
Oncogenic and Tumor Suppressive Components of the Cell Cycle in Breast Cancer Progression and Prognosis

 ,
, 
Abstract
:1. Introduction
2. Overview of Cell Cycle
3. Oncogenic Components of Cell Cycle
3.1. Cyclin D
3.2. Cyclin A
3.3. Cyclin E
3.4. Cyclin B
3.5. CDK2
4. Tumor Suppressive Components of Cell Cycle
4.1. p16(INK4A/MTS-1/CDKN2A)
4.2. p21 (WAF1/CIP1/SDI1/MDA-6)
4.3. p27 (Kip1)
4.4. p53 (Wild Type)
4.5. p53 (Mutant)
5. Future Perspectives
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 5GG | Pentagalloylglucose |
| ABCSG | Austrian breast and colorectal cancer study group |
| AC | Adenocarcinoma |
| c-AMP | Cyclic adenosine monophosphate |
| CCND1 | Cyclin D1 |
| CDC2 kinase | Cell division control 2 kinase |
| CDKs | Cyclin-dependent kinases |
| CI | Confidence interval |
| DC | Ductal carcinoma |
| DCIS | Ductal carcinoma in situ |
| DFS | Disease-free survival |
| DSS | Disease-specific survival |
| E2F1 | E2 promoter factor 1 |
| ER | Estrogen receptor |
| FISH | Fluorescent in situ hybridization |
| GLOBOCAN | Global cancer observatory |
| HDAC3 | Histone deacetylase 3 |
| HER2+ | Human epidermal growth factor receptor2+ |
| HR | Hazard ratio |
| IDC | Invasive ductal carcinoma |
| IHC | Immunohistochemistry |
| ILC | Invasive lobular carcinoma |
| LA | Luminal A |
| MU | Mutation |
| NBE | Normal breast epithelium |
| ND | Not decided |
| OS | Overall survival |
| PD | Proliferative disease |
| PR | Progesterone receptor |
| Rb | Retinoblastoma |
| RFS | Recurrence free survival |
| siRNA | Small interfering RNA |
| TGF-β | Transforming growth factor-β |
| TNBC | Triple negative breast cancer |
| WT | Wild type |
References
- Zhivotovsky, B.; Orrenius, S. Cell cycle and cell death in disease: Past, present and future. J. Intern. Med. 2010, 268, 395–409. [Google Scholar] [CrossRef]
- Witsch, E.; Sela, M.; Yarden, Y. Roles for Growth Factors in Cancer Progression. Physiology 2010, 25, 85–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- You, J.S.; Jones, P.A. Cancer Genetics and Epigenetics: Two Sides of the Same Coin? Cancer Cell 2012, 22, 9–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baylin, S.B.; Jones, P.A. Epigenetic Determinants of Cancer. Cold Spring Harb. Perspect. Biol. 2016, 8, a019505. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Goel, N. Yogita Integrative Analysis of Multi-Genomic Data for Kidney Renal Cell Carcinoma. Interdiscip. Sci. Comput. Life Sci. 2020, 12, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Goel, N.; Khandnor, P. TCGA: A multi-genomics material repository for cancer research. In Materials Today: Proceedings; Elsevier: Amsterdam, The Netherlands, 2020; pp. 1492–1495. [Google Scholar]
- Lee, S.Y.; Ju, M.K.; Jeon, H.M.; Lee, Y.J.; Kim, C.H.; Park, H.G.; Han, S.I.; Kang, H.S. Oncogenic Metabolism Acts as a Prerequisite Step for Induction of Cancer Metastasis and Cancer Stem Cell Phenotype. Oxidative Med. Cell. Longev. 2018, 2018, 1–28. [Google Scholar] [CrossRef] [Green Version]
- Kashyap, D.; Tuli, H.S.; Garg, V.K.; Goel, N.; Bishayee, A. Oncogenic and Tumor-Suppressive Roles of MicroRNAs with Special Reference to Apoptosis: Molecular Mechanisms and Therapeutic Potential. Mol. Diagn. Ther. 2018, 22, 179–201. [Google Scholar] [CrossRef]
- Kashyap, D.; Kaur, H. Cell-free miRNAs as non-invasive biomarkers in breast cancer: Significance in early diagnosis and metastasis prediction. Life Sci. 2020, 246, 117417. [Google Scholar] [CrossRef]
- Mitra, S.; Bal, A.; Kashyap, D.; Kumar, S.; Shrivastav, S.; Das, A.; Singh, G. Tumour angiogenesis and c-Met pathway activation –implications in breast cancer. APMIS 2020, 128, 316–325. [Google Scholar] [CrossRef]
- Wang, L.-H.; Wu, C.-F.; Rajasekaran, N.; Shin, Y.K. Loss of Tumor Suppressor Gene Function in Human Cancer: An Overview. Cell. Physiol. Biochem. 2018, 51, 2647–2693. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.Y.H.P.; Muller, W.J. Oncogenes and Tumor Suppressor Genes. Cold Spring Harb. Perspect. Biol. 2010, 2, a003236. [Google Scholar] [CrossRef] [Green Version]
- Velez, A.M.A.; Howard, M.S. Tumor-suppressor genes, cell cycle regulatory checkpoints, and the skin. N. Am. J. Med Sci. 2015, 7, 176–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deshpande, A.; Sicinski, P.; Hinds, P.W. Cyclins and cdks in development and cancer: A perspective. Oncogene 2005, 24, 2909–2915. [Google Scholar] [CrossRef]
- Law, M.E.; Corsino, P.E.; Narayan, S.; Law, B.K. Cyclin-Dependent Kinase Inhibitors as Anticancer Therapeutics. Mol. Pharmacol. 2015, 88, 846–852. [Google Scholar] [CrossRef] [Green Version]
- Yixiao, F.; Mia, S.; Shifeng, H.; Chengfu, Y.; Zongyue, Z.; Linghuan, Z.; Xiaojuan, J.; Wei, L.; Bo, H.; Wenping, L.; et al. Breast cancer development and progression: Risk factors, cancer stem cells, signaling pathways, genomics, and molecular pathogenesis. Genes Dis. 2018, 5, 77–106. [Google Scholar] [CrossRef]
- Aggarwal, V.; Kashyap, D.; Sak, K.; Tuli, H.S.; Jain, A.; Chaudhary, A.; Garg, V.K.; Sethi, G.; Yerer, M.B. Molecular Mechanisms of Action of Tocotrienols in Cancer: Recent Trends and Advancements. Int. J. Mol. Sci. 2019, 20, 656. [Google Scholar] [CrossRef] [Green Version]
- Mehta, S.; Shelling, A.; Muthukaruppan, A.; Lasham, A.; Blenkiron, C.; Laking, G.; Print, C. Predictive and prognostic molecular markers for cancer medicine. Ther. Adv. Med Oncol. 2010, 2, 125–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caldon, C.E.; Daly, R.J.; Sutherland, R.L.; Musgrove, E.A. Cell cycle control in breast cancer cells. J. Cell. Biochem. 2005, 97, 261–274. [Google Scholar] [CrossRef]
- Haase, S.B.; Wittenberg, C. Topology and Control of the Cell-Cycle-Regulated Transcriptional Circuitry. Genetics 2014, 196, 65–90. [Google Scholar] [CrossRef] [Green Version]
- Cheung, T.H.T.; Rando, T.A. Molecular regulation of stem cell quiescence. Nat. Rev. Mol. Cell Biol. 2013, 14, 329–340. [Google Scholar] [CrossRef] [PubMed]
- Enders, G.H. Mammalian Interphase Cdks: Dispensable Master Regulators of the Cell Cycle. Genes Cancer 2012, 3, 614–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertoli, C.; Skotheim, J.M.; De Bruin, R.A.M. Control of cell cycle transcription during G1 and S phases. Nat. Rev. Mol. Cell Biol. 2013, 14, 518–528. [Google Scholar] [CrossRef] [Green Version]
- Cynthia, O.; Paschal, W.; Debu, T. Oncogenes and tumor suppressor genes in breast cancer: Potential diagnostic and therapeutic applications. Oncologist. 2004, 9, 361–377. [Google Scholar] [CrossRef]
- Kato, J.; Matsushime, H.; Hiebert, S.W.; Ewen, E.M.; Sherr, C.J. Direct binding of cyclin D to the retinoblastoma gene product (pRb) and pRb phosphorylation by the cyclin D-dependent kinase CDK4. Genes Dev. 1993, 7, 331–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiman, K.G. The retinoblastoma gene: Role in cell cycle control and cell differentiation. FASEB J. 1993, 7, 841–845. [Google Scholar] [CrossRef]
- Coqueret, O. Linking cyclins to transcriptional control. Gene 2002, 299, 35–55. [Google Scholar] [CrossRef]
