrossRef]
- Zhang, X.; Dawson, C.W.; He, Z.; Huang, P. Immune evasion strategies of the human gamma-herpesviruses: implications for viral tumorigenesis. J. Med. Virol. 2012, 84, 272–281. [Google Scholar] [CrossRef]
- Xia, T.; O'Hara, A.; Araujo, I.; Barreto, J.; Carvalho, E.; Sapucaia, J.B.; Ramos, J. C.; Luz, E.; Pedroso, C.; Manrique, M.; et al. EBV microRNAs in primary lymphomas and targeting of CXCL-11 by ebv-mir-BHRF1-3. Cancer Res. 2008, 68, 1436–1442. [Google Scholar]
- Nachmani, D.; Stern-Ginossar, N.; Sarid, R.; Mandelboim, O. Diverse herpesvirus microRNAs target the stress-induced immune ligand MICB to escape recognition by natural killer cells. Cell Host Microbe 2009, 5, 376–385. [Google Scholar] [CrossRef]
- Germain, C.; Meier, A.; Jensen, T.; Knapnougel, P.; Poupon, G.; Lazzari, A.; Neisig, A.; Hakansson, K.; Dong, T.; Wagtmann, N.; et al. Induction of lectin-like transcript 1 (LLT1) protein cell surface expression by pathogens and interferon-gamma contributes to modulate immune responses. J. Biol. Chem. 2011, 286, 37964–37975. [Google Scholar]
- Gurer, C.; Strowig, T.; Brilot, F.; Pack, M.; Trumpfheller, C.; Arrey, F.; Park, C.G.; Steinman, R.M.; Munz, C. Targeting the nuclear antigen 1 of Epstein-Barr virus to the human endocytic receptor DEC-205 stimulates protective T-cell responses. Blood 2008, 112, 1231–1239. [Google Scholar] [CrossRef]
- Nakata, A.; Ogawa, K.; Sasaki, T.; Koyama, N.; Wada, K.; Kotera, J.; Kikkawa, H.; Omori, K.; Kaminuma, O. Potential role of phosphodiesterase 7 in human T cell function: comparative effects of two phosphodiesterase inhibitors. Clin. Exp. Immunol. 2002, 128, 460–466. [Google Scholar] [CrossRef]
- Goto, M.; Murakawa, M.; Kadoshima-Yamaoka, K.; Tanaka, Y.; Inoue, H.; Murafuji, H.; Hayashi, Y.; Miura, K.; Nakatsuka, T.; et al. Phosphodiesterase 7A inhibitor ASB16165 suppresses proliferation and cytokine production of NKT cells. Cell Immunol. 2009, 258, 147–151. [Google Scholar] [CrossRef]
- Yang, G.; McIntyre, K.W.; Townsend, R.M.; Shen, H.H.; Pitts, W.J.; Dodd, J.H.; Nadler, S.G.; McKinnon, M.; Watson, A.J. Phosphodiesterase 7A-deficient mice have functional T cells. J. Immunol. 2003, 171, 6414–6420. [Google Scholar]
- Jin, W.; Chang, M.; Sun, S.C. Peli: a family of signal-responsive E3 ubiquitin ligases mediating TLR signaling and T-cell tolerance. Cell Mol. Immunol. 2012, 9, 113–122. [Google Scholar] [CrossRef]
- Tavalai, N.; Stamminger, T. Interplay between Herpesvirus Infection and Host Defense by PML Nuclear Bodies. Viruses 2009, 1, 1240–1264. [Google Scholar] [CrossRef]
- Liang, D.; Gao, Y.; Lin, X.; He, Z.; Zhao, Q.; Deng, Q.; Lan, K. A human herpesvirus miRNA attenuates interferon signaling and contributes to maintenance of viral latency by targeting IKKepsilon. Cell Res. 2011, 21, 793–806. [Google Scholar] [CrossRef]
- Durand, B.; Sperisen, P.; Emery, P.; Barras, E.; Zufferey, M.; Mach, B.; Reith, W. RFXAP, a novel subunit of the RFX DNA binding complex is mutated in MHC class II deficiency. EMBO. J. 1997, 16, 1045–1055. [Google Scholar] [CrossRef]
