Abstract
Melatonin, an endogenously synthesized indolamine, is a powerful antioxidant exerting beneficial action in many pathological conditions. Melatonin protects from oxidative stress in ischemic/reperfusion injury, neurodegenerative diseases, and aging, decreases inflammation, modulates the immune system, inhibits proliferation, counteracts the Warburg effect, and promotes apoptosis in various cancer models. Melatonin stimulates antioxidant enzymes in the cells, protects mitochondrial membrane phospholipids, especially cardiolipin, from oxidation thus preserving integrity of the membranes, affects mitochondrial membrane potential, stimulates activity of respiratory chain enzymes, and decreases the opening of mitochondrial permeability transition pore and cytochrome c release. This review will focus on the molecular mechanisms of melatonin effects in the cells during normal and pathological conditions and possible melatonin clinical applications.
1. Introduction
The natural hormone melatonin (N-acetyl-5-methoxytryptamine) can be found in almost all species, such as fungi, plants, and animals [1]. The diet sources rich in melatonin include olives, rice, chamomile, green tea, coffee, tomatoes, and cereals [2,3]. Melatonin modulates many physiological functions, including sleep and circadian rhythm regulation, neuro- and cardioprotection, acting as powerful antioxidant and protecting from lipid peroxidation, inflammation, decreasing tumor growth, inducing apoptosis, and enhancing mitochondrial activity [1,4].
2. Melatonin Biosynthesis and Bioavailability
Melatonin is secreted primarily by the pineal gland, where it is synthesized from the amino acid tryptophan. Melatonin synthesis could also occur in other organs and cells, such as the bone marrow, brain, lens, skin, retina, and lymphocytes. Four essential enzymes are involved in melatonin synthesis: tryptophan-5-hydroxylase, 5-hydroxytryptophan decarboxylase, serotonin-N-acetyltransferase, and hydroxyindole-O-methyltransferase [5] (Figure 1). Around 30 µg/day of melatonin is secreted constantly in adult humans; however, in the evening, its synthesis increases reaching a maximum plasma peak in the middle of the dark period. As a neurohormone, melatonin is mainly involved in circadian rhythm control [1,6].
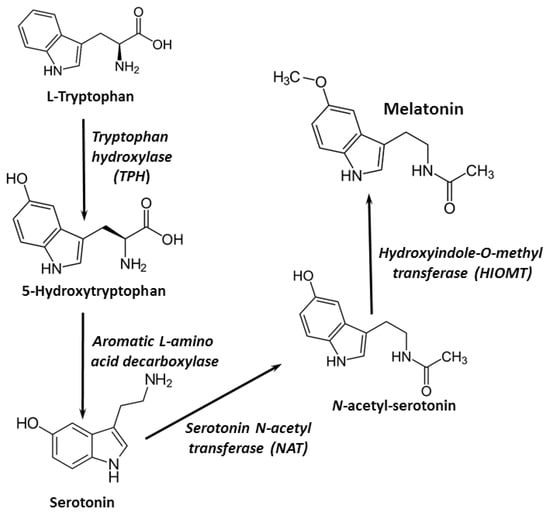
Figure 1.
Melatonin synthesis pathways.
Melatonin is not retained in the pineal gland; it is secreted to the blood and transported to the liver, where it is rapidly metabolized [7]. First, cytochrome P450 monooxygenases A2 and 1A hydroxylate melatonin at the C6 position, then melatonin is converted to its sulfate derivative (6-sulfatoxymelatonin), and later excreted in the urine [7].
Melatonin is transported bound to the serum albumin in the blood, although it could be bound by hemoglobin as well [8,9]. It is capable of crossing the blood–brain barrier [10,11] and cellular membranes [12], and is found in high amounts in mitochondria within the cells [13,14].
The distribution half-life of melatonin is 0.5–5.6 min after intravenous administration [15,16]. After the administration of usual oral dosages (1–5 mg), the plasma concentration peak is reached after 60 min, and then rapidly decreases in a biphasic manner, with the corresponding half-life values of 2 and 20 min [15,16]. Basal melatonin concentrations are reached in 4–8 h. In healthy volunteers, an administration of 0.5 mg of melatonin resulted in a plasma melatonin peak varying between 2 and 395 nmol/L and an elimination half-life of 47 min, whereas bioavailability was around 33%, ranging from 10 to 56% [15,16]. The low melatonin bioavailability is due to excessive hepatic first pass metabolism [15,16,17,18].
The synthesis of melatonin decreases with age, often causing insomnia; the largest decline could be observed in the cases of Alzheimer’s disease, cardiovascular issues, and cancer [19]. Usually, melatonin is safe and not toxic; even at high doses only mild side effects, such as dizziness, headaches, nausea, and sleepiness, could be observed [20].
3. Receptor-Dependent Effects of Melatonin
Melatonin exerts its effects within the cells via interactions with receptors and with intracellular targets, such as transporters, ion binding proteins, enzymes, cytoskeletal components and mitochondria [9,14,21,22], due to its ability to cross freely membranes as well as penetrate the blood–brain barrier [12] (Figure 2).
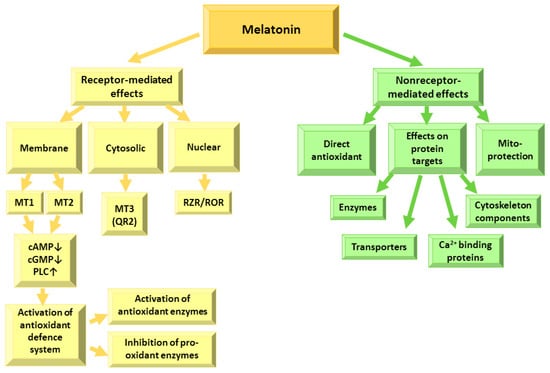
Figure 2.
Receptor-dependent and receptor-independent effects of melatonin. MT—melatonin receptor; cAMP—adenylyl cyclase; cGMP—guanylyl cyclase; PLC—phospholipase C; QR2—quinone reductase 2; RZR/ROR—nuclear retinoid Z receptor.
Melatonin receptors are spread in the retina, brain, kidneys, gastrointestinal tract, skin, immune, endocrine, reproductive, and cardiovascular systems [5]. They modulate adenylyl cyclase (cAMP), guanylyl cyclase (cGMP) and phospholipase C (PLC) activities—and subsequently, calcium and potassium fluxes in the cell [23,24]. Melatonin receptors 1 (MT1) and 2 (MT2) are composed of 350 amino acids and 362 amino acids, respectively, and are 60% homologous proteins belonging to the class of G-protein-coupled receptors (GPCR) [23] with four intracellular and four extracellular domains and 7-transmembrane-helices [25]. The MT1 receptor has an approximately three-fold higher affinity to melatonin than the MT2 receptor, and is thought to be responsible for the circadian effects of melatonin and modulation of the signal transduction in the reproductive system and regulation of peripheral vasoconstriction [26,27]. MT1 receptors are expressed in the retina, brain, cardiovascular system, immune system, skin, pancreas, kidney, liver, spleen, adrenal cortex, testes, ovary, placenta, and breast [26,27]. Most MT2 receptors are present in the retina, brain, immune system, adipose tissue, blood vessels, mammary glands, testes, kidney, and gastrointestinal tract [28]. Melatonin binding to MT1 and MT2 receptors results in the inhibition of the AC/cAMP/PKA/CREB signaling pathway [29], leading to the activation of calcium signaling by calmodulin kinases and PKC and regulation of hormone synthesis. Melatonin binding to the receptors could also activate the Rafs/MEK1/2/ERK1/2 signaling pathway [30,31], which is important for cell proliferation regulation [28]. The melatonin-modulated ERK–MAPK/JNK signaling pathway plays a major role in the oxidative stress [32]. Melatonin can activate the PI3K/Akt (PKB) signaling pathway [33,34] which is very important in cardioprotection [33]. Furthermore, melatonin can decrease tumor cell proliferation and viability by inhibiting the negative feedback from the downstream effector of the PI3K/Akt (PKB) signaling pathway—mammalian target of rapamycin complex 1 (mTOR) [35,36] and modulate apoptosis [37,38]—and inhibit the GC/cGMP/PKG pathway [39]. Melatonin receptor 3 (MT3) is found in animals and is a quinone reductase 2 (QR2) capable of neutralizing free radicals [40,41,42]. The melatonin effects on the nuclear retinoid Z receptor (ROR/RZR) protein family [9,43] play a role in melatonin immunomodulatory actions—T and B lymphocyte stimulation, inhibition of cytokine production, and suppression of nF-κB-dependent inflammation [43]. Thus, melatonin regulates many physiological functions via receptor-dependent signaling pathways.
The receptor-related melatonin effects comprise the indirect modulation of the activity of the enzymes involved in cellular protection against damage by reactive oxygen species (ROS) and reactive nitrogen species (RNS) [7,44,45]. Reduced glutathione (GSH) is one of the most important components of the first line of the antioxidant defense system in the cells capable of neutralizing ROS formed in a variety of redox reactions. The activities of enzymes that improve intracellular levels of reduced GSH are maintained by melatonin [46]. Melatonin increases the activity of γ-glutamylcysteine synthase, which is the rate-limiting enzyme of the GSH synthesis pathway, thus maintaining the cellular levels of GSH [47,48]. Furthermore, melatonin can regulate GSH/GSSG (oxidized glutathione) balance by the stimulation of glutathione reductase (GR), responsible for GSSG reduction to GSH [46]. Melatonin can enhance the gene expression of antioxidant enzymes, comprising superoxide dismutase (SOD) and glutathione peroxidase (GSH-Px) [49,50], and increase catalase (CAT) activity [19]. Melatonin as an indirect antioxidant can also downregulate pro-oxidative and pro-inflammatory enzymes [51]. Melatonin suppresses the nitric oxide synthase, responsible for nitric oxide, and lipoxygenase, for superoxide anion generation [52]. Downregulation of lipoxygenase activity by melatonin protects the cells from the hydroperoxidation of polyunsaturated fatty acids [53]. Melatonin modulates endoplasmic reticulum stress responses [54], activity of sirtuins [55], and processes of mitophagy and autophagy [56,57]. Thus, melatonin is a powerful, multifaceted, natural antioxidant capable of preventing the overproduction of ROS and RNS, to protect the functions of many biological molecules—DNA, lipids, and proteins—from oxidative damage, and to suppress the development of severe degenerative disorders such as cardiologic and neurological diseases, diabetes, and cancer [49,50] (Figure 3).
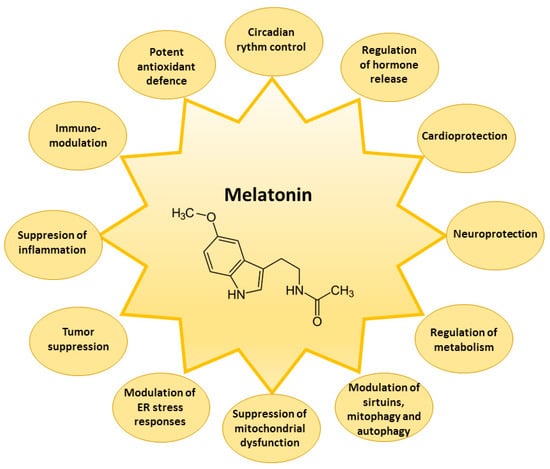
Figure 3.
Main biological effects of melatonin in the cell. ER—endoplasmic reticulum.
4. Receptor-Independent Effects of Melatonin
4.1. Direct Antioxidant Effects of Melatonin
Melatonin is a potent direct scavenger of free radicals, including singlet oxygen, hydroxyl, peroxyl radicals, hydrogen peroxide, nitric oxide and peroxynitrite. Melatonin effectively terminates the radical reaction chain without the formation of pro-oxidant metabolites [3,58] by donating one or more electrons to the free radicals [59]. It is very important that melatonin intermediates produced during free radical neutralization reactions—cyclic 3-hydroxymelatonin, N1-acetyl-N2-formyl-5-methoxykynuramine and N-acetyl-5-methoxykynuramine—are also strong antioxidants [19]; thus, one melatonin molecule has the ability to scavenge up to 10 ROS compared to the traditional antioxidants that typically neutralize one ROS [19,51]. Melatonin can react with hydroxyl radical resulting in the formation of the indolyl radical cation, which has low reactivity and toxicity [60]. Melatonin also directly scavenges the alkoxyl radical, produced during the transition metal-catalyzed degradation of lipid peroxides, thus preventing further lipid peroxidation [61,62]. Melatonin also can act as metal chelator capable of reducing metal-induced toxicity [63].
4.2. Effects of Melatonin on Target Proteins
Melatonin can directly interact with various proteins within the cell, including enzymes, transporters, Ca2+-binding proteins, and cytoskeleton components [9].