- Diehl, A.J.; Zindy, F.; Sherr, C.J. Inhibition of cyclin D1 phosphorylation on threonine-286 prevents its rapid degradation via the ubiquitin-proteasome pathway. Genes Dev. 1997, 11, 957–972. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.-K.; Gervais, J.L.M.; Zhang, H. Human CUL-1 associates with the SKP1/SKP2 complex and regulates p21CIP1/WAF1 and cyclin D proteins. Proc. Natl. Acad. Sci. USA 1998, 95, 11324–11329. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Li, Z.; Lu, Y.; Du, R.; Katiyar, S.; Yang, J.; Fu, M.; Leader, J.E.; Quong, A.; Novikoff, P.M.; et al. Cyclin D1 repression of nuclear respiratory factor 1 integrates nuclear DNA synthesis and mitochondrial function. Proc. Natl. Acad. Sci. USA 2006, 103, 11567–11572. [Google Scholar] [CrossRef] [Green Version]
- Hall, M.; Peters, G. Genetic Alterations of Cyclins, Cyclin-Dependent Kinases, and Cdk Inhibitors in Human Cancer. In Advances in Cancer Research; Elsevier: Amsterdam, The Netherlands, 1996; pp. 67–108. [Google Scholar]
- Yamamoto, M.; Tamakawa, S.; Yoshie, M.; Yaginuma, Y.; Ogawa, K. Neoplastic hepatocyte growth associated with cyclin D1 redistribution from the cytoplasm to the nucleus in mouse hepatocarcinogenesis. Mol. Carcinog. 2006, 45, 901–913. [Google Scholar] [CrossRef] [Green Version]
- Hodges, L.C.; Cook, J.D.; Lobenhofer, E.K.; Li, L.; Bennett, L.; Bushel, P.R.; Aldaz, C.M.; Afshari, A.C.; Walker, C.L. Tamoxifen functions as a molecular agonist inducing cell cycle-associated genes in breast cancer cells. Mol. Cancer Res. 2003, 1, 300–311. [Google Scholar]
- Kenny, F.S.; Hui, R.; Musgrove, E.A.; Gee, J.M.; Blamey, R.W.; Nicholson, R.I.; Sutherland, R.L.; Robertson, J.F. Overexpression of cyclin D1 messenger RNA predicts for poor prognosis in estrogen receptor-positive breast cancer. Clin Cancer Res. 1999, 5, 2069–2076. [Google Scholar] [PubMed]
- Mohammadizadeh, F.; Hani, M.; Ranaee, M.; Bagheri, M. Role of cyclin D1 in breast carcinoma. J. Res. Med Sci. 2013, 18, 1021–1025. [Google Scholar] [PubMed]
- Barnes, D.M.; Gillett, C.E. Cyclin D1 in Breast Cancer. Breast Cancer Res. Treat. 1998, 52, 1–15. [Google Scholar] [CrossRef] [PubMed]
- A Velasco-Velázquez, M.; Li, Z.; Casimiro, M.; Loro, E.; Homsi, N.; Pestell, R.G. Examining the role of cyclin D1 in breast cancer. Futur. Oncol. 2011, 7, 753–765. [Google Scholar] [CrossRef] [PubMed]
- Ravikumar, G.; Ananthamurthy, A. Cyclin D1 expression in ductal carcinoma of the breast and its correlation with other prognostic parameters. J. Cancer Res. Ther. 2014, 10, 671–675. [Google Scholar] [PubMed]
- Assem, M.; Youssef, E.A.; Rashad, R.M.; Yahia, M.A.-H. Immunohistochemical Expression of Cyclin D1 in Invasive Ductal Carcinoma of Human Breast. Oncomedicine 2017, 2, 80–87. [Google Scholar] [CrossRef]
- Buckley, M.F.; Sweeney, K.J.; Hamilton, A.J.; Sini, R.L.; Manning, D.L.; Nicholson, I.R.; DeFazio, A.; Watts, C.K.; Musgrove, A.E.; Sutherland, R.L. Expression and amplification of cyclin genes in human breast cancer. Oncogene 1993, 8, 2127–2133. [Google Scholar]
- Lebwohl, E.D.; Muise-Helmericks, R.; Sepp-Lorenzino, L.; Serve, S.; Timaul, M.; Bol, R.; Borgen, P.; Rosen, N. A truncated cyclin D1 gene encodes a stable mRNA in a human breast cancer cell line. Oncogene 1994, 9, 1925–1929. [Google Scholar]
- Zukerberg, L.R.; Yang, I.W.; Gadd, M.; Thor, A.D.; Koerner, F.C.; Schmidt, E.V.; Arnold, A. Cyclin D1 (PRAD1) protein expression in breast cancer: Approximately one-third of infiltrating mammary carcinomas show overexpression of the cyclin D1 oncogene. Mod. Pathol. 1995, 8, 560–567. [Google Scholar]
- Khan, N.; Syed, D.N.; Ahmad, N.; Mukhtar, H. Fisetin: A Dietary Antioxidant for Health Promotion. Antioxid. Redox Signal. 2013, 19, 151–162. [Google Scholar] [CrossRef]
- Rosendahl, A.H.; Perks, C.M.; Zeng, L.; Markkula, A.; Simonsson, M.; Rose, C.; Ingvar, C.; Holly, J.M.; Jernström, H. Caffeine and Caffeic Acid Inhibit Growth and Modify Estrogen Receptor and Insulin-like Growth Factor I Receptor Levels in Human Breast Cancer. Clin. Cancer Res. 2015, 21, 1877–1887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.-Y.; Caamano, J.; Cooper, F.; Guo, X.; Klein-Szanto, A.J. Immunohistochemistry of Cyclin D1 in Human Breast Cancer. Am. J. Clin. Pathol. 1994, 102, 695–698. [Google Scholar] [CrossRef] [Green Version]
- Wilcken, N.R.; Prall, O.W.; Musgrove, A.E.; Sutherland, R.L. Inducible overexpression of cyclin D1 in breast cancer cells reverses the growth-inhibitory effects of antiestrogens. Clin. Cancer Res. 1997, 3, 849–854. [Google Scholar] [PubMed]
- Jirström, K.; Stendahl, M.; Rydén, L.; Kronblad, A.; Bendahl, P.-O.; Stal, O.; Landberg, G. Adverse Effect of Adjuvant Tamoxifen in Premenopausal Breast Cancer with Cyclin D1 Gene Amplification. Cancer Res. 2005, 65, 8009–8016. [Google Scholar] [CrossRef] [Green Version]
- Courjal, F.; Louason, G.; Speiser, P.; Katsaros, D.; Zeillinger, R.; Theillet, C. Cyclin gene amplification and overexpression in breast and ovarian cancers: Evidence for the selection of cyclin D1 in breast and cyclin E in ovarian tumors. Int. J. Cancer 1996, 69, 247–253. [Google Scholar] [CrossRef]
- Umekita, Y.; Ohi, Y.; Sagara, Y.; Yoshida, H. Overexpression of cyclinD1 predicts for poor prognosis in estrogen receptor-negative breast cancer patients. Int. J. Cancer 2002, 98, 415–418. [Google Scholar] [CrossRef] [PubMed]
- Elsheikh, S.; Green, A.R.; Aleskandarany, M.A.; Grainge, M.; Paish, C.E.; Lambros, M.B.K.; Reis-Filho, J.S.; Ellis, I.O. CCND1 amplification and cyclin D1 expression in breast cancer and their relation with proteomic subgroups and patient outcome. Breast Cancer Res. Treat. 2007, 109, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Rudas, M.; Lehnert, M.; Huynh, A.; Jakesz, R.; Singer, C.; Lax, S.; Schippinger, W.; Dietze, O.; Greil, R.; Stiglbauer, W.; et al. Cyclin D1 Expression in Breast Cancer Patients Receiving Adjuvant Tamoxifen-Based Therapy. Clin. Cancer Res. 2008, 14, 1767–1774. [Google Scholar] [CrossRef] [Green Version]
- Alle, K.M.; Henshall, S.M.; Field, A.S.; Sutherland, R.L. Cyclin D1 protein is overexpressed in hyperplasia and intraductal carcinoma of the breast. Clin. Cancer Res. 1998, 4, 847–854. [Google Scholar] [PubMed]
- Heiss, C.; Schanz, A.; Amabile, N.; Jahn, S.; Chen, Q.; Wong, M.L.; Rassaf, T.; Heinen, Y.; Cortese-Krott, M.; Grossman, W.; et al. Nitric Oxide Synthase Expression and Functional Response to Nitric Oxide Are Both Important Modulators of Circulating Angiogenic Cell Response to Angiogenic Stimuli. Arter. Thromb. Vasc. Biol. 2010, 30, 2212–2218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oyama, T.; Kashiwabara, K.; Yoshimoto, K.; Arnold, A.; Koerner, F. Frequent overexpression of the cyclin D1 oncogene in invasive lobular carcinoma of the breast. Cancer Res. 1998, 58, 2876–2880. [Google Scholar] [PubMed]
