


A β-Cyclodextrin-Based Nanoparticle with Very High Transfection Efficiency Unveils siRNA-Activated TLR3 Responses in Human Prostate Cancer Cells
 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
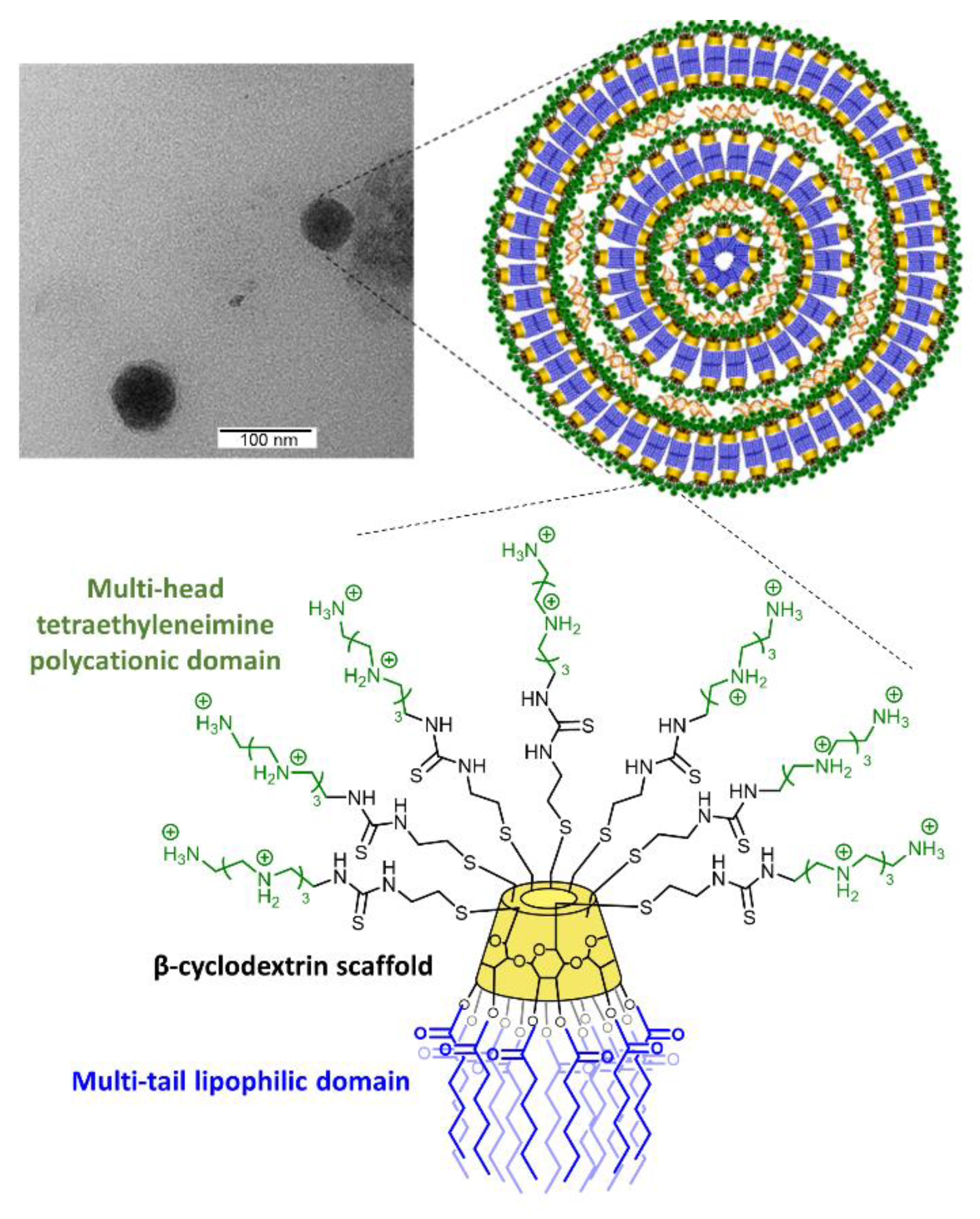
2.1. Molecular Vector Synthesis
2.2. Dynamic Light Scattering (DLS)
2.3. Transmission Electron Microscopy
2.4. Gel Retardation Assay
2.5. Small Interfering RNA Protection against RNAses
2.6. Cell Culture
2.7. Toxicity Assay
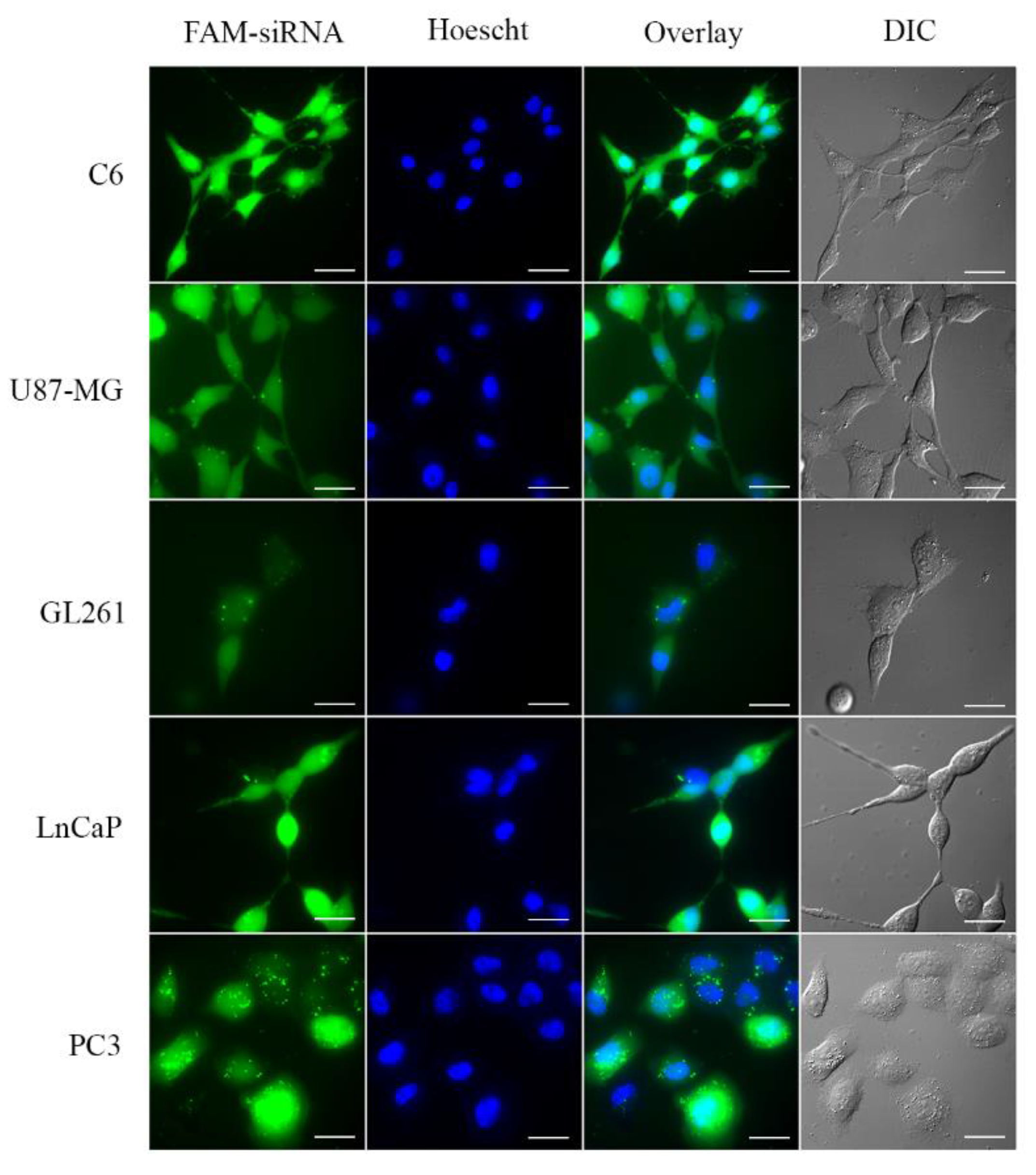
2.8. Small Interfering RNA Cellular Uptake
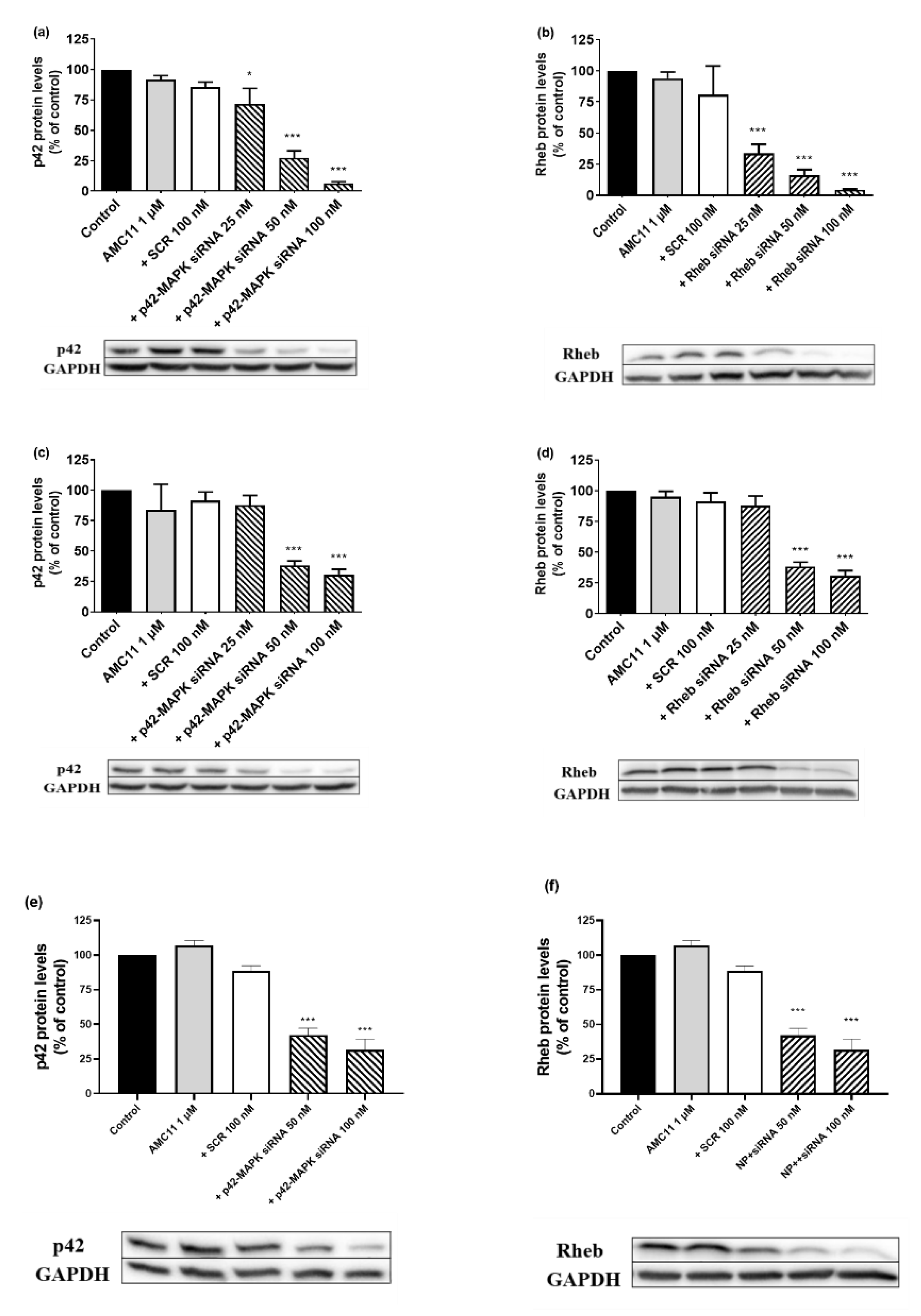
2.9. Small Interfering RNA Transfection and Western Blot Analysis
2.10. Isolation of Nuclear and Cytosolic Fractions
2.11. Caspase Activity
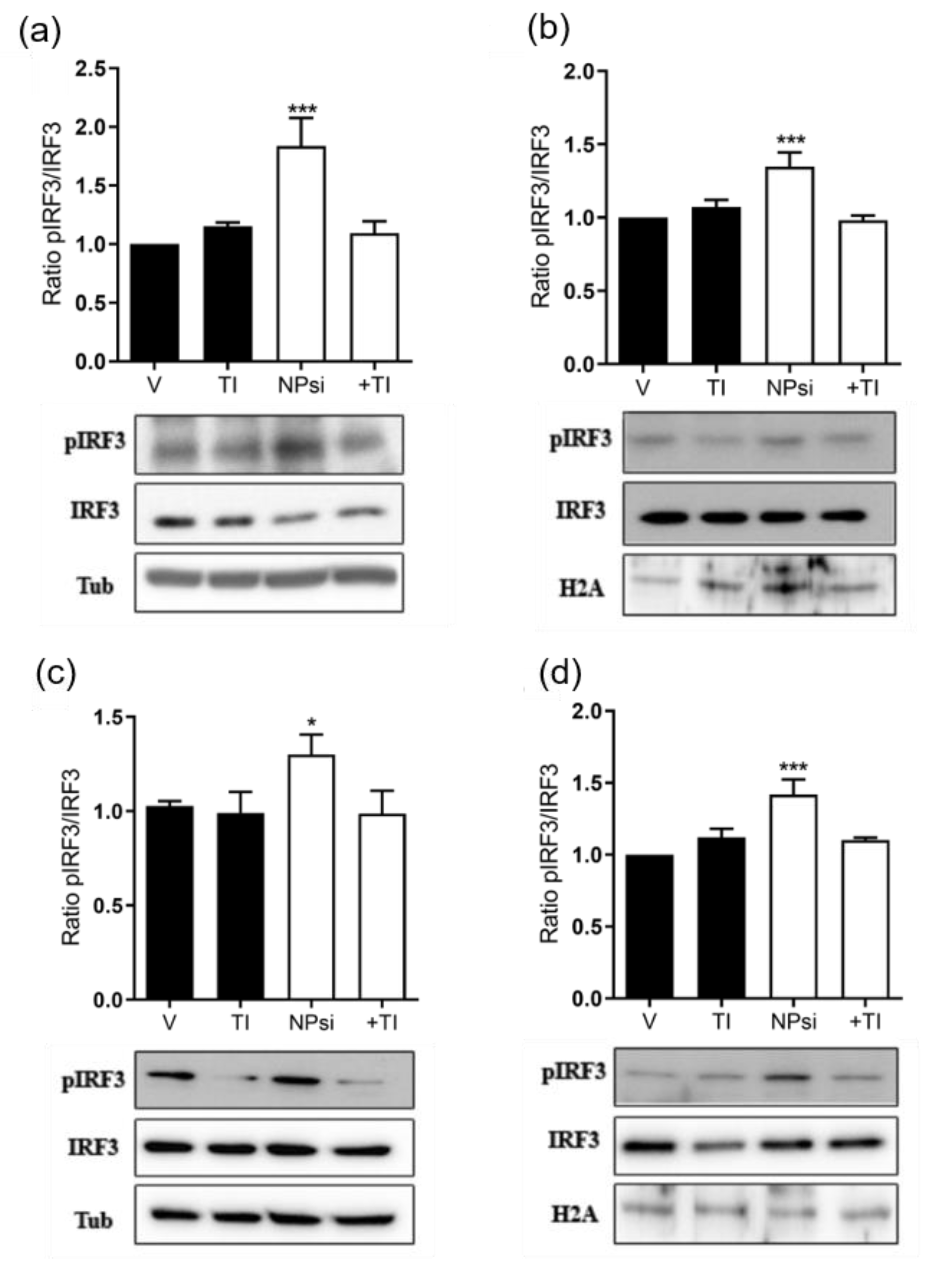
2.12. Interferon β-1a Quantification
2.13. Statistical Analysis
3. Results
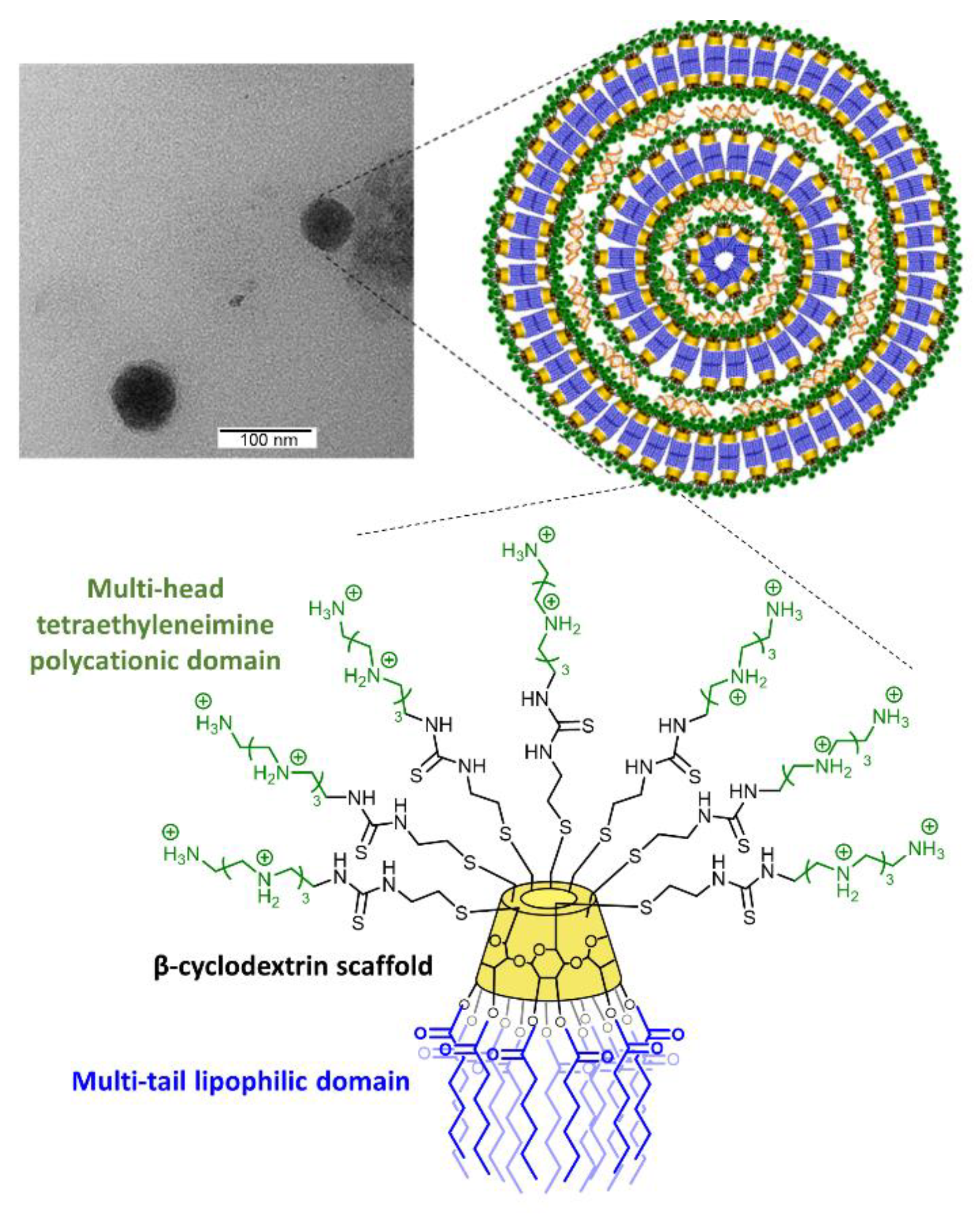
3.1. Synthesis, Hydrodynamic Diameter, and ζ-Potential of AMC11
3.2. AMC11–Small Interfering RNA Interaction
3.3. Cellular Toxicity
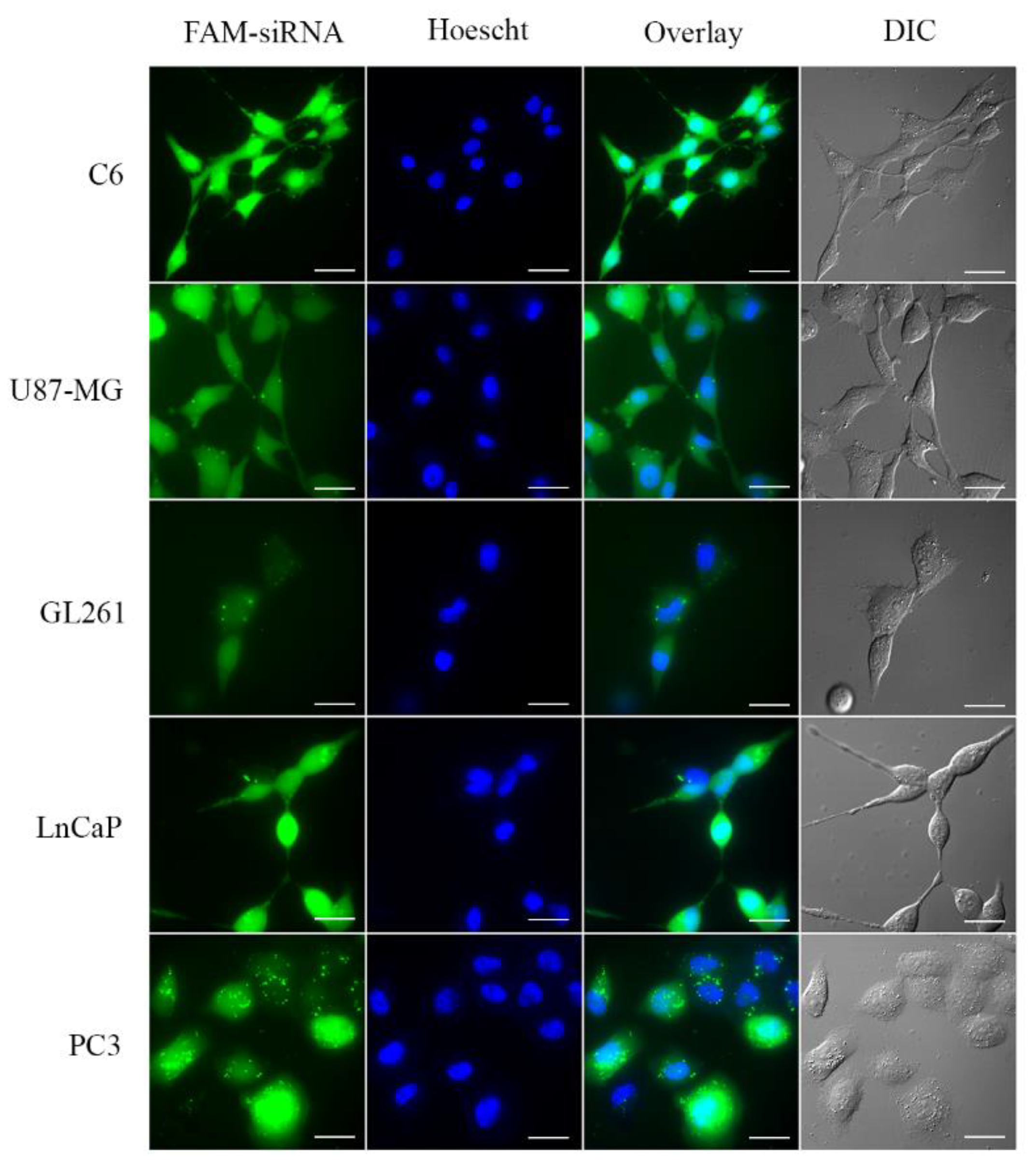
3.4. Small Interfering RNA Transport into the Cell
3.5. Transfection Efficiency of AMC11 βCD–Nucleic Acid Nanocomplexes
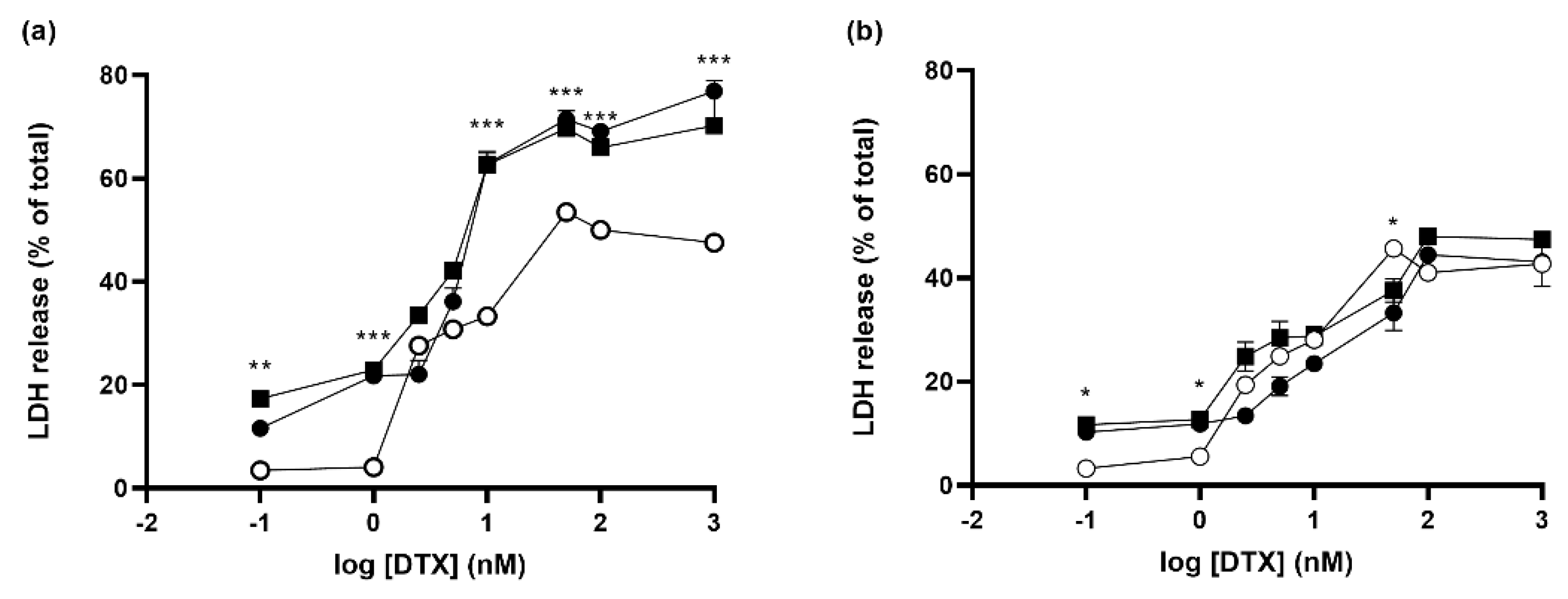
3.6. Effect of Knocking down Rheb and p42-MAPK on Docetaxel-Induced Cell Death in Prostate Cancer Cell Lines
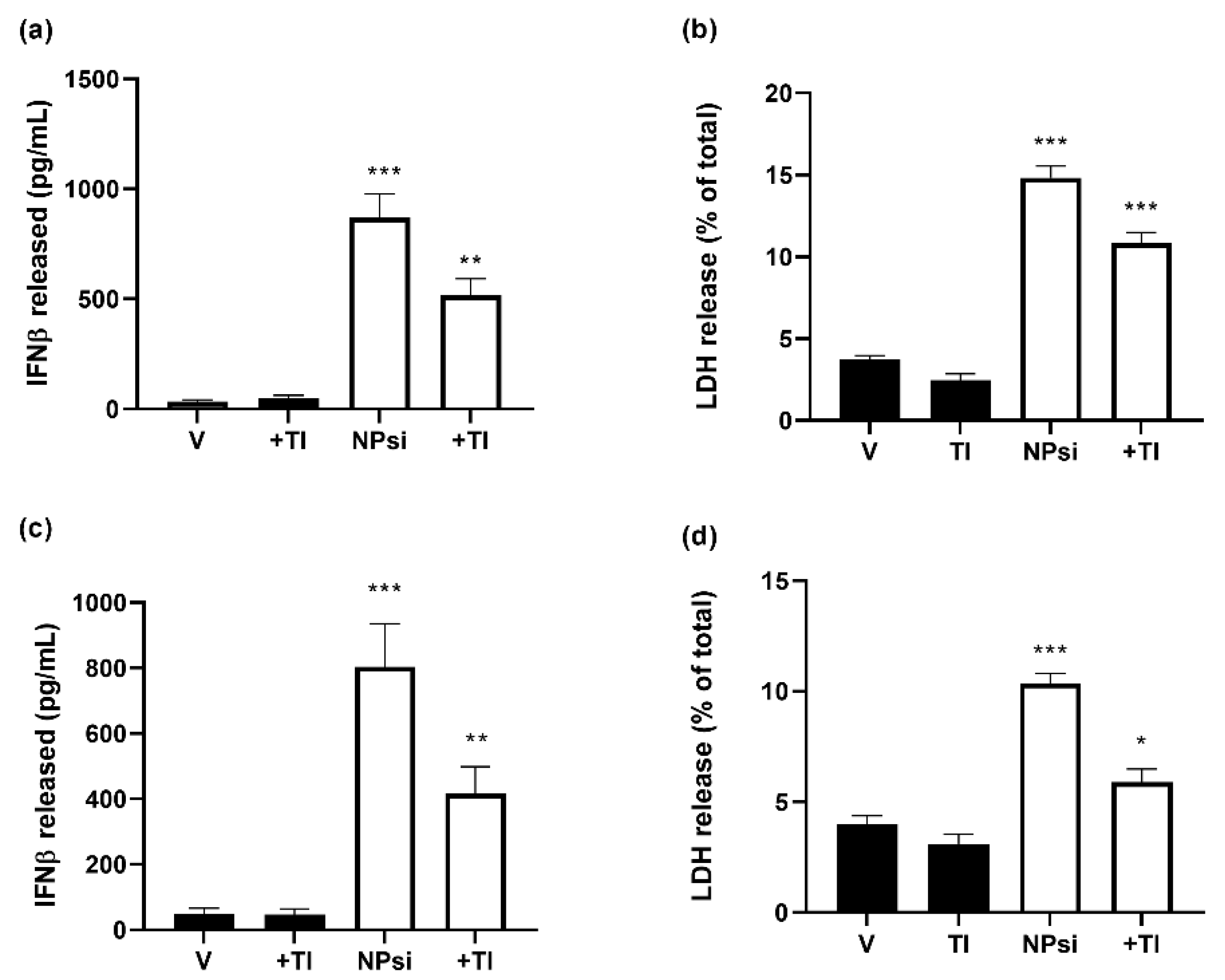
3.7. Molecular Mechanism of siRNA-Induced Toxicity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aagaard, L.; Rossi, J.J. RNAi therapeutics: Principles, prospects and challenges. Adv. Drug Deliv. Rev. 2007, 59, 75–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carthew, R.W.; Sontheimer, E.J. Origins and Mechanisms of miRNAs and siRNAs. Cell 2009, 136, 642–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anthiya, S.; Griveau, A.; Loussouarn, C.; Baril, P.; Garnett, M.; Issartel, J.P.; Garcion, E. MicroRNA-Based Drugs for Brain Tumors. Trends Cancer 2018, 4, 222–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahmoodi Chalbatani, G.; Dana, H.; Gharagouzloo, E.; Grijalvo, S.; Eritja, R.; Logsdon, C.D.; Memari, F.; Miri, S.R.; Rad, M.R.; Marmari, V. Small interfering RNAs (siRNAs) in cancer therapy: A nano-based approach. Int. J. Nanomed. 2019, 14, 3111–3128. [Google Scholar] [CrossRef] [Green Version]