The zinc-dependent matrix metalloproteinases (MMPs) which are related to extracellular matrix remodeling and important in pathological processes such as tumor proliferation and the formation of metastasis, atherosclerosis, rheumatoid arthritis, and gastric ulcers, have been suggested to be one of the targets of melatonin [64]. Potent anti-ulcer action of melatonin is attributed to its possible binding to pepsin—a stomach protease responsible for breaking down the ingested proteins into peptides [65,66]. Melatonin-induced inhibition of phosphoprotein phosphatase 2A protects from the hyperphosphorylation of neuronal proteins, protecting neuronal cells and alleviating neurodegenerative diseases [67].
Melatonin is highly lipophilic and can therefore be distributed in the body by the means of passive diffusion [68]. Besides this mechanism, melatonin can be transported across the plasma membrane by the glucose transporter GLUT1 [37,69] and across the mitochondrial membranes by the oligopeptide transporters PEPT1/2 [70]. The melatonin binding site on GLUT1 overlaps with glucose binding [69], which might play a role in the counteraction of the Warburg effect in cancer cells by melatonin [71]. In the plasma, melatonin is bound by serum albumin [8,9].
Melatonin is a high affinity ligand of a Ca2+-binding protein calmodulin, participating in many calcium signaling pathways [28,72]. The cytoskeletal effects of melatonin were suggested to be modulated by the Ca2+–calmodulin complex, although at higher concentrations melatonin has been observed to bind tubulin [73,74,75], leading to the inhibition of microtubule formation. Melatonin via interactions with cytosolic protein networks can control cell motility, division, and organelle function, as well as in mitogen-activated protein kinase (MAPK)-related scaffold protein trafficking and signaling functions [28]. Melatonin can inhibit cytoskeleton reorganization; thus, suppressing cancer proliferation and restoring normal mitochondrial functions [76,77,78]. Melatonin has been shown to bind calreticulin—a Ca2+-binding protein known for its chaperon action and responsible for the regulation of Ca2+ homeostasis [79]. More than 15 melatonin target proteins have been proposed, capable of binding melatonin at concentrations ranging from subnanomolar to millimolar, and indicating the wide variety of processes that could be modulated by melatonin in the cells [9].
4.3. Effects of Melatonin on Epigenetic Regulation
MicroRNAs (miRNAs) have emerged as modulators of gene expression, capable of reaching various parts of organisms via their distribution in exosomes and ectosomes [80], and playing important roles in the development of many pathological conditions [80]. Melatonin has been shown to exert the protective activity by inducing changes in miRNA expression [80,81,82,83].
In a murine model, melatonin reduced the impairment of alcoholic liver disease by enhancing miRNA-497 expression [84], alleviated liver steatosis by modulating miRNA-23a expression [85], and reduced primary sclerosing cholangitis in a murine model of liver fibrosis by inhibiting miRNA-200b expression in cholangiocytes and stellate cells [86].
In Alzheimer’s disease models, melatonin partially suppressed neuroinflammation by reducing an increase in miRNA-124 and enhancing miRNA-132 [87]. Melatonin alleviated neonatal brain inflammation in rats induced by bacterial lipopolysaccharide by restoring upregulated miRNA-34a and downregulated miRNA-146a and miR-126 activities [88].
In cancer models, melatonin suppressed miRNA-155 expression in several human glioma cell lines [89], inhibited angiogenesis modulated by miRNA-3195 and miRNA-374b expression in hypoxic PC-3 prostate cancer cells [90], inhibited the expression of 10 miRNA, and enhanced the expression of 12 miRNA in MCF-7 breast cancer cells [91]. Future directions for studies of the miRNA and melatonin interactions would be the clarification of whether melatonin could modulate the changes in the composition of exosomal and ectosomal miRNAs, thus affecting the progression of pathological conditions in the organism [80].
4.4. Effects of Melatonin on Mitochondrial Functions
Several studies indicate that, in subcellular compartments, melatonin may be differentially distributed. Cell nuclei and mitochondria, in particular, tend to contain higher levels of melatonin than other compartments, such as cytosol [70]. Mitochondria play a crucial role in metabolism, calcium homeostasis, apoptosis, and regulation of many physiological and pathological processes in the cells [92]. Mitochondria are essential organelles, responsible for cellular energy supply through oxidative phosphorylation as well as being the main site of ROS generation. ROS-induced impairment of mitochondrial and cellular functions can be controlled by either preventing the ROS formation or by scavenging them as soon as they are produced [93]. Mitochondria are the main sites for melatonin synthesis and metabolism [70,94], thus melatonin could be an ideal mitochondrial protector due to its close location to ROS production sites and its potent antioxidant properties (Figure 4).
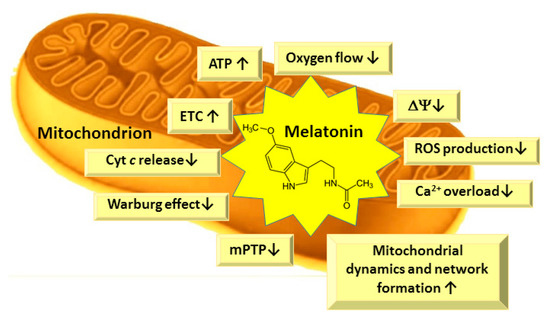
Figure 4.
Main biological effects of melatonin in mitochondria. ETC—electron transport chain; ΔΨ—mitochondrial membrane potential; ROS—reactive oxygen species; mPTP—mitochondrial permeability transition pore; Cyt—cytochrome.
Melatonin has been shown to improve mitochondrial functions, to stimulate the activity of the respiratory chain—mainly complexes I, III and IV [4,95,96,97,98,99]—and to increase mitochondrial ATP production in both normal and pathological conditions, although it did not change the activity of the ATP synthase [4,95,96,97,98,99]. The maintained ATP production at decreased oxygen flux and lower mitochondrial membrane potential is similar to the mild uncoupling, which is important for cardioprotection, neuroprotection, and the prevention of various pathological conditions due to decreased ROS generation [100,101]. Furthermore, the increased ATP production protects from the collapse of mitochondrial membrane potential and calcium overload and inhibits opening of the mitochondrial permeability transition pore (mPTP)—and subsequently, the cell death [102,103]. Peroxidation of mitochondrial membrane phospholipids leads to mitochondrial dysfunction in aging and is the main cause of age-associated disorders [97,104,105,106]. The structure of membrane phospholipids is altered during peroxidation, resulting in the structural changes of the lipid bilayer, and altered membrane fluidity and permeability. Changed membrane properties influence the impairments in the mitochondrial electron transport and oxidative phosphorylation, inner membrane impermeability, decreased mitochondrial membrane potential and effects on mitochondrial Ca2+ homeostasis [107,108,109]. There is a high content of unsaturated fatty acids, such as linoleic acid, in cardiolipin molecules in the heart and liver, and arachidonic and docosahexaenoic acids in the cardiolipin molecules of brain tissue mitochondria, which are highly sensitive to ROS-induced damage [104,110]. Cardiolipin is present in the inner mitochondrial membrane, especially in intermembrane contact sites, near to the sites of ROS production, i.e., the respiratory chain complexes I and III [104,110]. The oxidation or decrease in cardiolipin in mitochondrial inner membrane is supposed to be responsible for age-related decline in mitochondrial functions [102,104]. Oxidized cardiolipin does not tightly bind cytochrome c (Cyt c) and it is released into the intermembrane space, then due to alterations in lipid bilayer, mPTP is formed [111], and Cyt c can be released to cytosol as a signal molecule for apoptosis processes. Melatonin has been shown to prevent cardiolipin peroxidation in mitochondria both under in vitro and in vivo conditions [102,106,112], thus preventing mPTP opening and cytochrome c release from mitochondria [113].
Melatonin also maintains mitochondrial bioenergetics and redox homeostasis via the regulation of mitochondrial dynamics [114]. Studies have revealed the importance of mitochondrial dynamics in inflammatory responses and the maintenance of immune synapse stability. Mitochondrial dynamics have been implicated to modulate the antigen-specific activation and immune responses [115]. Mitochondria have been shown to move towards the immune synapse, driven by cell polarization and integrin adhesion [115,116,117], as well as the fission factor dynamin-related protein 1 (Drp1) regulating mitochondrial redistribution in response to T cell receptor assembly [116].
Melatonin has been shown to exert multiple anticancer effects related to mitochondrial function regulation. A fundamental protective mechanism for reducing pathologies may be the ability of melatonin to alter metabolism in cancer cells as well as other metabolically compromised cells due to the suppression of aerobic glycolysis and activation of oxidative phosphorylation [71,118,119]. Besides inhibiting aerobic glycolysis, melatonin also down-regulates survival signaling and metastasis formation [38]. Melatonin modulates the activity of pyruvate kinase complex and suppresses the pyruvate dehydrogenase kinase, which is considered to be a potential therapeutic target in cancer [120]. The restoration of the mitochondrial network by melatonin increases apoptosis, thus reducing cell growth in lung, breast, and colon cancers [121,122,123].
Thus, melatonin, via its direct ROS/RNS scavenging at its production site—mitochondria—indirect activation of antioxidant defense system, and preservation of the integrity of mitochondrial membranes by protecting them from cardiolipin loss and its peroxidation, has a crucial role in maintaining normal physiological functions of mitochondria and energy turnover in the cells. Melatonin is highly present in mitochondrial membranes; therefore, future studies of the effects of melatonin on the functions of mitochondria in various tissues and organs under normal and pathological conditions as well as investigations of mitochondria-targeted melatonin preparations could help to elucidate the detailed mechanisms of the protective actions of melatonin and its role in alleviating many pathological conditions.
5. Beneficial Effects of Melatonin in Pathological Conditions
5.1. Effects of Melatonin in Neurodegenerative Disoders
Age-related conditions that share mitochondrial dysfunction, oxidative/nitrosative stress, and apoptosis in multiple parts of the brain are neurodegenerative disorders such as Alzheimer’s disease and Parkinson’s disease [124,125]. Reduced melatonin levels are found in the blood and cerebrospinal fluid of Alzheimer’s patients, even in the early onset of the disease [126]. In a rat model of sporadic Alzheimer’s disease, melatonin attenuated memory decline, amyloid-β accumulation, and neurodegeneration [127]. In a Parkinson’s disease model, melatonin restored electron transport chain complex I and complex IV activities, and downregulated mitochondrial inducible nitric oxide synthase by decreasing nitric oxide radicals in mitochondria from substantia nigra and striatum [128]. Decreased local melatonin synthesis in neuronal and immune cells, as well as in the glia and gut, might be important to the etiology and management of Parkinson’s disease [129]. Melatonin has been shown to be neuroprotective in various models of neurodegenerative diseases (for reviews see refs [56,57,130,131,132]). Clinical trials have also been performed to investigate the role of melatonin supplementation on the alleviation of clinical symptoms during Alzheimer’s disease [130]. In studies evaluating sleep efficiency, contradictory results were obtained—melatonin did not show beneficial effects in several studies [133,134,135], probably due to the stability of the assessment method used [130]. A slight neuroprotective effect of melatonin was observed when subjective criteria (Pittsburg Sleep Quality Index) [136] or actigraphy [137] were used to evaluate the melatonin effects. Nevertheless, the majority of clinical investigations support the beneficial effects of melatonin on cognitive impairment and sleep disorders [130].
5.2. Cardioprotective Effects of Melatonin
Melatonin is a cardioprotective agent, reducing the cardiac damage and changes in cellular physiology during ischemia/reperfusion injury due to actions at mitochondrial level [99,112,138,139]. Melatonin treatment of an ischemic/reperfused rat heart significantly decreased the level of lipid peroxidation, restored decreased mitochondrial respiration rate in State 3, as well as complex 1 and complex III activities in isolated rat heart mitochondria [22,112]. Furthermore, melatonin treatment resulted in a decrease in H2O2 generation, protected from cardiolipin loss, and its peroxidation thus preserving the physiological functions of mitochondrial membranes, the ETC activity, and preventing mPTP opening and cytochrome c loss [112,113,139,140]. Melatonin pretreatment attenuates IR-induced mitochondrial oxidative damage via the activation of the JAK2/STAT3 signaling pathway [141]. The STAT3 is a transcription factor of the manganese superoxide dismutase gene, which can increase manganese superoxide dismutase antioxidant activity, thus improving mitochondrial antioxidant defense [142]. Moreover, melatonin can also up-regulate mitochondrial STAT3 through the SAFE signal transduction pathway to reduce myocardial ischemia/reperfusion injury [143]. During ischemia/reperfusion in diabetic rats, melatonin protected mitochondrial functions by alleviating mitochondrial oxidative stress and stimulating mitochondrial biogenesis via up-regulation of the AMPK–PGC-1α–silent information regulator 3 (SIRT3) axis [144].