- Ahnström, M.; Nordenskjöld, B.; Rutqvist, L.E.; Skoog, L.; Stål, O. Role of cyclin D1 in ErbB2-positive breast cancer and tamoxifen resistance. Breast Cancer Res. Treat. 2005, 91, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Simpson, J.F.; Quan, D.E.; O’Malley, F.; Odom-Maryon, T.; Clarke, P.E. Amplification of CCND1 and expression of its protein product, cyclin D1, in ductal carcinoma in situ of the breast. Am. J. Pathol. 1997, 151, 161–168. [Google Scholar]
- Tobin, N.P.; Bergh, J. Analysis of Cyclin D1 in Breast Cancer: A Call to Arms. Curr. Breast Cancer Rep. 2012, 4, 171–173. [Google Scholar] [CrossRef] [Green Version]
- Boström, P.; Söderström, M.; Palokangas, T.; Vahlberg, T.; Collan, Y.; Carpén, O.; Hirsimäki, P. Analysis of cyclins A, B1, D1 and E in breast cancer in relation to tumour grade and other prognostic factors. BMC Res. Notes 2009, 2, 140. [Google Scholar] [CrossRef] [Green Version]
- Dai, M.; Al-Odaini, A.A.; Fils-Aimé, N.; Villatoro, A.M.; Guo, J.; Arakelian, A.; Rabbani, A.S.; Ali, S.; Lebrun, J.J. Cyclin D1 cooperates with p21 to regulate TGFβ-mediated breast cancer cell migration and tumor local invasion. Breast Cancer Res. 2013, 15, R49. [Google Scholar] [CrossRef] [Green Version]
- Russell, A.F.; Thompson, A.M.; Hendley, J.; Trute, L.; Armes, E.J.; Germain, D.S. Cyclin D1 and D3 associate with the SCF complex and are coordinately elevated in breast cancer. Oncogene 1999, 18, 1983–1991. [Google Scholar] [CrossRef] [Green Version]
- Khandeparkar, S.G.S.; Lengare, P.V.; Joshi, A.R.; Gogate, B.P.; Solanke, S.G.; Gore, S.H. Immunohistochemical expression of cyclin D1 in invasive breast carcinoma and its correlation with clinicopathological parameters. Indian J. Pathol. Microbiol. 2020, 63, 376. [Google Scholar] [CrossRef]
- Husdal, A.; Bukholm, G.; Bukholm, I.R.K. The Prognostic Value and Overexpression of Cyclin A Is Correlated with Gene Amplification of both Cyclin A and Cyclin E in Breast Cancer Patient. Cell. Oncol. 2006, 28, 107–116. [Google Scholar] [CrossRef]
- Baldini, E.; Camerini, A.; Sgambato, A.; Prochilo, T.; Capodanno, A.; Pasqualetti, F.; Orlandini, C.; Resta, L.; Bevilacqua, G.; Collecchi, P. Cyclin A and E2F1 overexpression correlate with reduced disease-free survival in node-negative breast cancer patients. Anticancer. Res. 2007, 26, 4415–4421. [Google Scholar]
- Michalides, R.; Van Tinteren, H.; Balkenende, A.; Vermorken, J.B.; Benraadt, J.; Huldij, J.; Van Diest, P. Cyclin A is a prognostic indicator in early stage breast cancer with and without tamoxifen treatment. Br. J. Cancer 2002, 86, 402–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poikonen, P.; Sjöström, J.; Amini, R.-M.; Villman, K.; Ahlgren, J.; Blomqvist, C. Cyclin A as a marker for prognosis and chemotherapy response in advanced breast cancer. Br. J. Cancer 2005, 93, 515–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, N.; Arnerlöv, C.; Emdin, S.; Landberg, G. Cyclin E overexpression, a negative prognostic factor in breast cancer with strong correlation to oestrogen receptor status. Br. J. Cancer 1996, 74, 874–880. [Google Scholar] [CrossRef] [Green Version]
- Sieuwerts, A.M.; Look, M.P.; Gelder, M.E.M.-V.; Timmermans, M.; Trapman, A.M.; Garcia, R.R.; Arnold, M.; Goedheer, A.J.; De Weerd, V.; Portengen, H.; et al. Which Cyclin E Prevails as Prognostic Marker for Breast Cancer? Results from a Retrospective Study Involving 635 Lymph Node–Negative Breast Cancer Patients. Clin. Cancer Res. 2006, 12, 3319–3328. [Google Scholar] [CrossRef] [Green Version]
- Donnellan, R.; Kleinschmidt, I.; Chetty, R. Cyclin E immunoexpression in breast ductal carcinoma: Pathologic correlations and prognostic implications. Hum. Pathol. 2001, 32, 89–94. [Google Scholar] [CrossRef]
- Potemski, P.; Kusinska, R.; Watala, C.; Pluciennik, E.; Bednarek, A.K.; Kordek, R. Cyclin E expression in breast cancer correlates with negative steroid receptor status, HER2 expression, tumor grade and proliferation. J. Exp. Clin. Cancer Res. 2006, 25, 59–64. [Google Scholar] [PubMed]
- Han, S.; Park, K.; Bae, B.-N.; Kim, K.H.; Kim, H.-J.; Kim, Y.-D. Prognostic implication of cyclin E expression and its relationship with cyclin D1 and p27Kip1 expression on tissue microarrays of node negative breast cancer. J. Surg. Oncol. 2003, 83, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Potemski, P.; Kusińska, R.; Pasz-Walczak, G.; Piekarski, J.H.; Watała, C.; Płuciennik, E.; Bednarek, A.K.; Kordek, R. Prognostic relevance of cyclin E expression in operable breast cancer. Med Sci. Monit. 2009, 15, 34–40. [Google Scholar]
- Niméus-Malmström, E.; Koliadi, A.; Ahlin, C.; Holmqvist, M.; Holmberg, L.; Amini, R.-M.; Jirström, K.; Wärnberg, F.; Blomqvist, C.; Fernö, M.; et al. Cyclin B1 is a prognostic proliferation marker with a high reproducibility in a population-based lymph node negative breast cancer cohort. Int. J. Cancer 2009, 127, 961–967. [Google Scholar] [CrossRef]
- Sun, X.; Zhangyuan, G.; Shi, L.; Wang, Y.; Sun, B.; Ding, Q. Prognostic and clinicopathological significance of cyclin B expression in patients with breast cancer. Medicine 2017, 96, e6860. [Google Scholar] [CrossRef] [PubMed]
- Aaltonen, K.; Amini, R.-M.; Heikkilä, P.; Aittomäki, K.; Tamminen, A.; Nevanlinna, H.; Blomqvist, C. High cyclin B1 expression is associated with poor survival in breast cancer. Br. J. Cancer 2009, 100, 1055–1060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Göhring, U.-J.; Schöndorf, T.; Kiecker, V.R.; Becker, M.; Kurbacher, C.; Scharl, A. Immunohistochemical detection of H-ras protooncoprotein p21 indicates favorable prognosis in node-negative breast cancer patients. Tumor Biol. 1999, 20, 173–183. [Google Scholar] [CrossRef]
- Domagala, W.; Welcker, M.; Chosia, M.; Karbowniczek, M.; Harezga, B.; Bartkova, J.; Bartek, J.; Osborn, M. p21/WAF1/Cip1 expression in invasive ductal breast carcinoma: Relationship to p53, proliferation rate, and survival at 5 years. Virchows Arch. 2001, 439, 132–140. [Google Scholar] [CrossRef]
- Fayed, Y.M.; El-Sheikh, S.A.-M.; Abulkheir, I.L.H.; Khorshed, I.N.A.-H. Immunohistochemical expression of p27 in ductal carcinoma of breast and its correlation with HER2/neu expression and hormonal status. Egypt. J. Pathol. 2012, 32, 33–41. [Google Scholar] [CrossRef]
- Wei, C.-Y.; Tan, Q.-X.; Zhu, X.; Qin, Q.-H.; Zhu, F.-B.; Mo, Q.-G.; Yang, W.-P. Expression of CDKN1A/p21 and TGFBR2 in breast cancer and their prognostic significance. Int. J. Clin. Exp. Pathol. 2015, 8, 14619–14629. [Google Scholar]
- Alkarain, A.; Jordan, R.; Slingerland, J. p27 Deregulation in Breast Cancer: Prognostic Significance and Implications for Therapy. J. Mammary Gland. Biol. Neoplasia 2004, 9, 67–80. [Google Scholar] [CrossRef]
- Tsuchiya, A.; Zhang, G.J.; Kanno, M. Prognostic impact of cyclin-dependent kinase inhibitor p27kip1 in node-positive breast cancer. J. Surg. Oncol. 1999, 70, 230–234. [Google Scholar] [CrossRef]
- Traub, F.; Mengel, M.; Lück, H.J.; Kreipe, H.H.; Von Wasielewski, R. Prognostic impact of Skp2 and p27 in human breast cancer. Breast Cancer Res. Treat. 2006, 99, 185–191. [Google Scholar] [CrossRef]