- Posadas, I.; Guerra, F.J.; Cena, V. Nonviral vectors for the delivery of small interfering RNAs to the CNS. Nanomedicine 2010, 5, 1219–1236. [Google Scholar] [CrossRef]
- Adams, D.; Gonzalez-Duarte, A.; O’Riordan, W.D.; Yang, C.C.; Ueda, M.; Kristen, A.V.; Tournev, I.; Schmidt, H.H.; Coelho, T.; Berk, J.L.; et al. Patisiran, an RNAi Therapeutic, for Hereditary Transthyretin Amyloidosis. N. Engl. J. Med. 2018, 379, 11–21. [Google Scholar] [CrossRef]
- Balwani, M.; Sardh, E.; Ventura, P.; Peiro, P.A.; Rees, D.C.; Stolzel, U.; Bissell, D.M.; Bonkovsky, H.L.; Windyga, J.; Anderson, K.E.; et al. Phase 3 Trial of RNAi Therapeutic Givosiran for Acute Intermittent Porphyria. N. Engl. J. Med. 2020, 382, 2289–2301. [Google Scholar] [CrossRef]
- Khurana, A.; Allawadhi, P.; Khurana, I.; Allwadhi, S.; Weiskirchen, R.; Banothu, A.K.; Chhabra, D.; Joshi, K.; Bharani, K.K. Role of nanotechnology behind the success of mRNA vaccines for COVID-19. Nano Today 2021, 38, 101142. [Google Scholar] [CrossRef]
- Chung, Y.H.; Beiss, V.; Fiering, S.N.; Steinmetz, N.F. COVID-19 Vaccine Frontrunners and Their Nanotechnology Design. ACS Nano 2020, 14, 12522–12537. [Google Scholar] [CrossRef]
- Mansoori, B.; Mohammadi, A.; Shir Jang, S.; Baradaran, B. Mechanisms of immune system activation in mammalians by small interfering RNA (siRNA). Artif. Cells Nanomed. Biotechnol. 2016, 44, 1589–1596. [Google Scholar] [CrossRef]
- Meng, Z.; Lu, M. RNA Interference-Induced Innate Immunity, Off-Target Effect, or Immune Adjuvant? Front. Immunol. 2017, 8, 331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geng, W.C.; Huang, Q.; Xu, Z.; Wang, R.; Guo, D.S. Gene delivery based on macrocyclic amphiphiles. Theranostics 2019, 9, 3094–3106. [Google Scholar] [CrossRef] [PubMed]
- Blanco, J.L.J.; Benito, J.M.; Mellet, C.O.; Fernández, J.M.G. Molecular nanoparticle-based gene delivery systems. J. Drug Deliv. Sci. Technol. 2017, 42, 18–37. [Google Scholar] [CrossRef]
- Carbajo-Gordillo, A.I.; López-Fernández, J.; Benito, J.M.; Jiménez Blanco, J.L.; Santana-Armas, M.L.; Marcelo, G.; Di Giorgio, C.; Przybylski, C.; Ortiz Mellet, C.; Tros de Ilarduya, C.; et al. Enhanced Gene Delivery Triggered by Dual pH/Redox Responsive Host-Guest Dimerization of Cyclooligosaccharide Star Polycations. Macromol. Rapid Commun. 2022, 43, 2200145. [Google Scholar] [CrossRef]