- Keryer-Bibens, C.; Pioche-Durieu, C.; Villemant, C.; Souquere, S.; Nishi, N.; Hirashima, M.; Middeldorp, J.; Busson, P. Exosomes released by EBV-infected nasopharyngeal carcinoma cells convey the viral latent membrane protein 1 and the immunomodulatory protein galectin 9. BMC. Cancer 2006, 6, 283. [Google Scholar] [CrossRef]
- Pegtel, D. M.; Cosmopoulos, K.; Thorley-Lawson, D.A.; van Eijndhoven, M.A.; Hopmans, E.S.; Lindenberg, J.L.; de Gruijl, T.D.; Wurdinger, T.; Middeldorp, J.M. Functional delivery of viral miRNAs via exosomes. Proc. Natl. Acad. Sci. USA 2010, 107, 6328–6333. [Google Scholar]
- Abend, J.R.; Uldrick, T.; Ziegelbauer, J.M. Regulation of tumor necrosis factor-like weak inducer of apoptosis receptor protein (TWEAKR) expression by Kaposi's sarcoma-associated herpesvirus microRNA prevents TWEAK-induced apoptosis and inflammatory cytokine expression. J. Virol. 2010, 84, 12139–12151. [Google Scholar] [CrossRef]
- Suffert, G.; Malterer, G.; Hausser, J.; Viiliainen, J.; Fender, A.; Contrant, M.; Ivacevic, T.; Benes, V.; Gros, F.; Voinnet, O.; et al. Kaposi's sarcoma herpesvirus microRNAs target caspase 3 and regulate apoptosis. PLoS. Pathog. 2011, 7, e1002405. [Google Scholar] [CrossRef]
- Li, Z.; Chen, X.; Li, L.; Liu, S.; Yang, L.; Ma, X.; Tang, M.; Bode, A.M.; Dong, Z.; Sun, L.; et al. EBV encoded miR-BHRF1-1 potentiates viral lytic replication by downregulating host p53 in nasopharyngeal carcinoma. Int. J. Biochem. Cell Biol. 2012, 44, 275–279. [Google Scholar] [CrossRef]
- Skalsky, R.L.; Samols, M.A.; Plaisance, K.B.; Boss, I.W.; Riva, A.; Lopez, M.C.; Baker, H.V.; Renne, R. Kaposi's sarcoma-associated herpesvirus encodes an ortholog of miR-155. J. Virol. 2007, 81, 12836–12845. [Google Scholar] [CrossRef]
- Gottwein, E.; Cullen, B.R. A human herpesvirus microRNA inhibits p21 expression and attenuates p21-mediated cell cycle arrest. J. Virol. 2010, 84, 5229–5237. [Google Scholar] [CrossRef]
- Choy, E. Y.; Siu, K.L.; Kok, K.H.; Lung, R.W.; Tsang, C.M.; To, K.F.; Kwong, D.L.; Tsao, S.W.; Jin, D.Y. An Epstein-Barr virus-encoded microRNA targets PUMA to promote host cell survival. J. Exp. Med. 2008, 205, 2551–2560. [Google Scholar] [CrossRef]
- Youle, R.J.; Strasser, A. The BCL-2 protein family: opposing activities that mediate cell death. Nat. Rev. Mol. Cell Biol. 2008, 9, 47–59. [Google Scholar] [CrossRef]
- Marquitz, A.R.; Mathur, A.; Nam, C.S.; Raab-Traub, N. The Epstein-Barr Virus BART microRNAs target the pro-apoptotic protein Bim. Virology 2011, 412, 392–400. [Google Scholar] [CrossRef]
- Liu, Y.; Sun, R.; Lin, X.; Liang, D.; Deng, Q.; Lan, K. Kaposi's sarcoma-associated herpesvirus-encoded microRNA miR-K12-11 attenuates transforming growth factor beta signaling through suppression of SMAD5. J. Virol. 2012, 86, 1372–1381. [Google Scholar] [CrossRef]
- Di Bartolo, D.L.; Cannon, M.; Liu, Y.F.; Renne, R.; Chadburn, A.; Boshoff, C.; Cesarman, E. KSHV LANA inhibits TGF-beta signaling through epigenetic silencing of the TGF-beta type II receptor. Blood 2008, 111, 4731–4740. [Google Scholar] [CrossRef]
