Abstract
Diabetic foot ulcers (DFUs) are characterized by a lack of angiogenesis and distal limb diabetic neuropathy. This makes it possible for opportunistic pathogens to protect the biofilm-encased micro-communities, causing a delay in wound healing. The acute and chronic phases of DFU-associated infections are distinguished by the differential expression of innate proinflammatory cytokines and tumor necrosis factors (TNF-α and -β). Efforts are being made to reduce the microbial bioburden of wounds by using therapies such as debridement, hyperbaric oxygen therapy, shock wave therapy, and empirical antibiotic treatment. However, the constant evolution of pathogens limits the effectiveness of these therapies. In the wound-healing process, continuous homeostasis and remodeling processes by commensal microbes undoubtedly provide a protective barrier against diverse pathogens. Among commensal microbes, probiotics are beneficial microbes that should be administered orally or topically to regulate gut–skin interaction and to activate inflammation and proinflammatory cytokine production. The goal of this review is to bridge the gap between the role of probiotics in managing the innate immune response and the function of proinflammatory mediators in diabetic wound healing. We also highlight probiotic encapsulation or nanoformulations with prebiotics and extracellular vesicles (EVs) as innovative ways to tackle target DFUs.
1. Introduction
The term “neuropathy” in diabetes is a generic term that refers to the loss of sensation and steadiness of the distal limb. The loss of sensation affects peripheral blood movement. Therefore, dysregulating the protective sensation leads to systemic inflammatory status of diabetic foot ulcers (DFUs) [1]. Characterizing the type of wound—which considers various factors such as the length of the wound, the size of the ulcer, and its location—is a crucial step in the evaluation of DFUs. These parameters primarily indicate whether the ulcer is acute or chronic in nature [2]. From the acute to the gangrene stages, DFUs express differently. The acute stage begins with a recurring loss of sensation and subcutaneous hemorrhage of the lesion, which usually goes unnoticed and results in foot deformity. It normally heals in three weeks, but improper wound management in the acute stage leads to a chronic or infectious stage with inflammatory dysregulation [3].
However, due to the broad pathophysiology of DFU infection, relying on any single therapy [4] causes local inflammation, bleeding, and a high increase in blood oxygen levels. To overcome such limitations, therapies targeting immune cells using fibroblasts, stem cells, grafts, monoclonal antibodies, and bioactive molecules have been recently developed. However, these therapies have some limitations. A short half-life, the need for repeated administration, high costs of production, short-term bioactivity, insufficient data on large populations, limited downstream processing, renal clearance, and ineffectiveness against polymicrobial infections are all issues that must be addressed [5].
Gut immunological homeostasis, which is maintained by the interaction among intestinal microflora, is crucial in controlling the host’s inflammatory response. The gut microbiota influences various parts of the human body, including the brain, liver, and skin. Concerning DFU infection, commensal microbes (Staphylococcus epidermidis, Propionibacterium, and Cutibacterium species) on human skin have direct coordination with the gut microbiota and govern chronic responses during skin–pathogen contact [6]. Among gut microbiota, probiotics have been used to treat a variety of health issues since long ago and have also shown promising results as immunomodulators during chronic illness [7]. Probiotics already have several advantages, including non-toxicity, user-friendliness, strong immunity, a longer half-life, an easy route of administration, and simple downstreaming [8]. However, questions still remain about the use of probiotics as a DFU therapeutic. Constant remodeling of probiotics through encapsulation or nanoformulation using prebiotics and synbiotics, biogenic nanoparticles (NPs), and extracellular vesicle (EVs) originating from probiotics can open new avenues in the field of DFU management. These strategies are still unexplored.
In this review, we first introduce the importance of the gut–skin axis in maintaining skin homeostasis and emphasize it as a new route for DFU therapy using probiotics. Then, the role of probiotics in maintaining acute and chronic conditions of DFUs will be further discussed. Further, a novel idea on how probiotics can reprogram acute and chronic modulation during DFU infections will be introduced as a future application of probiotics. These include cutting-edge technologies such as the encapsulation or nanoformulation of probiotics using prebiotics and synbiotics, biogenic NPs, and EVs. Overall, our review will provide perspectives on probiotic-mediated therapeutics against infections in acute and chronic DFUs.
2. Method
For this review paper, we searched the databases (PubMed, Science Direct, Web of Science, and Scopus) for articles that majorly dealt with the following keywords: “diabetic foot ulcer”, “diabetes”, “biofilm”, “acute”, “chronic”, “pathogen”, “inflammatory cytokine”, “cytokine”, “macrophage”, “fibroblast growth factor”, “stem cell”, “neuropeptide”, “monoclonal antibody”, “ bioactive”, “gut skin axis”, “probiotic”, “symbiotic”, “extracellular vesicle”, “postbiotic”, and “paraprobiotics”. In addition, case studies, review studies, and retrospective studies involving randomized clinical trials relevant to diabetic foot ulcer and its therapeutic approaches were included. Moreover, we explored the bibliographies of all retrieved and relevant articles.