- Newman, L.; Xia, W.; Yang, H.-Y.; Sahin, A.; Bondy, M.; Lukmanji, F.; Hung, M.-C.; Lee, M.-H. Correlation of p27 protein expression with HER-2/neu expression in breast cancer. Mol. Carcinog. 2001, 30, 169–175. [Google Scholar] [CrossRef]
- Deepu, M.; Daniel, F.R.; Giorgio, I.; Anne, Z.J.; Joan, C.; Amber, G.; Rachael, A.S.; Silvia, C.F.; Michele, P.; Franco, M. Loss of p27KIP1 expression in fully-staged node-negative breast cancer: Association with lack of hormone receptors in T1a/b, but not T1c infiltrative ductal carcinoma. Anticancer Res. 2011, 12, 4401–4405. [Google Scholar]
- Leivonen, M.; Nordling, S.; Lundin, J.; Von Boguslawski, K.; Haglund, C. p27 expression correlates with short-term, but not with long-term prognosis in breast cancer. Breast Cancer Res. Treat. 2001, 67, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Barnes, A.; Pinder, S.; Bell, J.; Paish, E.; Wencyk, P.; Robertson, J.; Elston, C.; Ellis, I. Expression of p27kip1 in breast cancer and its prognostic significance. J. Pathol. 2003, 201, 451–459. [Google Scholar] [CrossRef]
- Pohl, G.; Rudas, M.; Dietze, O.; Lax, S.; Markis, E.; Pirker, R.; Zielinski, C.C.; Hausmaninger, H.; Kubista, E.; Samonigg, H.; et al. High p27Kip1 Expression Predicts Superior Relapse-Free and Overall Survival for Premenopausal Women With Early-Stage Breast Cancer Receiving Adjuvant Treatment With Tamoxifen Plus Goserelin. J. Clin. Oncol. 2003, 21, 3594–3600. [Google Scholar] [CrossRef] [PubMed]
- Nohara, T.; Ryo, T.; Iwamoto, S.; Gon, G.; Tanigawa, N. Expression of Cell-Cycle Regulator p27 Is Correlated to the Prognosis and ER Expression in Breast Carcinoma Patients. Oncology 2001, 60, 94–100. [Google Scholar] [CrossRef]
- Spataro, V.J.; Litman, H.; Viale, G.; Maffini, F.; Masullo, M.; Golouh, R.; Martínez-Tello, F.J.; Grigolato, P.; Shilkin, K.B.; Gusterson, B.A.; et al. Decreased immunoreactivity for p27 protein in patients with early-stage breast carcinoma is correlated with HER-2/neuoverexpression and with benefit from one course of perioperative chemotherapy in patients with negative lymph node status. Cancer 2003, 97, 1591–1600. [Google Scholar] [CrossRef]
- Davidoff, A.M.; Herndon, E.J.; Glover, N.S.; Kerns, B.J.; Pence, J.C.; Iglehart, J.D.; Marks, J.R. Relation between p53 overexpression and established prognostic factors in breast cancer. Surgery 1991, 110, 259–264. [Google Scholar]
- Porter, P.L.; Barlow, W.E.; Yeh, I.-T.; Lin, M.G.; Yuan, X.P.; Donato, E.; Sledge, G.W.; Shapiro, C.L.; Ingle, J.N.; Haskell, C.M.; et al. p27 Kip1 and Cyclin E Expression and Breast Cancer Survival After Treatment With Adjuvant Chemotherapy. J. Natl. Cancer Inst. 2006, 98, 1723–1731. [Google Scholar] [CrossRef] [Green Version]
- Chu, J.-S.; Huang, C.-S.; Chang, K.-J. p27 expression as a prognostic factor of breast cancer in Taiwan. Cancer Lett. 1999, 141, 123–130. [Google Scholar] [CrossRef]
- Cui, S.-P.; Wang, H.-L.; Peng, W.; Liu, H.-J.; Hou, L.; Zhang, B. Aberrant expression and correlative analysis of P16 in breast cancers. Beijing da xue xue bao. Yi xue ban = J. Peking Univ. Heal. Sci. 2012, 44, 755–759. [Google Scholar]
- Emig, R.; Magener, A.; Ehemann, V.; Meyer, A.; Stilgenbauer, F.; Volkmann, M.; Wallwiener, D.; Sinn, H.P. Aberrant cytoplasmic expression of the p16 protein in breast cancer is associated with accelerated tumour proliferation. Br. J. Cancer 1998, 78, 1661–1668. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.; Yuan, Y.; Liu, G.; Wei, Y. P53 and Ki-67 as prognostic markers in triple-negative breast cancer patients. PLoS ONE 2017, 12, e0172324. [Google Scholar] [CrossRef] [PubMed]
- Al-Joudi, F.S.; Iskandar, A.Z.; Rusli, J. The expression of p53 in invasive ductal carcinoma of the breast: A study in the North-East States of Malaysia. Med. J. Malays. 2008, 63, 96–99. [Google Scholar]
- Shan, M.; Zhang, X.; Liu, X.; Qin, Y.; Liu, T.; Liu, Y.; Wang, J.; Zhong, Z.; Zhang, Y.; Geng, J.; et al. P16 and P53 Play Distinct Roles in Different Subtypes of Breast Cancer. PLoS ONE 2013, 8, e76408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abou-Bakr, A.A.; Eldweny, I.H. P16 expression correlates with basal-like triple-negative breast carcinoma. Ecancermedicalscience 2013, 7, 317. [Google Scholar] [CrossRef] [PubMed]
- Harbhajanka, A.; Lamzabi, I.; Bitterman, P.; Reddy, V.B.; Ghai, R.; Gattuso, P. Correlation of p16 Expression on Cancer and Stromal Cells With Clinicopathologic and Immunohistochemical Features of Lobular Breast Carcinoma. Appl. Immunohistochem. Mol. Morphol. 2019, 27, 658–662. [Google Scholar] [CrossRef] [PubMed]
- Salmani, H.; Hosseini, A.; Azarnezhad, A.; Ahmad, H. PTEN and p53 gene expressions in breast cancer specimens and their clinicopathological significance. Middle East J. Cancer 2018, 9, 105–111. [Google Scholar] [CrossRef]
- Yang, P.; Du, C.W.; Kwan, M.; Liang, S.X.; Zhang, G.J. The impact of p53 in predicting clinical outcome of breast cancer patients with visceral metastasis. Sci. Rep. 2013, 3, srep02246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertheau, P.; Lehmann-Che, J.; Varna, M.; Dumay, A.; Poirot, B.; Porcher, R.; Turpin, E.; Plassa, L.-F.; De Roquancourt, A.; Bourstyn, E.; et al. p53 in breast cancer subtypes and new insights into response to chemotherapy. Breast 2013, 22, S27–S29. [Google Scholar] [CrossRef]
- Li, J.-P.; Zhang, X.-M.; Zhang, Z.; Zheng, L.-H.; Jindal, S.; Liu, Y.-J. Association of p53 expression with poor prognosis in patients with triple-negative breast invasive ductal carcinoma. Medicine 2019, 98, e15449. [Google Scholar] [CrossRef]
- Lee, D.-S.; Kim, S.H.; Suh, Y.J.; Kim, H.K.; Shim, B.Y. Clinical Implication of p53 Overexpression in Breast Cancer Patients Younger than 50 Years with a Triple-negative Subtype Who Undergo a Modified Radical Mastectomy. Jpn. J. Clin. Oncol. 2011, 41, 854–866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kikuchi, S.; Nishimura, R.; Osako, T.; Okumura, Y.; Nishiyama, Y.; Toyozumi, Y.; Arima, N. Definition of p53 overexpression and its association with the clinicopathological features in luminal/HER2-negative breast cancer. Anticancer. Res. 2013, 33, 3891–3898. [Google Scholar] [PubMed]
- Coates, A.S.; Millar, E.K.; O’Toole, S.A.; Molloy, T.J.; Viale, G.; Goldhirsch, A.; Regan, M.M.; Gelber, R.D.; Sun, Z.; Castiglione-Gertsch, M.; et al. Prognostic interaction between expression of p53 and estrogen receptor in patients with node-negative breast cancer: Results from IBCSG Trials VIII and IX. Breast Cancer Res. 2012, 14, R143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abubakar, M.; Guo, C.; Koka, H.; Sung, H.; Shao, N.; Guida, J.; Deng, J.; Li, M.; Hu, N.; Zhou, B.; et al. Clinicopathological and epidemiological significance of breast cancer subtype reclassification based on p53 immunohistochemical expression. NPJ Breast Cancer 2019, 5. [Google Scholar] [CrossRef]
- Muhammad, E.M.S.; Ahmad, A.N.; Guirguis, M.N.; Ali, A.-E.M. Immunohistochemical P53 Expression in Breast Carcinoma with Correlation to Clinico-Pathological Parameters. Med. J. Cairo Univ. 2012, 80, 179–189. [Google Scholar]