- Carbajo-Gordillo, A.I.; Gonzalez-Cuesta, M.; Jimenez Blanco, J.L.; Benito, J.M.; Santana-Armas, M.L.; Carmona, T.; Di Giorgio, C.; Przybylski, C.; Ortiz Mellet, C.; Tros de Ilarduya, C.; et al. Trifaceted Mickey Mouse Amphiphiles for Programmable Self-Assembly, DNA Complexation and Organ-Selective Gene Delivery. Chemistry 2021, 27, 9429–9438. [Google Scholar] [CrossRef]
- Carbajo-Gordillo, A.I.; Jimenez Blanco, J.L.; Benito, J.M.; Lana, H.; Marcelo, G.; Di Giorgio, C.; Przybylski, C.; Hinou, H.; Ceña, V.; Ortiz Mellet, C.; et al. Click Synthesis of Size- and Shape-Tunable Star Polymers with Functional Macrocyclic Cores for Synergistic DNA Complexation and Delivery. Biomacromolecules 2020, 21, 5173–5188. [Google Scholar] [CrossRef]
- Rivero-Barbarroja, G.; Benito, J.M.; Mellet, C.O.; Fernández, J.M.G. Cyclodextrin-Based Functional Glyconanomaterials. Nanomaterials 2020, 10, 2517. [Google Scholar] [CrossRef]
- Gallego-Yerga, L.; Benito, J.M.; Blanco-Fernández, L.; Martínez-Negro, M.; Vélaz, I.; Aicart, E.; Junquera, E.; Mellet, C.O.; de Ilarduya, C.T.; Fernández, J.M.G. Plasmid-Templated Control of DNA–Cyclodextrin Nanoparticle Morphology through Molecular Vector Design for Effective Gene Delivery. Chem. Eur. J. 2018, 24, 3825–3835. [Google Scholar] [CrossRef]
- Remaut, K.; De Clercq, E.; Andries, O.; Rombouts, K.; Van Gils, M.; Cicchelero, L.; Vandenbussche, I.; Van Praet, S.; Benito, J.M.; Fernandez, J.M.; et al. Aerosolized Non-viral Nucleic Acid Delivery in the Vaginal Tract of Pigs. Pharm. Res. 2016, 33, 384–394. [Google Scholar] [CrossRef]
- Aggarwal, R.; Huang, J.; Alumkal, J.J.; Zhang, L.; Feng, F.Y.; Thomas, G.V.; Weinstein, A.S.; Friedl, V.; Zhang, C.; Witte, O.N.; et al. Clinical and Genomic Characterization of Treatment-Emergent Small-Cell Neuroendocrine Prostate Cancer: A Multi-institutional Prospective Study. J. Clin. Oncol. 2018, 36, 2492–2503. [Google Scholar] [CrossRef] [Green Version]
- Ryan, C.J.; Smith, M.R.; de Bono, J.S.; Molina, A.; Logothetis, C.J.; de Souza, P.; Fizazi, K.; Mainwaring, P.; Piulats, J.M.; Ng, S.; et al. Abiraterone in metastatic prostate cancer without previous chemotherapy. N. Engl. J. Med. 2013, 368, 138–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sweeney, C.J.; Chen, Y.H.; Carducci, M.; Liu, G.; Jarrard, D.F.; Eisenberger, M.; Wong, Y.N.; Hahn, N.; Kohli, M.; Cooney, M.M.; et al. Chemohormonal Therapy in Metastatic Hormone-Sensitive Prostate Cancer. N. Engl. J. Med. 2015, 373, 737–746. [Google Scholar] [CrossRef] [PubMed]