3. Diabetes: A Constant Fluctuation in Blood Glucose Levels
According to the International Diabetes Federation Atlas (IDFA), diabetes is prominent in every part of the world. Over 537 million people suffered from diabetes around the world in 2019, and cases are expected to increase to 700–750 million worldwide by 2045 [9]. A global survey revealed 6.7 million deaths due to diabetes in 2021 (1 every 5 s) involving demographic locations including Africa (416,000), Europe (1.1 million), the Middle East (796,000), Southeast Asia (747,000), and the Western Pacific (2.3 million). Diabetes in the current scenario holds the position of being a chronic and long-term hypo/hyperglycemic condition that can result in multiple complications such as cardiovascular disorders, neuropathy, nephropathy, and retinopathy [10]. The lifestyle pattern and genetic disposition from different demographic locations normally explain the prognosis of diabetes, but substantial research indicated that the gut microbiota, which are crucial to human physiology, metabolism, and dysbiosis, have a direct role in type 2 diabetes mellitus (T2DM) [11]. T2DM is frequently associated with obesity; it is noteworthy that certain compositions of gut microbiota have been linked to insulin resistance and glycemic control problems [12]. Changes in the composition of gut bacteria may contribute to the etiology of diseases such as obesity, diabetes, and kidney disease, the prevalence of which are still rising globally [13]. These situations generally involve diabetic ketoacidosis and hyperosmolar state, which play a significant role in automatic and peripheral neuropathy [14]. The glycemic complications related to diabetes involve both hypoglycemia and hyperglycemia, but hyperglycemic DFUs account for most of the mortality and morbidity [15].
4. The Diabetic Foot Ulcer: A Long-Lasting Foot Deformity
The DFU, one of the major complications of diabetes, affects 25 to 30% of people during their lifetime. Among them, 20% undergo lower limb amputation with an annual mortality rate of 11% for DFUs and 22% for amputees [16]. Every 20 to 30 s, a lower limb gets amputated, with DFUs accounting for 85–95% of the cases [17]. Patients with high blood glucose levels exhibited severe pathological conditions such as tissue hypoxia, leading to inadequate blood supply to the vascular endothelial cells [18]. The prognosis involves a localized injury to the distal portion of the ankle, which goes unnoticed due to inefficient sensory response, peripheral neuropathy, and vascular lesions. The lesion paves the way for opportunistic pathogens that increase the chances of tissue hypoxia by reducing the process of angiogenesis and endothelial revascularization, which in turn delay the wound-healing process. This delay in the wound-healing process results in the chronic stages of DFUs and the formation of gangrene [19,20]. The process from neuropathy to gangrene follows a gradual infectious route, which leads to a high microbial bioburden [21] and delays the wound-healing process at chronic stages due to the formation of microbial communities known as biofilms. High blood glucose levels increase the concentration of proteins and carbohydrates, which form the basic core for persistent infections. About 40 to 80% of diabetic patients develop recurrent infections, and between 20 to 25% develop deep infections with osteomyelitis [22]. To select the appropriate treatment course and manage care for DFU patients, several classification systems have been developed to elucidate the characteristics of DFUs and the severity of infections as described below.
4.1. Evaluation and Classification of DFU Extent
The systematic evaluation of DFUs defines the constant prognosis of foot ulcers. Health professionals determine the etiology of the foot and verify the extent of lesions that involve acute Charcot foot or chronic ischemic foot [23]. Various classification systems have been developed to describe ulcer characteristics and its lower limb extremities.
4.1.1. Meggitt–Wagner (MW) Classification System
Developed in the 1970s, this system comprises six ulcer grades that range from 0 to 5. This system assesses the pro-ulcerative stage, superficial infection, subcutaneous infection, deep ulcer in tendons, forefoot gangrene, and whole-foot gangrene (50% foot infection) [24]. This system is simple and widely accepted for predicting lower extremity amputation. However, this system is not recommended for use in assessments of DFUs because it does not adequately address all DFU subtypes and the spectrum of infections. The major limitations of this system are the infection rate and tissue viability [25].
4.1.2. University of Texas (UT) Classification
This system is a modified version of the MW classification that has also been effective in predicting lower extremity amputation. Moreover, this system overcomes all of the shortcomings of MW, especially the depth of lesion and infection rate [26]. This system uses four grades (0 to 3; depth) and four stages (A–D; severity of wound) to classify DFUs by marking the presence of infection, ischemia, or both [27]. This system helps predict the infection rate and the rate of amputation and is used in practice; however, it is ineffective in determining the degree of neuropathy and microbial load differentiation.