- Bae, S.Y.; Lee, J.H.; Bae, J.W.; Jung, S.P. Differences in prognosis by p53 expression after neoadjuvant chemotherapy in triple-negative breast cancer. Ann. Surg. Treat. Res. 2020, 98, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Nema, S.; Mehta, P.; Narang, S. Role of p53 and Ki-67 in prognostication of carcinoma breast. Indian J. Pathol. Oncol. 2019, 6, 261–265. [Google Scholar] [CrossRef]
- Lara, J.F.; Thor, A.D.; Dressler, L.G.; Broadwater, G.; Bleiweiss, I.J.; Edgerton, S.; Cowan, D.; Goldstein, L.J.; Martino, S.; Ingle, J.N.; et al. p53 Expression in Node-Positive Breast Cancer Patients: Results from the Cancer and Leukemia Group B 9344 Trial (159905). Clin. Cancer Res. 2011, 17, 5170–5178. [Google Scholar] [CrossRef] [Green Version]
- Marchetti, P.; Cannita, K.; Ricevuto, E.; De Galitiis, F.; Di Rocco, Z.C.; Tessitore, A.; Bisegna, R.; Porzio, G.; De Rubeis, G.P.; Ventura, T.; et al. Prognostic value of p53 molecular status in high-risk primary breast cancer. Ann. Oncol. 2003, 14, 704–708. [Google Scholar] [CrossRef]
- Loo, L.W.M.; Gao, C.; Shvetsov, Y.B.; Okoro, D.R.; Hernandez, B.Y.; Bargonetti, J. MDM2, MDM2-C, and mutant p53 expression influence breast cancer survival in a multiethnic population. Breast Cancer Res. Treat. 2018, 174, 257–269. [Google Scholar] [CrossRef]
- Kim, J.-Y.; Park, K.; Jung, H.H.; Lee, E.; Cho, E.Y.; Lee, K.H.; Bae, S.Y.; Lee, S.K.; Kim, S.W.; Lee, J.E.; et al. Association between Mutation and Expression of TP53 as a Potential Prognostic Marker of Triple-Negative Breast Cancer. Cancer Res. Treat. 2016, 48, 1338–1350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zwijsen, R.M.; Klompmaker, R.; Wientjens, E.B.; Kristel, P.M.; Van Der Burg, B.; Michalides, R.J. Cyclin D1 triggers autonomous growth of breast cancer cells by governing cell cycle exit. Mol. Cell. Biol. 1996, 16, 2554–2560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lodén, M.; Stighall, M.; Nielsen, N.H.; Roos, G.; Emdin, S.O.; Östlund, H.; Landberg, G. The cyclin D1 high and cyclin E high subgroups of breast cancer: Separate pathways in tumorogenesis based on pattern of genetic aberrations and inactivation of the pRb node. Oncogene 2002, 21, 4680–4690. [Google Scholar] [CrossRef] [Green Version]
- Kilker, R.L.; Planas-Silva, M.D. Cyclin D1 Is Necessary for Tamoxifen-Induced Cell Cycle Progression in Human Breast Cancer Cells. Cancer Res. 2006, 66, 11478–11484. [Google Scholar] [CrossRef] [Green Version]
- Yde, C.W.; Issinger, O.-G. Enhancing cisplatin sensitivity in MCF-7 human breast cancer cells by down-regulation of Bcl-2 and cyclin D1. Int. J. Oncol. 2006, 29, 1397–1404. [Google Scholar] [CrossRef]
- Kilker, R.L.; Hartl, M.W.; Rutherford, T.M.; Planas-Silva, M.D. Cyclin D1 expression is dependent on estrogen receptor function in tamoxifen-resistant breast cancer cells. J. Steroid Biochem. Mol. Biol. 2004, 92, 63–71. [Google Scholar] [CrossRef]
- Joung, Y.H.; Lim, E.J.; Lee, M.Y.; Park, J.-H.; Ye, S.-K.; Park, E.U.; Kim, S.Y.; Zhang, Z.; Lee, K.J.; Park, N.K.; et al. Hypoxia activates the cyclin D1 promoter via the Jak2/STAT5b pathway in breast cancer cells. Exp. Mol. Med. 2005, 37, 353–364. [Google Scholar] [CrossRef] [PubMed]
- Keyomarsi, K.; Pardee, A.B. Redundant cyclin overexpression and gene amplification in breast cancer cells. Proc. Natl. Acad. Sci. USA 1993, 90, 1112–1116. [Google Scholar] [CrossRef] [Green Version]
- Scaltriti, M.; Eichhorn, P.J.; Cortés, J.; Prudkin, L.; Aura, C.; Jiménez, J.; Chandarlapaty, S.; Serra, V.; Prat, A.; Ibrahim, Y.H.; et al. Cyclin E amplification/overexpression is a mechanism of trastuzumab resistance in HER2+breast cancer patients. Proc. Natl. Acad. Sci. USA 2011, 108, 3761–3766. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.-J.; Chang, C.-Y.; Lin, J.-K. Induction of G1 phase arrest in MCF human breast cancer cells by pentagalloylglucose through the down-regulation of CDK4 and CDK2 activities and up-regulation of the CDK inhibitors p27(Kip) and p21(Cip). Biochem. Pharmacol. 2003, 65, 1777–1785. [Google Scholar] [CrossRef]
- Chen, X.; Danes, C.; Lowe, M.; Herliczek, T.W.; Keyomarsi, K. Activation of the Estrogen-Signaling Pathway by p21WAF1/CIP1 in Estrogen Receptor-Negative Breast Cancer Cells. J. Natl. Cancer Inst. 2000, 92, 1403–1413. [Google Scholar] [CrossRef] [Green Version]
- Fritah, A.; Saucier, C.; Mester, J.; Redeuilh, G.; Sabbah, M. p21WAF1/CIP1 Selectively Controls the Transcriptional Activity of Estrogen Receptor α. Mol. Cell. Biol. 2005, 25, 2419–2430. [Google Scholar] [CrossRef] [Green Version]
- Androic, I.; Krämer, A.; Yan, R.; Rödel, F.; Gätje, R.; Kaufmann, M.; Strebhardt, K.; Yuan, J. Targeting cyclin B1 inhibits proliferation and sensitizes breast cancer cells to taxol. BMC Cancer 2008, 8, 391. [Google Scholar] [CrossRef] [Green Version]
- Arima, Y.; Hayashi, N.; Hayashi, H.; Sasaki, M.; Kai, K.; Sugihara, E.; Abe, E.; Yoshida, A.; Mikami, S.; Nakamura, S.; et al. Loss of p16 expression is associated with the stem cell characteristics of surface markers and therapeutic resistance in estrogen receptor-negative breast cancer. Int. J. Cancer 2012, 130, 2568–2579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maria, K.; Vassilis, G.G.; George, Z.R.; Petros, L.; Christos, M.; John, G.; Christos, K. High expression levels of p27 correlate with lymph node status in a subset of advanced invasive breast carcinomas: Relation to E-cadherin alterations, proliferative activity, and ploidy of the tumors. Cancer. 2002, 94, 2454–2465. [Google Scholar] [CrossRef]
- Nahta, R.; Takahashi, T.; Ueno, N.T.; Hung, M.-C.; Esteva, F.J. P27kip1 Down-Regulation Is Associated with Trastuzumab Resistance in Breast Cancer Cells. Cancer Res. 2004, 64, 3981–3986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cariou, S.; Donovan, J.C.H.; Flanagan, W.M.; Milic, A.; Bhattacharya, N.; Slingerland, J.M. Down-regulation of p21WAF1/CIP1 or p27Kip1 abrogates antiestrogen-mediated cell cycle arrest in human breast cancer cells. Proc. Natl. Acad. Sci. USA 2000, 97, 9042–9046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, I.; Shalli, K.; McDonald, S.L.; Moir, E.S.; Hutcheon, A.W.; Heys, S.D.; Schofield, A.C. Reduced expression of p27 is a novel mechanism of docetaxel resistance in breast cancer cells. Breast Cancer Res. 2004, 6, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Dai, X.; Cheng, H.; Bai, Z.; Li, J. Breast Cancer Cell Line Classification and Its Relevance with Breast Tumor Subtyping. J. Cancer 2017, 8, 3131–3141. [Google Scholar] [CrossRef] [Green Version]
- Dickson, C.; Fantl, V.; Gillett, C.; Brookes, S.; Bartek, J.; Smith, R.; Fisher, C.; Barnes, D.; Peters, G. Amplification of chromosome band 11q13 and a role for cyclin D1 in human breast cancer. Cancer Lett. 1995, 90, 43–50. [Google Scholar] [CrossRef]
- Zhang, P. The cell cycle and development: Redundant roles of cell cycle regulators. Curr. Opin. Cell Biol. 1999, 11, 655–662. [Google Scholar] [CrossRef]