- Jativa, P.; Ceña, V. Use of nanoparticles for glioblastoma treatment: A new approach. Nanomedicine 2017, 12, 2533–2554. [Google Scholar] [CrossRef] [PubMed]
- Ronkina, N.; Gaestel, M. MAPK-Activated Protein Kinases: Servant or Partner? Annu. Rev. Biochem. 2022, 91, 505–540. [Google Scholar] [CrossRef]
- Nickols, N.G.; Nazarian, R.; Zhao, S.G.; Tan, V.; Uzunangelov, V.; Xia, Z.; Baertsch, R.; Neeman, E.; Gao, A.C.; Thomas, G.V.; et al. MEK-ERK signaling is a therapeutic target in metastatic castration resistant prostate cancer. Prostate Cancer Prostatic Dis. 2019, 22, 531–538. [Google Scholar] [CrossRef]
- Roudsari, N.M.; Lashgari, N.A.; Momtaz, S.; Abaft, S.; Jamali, F.; Safaiepour, P.; Narimisa, K.; Jackson, G.; Bishayee, A.; Rezaei, N.; et al. Inhibitors of the PI3K/Akt/mTOR Pathway in Prostate Cancer Chemoprevention and Intervention. Pharmaceutics 2021, 13, 1195. [Google Scholar] [CrossRef]
- Gupta, S.; Kumar, M.; Chaudhuri, S.; Kumar, A. The non-canonical nuclear functions of key players of the PI3K-AKT-MTOR pathway. J. Cell. Physiol. 2022, 237, 3181–3204. [Google Scholar] [CrossRef]
- Ashrafizadeh, M.; Hushmandi, K.; Rahmani Moghadam, E.; Zarrin, V.; Hosseinzadeh Kashani, S.; Bokaie, S.; Najafi, M.; Tavakol, S.; Mohammadinejad, R.; Nabavi, N.; et al. Progress in Delivery of siRNA-Based Therapeutics Employing Nano-Vehicles for Treatment of Prostate Cancer. Bioengineering 2020, 7, 91. [Google Scholar] [CrossRef]
- Martinez, A.; Bienvenu, C.; Jimenez Blanco, J.L.; Vierling, P.; Mellet, C.O.; Garcia Fernandez, J.M.; Di Giorgio, C. Amphiphilic oligoethyleneimine-beta-cyclodextrin “click” clusters for enhanced DNA delivery. J. Org. Chem. 2013, 78, 8143–8148. [Google Scholar] [CrossRef]
- Gallego-Yerga, L.; de la Torre, C.; Sansone, F.; Casnati, A.; Mellet, C.O.; Garcia Fernandez, J.M.; Ceña, V. Synthesis, self-assembly and anticancer drug encapsulation and delivery properties of cyclodextrin-based giant amphiphiles. Carbohydr. Polym. 2021, 252, 117135. [Google Scholar] [CrossRef]