4.1.3. Perfusion, Extent, Depth, Infection, and Sensation (PEDIS)
This system was developed in 2003 by International Working group of Diabetic Foot (IWGDF) solely for determining the rate of infection. It includes four categories. The first is uninfected or pro-ulcerative (the same as MW and UT); the second involves signs of infection, erythema, or colitis. The last two stages involve the severity of the infection and its related circumstances such as moderate to severe infection and systemic toxicity involving fever, chills, hypertension, and cardiovascular disorders [28]. It is a complex system that does not truly define the ulcer types but has an advantage in terms of choosing empirical antibiotic therapy against chronic infection.
4.1.4. Saint Elian Wound Score System (SEWSS)
This is an advanced version of the PEDIS system. This system uses three grades (I to III) based on 10 factors in the following categories: location, topographic aspects, the number of affected zones, ischemia, infection, edema, neuropathy, depth, area, and wound-healing phase. The total score is 6 to 30 points, and the score can switch to a grade. It is an advantageous system compared to others and can efficiently detect the outcome of DFUs (minor amputation and wound healing), but it is time-consuming [29].
4.1.5. Site, Ischemia, Neuropathy, Bacterial Infection, Area, and Depth (SINBAD)
This is a versatile and rapid system that uses five clinical features (site of infection, ischemia condition, neuropathy, bacterial infection, and depth), which are graded as either present (0) or absent (1). This system is primarily used in various rural regions where DFU occurrence is very prominent and does not require any medical setup for evaluation. SINBAD generally provides a high degree of versatility and also helps to differentiate between the acute and chronic conditions in the context of validating research and consistent results [30].
5. Timeline and Stages of DFUs
The open lesion and complete vascularization take 8 to 14 days for DFU wound closure [31], but the normal healing process depends on the stages of the foot ulcer, considering the previous classification of DFUs (Section 4.1) from the superficial wound stage to gangrene of whole foot. A wound can be defined as an acute or chronic type. The latter is the most infectious and non-healing condition and involves factors such as high infection and release of proinflammatory responses. The major pathways in acute and chronic conditions are shown in Figure 1.
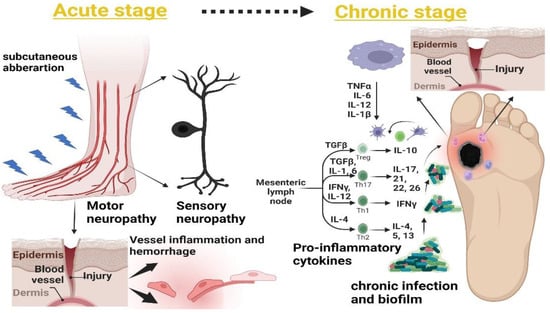
Figure 1.
Acute and chronic conditions of the DFU. The acute stage includes subcutaneous aberration, which is associated with loss of sensation at the distal part of the wound. The continuous aberration at the wound causes blood vessel angiogenesis to fail, resulting in inflammation at the wound site (left). The chronic stage is characterized by continuous wound damage and the movement of proinflammatory responses. This causes skin moistening, which allows opportunistic pathogens to form a biofilm, delaying the wound-healing mechanism during a DFU (right).
5.1. Sensory and Motor Neuropathy
Sensory and motor neuropathy involve constant pressure and local necrosis. The high pressure on the peripheral veins leads to constant injuries that result in autonomic tissue injury. There might be instances that can result in small areas of gangrene or ulceration [32]. Additionally, autonomic neuropathy includes thermoregulatory dysregulation that involves excessive sweating, cracking, and edema in the foot region, which further adds to the ulceration [33].
5.2. Subcutaneous Hemorrhage
The skin breakdown during the tissue damage leads to subcutaneous hematoma involving deep tissue cellulitis near bones and tendons, which is normally manifested as pain and tenderness of the epidermis and dermis [34]. The loss of sensory and motor reflexes results in the formation of thickened skin that is generally referred to as a “callus”, which causes constant pressure that leads to foot deformity. The formation of small lesions near the callus region can be treated by using debridement and fitting shoes, but improper management can subsequently lead to foot deformation and mechanical stress [35].