- Sutherland, R.L.; Musgrove, E.A. Cyclins and Breast Cancer. J. Mammary Gland. Biol. Neoplasia 2004, 9, 95–104. [Google Scholar] [CrossRef]
- Lundberg, A.; Lindström, L.S.; Li, J.; Harrell, J.C.; Darai-Ramqvist, E.; Sifakis, E.; Foukakis, T.; Perou, C.M.; Czene, K.; Bergh, J.; et al. The long-term prognostic and predictive capacity of cyclin D1 gene amplification in 2305 breast tumours. Breast Cancer Res. 2019, 21, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Z.; Wang, L.; Wang, C.; Ju, X.; Wang, M.; Chen, K.; Loro, E.; Li, Z.; Zhang, Y.; Wu, K.; et al. Cyclin D1 induction of Dicer governs microRNA processing and expression in breast cancer. Nat. Commun. 2013, 4, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, Z.; Yeow, W.-S.; Zou, C.; Wassell, R.; Wang, C.; Pestell, R.G.; Quong, J.N.; Quong, A.A. Cyclin D1/Cyclin-Dependent Kinase 4 Interacts with Filamin A and Affects the Migration and Invasion Potential of Breast Cancer Cells. Cancer Res. 2010, 70, 2105–2114. [Google Scholar] [CrossRef] [Green Version]
- Ravazoula, P.; Batistatou, A.; Aletra, C.; Ladopoulos, J.; Kourounis, G.; Tzigounis, B. Immunohistochemical expression of glucose transporter Glut1 and cyclin D1 in breast carcinomas with negatine lymph nodes. Eur. J. Gynaecol. Oncol. 2003, 24, 544–546. [Google Scholar]
- Ates, C.; Sevinçli, S.; Pohl, T. Electromagnetically induced transparency in strongly interacting Rydberg gases. Phys. Rev. A 2011, 83, 6380–6385. [Google Scholar] [CrossRef] [Green Version]
- Leng, X.; Connell-Crowley, L.; Goodrich, D.; Harper, J. S-Phase entry upon ectopic expression of G1 cyclin-dependent kinases in the absence of retinoblastoma protein phosphorylation. Curr. Biol. 1997, 7, 709–712. [Google Scholar] [CrossRef] [Green Version]
- Hwang, H.C.; Clurman, E.B. Cyclin E in normal and neoplastic cell cycles. Oncogene 2005, 24, 2776–2786. [Google Scholar] [CrossRef] [Green Version]
- Keyomarsi, K.; Tucker, S.L.; Buchholz, T.A.; Callister, M.; Ding, Y.; Hortobagyi, G.N.; Bedrosian, I.; Knickerbocker, C.; Toyofuku, W.; Lowe, M.; et al. Cyclin E and Survival in Patients with Breast Cancer. N. Engl. J. Med. 2002, 347, 1566–1575. [Google Scholar] [CrossRef]
- Dorée, M.; Galas, S. The cyclin-dependent protein kinases and the control of cell division. FASEB J. 1994, 8, 1114–1121. [Google Scholar] [CrossRef]
- Singhai, R.; Patil, A.V.; Patil, V.W.; Gurav, P.D. Triple-negative (ER, PgR, HER-2/neu) breast cancer in Indian women. Breast Cancer Targets Ther. 2011, 3, 9–19. [Google Scholar] [CrossRef] [Green Version]
- Kaldis, P.; Aleem, E. Cell Cycle Sibling Rivalry: Cdc2 Versus Cdk2. Cell Cycle 2005, 4, 1491–1494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hakem, A.; Sasaki, T.; Kozieradzki, I.; Penninger, J.M. The Cyclin-dependent Kinase Cdk2 Regulates Thymocyte Apoptosis. J. Exp. Med. 1999, 189, 957–968. [Google Scholar] [CrossRef] [Green Version]
- Carroll, J.S.; Prall, O.W.J.; Musgrove, E.A.; Sutherland, R.L. A Pure Estrogen Antagonist Inhibits Cyclin E-Cdk2 Activity in MCF-7 Breast Cancer Cells and Induces Accumulation of p130-E2F4 Complexes Characteristic of Quiescence. J. Biol. Chem. 2000, 275, 38221–38229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Poi, M.J.; Tsai, M.-D. Regulatory Mechanisms of Tumor Suppressor P16INK4Aand Their Relevance to Cancer. Biochemistry 2011, 50, 5566–5582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, S.J.; Reddy, E.P. CDK4: A Key Player in the Cell Cycle, Development, and Cancer. Genes Cancer 2012, 3, 658–669. [Google Scholar] [CrossRef] [Green Version]
- Geradts, J.; Wilson, P. High frequency of aberrant p16(INK4A) expression in human breast cancer. Am. J. Pathol. 1996, 149, 15–20. [Google Scholar]
- Gartel, A.L.; Serfas, M.S.; Tyner, A.L. p21--Negative Regulator of the Cell Cycle. Exp. Biol. Med. 1996, 213, 138–149. [Google Scholar] [CrossRef]
- Dai, M.; Al-Odaini, A.A.; Arakelian, A.; Rabbani, A.S.; Ali, S.; Lebrun, J.-J. A novel function for p21Cip1 and acetyltransferase p/CAF as critical transcriptional regulators of TGFβ-mediated breast cancer cell migration and invasion. Breast Cancer Res. 2012, 14, R127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winters, Z.; Hunt, N.; Bradburn, M.; Royds, J.; Turley, H.; Harris, A.; Norbury, C. Subcellular localisation of cyclin B, Cdc2 and p21WAF1/CIP1 in breast cancer: Association with prognosis. Eur. J. Cancer 2001, 37, 2405–2412. [Google Scholar] [CrossRef]
- Li, Y.; Dowbenko, D.; Lasky, L.A. AKT/PKB Phosphorylation of p21Cip/WAF1 Enhances Protein Stability of p21Cip/WAF1 and Promotes Cell Survival. J. Biol. Chem. 2002, 277, 11352–11361. [Google Scholar] [CrossRef] [Green Version]
- Rossig, L.; Jadidi, A.S.; Urbich, C.; Badorff, C.; Zeiher, A.M.; Dimmeler, S. Akt-Dependent Phosphorylation of p21Cip1 Regulates PCNA Binding and Proliferation of Endothelial Cells. Mol. Cell. Biol. 2001, 21, 5644–5657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Wang, R.; Ye, Z.; Wang, Y.; Li, X.; Chen, W.; Zhang, M.; Cai, C. PVT1 affects EMT and cell proliferation and migration via regulating p21 in triple-negative breast cancer cells cultured with mature adipogenic medium. Acta Biochim. Biophys. Sin. 2018, 50, 1211–1218. [Google Scholar] [CrossRef] [Green Version]
- Qian, X.; Hulit, J.; Suyama, K.; Eugenin, A.E.; Belbin, T.J.; Loudig, O.; Smirnova, T.; Zhou, Z.N.; Segall, J.; Locker, J.; et al. p21CIP1 mediates reciprocal switching between proliferation and invasion during metastasis. Oncogene 2012, 32, 2292–2303. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Tenorio, G.; Berglund, F.; Merca, A.E.; Nordenskjöld, B.; Rutqvist, L.E.; Skoog, L.; Stål, O. Cytoplasmic p21WAF1/CIP1 correlates with Akt activation and poor response to tamoxifen in breast cancer. Int. J. Oncol. 2006, 28, 1031–1042. [Google Scholar] [CrossRef]
- Vincent, A.J.; Ren, S.; Harris, L.G.; Devine, D.J.; Samant, R.S.; Fodstad, O.; Shevde, L.A. Cytoplasmic translocation of p21 mediates NUPR1-induced chemoresistance. FEBS Lett. 2012, 586, 3429–3434. [Google Scholar] [CrossRef] [Green Version]
- Sohn, D.; Essmann, F.; Schulze-Osthoff, K.; Jänicke, R.U. p21 Blocks Irradiation-Induced Apoptosis Downstream of Mitochondria by Inhibition of Cyclin-Dependent Kinase–Mediated Caspase-9 Activation. Cancer Res. 2006, 66, 11254–11262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neganova, I.; Lako, M. G1 to S phase cell cycle transition in somatic and embryonic stem cells. J. Anat. 2008, 213, 30–44. [Google Scholar] [CrossRef] [PubMed]
- Fredersdorf, S.; Burns, J.; Milne, A.M.; Packham, G.; Fallis, L.; Gillett, C.E.; Royds, J.A.; Peston, D.; Hall, P.A.; Hanby, A.M.; et al. High level expression of p27kip1 and cyclin D1 in some human breast cancer cells: Inverse correlation between the expression of p27kip1 and degree of malignancy in human breast and colorectal cancers. Proc. Natl. Acad. Sci. USA 1997, 94, 6380–6385. [Google Scholar] [CrossRef] [Green Version]