5.3. Effects of Melatonin in Diabetes, Obesity and Metabolic Diseases
Metabolic diseases—cardiovascular diseases, diabetes, obesity, and metabolic syndromes—are caused by the altered normal physiological processes in the organism [145,146,147]. Melatonin receptors MT1 and MT2 are expressed in islets of Langerhans and they can modulate α-cell-dependent glucagon secretion and β-cell-dependent insulin secretion [148]. Altered signaling of melatonin receptors, especially MT2 activation, could lead to the development of type-2 diabetes mellitus [148]. Melatonin concentration changes are important for the blood glucose-regulating function of the islets [145,148]. The inverse relationship between insulin and melatonin secretion was reported in the studies on diabetic rat models [148]. Furthermore, melatonin was able to inhibit glucose-induced insulin release in rodents [145,149]. Elevated insulin and leptin levels could suppress the increase in melatonin synthesis at night [150]. Additionally, the aging-related increase in insulin resistance could be prevented by melatonin as well [150]. Melatonin modulated body weight regulation and restored the insulin sensitivity in obese young rats on a high-fat diet [151,152]. Melatonin also decreased abdominal fat and plasma leptin levels in middle-aged rats [153,154]. Thus, melatonin could prevent body weight gain without affecting the food intake [145,154,155,156], due to elevated energy expenditure, mainly in brown adipose tissue [156,157] or a stimulation of physical activity or basal metabolic rate [145]. Hypo-lipidemic effects of melatonin were reported in rodents [152,158] and humans [159,160]. In peri- and post-menopausal women, melatonin up-regulated their high-density-lipoprotein cholesterol levels, but not the concentrations of total cholesterol [161]. Melatonin also decreased low-density-lipoprotein cholesterol and triglyceride levels simultaneously, increasing high-density-lipoprotein cholesterol levels in diabetic patients [159,160,162]. Further investigations of melatonin’s influence on mitochondrial functions in obesity or diabetes are expected to detail its mechanisms of alleviation of metabolic diseases [145,146,147].
5.4. Effects of Melatonin in Skeletal Muscle Disorders
The decreased level of melatonin in postmenopausal women was reported in sarcopenia [163,164]. Melatonin supplementation protected mitochondria from aging-caused impairment in cardiac and diaphragm muscles in the model of accelerated aging in mice [165]. Melatonin reduced the inflammation and increased glycolytic potential in skeletal muscles during sarcopenia [166]. Furthermore, melatonin protected from the development of sarcopenia in aged rodent models [167,168]. Melatonin protected from ischemic damage in rat cremaster and gracilis muscles during ischemia/reperfusion [169,170], and enhanced cell regeneration in rat muscle injury models [171,172]. In rodent fibromyalgia models, melatonin protected from ROS damage and restored mitochondrial dynamics parameters in the gastrocnemius muscle [164,173,174]. Melatonin, due to its antioxidant activity, was also effective in alleviating Duchenne muscular dystrophy in rodents and humans [175,176].
5.5. Immunomodulatory Effects of Melatonin
Melatonin exerts a potent immunomodulatory activity in vivo and in vitro [177,178,179]. Melatonin membrane receptors are present in T and B lymphocytes [69]. Melatonin promotes T cell differentiation from the type 1 helper T cells and regulates cytokine gene expression [180,181]. Furthermore, melatonin can enhance cytokine production in human peripheral blood mononuclear cells via its action on nuclear receptors [182].
Melatonin is a pleiotropic immunomodulatory agent, activating inflammation at an early phase via up-regulation of pro-inflammatory phosholipase A2, lipoxygenase, and cytokines (IL-1 and tumor necrosis factor alpha (TNF-α)) [183,184,185]. The anti-inflammatory action of melatonin is related to the suppression of Th1 function and activation of type 2 helper T lymphocytes which produce interleukin (IL)-4 [69].
Melatonin decreases NF-kB binding to DNA, probably by preventing its translocation to the nucleus, thus down-regulating cytokine production [186,187]. Melatonin suppresses chronic inflammation, inhibiting the expression of inducible nitric oxide synthase (iNOS), cyclooxygenase, as well as protein lipase A2, lipoxygenase and cytokine activities due to its antioxidant properties [188]. This activity is similar to that of flavonoids, acting as antioxidants under physiological and as pro-oxidants under pathological conditions.
5.6. Effects of Melatonin in Gastrointestinal Tract
Alterations in gut microbiota and increased gut permeability are implicated in the etiology, course, and treatment of many neurodegenerative diseases [129]. Moreover, the exposure to stress induces changes in the brain–gut interactions, leading to the development of gastrointestinal disorders [189]. Melatonin is present in significant amounts in the gastrointestinal tract, directly affecting its tissues and indirectly modulating the brain–gut axis [129,190,191]. Melatonin protects from the increase in the ROS-induced lipid peroxidation and up-regulates key anti-oxidizing enzymes such as superoxide dismutase [129]. Melatonin restores gastric and pancreatic functions and is implicated in the healing of gastric ulcers, thus beneficially affecting the brain–gut axis [190,191,192,193].
5.7. The Role of Melatonin in the Treatment of the Protozoan Parasitic Infections
Melatonin has been shown to be beneficial in bacterial, viral, and parasitic infections due to its potent antioxidant immunomodulatory effects and direct effects on mitochondrial functions [194]. Melatonin suppressed the development of Plasmodium parasites in malaria due to regulation of cAMP–PKA and IP3–Ca2+ signaling pathways [195,196,197,198]. In Chagas’ disease [199], toxoplasmosis [200] and African trypanosomiasis [201], melatonin stimulated the host’s immune response against the parasite, up-regulating the production of inflammatory mediators. Melatonin inhibited leishmaniasis parasite replication via direct effects on mitochondrial functions—activating the efflux of Ca2+ from mitochondria, increasing the level of mitochondrial nitrites, and suppressing the superoxide dismutase activity [202]. Serum melatonin levels were elevated in giardiasis patients, implicating that the increased melatonin concentrations could modulated phagocytic activity [203]. In amoebiasis, melatonin decreased the amoebic lesions and enhanced the leukophagocytosis [204].
Melatonin, such as many other antiparasitic drugs—ivermectin [205,206,207,208,209], doxycycline [210,211,212], metronidazole [213], mefloquine [214], and hydroxychloroquine [215,216]—has been suggested as a possible adjuvant agent in the treatment of coronavirus disease 2019 (COVID-19) infection [217,218,219,220,221]. Due to its anti-inflammatory, antioxidant and potent mitoprotective properties, melatonin might be beneficial in the preservation of mitochondrial functions and neuro- and cardioprotection during infections, whereas clarification of its mechanism of action as antiparasitic agent would be an interesting topic for further research.
6. Conclusions and Future Perspectives
Melatonin, an endogenous hormone synthetized from tryptophan in the pineal gland mainly during the dark period of the day, regulates the circadian rhythm, can cross cellular membranes and the blood–brain barrier, and is finally driven to mitochondria, where it protects from ROS directly at their generation site—the mitochondrial electron transport chain. Melatonin is most powerful natural antioxidant, alleviating cellular injury during many pathological conditions, such as ischemia/reperfusion, neurodegenerative diseases, aging, and exerting anti-proliferative, anti-inflammatory, and anti-cancer effects.
Melatonin acts via receptor-dependent and independent pathways, modulating various signal transduction steps and supporting the healthy state of mitochondria, preventing the cellular energy supply from impairment. Melatonin is well tolerated as a supplement even at high doses (1–5 mg, [15,16,17,18]), and could serve as a potential adjuvant remedy in many degenerative and infectious diseases and for the sleep regulation. The novel pharmacological melatonin formulations such as prolonged-release tablets or melatonin liposomes, niosomes, or nanoparticles for alternative administration routes—e.g., intranasal or transdermal delivery—could be developed to overcome the problems related to the short therapeutic window of melatonin caused by the poor oral absorption and low bioavailability due to the high first-pass metabolism.
Author Contributions
Conceptualization, D.M.K. and J.B.; literature review and resources, D.M.K. and J.B.; writing—original draft preparation, D.M.K. and J.B.; writing—review and editing, D.M.K. and J.B.; visualization, D.M.K. and J.B.; supervision, J.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors wish to thank the Pharmaceutical and Health Technology Open Access Center for support of this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hardeland, R.; Cardinali, D.P.; Srinivasan, V.; Spence, D.W.; Brown, G.M.; Pandi-Perumal, S.R. Melatonin—A pleiotropic, orchestrating regulator molecule. Prog. Neurobiol. 2011, 93, 350–384. [Google Scholar] [CrossRef] [PubMed]
- Bonomini, F.; Borsani, E.; Favero, G.; Rodella, L.F.; Rezzani, R. Dietary Melatonin Supplementation Could Be a Promising Preventing/Therapeutic Approach for a Variety of Liver Diseases. Nutrients 2018, 10, 1135. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Xie, Y.; Zhang, Z.; Chen, L. Melatonin: A Multifunctional Factor in Plants. Int. J. Mol. Sci. 2018, 19, 1528. [Google Scholar] [CrossRef] [PubMed]
- Acuña Castroviejo, D.; López, L.C.; Escames, G.; López, A.; García, J.A.; Reiter, R.J. Melatonin-mitochondria interplay in health and disease. Curr. Top. Med. Chem. 2011, 11, 221–240. [Google Scholar] [CrossRef] [PubMed]
- Dubocovich, M.L.; Markowska, M. Functional MT1 and MT2 melatonin receptors in mammals. Endocrine 2005, 27, 101–110. [Google Scholar] [CrossRef]