- Rodrigo, A.C.; Rivilla, I.; Perez-Martinez, F.C.; Monteagudo, S.; Ocana, V.; Guerra, J.; Garcia-Martinez, J.C.; Merino, S.; Sanchez-Verdu, P.; Ceña, V.; et al. Efficient, Non-Toxic Hybrid PPV-PAMAM Dendrimer as a Gene Carrier for Neuronal Cells. Biomacromolecules 2011, 12, 1205–1213. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-Carrion, M.D.; Perez-Martinez, F.C.; Merino, S.; Sanchez-Verdu, P.; Martinez-Hernandez, J.; Lujan, R.; Ceña, V. Dendrimer-mediated siRNA delivery knocks down Beclin 1 and potentiates NMDA-mediated toxicity in rat cortical neurons. J. Neurochem. 2012, 120, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Perez-Carrion, M.D.; Ceña, V. Knocking down HMGB1 using dendrimer-delivered siRNA unveils its key role in NMDA-induced autophagy in rat cortical neurons. Pharm. Res. 2013, 30, 2584–2595. [Google Scholar] [CrossRef] [PubMed]
- Manzanares, D.; Perez-Carrion, M.D.; Jimenez Blanco, J.L.; Ortiz Mellet, C.; Garcia Fernandez, J.M.; Ceña, V. Cyclodextrin-Based Nanostructure Efficiently Delivers siRNA to Glioblastoma Cells Preferentially via Macropinocytosis. Int. J. Mol. Sci. 2020, 21, 9306. [Google Scholar] [CrossRef]
- Lopez-Hernandez, B.; Ceña, V.; Posadas, I. The endoplasmic reticulum stress and the HIF-1 signalling pathways are involved in the neuronal damage caused by chemical hypoxia. Br. J. Pharmacol. 2015, 172, 2838–2851. [Google Scholar] [CrossRef] [Green Version]
- Mellet, C.O.; Fernández, J.G.; Benito, J.M. Cyclodextrins for Pharmaceutical and Biomedical Applications. In Supramolecular Systems in Biomedical Fields; Schneider, H.-J., Ed.; RSC Publishing: Cambridge, UK, 2013; pp. 94–139. [Google Scholar]
- Manzanares, D.; Ceña, V. Endocytosis: The Nanoparticle and Submicron Nanocompounds Gateway into the Cell. Pharmaceutics 2020, 12, 371. [Google Scholar] [CrossRef] [Green Version]
- Rana, Z.; Diermeier, S.; Walsh, F.P.; Hanif, M.; Hartinger, C.G.; Rosengren, R.J. Anti-Proliferative, Anti-Angiogenic and Safety Profiles of Novel HDAC Inhibitors for the Treatment of Metastatic Castration-Resistant Prostate Cancer. Pharmaceuticals 2021, 14, 1020. [Google Scholar] [CrossRef]
- Zainol, M.I.B.; Kawasaki, T.; Monwan, W.; Murase, M.; Sueyoshi, T.; Kawai, T. Innate immune responses through Toll-like receptor 3 require human-antigen-R-mediated Atp6v0d2 mRNA stabilization. Sci. Rep. 2019, 9, 20406. [Google Scholar] [CrossRef] [Green Version]
- Smith, E.S.; Whitty, E.; Yoo, B.; Moore, A.; Sempere, L.F.; Medarova, Z. Clinical Applications of Short Non-Coding RNA-Based Therapies in the Era of Precision Medicine. Cancers 2022, 14, 1588. [Google Scholar] [CrossRef]