- Guan, X.; Wang, Y.; Xie, R.; Chen, L.; Bai, J.; Lu, J.; Kuo, M.T. p27Kip1 as a prognostic factor in breast cancer: A systematic review and meta-analysis. J. Cell. Mol. Med. 2009, 14, 944–953. [Google Scholar] [CrossRef] [Green Version]
- Filipits, M.; Rudas, M.; Heinzl, H.; Jakesz, R.; Kubista, E.; Lax, S.; Schippinger, W.; Dietze, O.; Greil, R.; Stiglbauer, W.; et al. Low p27 Expression Predicts Early Relapse and Death in Postmenopausal Hormone Receptor–Positive Breast Cancer Patients Receiving Adjuvant Tamoxifen Therapy. Clin. Cancer Res. 2009, 15, 5888–5894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zilfou, J.T.; Lowe, S.W. Tumor Suppressive Functions of p53. Cold Spring Harb. Perspect. Biol. 2009, 1, a001883. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA A Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef]
- Mathur, P.; Sathishkumar, K.; Chaturvedi, M.; Das, P.; Sudarshan, K.L.; Santhappan, S.; Nallasamy, V.; John, A.; Narasimhan, S.; Roselind, F.S.; et al. Cancer Statistics, 2020: Report From National Cancer Registry Programme, India. JCO Glob. Oncol. 2020, 6, 1063–1075. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Li, Y.; Qin, N.; Wang, F.; Du, J.; Wang, C.; Du, F.; Jiang, T.; Jiang, Y.; Dai, J.; et al. RNA-seq analysis identified hormone-related genes associated with prognosis of triple negative breast cancer. J. Biomed. Res. 2020, 34, 129–138. [Google Scholar] [CrossRef]
- Yu, Z.; Wang, Z.; Yu, X.; Zhang, Z. RNA-Seq-Based Breast Cancer Subtypes Classification Using Machine Learning Approaches. Comput. Intell. Neurosci. 2020, 2020, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Pennock, N.D.; Jindal, S.; Horton, W.; Sun, D.; Narasimhan, J.; Carbone, L.; Fei, S.S.; Searles, R.; Harrington, C.A.; Burchard, J.; et al. RNA-seq from archival FFPE breast cancer samples: Molecular pathway fidelity and novel discovery. BMC Med Genom. 2019, 12, 1–18. [Google Scholar] [CrossRef]
- Zhou, S.; Huang, Y.-E.; Liu, H.; Zhou, X.; Yuan, M.; Hou, F.; Wang, L.; Jiang, W. Single-cell RNA-seq dissects the intratumoral heterogeneity of triple-negative breast cancer based on gene regulatory networks. Mol. Ther.-Nucleic Acids 2021, 23, 682–690. [Google Scholar] [CrossRef]
- Karaayvaz, M.; Cristea, S.; Gillespie, S.M.; Patel, A.P.; Mylvaganam, R.; Luo, C.C.; Specht, M.C.; Bernstein, B.E.; Michor, F.; Ellisen, L.W. Unravelling subclonal heterogeneity and aggressive disease states in TNBC through single-cell RNA-seq. Nat. Commun. 2018, 9, 1–10. [Google Scholar] [CrossRef] [Green Version]
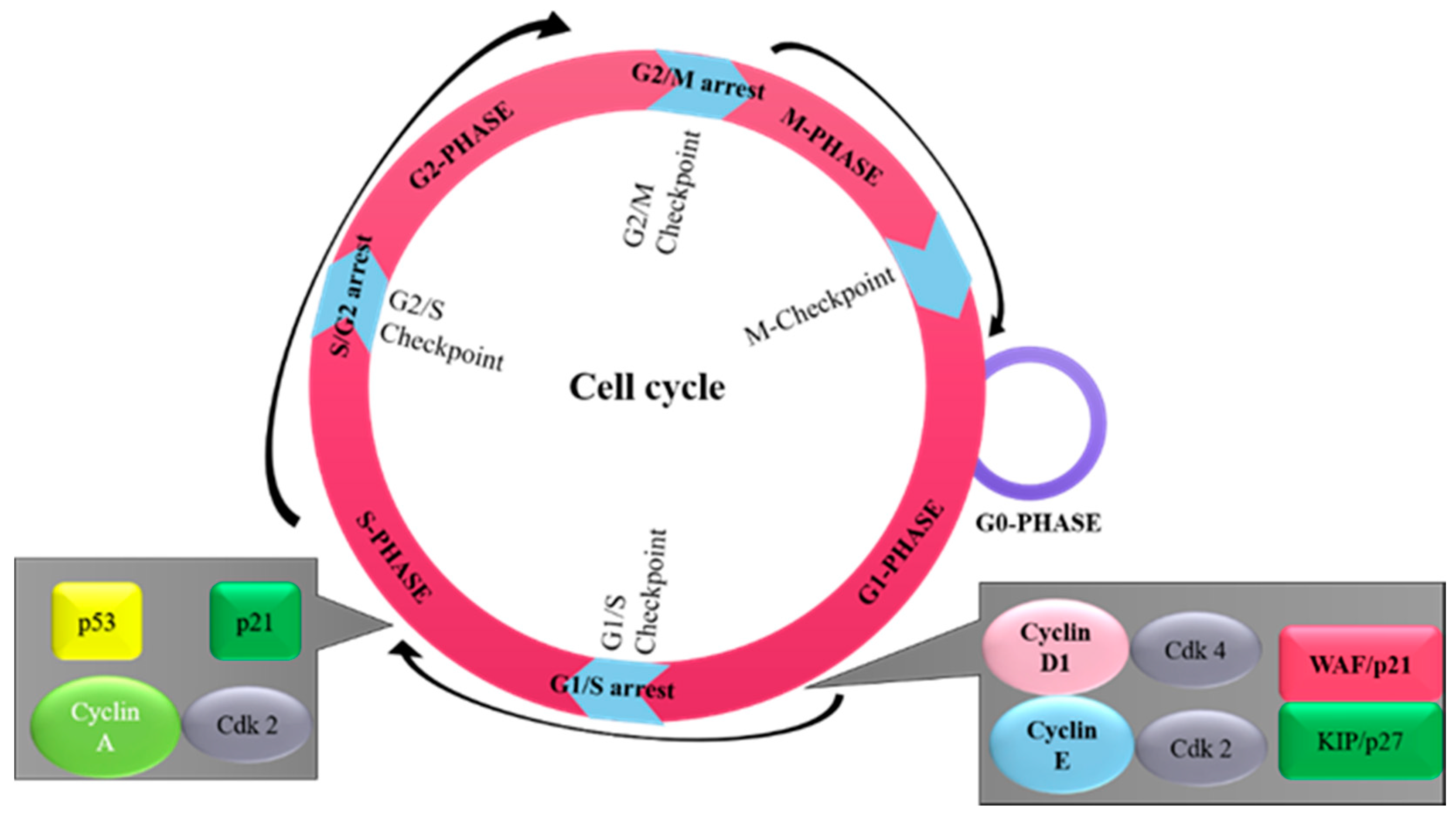


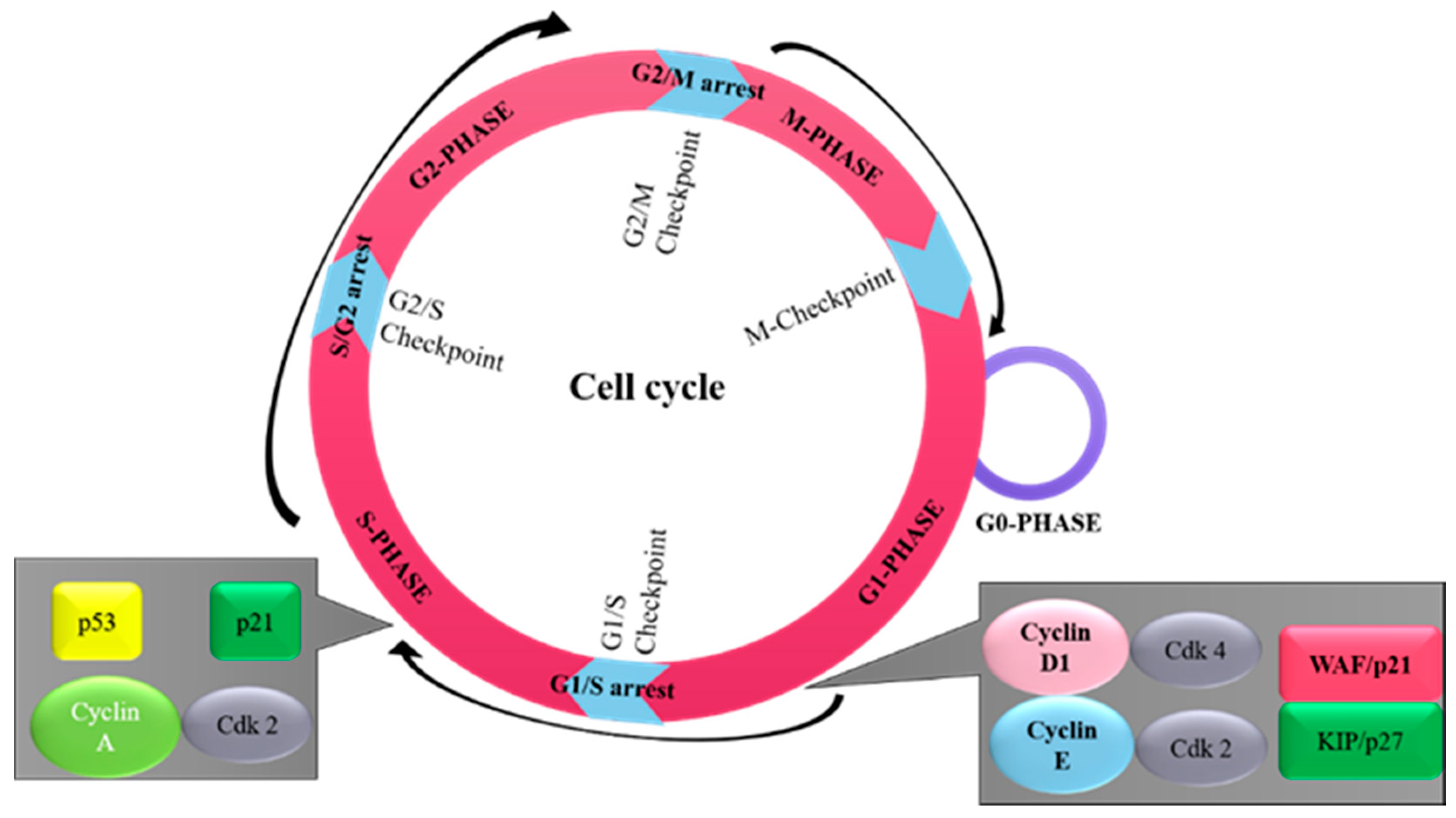{kind=link}
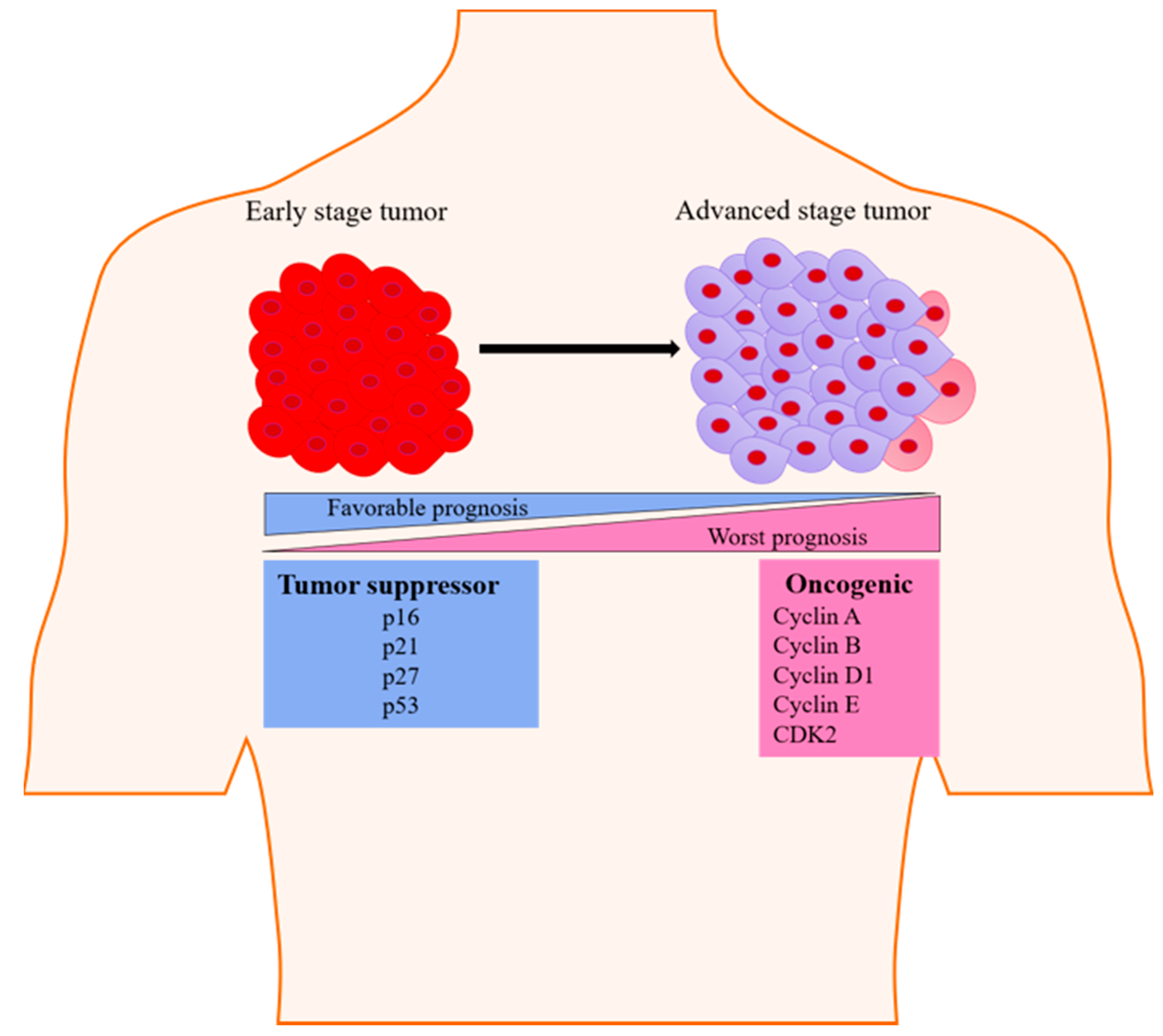{kind=link}
| Marker | Expression | Consequences | Receptor Status | Ref |
|---|---|---|---|---|
| Cyclin D | Overexpression | High risk of replace, Local reoccurrence, Metastasis | ER+/ER- | [45] |
| Overexpression | High tumor grade | ER+/ER-/PR+/PR-/HER2+ | [35] | |
| Overexpression | High proliferation | ER+/ER-/PR+/PR-/TNBC | [38] | |
| Overexpression | High proliferation | ER+/ER-/PR+/PR- | [39] | |
| Overexpression | High proliferation | ER+/ER- | [40] | |
| Overexpression | High proliferation | ER+/ER-/PR+/PR- | [42] | |
| Overexpression | Metastasis | [43] | ||
| Gene amplification Overexpression | High proliferation | [44] | ||
| Overexpression | High proliferation | ER+ | [46] | |
| Gene amplification | High risk for recurrence | ER+/ER- | [47] | |
| Overexpression, Gene amplification | High proliferation | ER+/ER-/PR+/PR- | [48] | |
| Overexpression | High proliferation, Short overall survival, Large tumor size, Lymph node metastasis | ER+/ER-/PR+/PR- | [49] | |
| Overexpression, Gene amplification | High proliferation | ER+/ER-/Basal like | [50] | |
| Overexpression | Reduce relapse-free survival | ER+/PR+ | [51] | |
| Overexpression | High proliferation | ER+/PR+/ER-/PR-/HER2+ | [52] | |
| Overexpression | High proliferation | ER+/PR+/ER-/PR-/HER2+ | [53] | |
| Overexpression | High proliferation | ER+/ER- | [54] | |
| Overexpression | High risk of recurrence | ER+ER-/HER2+/HER2- | [55] | |
| Gene amplification | High proliferation | ER+/ER- | [56] | |
| Gene amplification | Reduce patient survival time, therapy resistance | ER+ | [57] | |
| Overexpression | Poor prognosis | ER+/PR+/ER-/PR-/HER2+/Basal like | [58] | |
| Overexpression | Invasiveness, metastasis | TNBC | [59] | |
| Overexpression | High proliferation | ER+ | [60] | |
| Reduce expression | High tumor grade, Nodal positive status, Invasion | ER+/PR+/ER-/PR-/HER2+/HER2- | [61] | |
| Cyclin A | Overexpression, Gene amplification | Poor prognosis | [62] | |
| Overexpression | Relapse, Shorter disease-free survival | ER+/ER- | [63] | |
| Overexpression | Worst prognosis | ER+/ER- | [64] | |
| Overexpression | Shorter relapse time | ER+/ER- | [65] | |
| Overexpression | Less survival rate, High relapse rate | ER+/PR+/ER-/PR- | [66] | |
| Overexpression | Short distant metastasis-free survival | ER+/PR+/ER-/PR- | [67] | |
| Overexpression | Poor prognosis | ER+/ER- | [68] | |
| Overexpression | High tumor grade, High proliferation index | HER2+/HER2- | [69] | |
| Overexpression | Poor survival | ER+/ER- | [70] | |
| Overexpression | Poor prognosis, Decrease survival rate | ER+/PR+/ER-/PR-/HER2+ | [71] | |
| Cyclin B1 | Overexpression | Decrease survival | ER+/PR+/ER-/PR-/HER2+/HER2- | [72] |
| Overexpression | Reduce overall survival, Disease free survival, Lymphatic invasion | ER+/PR+/ER-/PR-/HER2+/HER2- | [73] | |
| p21(WAF1/Cip1) | Overexpression | High tumor grade, Large tumor size, Positive lymph node, High Ki-67 expression | ER+/PR+/ER-/PR-/HER2+/HER2- | [74] |
| Overexpression | Favorable prognosis | ER+/PR+/ER-/PR- | [75] | |
| Overexpression | Better survival | ER+/PR+/ER-/PR-/HER2+ | [76] | |
| Overexpression | Better survival | ER+/PR+/ER-/PR-/HER2+/HER2- | [77] | |
| Overexpression | Large tumor size, High tumor grade, Lymph node metastasis | ER+/PR+/ER-/PR-/HER2+/HER2- | [78] | |
| p27 (Kip1) | Reduced expression | High tumor grade, Lack of tumor differentiation, Poor prognosis | ER+/PR+/ER-/PR-/HER2+/HER2- | [79] |
| Overexpression | Better prognosis | ER+/ER- | [80] | |
| Overexpression | Favorable prognosis | ER+/PR+/HER2+ | [81] | |
| Reduced expression | Poor prognosis | ER+/PR+/ER-/PR-/HER2+/HER2- | [82] | |
| Overexpression | Long disease-free survival, overall survival | ER+/PR+/ER-/PR- | [83] | |
| Reduced expression | Poor prognosis | ER+/PR+/ER-/PR- | [84] | |
| Reduced expression | Large tumor size, high tumor grade, lymph node metastasis | ER+/ER- | [85] | |
| Overexpression |
Long relapse-free survival,
Overall survival | ER+/PR+ | [86] | |
| Reduced expression | Poor prognosis | ER+/PR+ | [87] | |
| Reduced expression | Increase proliferation | ER+/PR+/ER-/PR-/HER2+/HER2- | [88] | |
| Overexpression | Favorable prognosis | ER+/PR+/ER-/PR-/HER2+/HER2- | [89] | |
| Lower expression | Worst overall survival, Worst disease-free survival | ER+/PR+/ER-/PR- | [90] | |