- Carpentieri, A.; Díaz de Barboza, G.; Areco, V.; Peralta López, M.; Tolosa de Talamoni, N. New perspectives in melatonin uses. Pharmacol. Res. 2012, 65, 437–444. [Google Scholar] [CrossRef]
- Acuña-Castroviejo, D.; Escames, G.; Venegas, C.; Díaz-Casado, M.E.; Lima-Cabello, E.; López, L.C.; Rosales-Corral, S.; Tan, D.X.; Reiter, R.J. Extrapineal melatonin: Sources, regulation, and potential functions. Cell. Mol. Life Sci. CMLS 2014, 71, 2997–3025. [Google Scholar] [CrossRef]
- Cardinali, D.P.; Lynch, H.J.; Wurtman, R.J. Binding of melatonin to human and rat plasma proteins. Endocrinology 1972, 91, 1213–1218. [Google Scholar] [CrossRef]
- Liu, L.; Labani, N.; Cecon, E.; Jockers, R. Melatonin Target Proteins: Too Many or Not Enough? Front. Endocrinol. 2019, 10, 791. [Google Scholar] [CrossRef]
- Castagnola, E.; Robbins, E.M.; Woeppel, K.M.; McGuier, M.; Golabchi, A.; Taylor, I.M.; Michael, A.C.; Cui, X.T. Real-Time Fast Scan Cyclic Voltammetry Detection and Quantification of Exogenously Administered Melatonin in Mice Brain. Front. Bioeng. Biotechnol. 2020, 8, 602216. [Google Scholar] [CrossRef]
- Pardridge, W.M.; Mietus, L.J. Transport of albumin-bound melatonin through the blood-brain barrier. J. Neurochem. 1980, 34, 1761–1763. [Google Scholar] [CrossRef] [PubMed]
- Legros, C.; Chesneau, D.; Boutin, J.A.; Barc, C.; Malpaux, B. Melatonin from cerebrospinal fluid but not from blood reaches sheep cerebral tissues under physiological conditions. J. Neuroendocrinol. 2014, 26, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.X.; Mayo, J.C.; Sainz, R.M.; Leon, J.; Czarnocki, Z. Melatonin as an antioxidant: Biochemical mechanisms and pathophysiological implications in humans. Acta Biochim. Pol. 2003, 50, 1129–1146. [Google Scholar] [CrossRef]
- Acuña-Castroviejo, D.; Martín, M.; Macías, M.; Escames, G.; León, J.; Khaldy, H.; Reiter, R.J. Melatonin, mitochondria, and cellular bioenergetics. J. Pineal Res. 2001, 30, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Iguchi, H.; Kato, K.I.; Ibayashi, H. Melatonin serum levels and metabolic clearance rate in patients with liver cirrhosis. J. Clin. Endocrinol. Metab. 1982, 54, 1025–1027. [Google Scholar] [CrossRef]
- Di, W.L.; Kadva, A.; Johnston, A.; Silman, R. Variable bioavailability of oral melatonin. N. Engl. J. Med. 1997, 336, 1028–1029. [Google Scholar] [CrossRef]
- Waldhauser, F.; Saletu, B.; Trinchard-Lugan, I. Sleep laboratory investigations on hypnotic properties of melatonin. Psychopharmacology 1990, 100, 222–226. [Google Scholar] [CrossRef]
- Claustrat, B.; Brun, J.; Chazot, G. The basic physiology and pathophysiology of melatonin. Sleep Med. Rev. 2005, 9, 11–24. [Google Scholar] [CrossRef]
- Reiter, R.J.; Mayo, J.C.; Tan, D.X.; Sainz, R.M.; Alatorre-Jimenez, M.; Qin, L. Melatonin as an antioxidant: Under promises but over delivers. J. Pineal Res. 2016, 61, 253–278. [Google Scholar] [CrossRef]
- Al-Omary, F.A. Melatonin: Comprehensive profile. Profiles Drug Subst. Excip. Relat. Methodol. 2013, 38, 159–226. [Google Scholar] [CrossRef]
- López, A.; García, J.A.; Escames, G.; Venegas, C.; Ortiz, F.; López, L.C.; Acuña-Castroviejo, D. Melatonin protects the mitochondria from oxidative damage reducing oxygen consumption, membrane potential, and superoxide anion production. J. Pineal Res. 2009, 46, 188–198. [Google Scholar] [CrossRef] [PubMed]
- Leon, J.; Acuña-Castroviejo, D.; Sainz, R.M.; Mayo, J.C.; Tan, D.X.; Reiter, R.J. Melatonin and mitochondrial function. Life Sci. 2004, 75, 765–790. [Google Scholar] [CrossRef] [PubMed]
- Stauch, B.; Johansson, L.C.; Cherezov, V. Structural insights into melatonin receptors. FEBS J. 2020, 287, 1496–1510. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Clough, S.J.; Hutchinson, A.J.; Adamah-Biassi, E.B.; Popovska-Gorevski, M.; Dubocovich, M.L. MT1 and MT2 Melatonin Receptors: A Therapeutic Perspective. Annu. Rev. Pharmacol. Toxicol. 2016, 56, 361–383. [Google Scholar] [CrossRef] [PubMed]
- Dubocovich, M.L.; Rivera-Bermudez, M.A.; Gerdin, M.J.; Masana, M.I. Molecular pharmacology, regulation and function of mammalian melatonin receptors. Front. Biosci. A J. Virtual Libr. 2003, 8, d1093–d1108. [Google Scholar] [CrossRef]
- Pala, D.; Lodola, A.; Bedini, A.; Spadoni, G.; Rivara, S. Homology models of melatonin receptors: Challenges and recent advances. Int. J. Mol. Sci. 2013, 14, 8093–8121. [Google Scholar] [CrossRef]
- Jockers, R.; Delagrange, P.; Dubocovich, M.L.; Markus, R.P.; Renault, N.; Tosini, G.; Cecon, E.; Zlotos, D.P. Update on melatonin receptors: IUPHAR Review 20. Br. J. Pharmacol. 2016, 173, 2702–2725. [Google Scholar] [CrossRef]
- Luchetti, F.; Canonico, B.; Betti, M.; Arcangeletti, M.; Pilolli, F.; Piroddi, M.; Canesi, L.; Papa, S.; Galli, F. Melatonin signaling and cell protection function. FASEB J. 2010, 24, 3603–3624. [Google Scholar] [CrossRef]
- Godson, C.; Reppert, S.M. The Mel1a melatonin receptor is coupled to parallel signal transduction pathways. Endocrinology 1997, 138, 397–404. [Google Scholar] [CrossRef]
- Zhu, H.Q.; Cheng, X.W.; Xiao, L.L.; Jiang, Z.K.; Zhou, Q.; Gui, S.Y.; Wei, W.; Wang, Y. Melatonin prevents oxidized low-density lipoprotein-induced increase of myosin light chain kinase activation and expression in HUVEC through ERK/MAPK signal transduction. J. Pineal Res. 2008, 45, 328–334. [Google Scholar] [CrossRef]
- Luchetti, F.; Betti, M.; Canonico, B.; Arcangeletti, M.; Ferri, P.; Galli, F.; Papa, S. ERK MAPK activation mediates the antiapoptotic signaling of melatonin in UVB-stressed U937 cells. Free Radic. Biol. Med. 2009, 46, 339–351. [Google Scholar] [CrossRef] [PubMed]
- Kimball, S.R.; Abbas, A.; Jefferson, L.S. Melatonin represses oxidative stress-induced activation of the MAP kinase and mTOR signaling pathways in H4IIE hepatoma cells through inhibition of Ras. J. Pineal Res. 2008, 44, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Luo, G.P.; Jian, Z.; Ma, R.Y.; Cao, Z.Z.; Zhu, Y.; Zhu, Y.; Tang, F.Q.; Xiao, Y.B. Melatonin alleviates hypoxia-induced cardiac apoptosis through PI3K/Akt pathway. Int. J. Clin. Exp. Pathol. 2018, 11, 5840–5849. [Google Scholar] [PubMed]
- Beker, M.C.; Caglayan, B.; Caglayan, A.B.; Kelestemur, T.; Yalcin, E.; Caglayan, A.; Kilic, U.; Baykal, A.T.; Reiter, R.J.; Kilic, E. Interaction of melatonin and Bmal1 in the regulation of PI3K/AKT pathway components and cellular survival. Sci. Rep. 2019, 9, 19082. [Google Scholar] [CrossRef]
- Kim, H.S.; Kim, T.J.; Yoo, Y.M. Melatonin combined with endoplasmic reticulum stress induces cell death via the PI3K/Akt/mTOR pathway in B16F10 melanoma cells. PLoS ONE 2014, 9, e92627. [Google Scholar] [CrossRef]
- Shen, Y.Q.; Guerra-Librero, A.; Fernandez-Gil, B.I.; Florido, J.; García-López, S.; Martinez-Ruiz, L.; Mendivil-Perez, M.; Soto-Mercado, V.; Acuña-Castroviejo, D.; Ortega-Arellano, H.; et al. Combination of melatonin and rapamycin for head and neck cancer therapy: Suppression of AKT/mTOR pathway activation, and activation of mitophagy and apoptosis via mitochondrial function regulation. J. Pineal Res. 2018, 64. [Google Scholar] [CrossRef]
- Zawilska, J.B.; Skene, D.J.; Arendt, J. Physiology and pharmacology of melatonin in relation to biological rhythms. Pharmacol. Rep. 2009, 61, 383–410. [Google Scholar] [CrossRef]
- Mortezaee, K.; Najafi, M.; Farhood, B.; Ahmadi, A.; Potes, Y.; Shabeeb, D.; Musa, A.E. Modulation of apoptosis by melatonin for improving cancer treatment efficiency: An updated review. Life Sci. 2019, 228, 228–241. [Google Scholar] [CrossRef]
- Spangle, J.M.; Roberts, T.M.; Zhao, J.J. The emerging role of PI3K/AKT-mediated epigenetic regulation in cancer. Biochim. Biophys. Acta Rev. Cancer 2017, 1868, 123–131. [Google Scholar] [CrossRef]
- Černyšiov, V.; Mauricas, M.; Girkontaite, I. Melatonin inhibits granulocyte adhesion to ICAM via MT3/QR2 and MT2 receptors. Int. Immunol. 2015, 27, 599–608. [Google Scholar] [CrossRef]
- Nosjean, O.; Nicolas, J.P.; Klupsch, F.; Delagrange, P.; Canet, E.; Boutin, J.A. Comparative pharmacological studies of melatonin receptors: MT1, MT2 and MT3/QR2. Tissue distribution of MT3/QR2. Biochem. Pharmacol. 2001, 61, 1369–1379. [Google Scholar] [CrossRef]
- Nosjean, O.; Ferro, M.; Coge, F.; Beauverger, P.; Henlin, J.M.; Lefoulon, F.; Fauchere, J.L.; Delagrange, P.; Canet, E.; Boutin, J.A. Identification of the melatonin-binding site MT3 as the quinone reductase 2. J. Biol. Chem. 2000, 275, 31311–31317. [Google Scholar] [CrossRef] [PubMed]
- Smirnov, A.N. Nuclear melatonin receptors. Biochem. Biokhimiia 2001, 66, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Acuña-Castroviejo, D.; Escames, G.; López, L.C.; Hitos, A.B.; León, J. Melatonin and nitric oxide: Two required antagonists for mitochondrial homeostasis. Endocrine 2005, 27, 159–168. [Google Scholar] [CrossRef]
- Gilad, E.; Cuzzocrea, S.; Zingarelli, B.; Salzman, A.L.; Szabó, C. Melatonin is a scavenger of peroxynitrite. Life Sci. 1997, 60, PL169–PL174. [Google Scholar] [CrossRef]
- Pablos, M.I.; Agapito, M.T.; Gutierrez, R.; Recio, J.M.; Reiter, R.J.; Barlow-Walden, L.; Acuña-Castroviejo, D.; Menendez-Pelaez, A. Melatonin stimulates the activity of the detoxifying enzyme glutathione peroxidase in several tissues of chicks. J. Pineal Res. 1995, 19, 111–115. [Google Scholar] [CrossRef]
- Escames, G.; López, L.C.; Tapias, V.; Utrilla, P.; Reiter, R.J.; Hitos, A.B.; León, J.; Rodríguez, M.I.; Acuña-Castroviejo, D. Melatonin counteracts inducible mitochondrial nitric oxide synthase-dependent mitochondrial dysfunction in skeletal muscle of septic mice. J. Pineal Res. 2006, 40, 71–78. [Google Scholar] [CrossRef]
- Urata, Y.; Honma, S.; Goto, S.; Todoroki, S.; Iida, T.; Cho, S.; Honma, K.; Kondo, T. Melatonin induces gamma-glutamylcysteine synthetase mediated by activator protein-1 in human vascular endothelial cells. Free Radic. Biol. Med. 1999, 27, 838–847. [Google Scholar] [CrossRef]
- Reiter, R.J.; Tan, D.X.; Osuna, C.; Gitto, E. Actions of melatonin in the reduction of oxidative stress. A review. J. Biomed. Sci. 2000, 7, 444–458. [Google Scholar] [CrossRef]
- Bonnefont-Rousselot, D.; Collin, F. Melatonin: Action as antioxidant and potential applications in human disease and aging. Toxicology 2010, 278, 55–67. [Google Scholar] [CrossRef]