- Iizasa, H.; Wulff, B.E.; Alla, N.R.; Maragkakis, M.; Megraw, M.; Hatzigeorgiou, A.; Iwakiri, D.; Takada, K.; Wiedmer, A.; Showe, L.; et al. Editing of Epstein-Barr virus-encoded BART6 microRNAs controls their dicer targeting and consequently affects viral latency. J. Biol. Chem. 2010, 285, 33358–33370. [Google Scholar]
- Lu, C.C.; Li, Z.; Chu, C.Y.; Feng, J.; Sun, R.; Rana, T.M. MicroRNAs encoded by Kaposi's sarcoma-associated herpesvirus regulate viral life cycle. EMBO. Rep. 2010, 11, 784–790. [Google Scholar] [CrossRef]
- Bellare, P.; Ganem, D. Regulation of KSHV lytic switch protein expression by a virus-encoded microRNA: an evolutionary adaptation that fine-tunes lytic reactivation. Cell Host Microbe 2009, 6, 570–575. [Google Scholar] [CrossRef]
- Lin, X.; Liang, D.; He, Z.; Deng, Q.; Robertson, E.S.; Lan, K. miR-K12-7-5p encoded by Kaposi's sarcoma-associated herpesvirus stabilizes the latent state by targeting viral ORF50/RTA. PLoS. One. 2011, 6, e16224. [Google Scholar]
- Barth, S.; Pfuhl, T.; Mamiani, A.; Ehses, C.; Roemer, K.; Kremmer, E.; Jaker, C.; Hock, J.; Meister, G.; Grasser, F.A. Epstein-Barr virus-encoded microRNA miR-BART2 down-regulates the viral DNA polymerase BALF5. Nucleic. Acids. Res. 2008, 36, 666–675. [Google Scholar]
- Lo, M.; Wang, Y.Z.; Gout, P.W. The x(c)- cystine/glutamate antiporter: a potential target for therapy of cancer and other diseases. J.. Cell Physiol. 2008, 215, 593–602. [Google Scholar] [CrossRef]
- Kaleeba, J.A.; Berger, E.A. Kaposi's sarcoma-associated herpesvirus fusion-entry receptor: cystine transporter xCT. Science 2006, 311, 1921–1924. [Google Scholar] [CrossRef]
- Qin, Z.; Freitas, E.; Sullivan, R.; Mohan, S.; Bacelieri, R.; Branch, D.; Romano, M.; Kearney, P.; Oates, J.; Plaisance, K.; et al. Upregulation of xCT by KSHV-encoded microRNAs facilitates KSHV dissemination and persistence in an environment of oxidative stress. PLoS. Pathog. 2010, 6, e1000742. [Google Scholar] [CrossRef]
- Bellot, G.; Cartron, P.F.; Er, E.; Oliver, L.; Juin, P.; Armstrong, L.C.; Bornstein, P.; Mihara, K.; Manon, S.; Vallette, F.M. TOM22, a core component of the mitochondria outer membrane protein translocation pore, is a mitochondrial receptor for the proapoptotic protein Bax. Cell Death Differ 2007, 14, 785–794. [Google Scholar] [CrossRef]
- Yang, I.V.; Wade, C.M.; Kang, H.M.; Alper, S.; Rutledge, H.; Lackford, B.; Eskin, E.; Daly, M. J.; Schwartz, D.A. Identification of novel genes that mediate innate immunity using inbred mice. Genetics 2009, 183, 1535–1544. [Google Scholar] [CrossRef]
- Jin, Y.; Sultana, A.; Gandhi, P.; Franklin, E.; Hamamoto, S.; Khan, A.R.; Munson, M.; Schekman, R.; Weisman, L.S. Myosin V transports secretory vesicles via a Rab GTPase cascade and interaction with the exocyst complex. Dev. Cell 2011, 21, 1156–1170. [Google Scholar] [CrossRef]
- Takahashi, S.; Kubo, K.; Waguri, S.; Yabashi, A.; Shin, H.W.; Katoh, Y.; Nakayama, K. Rab11 regulates exocytosis of recycling vesicles at the plasma membrane. J. Cell Sci. 2012. [Google Scholar] [CrossRef]