- Sanati, M.; Afshari, A.R.; Kesharwani, P.; Sukhorukov, V.N.; Sahebkar, A. Recent trends in the application of nanoparticles in cancer therapy: The involvement of oxidative stress. J. Control. Release 2022, 348, 287–304. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, T.; Liu, F.F.; Lu, B.; Lip, H.; Park, E.; Alradwan, I.; Liu, J.F.; He, C.; Zetrini, A.; Zhang, T.; et al. Advances in Nanomedicine Design: Multidisciplinary Strategies for Unmet Medical Needs. Mol. Pharm. 2022, 19, 1722–1765. [Google Scholar] [CrossRef] [PubMed]
- Ai, X.; Wang, S.; Duan, Y.; Zhang, Q.; Chen, M.S.; Gao, W.; Zhang, L. Emerging Approaches to Functionalizing Cell Membrane-Coated Nanoparticles. Biochemistry 2021, 60, 941–955. [Google Scholar] [CrossRef] [PubMed]
- Bolukbas, D.A.; Datz, S.; Meyer-Schwickerath, C.; Morrone, C.; Doryab, A.; Gossl, D.; Vreka, M.; Yang, L.; Argyo, C.; van Rijt, S.H.; et al. Organ-restricted vascular delivery of nanoparticles for lung cancer therapy. Adv. Ther. 2020, 3, 2000017. [Google Scholar] [CrossRef]
- Thomas, C.E.; Ehrhardt, A.; Kay, M.A. Progress and problems with the use of viral vectors for gene therapy. Nat. Rev. Genet. 2003, 4, 346–358. [Google Scholar] [CrossRef]
- Manzanares, D.; Araya-Duran, I.; Gallego-Yerga, L.; Jativa, P.; Marquez-Miranda, V.; Canan, J.; Jimenez Blanco, J.L.; Mellet, C.O.; Gonzalez-Nilo, F.D.; Garcia Fernandez, J.M.; et al. Molecular determinants for cyclo-oligosaccharide-based nanoparticle-mediated effective siRNA transfection. Nanomedicine 2017, 12, 1607–1621. [Google Scholar] [CrossRef]
- Guo, Y.J.; Pan, W.W.; Liu, S.B.; Shen, Z.F.; Xu, Y.; Hu, L.L. ERK/MAPK signalling pathway and tumorigenesis. Exp. Ther. Med. 2020, 19, 1997–2007. [Google Scholar] [CrossRef] [Green Version]
- Braicu, C.; Buse, M.; Busuioc, C.; Drula, R.; Gulei, D.; Raduly, L.; Rusu, A.; Irimie, A.; Atanasov, A.G.; Slaby, O.; et al. A Comprehensive Review on MAPK: A Promising Therapeutic Target in Cancer. Cancers 2019, 11, 1618. [Google Scholar] [CrossRef] [Green Version]
- Munkley, J.; Rajan, P.; Lafferty, N.P.; Dalgliesh, C.; Jackson, R.M.; Robson, C.N.; Leung, H.Y.; Elliott, D.J. A novel androgen-regulated isoform of the TSC2 tumour suppressor gene increases cell proliferation. Oncotarget 2014, 5, 131–139. [Google Scholar] [CrossRef] [Green Version]