- Zhang, H.M.; Zhang, Y. Melatonin: A well-documented antioxidant with conditional pro-oxidant actions. J. Pineal Res. 2014, 57, 131–146. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.X.; Manchester, L.C.; Pilar Terron, M.; Flores, L.J.; Koppisepi, S. Medical implications of melatonin: Receptor-mediated and receptor-independent actions. Adv. Med. Sci. 2007, 52, 11–28. [Google Scholar] [PubMed]
- Blokhina, O.; Virolainen, E.; Fagerstedt, K.V. Antioxidants, oxidative damage and oxygen deprivation stress: A review. Ann. Bot. 2003, 91, 179–194. [Google Scholar] [CrossRef] [PubMed]
- Fernández, A.; Ordóñez, R.; Reiter, R.J.; González-Gallego, J.; Mauriz, J.L. Melatonin and endoplasmic reticulum stress: Relation to autophagy and apoptosis. J. Pineal Res. 2015, 59, 292–307. [Google Scholar] [CrossRef] [PubMed]
- Mayo, J.C.; Sainz, R.M.; González Menéndez, P.; Cepas, V.; Tan, D.X.; Reiter, R.J. Melatonin and sirtuins: A “not-so unexpected” relationship. J. Pineal Res. 2017, 62. [Google Scholar] [CrossRef]
- Luo, F.; Sandhu, A.F.; Rungratanawanich, W.; Williams, G.E.; Akbar, M.; Zhou, S.; Song, B.J.; Wang, X. Melatonin and Autophagy in Aging-Related Neurodegenerative Diseases. Int. J. Mol. Sci. 2020, 21, 7174. [Google Scholar] [CrossRef]
- Stacchiotti, A.; Corsetti, G. Natural Compounds and Autophagy: Allies Against Neurodegeneration. Front. Cell Dev. Biol. 2020, 8, 555409. [Google Scholar] [CrossRef]
- Mortezaee, K.; Potes, Y.; Mirtavoos-Mahyari, H.; Motevaseli, E.; Shabeeb, D.; Musa, A.E.; Najafi, M.; Farhood, B. Boosting immune system against cancer by melatonin: A mechanistic viewpoint. Life Sci. 2019, 238, 116960. [Google Scholar] [CrossRef]
- Reiter, R.J.; Guerrero, J.M.; Garcia, J.J.; Acuña-Castroviejo, D. Reactive oxygen intermediates, molecular damage, and aging. Relation to melatonin. Ann. N. Y. Acad. Sci. 1998, 854, 410–424. [Google Scholar] [CrossRef]
- Galano, A.; Tan, D.X.; Reiter, R.J. Melatonin as a natural ally against oxidative stress: A physicochemical examination. J. Pineal Res. 2011, 51, 1–16. [Google Scholar] [CrossRef]
- Reiter, R.J.; Tan, D.X.; Galano, A. Melatonin reduces lipid peroxidation and membrane viscosity. Front. Physiol. 2014, 5, 377. [Google Scholar] [CrossRef] [PubMed]
- García, J.J.; López-Pingarrón, L.; Almeida-Souza, P.; Tres, A.; Escudero, P.; García-Gil, F.A.; Tan, D.X.; Reiter, R.J.; Ramírez, J.M.; Bernal-Pérez, M. Protective effects of melatonin in reducing oxidative stress and in preserving the fluidity of biological membranes: A review. J. Pineal Res. 2014, 56, 225–237. [Google Scholar] [CrossRef] [PubMed]
- Romero, A.; Ramos, E.; de Los Ríos, C.; Egea, J.; Del Pino, J.; Reiter, R.J. A review of metal-catalyzed molecular damage: Protection by melatonin. J. Pineal Res. 2014, 56, 343–370. [Google Scholar] [CrossRef] [PubMed]
- Swarnakar, S.; Paul, S.; Singh, L.P.; Reiter, R.J. Matrix metalloproteinases in health and disease: Regulation by melatonin. J. Pineal Res. 2011, 50, 8–20. [Google Scholar] [CrossRef] [PubMed]
- Bubenik, G.A. Localization, physiological significance and possible clinical implication of gastrointestinal melatonin. Biol. Signals Recept. 2001, 10, 350–366. [Google Scholar] [CrossRef]
- Majka, J.; Wierdak, M.; Brzozowska, I.; Magierowski, M.; Szlachcic, A.; Wojcik, D.; Kwiecien, S.; Magierowska, K.; Zagajewski, J.; Brzozowski, T. Melatonin in Prevention of the Sequence from Reflux Esophagitis to Barrett’s Esophagus and Esophageal Adenocarcinoma: Experimental and Clinical Perspectives. Int. J. Mol. Sci. 2018, 19, 2033. [Google Scholar] [CrossRef]
- Gong, Y.H.; Hua, N.; Zang, X.; Huang, T.; He, L. Melatonin ameliorates Aβ(1-42) -induced Alzheimer’s cognitive deficits in mouse model. J. Pharm. Pharmacol. 2018, 70, 70–80. [Google Scholar] [CrossRef]
- Yu, H.; Dickson, E.J.; Jung, S.R.; Koh, D.S.; Hille, B. High membrane permeability for melatonin. J. Gen. Physiol. 2016, 147, 63–76. [Google Scholar] [CrossRef]
- Ren, W.; Liu, G.; Chen, S.; Yin, J.; Wang, J.; Tan, B.; Wu, G.; Bazer, F.W.; Peng, Y.; Li, T.; et al. Melatonin signaling in T cells: Functions and applications. J. Pineal Res. 2017, 62. [Google Scholar] [CrossRef]
- Mayo, J.C.; Sainz, R.M.; González-Menéndez, P.; Hevia, D.; Cernuda-Cernuda, R. Melatonin transport into mitochondria. Cell. Mol. Life Sci. 2017, 74, 3927–3940. [Google Scholar] [CrossRef]
- Reiter, R.J.; Sharma, R.; Ma, Q.; Rorsales-Corral, S.; de Almeida Chuffa, L.G. Melatonin inhibits Warburg-dependent cancer by redirecting glucose oxidation to the mitochondria: A mechanistic hypothesis. Cell. Mol. Life Sci. 2020, 77, 2527–2542. [Google Scholar] [CrossRef]
- Benítez-King, G.; Antón-Tay, F. Calmodulin mediates melatonin cytoskeletal effects. Experientia 1993, 49, 635–641. [Google Scholar] [CrossRef] [PubMed]
- Huerto-Delgadillo, L.; Antón-Tay, F.; Benítez-King, G. Effects of melatonin on microtubule assembly depend on hormone concentration: Role of melatonin as a calmodulin antagonist. J. Pineal Res. 1994, 17, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Freire, F.; Cardinali, D.P. Effects of melatonin treatment and environmental lighting on the ultrastructural appearence, melatonin synthesis, norepinephrine turnover and microtubule protein content of the rat pineal gland. J. Neural Transm. 1975, 37, 237–257. [Google Scholar] [CrossRef]
- Jarzynka, M.J.; Passey, D.K.; Johnson, D.A.; Konduru, N.V.; Fitz, N.F.; Radio, N.M.; Rasenick, M.; Benloucif, S.; Melan, M.A.; Witt-Enderby, P.A. Microtubules modulate melatonin receptors involved in phase-shifting circadian activity rhythms: In vitro and in vivo evidence. J. Pineal Res. 2009, 46, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Yoon, Y.M.; Go, G.; Yun, C.W.; Lim, J.H.; Lee, J.H.; Lee, S.H. Melatonin Suppresses Renal Cortical Fibrosis by Inhibiting Cytoskeleton Reorganization and Mitochondrial Dysfunction through Regulation of miR-4516. Int. J. Mol. Sci. 2020, 21, 5323. [Google Scholar] [CrossRef]
- Alvarez-Artime, A.; Cernuda-Cernuda, R.; Francisco Artime, N.; Cepas, V.; Gonzalez-Menendez, P.; Fernadez-Vega, S.; Quiros-Gonzalez, I.; Sainz, R.M.; Mayo, J.C. Melatonin-Induced Cytoskeleton Reorganization Leads to Inhibition of Melanoma Cancer Cell Proliferation. Int. J. Mol. Sci. 2020, 21, 548. [Google Scholar] [CrossRef]
- Su, S.C.; Hsieh, M.J.; Yang, W.E.; Chung, W.H.; Reiter, R.J.; Yang, S.F. Cancer metastasis: Mechanisms of inhibition by melatonin. J. Pineal Res. 2017, 62. [Google Scholar] [CrossRef]
- Macías, M.; Escames, G.; Leon, J.; Coto, A.; Sbihi, Y.; Osuna, A.; Acuña-Castroviejo, D. Calreticulin-melatonin. An unexpected relationship. Eur. J. Biochem. 2003, 270, 832–840. [Google Scholar] [CrossRef]
- Hardeland, R. Aging, Melatonin, and the Pro- and Anti-Inflammatory Networks. Int. J. Mol. Sci. 2019, 20, 1223. [Google Scholar] [CrossRef]
- Korkmaz, A.; Rosales-Corral, S.; Reiter, R.J. Gene regulation by melatonin linked to epigenetic phenomena. Gene 2012, 503, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Hardeland, R. Melatonin, noncoding RNAs, messenger RNA stability and epigenetics—Evidence, hints, gaps and perspectives. Int. J. Mol. Sci. 2014, 15, 18221–18252. [Google Scholar] [CrossRef] [PubMed]
- Hardeland, R. Melatonin and the pathologies of weakened or dysregulated circadian oscillators. J. Pineal Res. 2017, 62. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.D.; Hwang, S.L.; Lee, E.J.; Kim, H.M.; Chung, M.J.; Elfadl, A.K.; Lee, S.E.; Nedumaran, B.; Harris, R.A.; Jeong, K.S. Melatonin ameliorates alcohol-induced bile acid synthesis by enhancing miR-497 expression. J. Pineal Res. 2017, 62. [Google Scholar] [CrossRef]
- Kim, S.J.; Kang, H.S.; Lee, J.H.; Park, J.H.; Jung, C.H.; Bae, J.H.; Oh, B.C.; Song, D.K.; Baek, W.K.; Im, S.S. Melatonin ameliorates ER stress-mediated hepatic steatosis through miR-23a in the liver. Biochem. Biophys. Res. Commun. 2015, 458, 462–469. [Google Scholar] [CrossRef]
- Wu, N.; Meng, F.; Zhou, T.; Han, Y.; Kennedy, L.; Venter, J.; Francis, H.; DeMorrow, S.; Onori, P.; Invernizzi, P.; et al. Prolonged darkness reduces liver fibrosis in a mouse model of primary sclerosing cholangitis by miR-200b down-regulation. FASEB J. 2017, 31, 4305–4324. [Google Scholar] [CrossRef]
- Hardeland, R.; Cardinali, D.P.; Brown, G.M.; Pandi-Perumal, S.R. Melatonin and brain inflammaging. Prog. Neurobiol. 2015, 127-128, 46–63. [Google Scholar] [CrossRef]
- Wang, X.; Wang, Z.H.; Wu, Y.Y.; Tang, H.; Tan, L.; Wang, X.; Gao, X.Y.; Xiong, Y.S.; Liu, D.; Wang, J.Z.; et al. Melatonin attenuates scopolamine-induced memory/synaptic disorder by rescuing EPACs/miR-124/Egr1 pathway. Mol. Neurobiol. 2013, 47, 373–381. [Google Scholar] [CrossRef]
- Gu, J.; Lu, Z.; Ji, C.; Chen, Y.; Liu, Y.; Lei, Z.; Wang, L.; Zhang, H.T.; Li, X. Melatonin inhibits proliferation and invasion via repression of miRNA-155 in glioma cells. Biomed. Pharmacother. 2017, 93, 969–975. [Google Scholar] [CrossRef]
- Sohn, E.J.; Won, G.; Lee, J.; Lee, S.; Kim, S.H. Upregulation of miRNA3195 and miRNA374b Mediates the Anti-Angiogenic Properties of Melatonin in Hypoxic PC-3 Prostate Cancer Cells. J. Cancer 2015, 6, 19–28. [Google Scholar] [CrossRef]
- Lee, S.E.; Kim, S.J.; Youn, J.P.; Hwang, S.Y.; Park, C.S.; Park, Y.S. MicroRNA and gene expression analysis of melatonin-exposed human breast cancer cell lines indicating involvement of the anticancer effect. J. Pineal Res. 2011, 51, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Abate, M.; Festa, A.; Falco, M.; Lombardi, A.; Luce, A.; Grimaldi, A.; Zappavigna, S.; Sperlongano, P.; Irace, C.; Caraglia, M.; et al. Mitochondria as playmakers of apoptosis, autophagy and senescence. Semin. Cell Dev. Biol. 2020, 98, 139–153. [Google Scholar] [CrossRef] [PubMed]
- Dröse, S.; Brandt, U.; Wittig, I. Mitochondrial respiratory chain complexes as sources and targets of thiol-based redox-regulation. Biochim. Biophys. Acta 2014, 1844, 1344–1354. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Rosales-Corral, S.; Tan, D.X.; Jou, M.J.; Galano, A.; Xu, B. Melatonin as a mitochondria-targeted antioxidant: One of evolution’s best ideas. Cell. Mol. Life Sci. 2017, 74, 3863–3881. [Google Scholar] [CrossRef] [PubMed]
- Acuña Castroviejo, D.; Escames, G.; Carazo, A.; León, J.; Khaldy, H.; Reiter, R.J. Melatonin, mitochondrial homeostasis and mitochondrial-related diseases. Curr. Top. Med. Chem. 2002, 2, 133–151. [Google Scholar] [CrossRef]