- Warnatz, H.J.; Schmidt, D.; Manke, T.; Piccini, I.; Sultan, M.; Borodina, T.; Balzereit, D.; Wruck, W.; Soldatov, A.; Vingron, M.; et al. The BTB and CNC homology 1 (BACH1) target genes are involved in the oxidative stress response and in control of the cell cycle. J. Biol. Chem. 2011, 286, 23521–23532. [Google Scholar]
- Boss, I.W.; Nadeau, P.E.; Abbott, J.R.; Yang, Y.; Mergia, A.; Renne, R. A Kaposi's sarcoma-associated herpesvirus-encoded ortholog of microRNA miR-155 induces human splenic B-cell expansion in NOD/LtSz-scid IL2Rgammanull mice. J. Virol. 2011, 85, 9877–9886. [Google Scholar] [CrossRef]
- Rai, D.; Kim, S.W.; McKeller, M.R.; Dahia, P.L.; Aguiar, R.C. Targeting of SMAD5 links microRNA-155 to the TGF-beta pathway and lymphomagenesis. Proc. Natl. Acad. Sci. USA 2010, 107, 3111–3116. [Google Scholar]
- Eis, P.S.; Tam, W.; Sun, L.; Chadburn, A.; Li, Z.; Gomez, M.F.; Lund, E.; Dahlberg, J.E. Accumulation of miR-155 and BIC RNA in human B cell lymphomas. Proc. Natl. Acad. Sci. USA 2005, 102, 3627–3632. [Google Scholar]
- Kluiver, J.; Poppema, S.; de Jong, D.; Blokzijl, T.; Harms, G.; Jacobs, S.; Kroesen, B.J.; van den Berg, A. BIC and miR-155 are highly expressed in Hodgkin, primary mediastinal and diffuse large B cell lymphomas. J. Pathol. 2005, 207, 243–249. [Google Scholar] [CrossRef]
- Linnstaedt, S.D.; Gottwein, E.; Skalsky, R.L.; Luftig, M.A.; Cullen, B.R. Virally induced cellular microRNA miR-155 plays a key role in B-cell immortalization by Epstein-Barr virus. J. Virol. 2010, 84, 11670–11678. [Google Scholar] [CrossRef]
- Zhao, Y.; Yao, Y.; Xu, H.; Lambeth, L.; Smith, L.P.; Kgosana, L.; Wang, X.; Nair, V. A functional MicroRNA-155 ortholog encoded by the oncogenic Marek's disease virus. J. Virol. 2009, 83, 489–492. [Google Scholar]
- Wang, H.W.; Trotter, M.W.; Lagos, D.; Bourboulia, D.; Henderson, S.; Makinen, T.; Elliman, S.; Flanagan, A.M.; Alitalo, K.; Boshoff, C. Kaposi sarcoma herpesvirus-induced cellular reprogramming contributes to the lymphatic endothelial gene expression in Kaposi sarcoma. Nat. Genet 2004, 36, 687–693. [Google Scholar]
- Hansen, A.; Henderson, S.; Lagos, D.; Nikitenko, L.; Coulter, E.; Roberts, S.; Gratrix, F.; Plaisance, K.; Renne, R.; Bower, M.; et al. KSHV-encoded miRNAs target MAF to induce endothelial cell reprogramming. Genes Dev. 2010, 24, 195–205. [Google Scholar] [CrossRef]
- Skalsky, R.L.; Corcoran, D.L.; Gottwein, E.; Frank, C.L.; Kang, D.; Hafner, M.; Nusbaum, J.D.; Feederle, R.; Delecluse, H.J.; Luftig, M.A.; et al. The viral and cellular microRNA targetome in lymphoblastoid cell lines. PLoS. Pathog. 2012, 8, e1002484. [Google Scholar] [CrossRef]
- Riley, K.J.; Rabinowitz, G. S.; Yario, T.A.; Luna, J.M.; Darnell, R.B.; Steitz, J.A. EBV and human microRNAs co-target oncogenic and apoptotic viral and human genes during latency. EMBO. J. 2012, 31, 2207–2221. [Google Scholar] [CrossRef]
- Dolken, L.; Malterer, G.; Erhard, F.; Kothe, S.; Friedel, C.C.; Suffert, G.; Marcinowski, L.; Motsch, N.; Barth, S.; Beitzinger, M.; et al. Systematic analysis of viral and cellular microRNA targets in cells latently infected with human gamma-herpesviruses by RISC immunoprecipitation assay. Cell Host Microbe 2010, 7, 324–334. [Google Scholar] [CrossRef]