| Lower expression | Worst overall survival | ER+/PR+/ER-/PR- | [91] | |
| p16 (ink4a) | Overexpression | High proliferation index | ER+/PR+/ER-/PR-/HER2+/HER2- | [92] |
| Overexpression | Favorable prognosis | ER+/PR+/ER-/PR- | [93] | |
| Overexpression | Favorable prognosis | ER-/PR-/HER2- | [94] | |
| Overexpression | Favorable prognosis | ER+/PR+/ER-/PR- | [95] | |
| Overexpression | Disease progression | ER+/PR+/ER-/PR- | [96] | |
| Overexpression | Lung and brain metastasis | ER+/PR+/ER-/PR-/HER2+/HER2- | [97] | |
| Reduced expression | Metastasis | ER+/PR+/ER-/PR-/HER2+/HER2- | [98] | |
| p53 (wild type) | Overexpression | Favorable prognosis | ER+/PR+/ER-/PR-/HER2+/HER2- | [99] |
| Overexpression | Better disease-free survival | ER+/PR+/ER-/PR- | [100] | |
| Overexpression | Favorable prognosis | ER+/PR+/ER-/PR-/HER2+/HER2-/Basal like | [101] | |
| Overexpression | Favorable prognosis | ER+/PR+/ER-/PR- | [102] | |
| Overexpression | Worst prognosis | ER-/PR-/HER2- | [103] | |
| Overexpression | Worst prognosis | ER+/PR+/ER-/PR-/HER2+/HER2- | [104] | |
| Overexpression | Worst prognosis associated with ER expression | ER+/PR+/ER-/PR- | [105] | |
| Overexpression | Worst prognosis associated with HER2+/TNBC subtypes | ER+/PR+/ER-/PR-/HER2+/HER2- | [106] | |
| Overexpression | Worst prognosis, High tumor grade, Lymph vascular invasion, Lymphocyte infiltration | ER+/PR+/ER-/PR- | [107] | |
| Overexpression | Better overall survival in TNBC | ER-/PR-/HER2- | [108] | |
| Overexpression | Correlated with HER2 overexpression, High tumor grade | ER+/PR+/ER-/PR-/HER2+/HER2- | [109] | |
| Overexpression | Worst overall survival, Reoccurrence free survival | ER+/PR+/ER-/PR- | [110] | |
| p53 (Mutant) | Overexpression | Early relapse | ER+/PR+/ER-/PR-/HER2+/HER2- | [111] |
| Overexpression | Poor prognosis | ER+/PR+/ER-/PR- | [112] | |
| Overexpression | Less 5-years reoccurrence free survival | ER+/PR+/ER-/PR- | [113] |
| Marker | Expression | Consequences | Model | Histology | Ref |
|---|---|---|---|---|---|
| Cyclin D | Gene amplification mRNA Overexpression | Proliferation | MCF-7 | ER+ | [33] |
| Overexpression | Malignant transformation | MDA-MB-453 | HER2+ | [41] | |
| Overexpression | Increase proliferation | T-47D, MCF-7 | ER+/HER2+ | [34] | |
| Overexpression | Increase proliferation | MCF-7 | ER+ | [114] | |
| Overexpression | Increase proliferation | MCF-7, T-47D, MDA-MB-468, BT-549 | ER+/ER-/PR-/HER2- | [115] | |
| Overexpression | Increase proliferation | HBL-100, MDA-MB-23l, T-47D, MCF-7, MDA-MB-134, HMEC-184 | ER+/PR+/ER-/PR-/HER2-/HER2+ | [52] | |
| Overexpression | Increase proliferation | MCF-7T, | Tamoxifen-resistant | [116] | |
| Downregulation | Cell death, Growth arrest | MCF-7, MDA-MB-231, MDA-MB-435, HCC-1937, CAL-148 | ER+/ER-/PR-/HER2- | [117] | |
| Overexpression | Invasiveness, metastasis | MDA-MB-231 | ER-/PR-/HER2- | [59] | |
| Overexpression | Increase proliferation | MCF-7 | ER+ | [118] | |
| Overexpression | Increase proliferation | MCF-7 | ER+ | [60] | |
| Overexpression | Increase proliferation | MCF-7 | ER+ | [119] | |
| Overexpression | Increase proliferation | ZR75-1-2, ZR-75-1, MDAMB-157, MDA-MB-231, MDA-MB-436, T-47D, BT-20, HBL-100, Hs578T, SK-BR3 | ER+/ER-/PR+/PR-/HER2-/HER2+ | [120] | |
| Cyclin E | Overexpression, Gene amplification | Increase proliferation | BT-474, BT-474R | HER2+ | [121] |
| Downregulation | Suppression of cell cycle progression | MCF-7 | ER+ | [122] | |
| Cyclin B | Overexpression | Associated with ER+ status | MCF-7, MDA-MB-231, MDA-MB-436, Hs578 T | ER+/PR+/ER-/PR-/HER2-/HER2+ | [123,124] |
| Downregulation | Apoptosis, Anti-proliferation | MCF-7, BT-474, SK-BR-3, MDA-MB-231 | ER+/ER-/PR+/PR-/HER2-/HER2+ | [125] | |
| p16(WAF1/Cip1) | Reduced expression | Therapy resistance | HCC-1428, T-47D, MCF-7, MDA-MB-436, BT-549, MDA-MB-157, MDA-MB-231, MDA-MB-435S, Hs578T, HCC-1937, BT-20, SK-BR-3 | ER+/PR+/ER-/PR_/HER2-/HER2+ | [126] |
| Overexpression | Good survival | MCF-7, BT-549, MDA-MB-134, MDA-MB-157, MDA-MB-231, MDA-MB-453, MDA-MB-468, ZR-75-1, BT-20, SK-BR-3, T-47D | ER+/PR+/ER-/PR-/HER2-/HER2+ | [76] | |
| p21(WAF1/Cip1) | Overexpression | High proliferation rate |
ZR75-1,
ZR75-30, MCF-7, MDA-MB-453, T-47D, Cal51, SK-BR-5, SK-BR-7, CAMA-1, BT-20 | ER+/PR+/ER-/PR-/HER2+ | [127] |
| Reduced expression | Trastuzumab resistance | SK-BR-3 | HER2+ | [128] | |
| Overexpression | Cell cycle arrest | MCF-7 | ER+ | [129] | |
| Reduced expression | Acquired resistance to docetaxel | MCF-7, MDA-MB-231 | ER+/ER-/PR-/HER2- | [130] | |
| Overexpression | Associated with ER+ status | MCF-7, MDA-MB-231, MDA-MB-436, Hs578T | ER+/PR+/ER-/PR-/HER2-/HER2+ | [123,124] |
| Cell Line | ER | PR | HER2/neu+ | Subtype | BRAC1 Mutation | p53 Mutation | Tumor |
|---|---|---|---|---|---|---|---|
| CAMA-1 | + | +/- | - | LA | WT | MU | AC |
| HCC1428 | + | + | - | LA | ND | ND | AC |
| MCF-7 | + | - | - | LA | ND | WT | IDC |
| MDA-MB-134 | + | - | - | LA | ND | MU | IDC |
| T-47D | + | + | - | LA | WT | MU | IDC |
| ZR75-1 | + | +/- | - | LA | WT | WT | IDC |
| BT-474 | + | + | + | LB | WT | MU | IDC |
| ZR75-30 | + | - | + | LB | WT | WT | IDC |
| MDA-MB-453 | - | - | + | Her2+ | WT | MU | AC |
| SK-BR-3 | - | - | + | Her2+ | WT | MU | AC |
| SK-BR-5 | - | - | + | Her2+ | WT | MU | AC |
| BT-20 | - | - | - | TNBC | WT | MU | IDC |
| BT-549 | - | - | - | TNBC | WT | MU | IDC |
| CAL-51 | - | - | - | TNBC | WT | MU | AC |
| CAL-148 | - | - | - | TNBC | WT | MU | AC |
| HCC1937 | - | - | - | TNBC | MU | MU | DC |
| Hs578T | - | - | - | TNBC | WT | MU | IDC |
| MDA-MB-157 | - | - | - | TNBC | WT | MU | MC |
| MDA-MB-231 | - | - | - | TNBC | WT | MU | AC |
| MDA-MB-435 | - | - | - | TNBC | WT | MU | AC |
| MDA-MB-436 | - | - | - | TNBC | MU | MU | AC |
| MDA-MB-468 | - | - | - | TNBC | WT | MU | AC |
| SK-BR-7 | - | - | - | TNBC | WT | WT | AC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kashyap, D.; Garg, V.K.; Sandberg, E.N.; Goel, N.; Bishayee, A. Oncogenic and Tumor Suppressive Components of the Cell Cycle in Breast Cancer Progression and Prognosis. Pharmaceutics 2021, 13, 569. https://doi.org/10.3390/pharmaceutics13040569
Kashyap D, Garg VK, Sandberg EN, Goel N, Bishayee A. Oncogenic and Tumor Suppressive Components of the Cell Cycle in Breast Cancer Progression and Prognosis. Pharmaceutics. 2021; 13(4):569. https://doi.org/10.3390/pharmaceutics13040569
Chicago/Turabian StyleKashyap, Dharambir, Vivek Kumar Garg, Elise N. Sandberg, Neelam Goel, and Anupam Bishayee. 2021. "Oncogenic and Tumor Suppressive Components of the Cell Cycle in Breast Cancer Progression and Prognosis" Pharmaceutics 13, no. 4: 569. https://doi.org/10.3390/pharmaceutics13040569
APA StyleKashyap, D., Garg, V. K., Sandberg, E. N., Goel, N., & Bishayee, A. (2021). Oncogenic and Tumor Suppressive Components of the Cell Cycle in Breast Cancer Progression and Prognosis. Pharmaceutics, 13(4), 569. https://doi.org/10.3390/pharmaceutics13040569