- Acuña-Castroviejo, D.; Escames, G.; León, J.; Carazo, A.; Khaldy, H. Mitochondrial regulation by melatonin and its metabolites. Adv. Exp. Med. Biol. 2003, 527, 549–557. [Google Scholar] [CrossRef]
- Paradies, G.; Paradies, V.; Ruggiero, F.M.; Petrosillo, G. Protective role of melatonin in mitochondrial dysfunction and related disorders. Arch. Toxicol. 2015, 89, 923–939. [Google Scholar] [CrossRef]
- Paradies, G.; Paradies, V.; Ruggiero, F.M.; Petrosillo, G. Mitochondrial bioenergetics decay in aging: Beneficial effect of melatonin. Cell. Mol. Life Sci. 2017, 74, 3897–3911. [Google Scholar] [CrossRef]
- Petrosillo, G.; Di Venosa, N.; Pistolese, M.; Casanova, G.; Tiravanti, E.; Colantuono, G.; Federici, A.; Paradies, G.; Ruggiero, F.M. Protective effect of melatonin against mitochondrial dysfunction associated with cardiac ischemia- reperfusion: Role of cardiolipin. FASEB J. 2006, 20, 269–276. [Google Scholar] [CrossRef]
- Starkov, A.A. “Mild” uncoupling of mitochondria. Biosci. Rep. 1997, 17, 273–279. [Google Scholar] [CrossRef]
- Skulachev, V.P. Uncoupling: New approaches to an old problem of bioenergetics. Biochim. Biophys. Acta 1998, 1363, 100–124. [Google Scholar] [CrossRef]
- Petrosillo, G.; Moro, N.; Ruggiero, F.M.; Paradies, G. Melatonin inhibits cardiolipin peroxidation in mitochondria and prevents the mitochondrial permeability transition and cytochrome c release. Free Radic. Biol. Med. 2009, 47, 969–974. [Google Scholar] [CrossRef] [PubMed]
- Petrosillo, G.; Ruggiero, F.M.; Paradies, G. Role of reactive oxygen species and cardiolipin in the release of cytochrome c from mitochondria. FASEB J. 2003, 17, 2202–2208. [Google Scholar] [CrossRef]
- Paradies, G.; Paradies, V.; Ruggiero, F.M.; Petrosillo, G. Cardiolipin and mitochondrial function in health and disease. Antioxid. Redox Signal. 2014, 20, 1925–1953. [Google Scholar] [CrossRef] [PubMed]
- Paradies, G.; Petrosillo, G.; Paradies, V.; Ruggiero, F.M. Oxidative stress, mitochondrial bioenergetics, and cardiolipin in aging. Free Radic. Biol. Med. 2010, 48, 1286–1295. [Google Scholar] [CrossRef]
- Paradies, G.; Ruggiero, F.M.; Petrosillo, G.; Quagliariello, E. Peroxidative damage to cardiac mitochondria: Cytochrome oxidase and cardiolipin alterations. FEBS Lett. 1998, 424, 155–158. [Google Scholar] [CrossRef]
- Paradies, G.; Paradies, V.; Ruggiero, F.M.; Petrosillo, G. Role of Cardiolipin in Mitochondrial Function and Dynamics in Health and Disease: Molecular and Pharmacological Aspects. Cells 2019, 8, 728. [Google Scholar] [CrossRef]
- Paradies, G.; Petrosillo, G.; Paradies, V.; Reiter, R.J.; Ruggiero, F.M. Melatonin, cardiolipin and mitochondrial bioenergetics in health and disease. J. Pineal Res. 2010, 48, 297–310. [Google Scholar] [CrossRef]
- Petrosillo, G.; Casanova, G.; Matera, M.; Ruggiero, F.M.; Paradies, G. Interaction of peroxidized cardiolipin with rat-heart mitochondrial membranes: Induction of permeability transition and cytochrome c release. FEBS Lett. 2006, 580, 6311–6316. [Google Scholar] [CrossRef]
- Paradies, G.; Paradies, V.; De Benedictis, V.; Ruggiero, F.M.; Petrosillo, G. Functional role of cardiolipin in mitochondrial bioenergetics. Biochim. Biophys. Acta 2014, 1837, 408–417. [Google Scholar] [CrossRef]
- Kagan, V.E.; Chu, C.T.; Tyurina, Y.Y.; Cheikhi, A.; Bayir, H. Cardiolipin asymmetry, oxidation and signaling. Chem. Phys. Lipids 2014, 179, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Petrosillo, G.; Colantuono, G.; Moro, N.; Ruggiero, F.M.; Tiravanti, E.; Di Venosa, N.; Fiore, T.; Paradies, G. Melatonin protects against heart ischemia-reperfusion injury by inhibiting mitochondrial permeability transition pore opening. Am. J. Physiol. Heart Circ. Physiol. 2009, 297, H1487–H1493. [Google Scholar] [CrossRef] [PubMed]
- Paradies, G.; Petrosillo, G.; Paradies, V.; Ruggiero, F.M. Role of cardiolipin peroxidation and Ca2+ in mitochondrial dysfunction and disease. Cell Calcium 2009, 45, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Parameyong, A.; Govitrapong, P.; Chetsawang, B. Melatonin attenuates the mitochondrial translocation of mitochondrial fission proteins and Bax, cytosolic calcium overload and cell death in methamphetamine-induced toxicity in neuroblastoma SH-SY5Y cells. Mitochondrion 2015, 24, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Martín-Cófreces, N.B.; Baixauli, F.; Sánchez-Madrid, F. Immune synapse: Conductor of orchestrated organelle movement. Trends Cell Biol. 2014, 24, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Quintana, A.; Schwindling, C.; Wenning, A.S.; Becherer, U.; Rettig, J.; Schwarz, E.C.; Hoth, M. T cell activation requires mitochondrial translocation to the immunological synapse. Proc. Natl. Acad. Sci. USA 2007, 104, 14418–14423. [Google Scholar] [CrossRef]
- Baixauli, F.; Martín-Cófreces, N.B.; Morlino, G.; Carrasco, Y.R.; Calabia-Linares, C.; Veiga, E.; Serrador, J.M.; Sánchez-Madrid, F. The mitochondrial fission factor dynamin-related protein 1 modulates T-cell receptor signalling at the immune synapse. EMBO J. 2011, 30, 1238–1250. [Google Scholar] [CrossRef]
- Proietti, S.; Cucina, A.; Minini, M.; Bizzarri, M. Melatonin, mitochondria, and the cancer cell. Cell. Mol. Life Sci. 2017, 74, 4015–4025. [Google Scholar] [CrossRef]
- Reiter, R.J.; Sharma, R.; Ma, Q. Switching diseased cells from cytosolic aerobic glycolysis to mitochondrial oxidative phosphorylation: A metabolic rhythm regulated by melatonin? J. Pineal Res. 2020, e12677. [Google Scholar] [CrossRef]
- Ghareghani, M.; Scavo, L.; Jand, Y.; Farhadi, N.; Sadeghi, H.; Ghanbari, A.; Mondello, S.; Arnoult, D.; Gharaghani, S.; Zibara, K. Melatonin Therapy Modulates Cerebral Metabolism and Enhances Remyelination by Increasing PDK4 in a Mouse Model of Multiple Sclerosis. Front. Pharmacol. 2019, 10, 147. [Google Scholar] [CrossRef]
- Rehman, J.; Zhang, H.J.; Toth, P.T.; Zhang, Y.; Marsboom, G.; Hong, Z.; Salgia, R.; Husain, A.N.; Wietholt, C.; Archer, S.L. Inhibition of mitochondrial fission prevents cell cycle progression in lung cancer. FASEB J. 2012, 26, 2175–2186. [Google Scholar] [CrossRef] [PubMed]
- Hagenbuchner, J.; Kuznetsov, A.V.; Obexer, P.; Ausserlechner, M.J. BIRC5/Survivin enhances aerobic glycolysis and drug resistance by altered regulation of the mitochondrial fusion/fission machinery. Oncogene 2013, 32, 4748–4757. [Google Scholar] [CrossRef]
- Inoue-Yamauchi, A.; Oda, H. Depletion of mitochondrial fission factor DRP1 causes increased apoptosis in human colon cancer cells. Biochem. Biophys. Res. Commun. 2012, 421, 81–85. [Google Scholar] [CrossRef] [PubMed]
- Hardeland, R. Recent Findings in Melatonin Research and Their Relevance to the CNS. Cent. Nerv. Syst. Agents Med. Chem. 2018, 18, 102–114. [Google Scholar] [CrossRef] [PubMed]
- Balmik, A.A.; Chinnathambi, S. Multi-Faceted Role of Melatonin in Neuroprotection and Amelioration of Tau Aggregates in Alzheimer’s Disease. J. Alzheimer’s Dis. 2018, 62, 1481–1493. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.Y.; Zhou, J.N.; van Heerikhuize, J.; Hofman, M.A.; Swaab, D.F. Decreased melatonin levels in postmortem cerebrospinal fluid in relation to aging, Alzheimer’s disease, and apolipoprotein E-epsilon4/4 genotype. J. Clin. Endocrinol. Metab. 1999, 84, 323–327. [Google Scholar] [CrossRef]
- Rudnitskaya, E.A.; Muraleva, N.A.; Maksimova, K.Y.; Kiseleva, E.; Kolosova, N.G.; Stefanova, N.A. Melatonin Attenuates Memory Impairment, Amyloid-β Accumulation, and Neurodegeneration in a Rat Model of Sporadic Alzheimer’s Disease. J. Alzheimer’s Dis. 2015, 47, 103–116. [Google Scholar] [CrossRef]
- Tapias, V.; Escames, G.; López, L.C.; López, A.; Camacho, E.; Carrión, M.D.; Entrena, A.; Gallo, M.A.; Espinosa, A.; Acuña-Castroviejo, D. Melatonin and its brain metabolite N(1)-acetyl-5-methoxykynuramine prevent mitochondrial nitric oxide synthase induction in parkinsonian mice. J. Neurosci. Res. 2009, 87, 3002–3010. [Google Scholar] [CrossRef]
- Anderson, G.; Seo, M.; Berk, M.; Carvalho, A.F.; Maes, M. Gut Permeability and Microbiota in Parkinson’s Disease: Role of Depression, Tryptophan Catabolites, Oxidative and Nitrosative Stress and Melatonergic Pathways. Curr. Pharm. Des. 2016, 22, 6142–6151. [Google Scholar] [CrossRef]
- Chen, D.; Zhang, T.; Lee, T.H. Cellular Mechanisms of Melatonin: Insight from Neurodegenerative Diseases. Biomolecules 2020, 10, 1158. [Google Scholar] [CrossRef]
- Lee, J.G.; Woo, Y.S.; Park, S.W.; Seog, D.H.; Seo, M.K.; Bahk, W.M. The Neuroprotective Effects of Melatonin: Possible Role in the Pathophysiology of Neuropsychiatric Disease. Brain Sci. 2019, 9, 285. [Google Scholar] [CrossRef] [PubMed]
- Shukla, M.; Govitrapong, P.; Boontem, P.; Reiter, R.J.; Satayavivad, J. Mechanisms of Melatonin in Alleviating Alzheimer’s Disease. Curr. Neuropharmacol. 2017, 15, 1010–1031. [Google Scholar] [CrossRef] [PubMed]
- Alves, G.S.; Carvalho, A.F.; de Amorim de Carvalho, L.; Sudo, F.K.; Siqueira-Neto, J.I.; Oertel-Knochel, V.; Jurcoane, A.; Knochel, C.; Boecker, H.; Laks, J.; et al. Neuroimaging Findings Related to Behavioral Disturbances in Alzheimer’s Disease: A Systematic Review. Curr. Alzheimer Res. 2017, 14, 61–75. [Google Scholar] [CrossRef]
- Serfaty, M.; Kennell-Webb, S.; Warner, J.; Blizard, R.; Raven, P. Double blind randomised placebo controlled trial of low dose melatonin for sleep disorders in dementia. Int. J. Geriatr. Psychiatry 2002, 17, 1120–1127. [Google Scholar] [CrossRef] [PubMed]
- Singer, C.; Tractenberg, R.E.; Kaye, J.; Schafer, K.; Gamst, A.; Grundman, M.; Thomas, R.; Thal, L.J. A multicenter, placebo-controlled trial of melatonin for sleep disturbance in Alzheimer’s disease. Sleep 2003, 26, 893–901. [Google Scholar] [CrossRef]
- Wade, A.G.; Farmer, M.; Harari, G.; Fund, N.; Laudon, M.; Nir, T.; Frydman-Marom, A.; Zisapel, N. Add-on prolonged-release melatonin for cognitive function and sleep in mild to moderate Alzheimer’s disease: A 6-month, randomized, placebo-controlled, multicenter trial. Clin. Interv. Aging 2014, 9, 947–961. [Google Scholar] [CrossRef]
- Asayama, K.; Yamadera, H.; Ito, T.; Suzuki, H.; Kudo, Y.; Endo, S. Double blind study of melatonin effects on the sleep-wake rhythm, cognitive and non-cognitive functions in Alzheimer type dementia. J. Nippon Med. School 2003, 70, 334–341. [Google Scholar] [CrossRef]