- Jurado, J.O.; Alvarez, I.B.; Pasquinelli, V.; Martinez, G.J.; Quiroga, M.F.; Abbate, E.; Musella, R.M.; Chuluyan, H.E.; Garcia, V.E. Programmed death (PD)-1:PD-ligand 1/PD-ligand 2 pathway inhibits T cell effector functions during human tuberculosis. J. Immunol. 2008, 181, 116–125. [Google Scholar]
- Scibetta, A.G.; Santangelo, S.; Coleman, J.; Hall, D.; Chaplin, T.; Copier, J.; Catchpole, S.; Burchell, J.; Taylor-Papadimitriou, J. Functional analysis of the transcription repressor PLU-1/JARID1B. Mol. Cell Biol. 2007, 27, 7220–7235. [Google Scholar]
- Jones, S.A.; White, C.A.; Robb, L.; Alexander, W.S.; Tarlinton, D.M. SOCS3 deletion in B cells alters cytokine responses and germinal center output. J. Immunol. 2011, 187, 6318–6326. [Google Scholar] [CrossRef]
- Grossmann, C.; Podgrabinska, S.; Skobe, M.; Ganem, D. Activation of NF-kappaB by the latent vFLIP gene of Kaposi's sarcoma-associated herpesvirus is required for the spindle shape of virus-infected endothelial cells and contributes to their proinflammatory phenotype. J. Virol. 2006, 80, 7179–7185. [Google Scholar] [CrossRef]
- Mansouri, M.; Rose, P.P.; Moses, A.V.; Fruh, K. Remodeling of endothelial adherens junctions by Kaposi's sarcoma-associated herpesvirus. J. Virol. 2008, 82, 9615–9628. [Google Scholar] [CrossRef]
- Greene, W.; Gao, S.J. Actin dynamics regulate multiple endosomal steps during Kaposi's sarcoma-associated herpesvirus entry and trafficking in endothelial cells. PLoS. Pathog. 2009, 5, e1000512. [Google Scholar] [CrossRef]
- Si, H.; Robertson, E.S. Kaposi's sarcoma-associated herpesvirus-encoded latency-associated nuclear antigen induces chromosomal instability through inhibition of p53 function. J. Virol. 2006, 80, 697–709. [Google Scholar] [CrossRef]
- Shou, Z.; Lin, L.; Liang, J.; Li, J.L.; Chen, H.Y. Expression and prognosis of FOXO3a and HIF-1alpha in nasopharyngeal carcinoma. J. Cancer Res. Clin. Oncol. 2012, 138, 585–593. [Google Scholar] [CrossRef]
- Holland, J.J.; Peterson, J.A. Nucleic Acid and Protein Synthesis during Poliovirus Infection of Human Cells. J. Mol. Biol. 1964, 8, 556–575. [Google Scholar] [CrossRef]
- Malterer, G.; Dolken, L.; Haas, J. The miRNA-targetome of KSHV and EBV in human B-cells. RNA. Biol. 2011, 8, 30–34. [Google Scholar] [CrossRef]
- Abend, J.R.; Ramalingam, D.; Kieffer-Kwon, P.; Uldrick, T.S.; Yarchoan, R.; Ziegelbauer, J.M. KSHV microRNAs target two components of the TLR/IL-1R signaling cascade, IRAK1 and MYD88, to reduce inflammatory cytokine expression. J. Virol. 2012. [Google Scholar] [CrossRef]
- Forte, E.; Salinas, R.E.; Chang, C.; Zhou, T.; Linnstaedt, S.D.; Gottwein, E.; Jacobs, C.; Jima, D.; Li, Q.J.; Dave, S.S.; et al. The Epstein-Barr virus (EBV)-induced tumor suppressor microRNA MiR-34a is growth promoting in EBV-infected B cells. J. Virol. 2012, 86, 6889–6898. [Google Scholar] [CrossRef]
- Forte, E.; Luftig, M.A. The role of microRNAs in Epstein-Barr virus latency and lytic reactivation. Microbes Infect 2011, 13, 1156–1167. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).