- Amaral, C.L.; Freitas, L.B.; Tamura, R.E.; Tavares, M.R.; Pavan, I.C.; Bajgelman, M.C.; Simabuco, F.M. S6Ks isoforms contribute to viability, migration, docetaxel resistance and tumor formation of prostate cancer cells. BMC Cancer 2016, 16, 602. [Google Scholar] [CrossRef]
- Sawada, T.; Kanemoto, Y.; Amano, R.; Hayakawa, A.; Kurokawa, T.; Mori, J.; Kato, S. Antagonistic action of a synthetic androgen ligand mediated by chromatin remodeling in a human prostate cancer cell line. Biochem. Biophys. Res. Commun. 2022, 612, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Prigol, A.N.; Rode, M.P.; Silva, A.H.; Cisilotto, J.; Creczynski-Pasa, T.B. Pro-angiogenic effect of PC-3 exosomes in endothelial cells in vitro. Cell. Signal. 2021, 87, 110126. [Google Scholar] [CrossRef] [PubMed]
- de Bouteiller, O.; Merck, E.; Hasan, U.A.; Hubac, S.; Benguigui, B.; Trinchieri, G.; Bates, E.E.; Caux, C. Recognition of double-stranded RNA by human toll-like receptor 3 and downstream receptor signaling requires multimerization and an acidic pH. J. Biol. Chem. 2005, 280, 38133–38145. [Google Scholar] [CrossRef] [Green Version]
- Bugge, M.; Bergstrom, B.; Eide, O.K.; Solli, H.; Kjonstad, I.F.; Stenvik, J.; Espevik, T.; Nilsen, N.J. Surface Toll-like receptor 3 expression in metastatic intestinal epithelial cells induces inflammatory cytokine production and promotes invasiveness. J. Biol. Chem. 2017, 292, 15408–15425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robbins, M.; Judge, A.; MacLachlan, I. siRNA and innate immunity. Oligonucleotides 2009, 19, 89–102. [Google Scholar] [CrossRef] [PubMed]
- Fujisawa, M.; Kanda, T.; Shibata, T.; Sasaki, R.; Masuzaki, R.; Matsumoto, N.; Nirei, K.; Imazu, H.; Kuroda, K.; Sugitani, M.; et al. Involvement of the Interferon Signaling Pathways in Pancreatic Cancer Cells. Anticancer Res. 2020, 40, 4445–4455. [Google Scholar] [CrossRef]
- Deveci Ozkan, A.; Guney Eskiler, G.; Kaleli, S.; Sahin, E. Immunotherapeutic role of cabazitaxel treatment in the activation of TLR3 signalling in metastatic castration-resistant prostate cancer in vitro. Mol. Biol. Rep. 2022, 49, 1261–1271. [Google Scholar] [CrossRef]
- Kourko, O.; Smyth, R.; Cino, D.; Seaver, K.; Petes, C.; Eo, S.Y.; Basta, S.; Gee, K. Poly(I:C)-Mediated Death of Human Prostate Cancer Cell Lines Is Induced by Interleukin-27 Treatment. J. Interferon Cytokine Res. 2019, 39, 483–494. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Basal (% LDH Released) | Emax (% LDH Released) | EC50 (nM) |