- Aslan, G.; Gül, H.F.; Tektemur, A.; Sahna, E. Ischemic postconditioning reduced myocardial ischemia-reperfusion injury: The roles of melatonin and uncoupling protein 3. Anatol. J. Cardiol. 2020, 23, 19–27. [Google Scholar] [CrossRef]
- Ma, Z.; Xin, Z.; Di, W.; Yan, X.; Li, X.; Reiter, R.J.; Yang, Y. Melatonin and mitochondrial function during ischemia/reperfusion injury. Cell. Mol. Life Sci. 2017, 74, 3989–3998. [Google Scholar] [CrossRef]
- León, J.; Acuña-Castroviejo, D.; Escames, G.; Tan, D.X.; Reiter, R.J. Melatonin mitigates mitochondrial malfunction. J. Pineal Res. 2005, 38, 1–9. [Google Scholar] [CrossRef]
- Yang, B.; Zang, L.E.; Cui, J.W.; Zhang, M.Y.; Ma, X.; Wei, L.L. Melatonin Plays a Protective Role by Regulating miR-26a-5p-NRSF and JAK2-STAT3 Pathway to Improve Autophagy, Inflammation and Oxidative Stress of Cerebral Ischemia-Reperfusion Injury. Drug Des. Dev. Ther. 2020, 14, 3177–3188. [Google Scholar] [CrossRef] [PubMed]
- Zhen, G.; Liang, W.; Jia, H.; Zheng, X. Melatonin relieves sepsis-induced myocardial injury via regulating JAK2/STAT3 signaling pathway. Minerva Med. 2020. [Google Scholar] [CrossRef]
- Lochner, A.; Huisamen, B.; Nduhirabandi, F. Cardioprotective effect of melatonin against ischaemia/reperfusion damage. Front. Biosci. 2013, 5, 305–315. [Google Scholar] [CrossRef]
- Yu, L.; Gong, B.; Duan, W.; Fan, C.; Zhang, J.; Li, Z.; Xue, X.; Xu, Y.; Meng, D.; Li, B.; et al. Melatonin ameliorates myocardial ischemia/reperfusion injury in type 1 diabetic rats by preserving mitochondrial function: Role of AMPK-PGC-1α-SIRT3 signaling. Sci. Rep. 2017, 7, 41337. [Google Scholar] [CrossRef]
- Navarro-Alarcón, M.; Ruiz-Ojeda, F.J.; Blanca-Herrera, R.M.; MM, A.S.; Acuña-Castroviejo, D.; Fernández-Vázquez, G.; Agil, A. Melatonin and metabolic regulation: A review. Food Funct. 2014, 5, 2806–2832. [Google Scholar] [CrossRef]
- Karamitri, A.; Jockers, R. Melatonin in type 2 diabetes mellitus and obesity. Nat. Rev. Endocrinol. 2019, 15, 105–125. [Google Scholar] [CrossRef] [PubMed]
- Cardinali, D.P.; Vigo, D.E. Melatonin, mitochondria, and the metabolic syndrome. Cell. Mol. Life Sci. 2017, 74, 3941–3954. [Google Scholar] [CrossRef] [PubMed]
- Peschke, E.; Bähr, I.; Mühlbauer, E. Melatonin and pancreatic islets: Interrelationships between melatonin, insulin and glucagon. Int. J. Mol. Sci. 2013, 14, 6981–7015. [Google Scholar] [CrossRef] [PubMed]
- Peschke, E.; Wolgast, S.; Bazwinsky, I.; Pönicke, K.; Muhlbauer, E. Increased melatonin synthesis in pineal glands of rats in streptozotocin induced type 1 diabetes. J. Pineal Res. 2008, 45, 439–448. [Google Scholar] [CrossRef]
- Rasmussen, D.D.; Mitton, D.R.; Larsen, S.A.; Yellon, S.M. Aging-dependent changes in the effect of daily melatonin supplementation on rat metabolic and behavioral responses. J. Pineal Res. 2001, 31, 89–94. [Google Scholar] [CrossRef]
- Prunet-Marcassus, B.; Desbazeille, M.; Bros, A.; Louche, K.; Delagrange, P.; Renard, P.; Casteilla, L.; Pénicaud, L. Melatonin reduces body weight gain in Sprague Dawley rats with diet-induced obesity. Endocrinology 2003, 144, 5347–5352. [Google Scholar] [CrossRef] [PubMed]
- Agil, A.; Rosado, I.; Ruiz, R.; Figueroa, A.; Zen, N.; Fernández-Vázquez, G. Melatonin improves glucose homeostasis in young Zucker diabetic fatty rats. J. Pineal Res. 2012, 52, 203–210. [Google Scholar] [CrossRef]
- Rasmussen, D.D.; Boldt, B.M.; Wilkinson, C.W.; Yellon, S.M.; Matsumoto, A.M. Daily melatonin administration at middle age suppresses male rat visceral fat, plasma leptin, and plasma insulin to youthful levels. Endocrinology 1999, 140, 1009–1012. [Google Scholar] [CrossRef] [PubMed]
- Wolden-Hanson, T.; Mitton, D.R.; McCants, R.L.; Yellon, S.M.; Wilkinson, C.W.; Matsumoto, A.M.; Rasmussen, D.D. Daily melatonin administration to middle-aged male rats suppresses body weight, intraabdominal adiposity, and plasma leptin and insulin independent of food intake and total body fat. Endocrinology 2000, 141, 487–497. [Google Scholar] [CrossRef]
- Raskind, M.A.; Burke, B.L.; Crites, N.J.; Tapp, A.M.; Rasmussen, D.D. Olanzapine-induced weight gain and increased visceral adiposity is blocked by melatonin replacement therapy in rats. Neuropsychopharmacology 2007, 32, 284–288. [Google Scholar] [CrossRef]
- Tan, D.X.; Manchester, L.C.; Fuentes-Broto, L.; Paredes, S.D.; Reiter, R.J. Significance and application of melatonin in the regulation of brown adipose tissue metabolism: Relation to human obesity. Obes. Rev. 2011, 12, 167–188. [Google Scholar] [CrossRef]
- Cannon, B.; Nedergaard, J. Brown adipose tissue: Function and physiological significance. Physiol. Rev. 2004, 84, 277–359. [Google Scholar] [CrossRef]
- Hussain, S.A. Effect of melatonin on cholesterol absorption in rats. J. Pineal Res. 2007, 42, 267–271. [Google Scholar] [CrossRef] [PubMed]
- Torii, H.; Shimizu, R.; Tanizaki, Y.; Omiya, Y.; Yamamoto, M.; Kamiike, S.; Yasuda, D.; Hiraoka, Y.; Hashida, T.; Kume, N. Effects of Ramelteon and Other Sleep-Promoting Drugs on Serum Low-Density Lipoprotein and Non-high-density Lipoprotein Cholesterol: A Retrospective Comparative Pilot Study. Biol. Pharm. Bull. 2018, 41, 1778–1790. [Google Scholar] [CrossRef] [PubMed]
- Halpern, B.; Mancini, M.C.; Bueno, C.; Barcelos, I.P.; de Melo, M.E.; Lima, M.S.; Carneiro, C.G.; Sapienza, M.T.; Buchpiguel, C.A.; do Amaral, F.G.; et al. Melatonin Increases Brown Adipose Tissue Volume and Activity in Patients With Melatonin Deficiency: A Proof-of-Concept Study. Diabetes 2019, 68, 947–952. [Google Scholar] [CrossRef] [PubMed]
- Tamura, H.; Nakamura, Y.; Narimatsu, A.; Yamagata, Y.; Takasaki, A.; Reiter, R.J.; Sugino, N. Melatonin treatment in peri- and postmenopausal women elevates serum high-density lipoprotein cholesterol levels without influencing total cholesterol levels. J. Pineal Res. 2008, 45, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Tamura, H.; Takasaki, A.; Miwa, I.; Taniguchi, K.; Maekawa, R.; Asada, H.; Taketani, T.; Matsuoka, A.; Yamagata, Y.; Shimamura, K.; et al. Oxidative stress impairs oocyte quality and melatonin protects oocytes from free radical damage and improves fertilization rate. J. Pineal Res. 2008, 44, 280–287. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Kim, J.H.; Lee, D.C. Urine melatonin levels are inversely associated with sarcopenia in postmenopausal women. Menopause 2014, 21, 39–44. [Google Scholar] [CrossRef]
- Stacchiotti, A.; Favero, G.; Rodella, L.F. Impact of Melatonin on Skeletal Muscle and Exercise. Cells 2020, 9, 288. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, M.I.; Escames, G.; López, L.C.; García, J.A.; Ortiz, F.; López, A.; Acuña-Castroviejo, D. Melatonin administration prevents cardiac and diaphragmatic mitochondrial oxidative damage in senescence-accelerated mice. J. Endocrinol. 2007, 194, 637–643. [Google Scholar] [CrossRef]
- McBride, M.J.; Foley, K.P.; D’Souza, D.M.; Li, Y.E.; Lau, T.C.; Hawke, T.J.; Schertzer, J.D. The NLRP3 inflammasome contributes to sarcopenia and lower muscle glycolytic potential in old mice. Am. J. Physiol. Endocrinol. Metab. 2017, 313, E222–E232. [Google Scholar] [CrossRef]
- Sayed, R.K.A.; Fernández-Ortiz, M.; Diaz-Casado, M.E.; Rusanova, I.; Rahim, I.; Escames, G.; López, L.C.; Mokhtar, D.M.; Acuña-Castroviejo, D. The Protective Effect of Melatonin Against Age-Associated, Sarcopenia-Dependent Tubular Aggregate Formation, Lactate Depletion, and Mitochondrial Changes. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2018, 73, 1330–1338. [Google Scholar] [CrossRef]
- Sayed, R.K.A.; Fernández-Ortiz, M.; Diaz-Casado, M.E.; Aranda-Martínez, P.; Fernández-Martínez, J.; Guerra-Librero, A.; Escames, G.; López, L.C.; Alsaadawy, R.M.; Acuña-Castroviejo, D. Lack of NLRP3 Inflammasome Activation Reduces Age-Dependent Sarcopenia and Mitochondrial Dysfunction, Favoring the Prophylactic Effect of Melatonin. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2019, 74, 1699–1708. [Google Scholar] [CrossRef]
- Wang, W.Z.; Fang, X.H.; Stephenson, L.L.; Baynosa, R.C.; Khiabani, K.T.; Zamboni, W.A. Microcirculatory effects of melatonin in rat skeletal muscle after prolonged ischemia. J. Pineal Res. 2005, 39, 57–65. [Google Scholar] [CrossRef]
- Wang, W.Z.; Fang, X.H.; Stephenson, L.L.; Zhang, X.; Khiabani, K.T.; Zamboni, W.A. Melatonin attenuates I/R-induced mitochondrial dysfunction in skeletal muscle. J. Surg. Res. 2011, 171, 108–113. [Google Scholar] [CrossRef]
- Stratos, I.; Richter, N.; Rotter, R.; Li, Z.; Zechner, D.; Mittlmeier, T.; Vollmar, B. Melatonin restores muscle regeneration and enhances muscle function after crush injury in rats. J. Pineal Res. 2012, 52, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Ostjen, C.A.; Rosa, C.G.S.; Hartmann, R.M.; Schemitt, E.G.; Colares, J.R.; Marroni, N.P. Anti-inflammatory and antioxidant effect of melatonin on recovery from muscular trauma induced in rats. Exp. Mol. Pathol. 2019, 106, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Favero, G.; Trapletti, V.; Bonomini, F.; Stacchiotti, A.; Lavazza, A.; Rodella, L.F.; Rezzani, R. Oral Supplementation of Melatonin Protects against Fibromyalgia-Related Skeletal Muscle Alterations in Reserpine-Induced Myalgia Rats. Int. J. Mol. Sci. 2017, 18, 1389. [Google Scholar] [CrossRef] [PubMed]
- Favero, G.; Bonomini, F.; Franco, C.; Rezzani, R. Mitochondrial Dysfunction in Skeletal Muscle of a Fibromyalgia Model: The Potential Benefits of Melatonin. Int. J. Mol. Sci. 2019, 20, 765. [Google Scholar] [CrossRef]
- Hibaoui, Y.; Reutenauer-Patte, J.; Patthey-Vuadens, O.; Ruegg, U.T.; Dorchies, O.M. Melatonin improves muscle function of the dystrophic mdx5Cv mouse, a model for Duchenne muscular dystrophy. J. Pineal Res. 2011, 51, 163–171. [Google Scholar] [CrossRef]
- McCormick, R.; Vasilaki, A. Age-related changes in skeletal muscle: Changes to life-style as a therapy. Biogerontology 2018, 19, 519–536. [Google Scholar] [CrossRef]
- Ferlazzo, N.; Andolina, G.; Cannata, A.; Costanzo, M.G.; Rizzo, V.; Currò, M.; Ientile, R.; Caccamo, D. Is Melatonin the Cornucopia of the 21st Century? Antioxidants 2020, 9, 1088. [Google Scholar] [CrossRef]
- Moradkhani, F.; Moloudizargari, M.; Fallah, M.; Asghari, N.; Heidari Khoei, H.; Asghari, M.H. Immunoregulatory role of melatonin in cancer. J. Cell. Physiol. 2020, 235, 745–757. [Google Scholar] [CrossRef]