|---|---|---|---|
| DTX | 4.2 ± 0.1 | 50.6 ± 1.1 | 7.7 ± 1.3 |
| DTX + SCR-siRNA (100 nM) | 24.9 ± 3.2 | 68.1 ± 2.4 | 5.6 ± 0.6 |
| DTX + p42-siRNA (100 nM) | 14.3 ± 2.7 | 78.9 ± 2.2 | 6.1 ± 1.1 |
| DTX + Rheb-siRNA (100 nM) | 15.5 ± 3.7 | 71.8 ± 2.3 | 5.9 ± 0.7 |
| DTX + p42-siRNA (50 nM) + Rheb-siRNA (50 nM) | 18.3 ± 4.2 | 73.8 ± 2.1 | 5.8 ± 0.7 |
| Treatment | Basal (% LDH Released) | Emax (% LDH Released) | EC50 (nM) |
|---|---|---|---|
| DTX | 3.7 ± 0.7 | 42.7 ± 1.3 | 5.3 ± 0.6 |
| DTX + SCR-siRNA (100 nM) | 14.3 ± 1.7 | 47.1 ± 1.6 | 22.1 ± 4.3 |
| DTX + p42-siRNA (100 nM) | 12.6 ± 3.2 | 42.4 ± 3.8 | 9.1 ± 4.6 |
| DTX + Rheb-siRNA (100 nM) | 12.1 ± 2.3 | 41.2 ± 1.9 | 8.9 ± 2.7 |
| DTX + p42-siRNA (50 nM) + Rheb-siRNA (50 nM) | 15.2 ± 1.9 | 44.5 ± 3.5 | 14.1 ± 6.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de la Torre, C.; Játiva, P.; Posadas, I.; Manzanares, D.; Blanco, J.L.J.; Mellet, C.O.; Fernández, J.M.G.; Ceña, V. A β-Cyclodextrin-Based Nanoparticle with Very High Transfection Efficiency Unveils siRNA-Activated TLR3 Responses in Human Prostate Cancer Cells. Pharmaceutics 2022, 14, 2424. https://doi.org/10.3390/pharmaceutics14112424
de la Torre C, Játiva P, Posadas I, Manzanares D, Blanco JLJ, Mellet CO, Fernández JMG, Ceña V. A β-Cyclodextrin-Based Nanoparticle with Very High Transfection Efficiency Unveils siRNA-Activated TLR3 Responses in Human Prostate Cancer Cells. Pharmaceutics. 2022; 14(11):2424. https://doi.org/10.3390/pharmaceutics14112424
Chicago/Turabian Stylede la Torre, Cristina, Pablo Játiva, Inmaculada Posadas, Darío Manzanares, José L. Jiménez Blanco, Carmen Ortiz Mellet, José Manuel García Fernández, and Valentín Ceña. 2022. "A β-Cyclodextrin-Based Nanoparticle with Very High Transfection Efficiency Unveils siRNA-Activated TLR3 Responses in Human Prostate Cancer Cells" Pharmaceutics 14, no. 11: 2424. https://doi.org/10.3390/pharmaceutics14112424
APA Stylede la Torre, C., Játiva, P., Posadas, I., Manzanares, D., Blanco, J. L. J., Mellet, C. O., Fernández, J. M. G., & Ceña, V. (2022). A β-Cyclodextrin-Based Nanoparticle with Very High Transfection Efficiency Unveils siRNA-Activated TLR3 Responses in Human Prostate Cancer Cells. Pharmaceutics, 14(11), 2424. https://doi.org/10.3390/pharmaceutics14112424