- Carrascal, L.; Nunez-Abades, P.; Ayala, A.; Cano, M. Role of Melatonin in the Inflammatory Process and its Therapeutic Potential. Curr. Pharm. Des. 2018, 24, 1563–1588. [Google Scholar] [CrossRef]
- Gonzalez-Haba, M.G.; Garcia-Mauriño, S.; Calvo, J.R.; Goberna, R.; Guerrero, J.M. High-affinity binding of melatonin by human circulating T lymphocytes (CD4+). FASEB J. 1995, 9, 1331–1335. [Google Scholar] [CrossRef]
- Lopez-Gonzalez, M.A.; Guerrero, J.M.; Sanchez, B.; Delgado, F. Melatonin restores and enhances the human type B tonsillar lymphocyte subset in recurrent acute tonsillitis. Neurosci. Lett. 1998, 247, 131–134. [Google Scholar] [CrossRef]
- García-Mauriño, S.; Pozo, D.; Calvo, J.R.; Guerrero, J.M. Correlation between nuclear melatonin receptor expression and enhanced cytokine production in human lymphocytic and monocytic cell lines. J. Pineal Res. 2000, 29, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Acuña-Castroviejo, D.; Rahim, I.; Acuña-Fernández, C.; Fernández-Ortiz, M.; Solera-Marín, J.; Sayed, R.K.A.; Díaz-Casado, M.E.; Rusanova, I.; López, L.C.; Escames, G. Melatonin, clock genes and mitochondria in sepsis. Cell. Mol. Life Sci. 2017, 74, 3965–3987. [Google Scholar] [CrossRef] [PubMed]
- Mauriz, J.L.; Collado, P.S.; Veneroso, C.; Reiter, R.J.; González-Gallego, J. A review of the molecular aspects of melatonin’s anti-inflammatory actions: Recent insights and new perspectives. J. Pineal Res. 2013, 54, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Calvo, J.R.; Karbownik, M.; Qi, W.; Tan, D.X. Melatonin and its relation to the immune system and inflammation. Ann. N. Y. Acad. Sci. 2000, 917, 376–386. [Google Scholar] [CrossRef]
- Chuang, J.I.; Mohan, N.; Meltz, M.L.; Reiter, R.J. Effect of melatonin on NF-kappa-B DNA-binding activity in the rat spleen. Cell Biol. Int. 1996, 20, 687–692. [Google Scholar] [CrossRef] [PubMed]
- Mohan, N.; Sadeghi, K.; Reiter, R.J.; Meltz, M.L. The neurohormone melatonin inhibits cytokine, mitogen and ionizing radiation induced NF-kappa B. Biochem. Mol. Biol. Int. 1995, 37, 1063–1070. [Google Scholar]
- Deng, W.G.; Tang, S.T.; Tseng, H.P.; Wu, K.K. Melatonin suppresses macrophage cyclooxygenase-2 and inducible nitric oxide synthase expression by inhibiting p52 acetylation and binding. Blood 2006, 108, 518–524. [Google Scholar] [CrossRef]
- Konturek, P.C.; Brzozowski, T.; Konturek, S.J. Stress and the gut: Pathophysiology, clinical consequences, diagnostic approach and treatment options. J. Physiol. Pharmacol. 2011, 62, 591–599. [Google Scholar]
- Konturek, S.J.; Konturek, P.C.; Brzozowski, T. Melatonin in gastroprotection against stress-induced acute gastric lesions and in healing of chronic gastric ulcers. J. Physiol. Pharmacol. 2006, 57 (Suppl. 5), 51–66. [Google Scholar]
- Jaworek, J.; Nawrot-Porabka, K.; Leja-Szpak, A.; Konturek, S.J. Brain-gut axis in the modulation of pancreatic enzyme secretion. J. Physiol. Pharmacol. 2010, 61, 523–531. [Google Scholar]
- Sgambato, D.; Capuano, A.; Sullo, M.G.; Miranda, A.; Federico, A.; Romano, M. Gut-Brain Axis in Gastric Mucosal Damage and Protection. Curr. Neuropharmacol. 2016, 14, 959–966. [Google Scholar] [CrossRef] [PubMed]
- Jaworek, J.; Brzozowski, T.; Konturek, S.J. Melatonin as an organoprotector in the stomach and the pancreas. J. Pineal Res. 2005, 38, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Daryani, A.; Montazeri, M.; Pagheh, A.S.; Sharif, M.; Sarvi, S.; Hosseinzadeh, A.; Reiter, R.J.; Hadighi, R.; Joghataei, M.T.; Ghaznavi, H.; et al. The potential use of melatonin to treat protozoan parasitic infections: A review. Biomed. Pharmacother. 2018, 97, 948–957. [Google Scholar] [CrossRef] [PubMed]
- Scarpelli, P.H.; Tessarin-Almeida, G.; Viçoso, K.L.; Lima, W.R.; Borges-Pereira, L.; Meissner, K.A.; Wrenger, C.; Raffaello, A.; Rizzuto, R.; Pozzan, T.; et al. Melatonin activates FIS1, DYN1, and DYN2 Plasmodium falciparum related-genes for mitochondria fission: Mitoemerald-GFP as a tool to visualize mitochondria structure. J. Pineal Res. 2019, 66, e12484. [Google Scholar] [CrossRef] [PubMed]
- Gazarini, M.L.; Garcia, C.R. The malaria parasite mitochondrion senses cytosolic Ca2+ fluctuations. Biochem. Biophys. Res. Commun. 2004, 321, 138–144. [Google Scholar] [CrossRef]
- Srinivasan, V.; Zakaria, R.; Mohamed, M.; Saleh, R.M. Effects of melatonin derivatives on human malaria parasite Plasmodium falciparum. Recent Pat. Endocr. Metab. Immune Drug Discov. 2014, 8, 102–108. [Google Scholar] [CrossRef]
- Srinivasan, V.; Mohamed, M.; Zakaria, R.; Ahmad, A.H. Malaria, anti malarial drugs and the role of melatonin. Infect. Disord. Drug Targets 2012, 12, 371–379. [Google Scholar] [CrossRef]
- Santello, F.H.; Frare, E.O.; Caetano, L.C.; AlonsoToldo, M.P.; do Prado, J.C., Jr. Melatonin enhances pro-inflammatory cytokine levels and protects against Chagas disease. J. Pineal Res. 2008, 45, 79–85. [Google Scholar] [CrossRef]
- Machado, N.I.; Dos Santos, T.A.T.; de Souza, W.; DaMatta, R.A.; Seabra, S.H. Treatment with melatonin induces a reduction of Toxoplasma gondii development in LLC-MK2 cells. Parasitol. Res. 2020, 119, 2703–2711. [Google Scholar] [CrossRef]
- Kristensson, K.; Claustrat, B.; Mhlanga, J.D.; Møller, M. African trypanosomiasis in the rat alters melatonin secretion and melatonin receptor binding in the suprachiasmatic nucleus. Brain Res. Bull. 1998, 47, 265–269. [Google Scholar] [CrossRef]
- Elmahallawy, E.K.; Jiménez-Aranda, A.; Martínez, A.S.; Rodriguez-Granger, J.; Navarro-Alarcón, M.; Gutiérrez-Fernández, J.; Agil, A. Activity of melatonin against Leishmania infantum promastigotes by mitochondrial dependent pathway. Chem. Biol. Interact. 2014, 220, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Pereira, Q.L.C.; Hara, C.C.P.; Fernandes, R.T.S.; Fagundes, D.L.G.; França-Botelho, A.D.C.; Gomes, M.A.; França, E.L.; Honorio-França, A.C. Human colostrum action against Giardia lamblia infection influenced by hormones and advanced maternal age. Parasitol. Res. 2018, 117, 1783–1791. [Google Scholar] [CrossRef] [PubMed]
- França-Botelho, A.C.; França, J.L.; Oliveira, F.M.; Franca, E.L.; Honório-França, A.C.; Caliari, M.V.; Gomes, M.A. Melatonin reduces the severity of experimental amoebiasis. Parasites Vectors 2011, 4, 62. [Google Scholar] [CrossRef]
- Sen Gupta, P.S.; Rana, M.K. Ivermectin, Famotidine, and Doxycycline: A Suggested Combinatorial Therapeutic for the Treatment of COVID-19. ACS Pharmacol. Transl. Sci. 2020, 3, 1037–1038. [Google Scholar] [CrossRef]
- Khan, M.S.I.; Khan, M.S.I.; Debnath, C.R.; Nath, P.N.; Mahtab, M.A.; Nabeka, H.; Matsuda, S.; Akbar, S.M.F. Ivermectin Treatment May Improve the Prognosis of Patients With COVID-19. Arch. Bronconeumol. 2020, 56, 828–830. [Google Scholar] [CrossRef]
- Hellwig, M.D.; Maia, A. A COVID-19 Prophylaxis? Lower incidence associated with prophylactic administration of Ivermectin. Int. J. Antimicrob. Agents 2020, 106248. [Google Scholar] [CrossRef]
- Mittal, N.; Mittal, R. Inhaled route and anti-inflammatory action of ivermectin: Do they hold promise in fighting against COVID-19? Med. Hypotheses 2020, 110364. [Google Scholar] [CrossRef]
- Padhy, B.M.; Mohanty, R.R.; Das, S.; Meher, B.R. Therapeutic potential of ivermectin as add on treatment in COVID 19: A systematic review and meta-analysis. J. Pharm. Pharm. Sci. 2020, 23, 462–469. [Google Scholar] [CrossRef]
- Yates, P.A.; Newman, S.A.; Oshry, L.J.; Glassman, R.H.; Leone, A.M.; Reichel, E. Doxycycline treatment of high-risk COVID-19-positive patients with comorbid pulmonary disease. Ther. Adv. Respir. Dis. 2020, 14. [Google Scholar] [CrossRef]
- Milionis, H.; Kostikas, K. Is it prime time to consider a clinical trial of doxycycline for the management of COVID-19? Postgrad. Med. J. 2020. [Google Scholar] [CrossRef] [PubMed]
- Gendrot, M.; Andreani, J.; Jardot, P.; Hutter, S.; Delandre, O.; Boxberger, M.; Mosnier, J.; Le Bideau, M.; Duflot, I.; Fonta, I.; et al. In Vitro Antiviral Activity of Doxycycline against SARS-CoV-2. Molecules 2020, 25, 5064. [Google Scholar] [CrossRef] [PubMed]
- Gharebaghi, R.; Heidary, F.; Moradi, M.; Parvizi, M. Metronidazole; a Potential Novel Addition to the COVID-19 Treatment Regimen. Arch. Acad. Emerg. Med. 2020, 8, e40. [Google Scholar] [CrossRef] [PubMed]
- Gendrot, M.; Duflot, I.; Boxberger, M.; Delandre, O.; Jardot, P.; Le Bideau, M.; Andreani, J.; Fonta, I.; Mosnier, J.; Rolland, C.; et al. Antimalarial artemisinin-based combination therapies (ACT) and COVID-19 in Africa: In Vitro inhibition of SARS-CoV-2 replication by mefloquine-artesunate. Int. J. Infect. Dis. 2020, 99, 437–440. [Google Scholar] [CrossRef] [PubMed]
- El Ouarradi, A.; Abdeladim, S.; Oualim, S.; Filali, R.A.; Bensahi, I.; Elharass, M.; Hafid, S.; Tazi, H.; Naitlhou, A.; Bouaiti, E.A.; et al. Hydroxychloroquine and Azithromycin as a Treatment of COVID-19: Electrocardiogram Variability. J. Saudi Heart Assoc. 2020, 32, 350–357. [Google Scholar] [CrossRef] [PubMed]
- Shamshirian, A.; Hessami, A.; Heydari, K.; Navaei, R.A.; Ebrahimzadeh, M.A.; Yip, G.W.; Ghasemian, R.; Sedaghat, M.; Baradaran, H.; Yazdi, S.M.; et al. The Role of Hydroxychloroquine in COVID-19: A Systematic Review and Meta-Analysis. Ann. Acad. Med. Singap. 2020, 49, 789–800. [Google Scholar] [CrossRef] [PubMed]
- Martín Giménez, V.M.; Prado, N.; Diez, E.; Manucha, W.; Reiter, R.J. New proposal involving nanoformulated melatonin targeted to the mitochondria as a potential COVID-19 treatment. Nanomedicine 2020. [Google Scholar] [CrossRef]
- Cardinali, D.P.; Brown, G.M.; Pandi-Perumal, S.R. Can Melatonin Be a Potential “Silver Bullet” in Treating COVID-19 Patients? Diseases 2020, 8, 44. [Google Scholar] [CrossRef]
- Kleszczyński, K.; Slominski, A.T.; Steinbrink, K.; Reiter, R.J. Clinical Trials for Use of Melatonin to Fight against COVID-19 are Urgently Needed. Nutrients 2020, 12, 2561. [Google Scholar] [CrossRef]
- Öztürk, G.; Akbulut, K.G.; Güney, Ş. Melatonin, aging, and COVID-19: Could melatonin be beneficial for COVID-19 treatment in the elderly? Turk. J. Med. Sci. 2020, 50, 1504–1512. [Google Scholar] [CrossRef]
- Romero, A.; Ramos, E.; López-Muñoz, F.; Gil-Martín, E.; Escames, G.; Reiter, R.J. Coronavirus Disease 2019 (COVID-19) and Its Neuroinvasive Capacity: Is It Time for Melatonin? Cell. Mol. Neurobiol. 2020, 1–12. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).