5.3. Foot Deformity
Various risk factors such as deep tissue necrosis and motor and sensory polyneuropathy lead to diabetic neuro-osteoarthropathy, a term related to deformity in the tarsus and metatarsus region of the foot. The classic example is Charcot foot associated with the disorientation of bones and joints, which are further related to bone resorption and site-specific inflammation [36]. A study conducted in a Copenhagen wound-healing center from 1995 to 2015 showed a significant relationship between acute Charcot foot and the prognosis of ulcer. This study highlighted the social economic impact of Charcot foot ulcers on type I diabetic patients. Out of 173 patients, only 4% were able to perform their normal routine jobs, while 60–65% were below the basic social economic conditions [37].
5.4. Acute Inflammatory Responses (Neutrophils and Macrophages) and Accumulation of Extracellular Matrix (ECM)
A hyperglycemic condition always shows a low-grade inflammatory response to the microvascular environment during DFUs. The increased diabetic stress results in high inflammatory responses that result in the release of white blood cells (WBS) and acute phase proteins including C-reactive proteins, fibrinogen, chemokines, and interferon-γ [38]. A high glycemic index and localized inflammation result in the production of interleukins such as IL-6 from the macrophage. IL-6 is known for its proinflammatory activities; its constitutive expression results in a delay in wound healing [39]. Muller et al. [40] showed the effects on type 2 diabetic patients of an increased release of IL-6 and tumor necrosis factor (TNF) receptors, highlighting the role of IL-6 modulation as a diagnostic marker in the evaluation of inflammatory response during DFUs. Localized inflammation is related to innate and proinflammatory responses during chronic conditions, including a higher count of neutrophils, T cells, and mast cells, which represent the markers and generally delay the wound-healing process. The high expression of neutrophils is further related to revascularization via protease production, collagen, cathepsin, and matrix-degrading substances such as metalloproteinase [41].
5.5. Abnormal Matrix Metalloproteinase (MMP) Expression
Matrix metalloproteinases are important factors in wound healing. The expression of MMPs regulates the inflammatory phase and the endothelial basement for angiogenesis, but in cases of chronic DFU, the abnormal expression of MMP-9 significantly delays the wound-healing process [42]. The overexpression of MMP-9 upregulates the protease production, which results in the accumulation of macrophages and neutrophils, which in turn results in excessive collagen production that hampers the normal wound-healing process [43].
5.6. Bacterial Invasion and Persistent Infection
The overall prognosis of a DFU lies in the management of foot ulcers, but the bottom line of the management depends on the severity of the DFU. Inadequate management during sensory and motor neuropathy and constant friction of the foot leads to the moistening of the foot region that involves the movement of opportunistic pathogens. A total of 80% of traumatic wounds lead to amputation; the remaining 20% lead to recurrent bacterial infection [44]. Inflammation and persistent infection, which are the major causes of amputation, involve constant chemotaxis movement of T and B cells during infection. Infection is the only factor in DFUs that involves both the acute and chronic stages of DFUs; i.e., ischemia, callus formation, and hyperproduction of the proinflammation cytokine [45].
6. Microbial Bioburden in DFUs
The microbial spectra in DFUs are often related to the moistness and breaking of the cutaneous region of the wound, which in due course adversely affect the normal healing of the wound [46]. Normally, the onset of foot infections involves superficial or subcutaneous categories, but these infections can spread to the tendons, joints, and bones, causing severe osteomyelitis [47]. The microbial diversification varies according to the extent of the DFU infection. According to the guidelines of the Infectious Disease Society of America (IDSA), superficial infections generally involve aerobic microorganisms such as Staphylococcus, Micrococcus, Corynebacterium, Pseudomonas, and Klebsiella spp. [45]. A deep tissue or osteomyelitis condition involves the presence of some facultative anaerobic and strictly anaerobic microorganisms including Peptostreptococcus spp., Bacteroides spp., Prevotella, Fusobacterium, and Pseudomonas spp. [48]. The heterogeneous nature of wounds involves polymicrobial interaction, which corresponds to the high microbial load during chronic infection [49]. However, Gram-negative bacteria have a prevailing existence in the wound microbiome compared to Gram-positive bacteria. The various microbial communities coexisting in DFUs play different roles and are therefore essential in the multiple stages of DFUs. Chronic wounds often involve various small molecules or autoinducers that generally act as biomarkers in metagenomic studies of wounds [50]. Polymicrobial infections often involve cell-to-cell communication via these autoinducers, which enables the production of virulence factors such as the protease, elastase, rhamnolipid, and extracellular polysaccharide matrix (EPS) [51]. The synergistic adaptation of microbes to the wound site includes embedded micro-communities in a polymeric matrix known as a biofilm. The term biofilm is often related to the chronicity of a wound; i.e., the higher the microbial load, the more compact the structure of the biofilm will be [52]. The presence of a biofilm in DFU cases involves 60 to 80% of chronic cases and 8 to 10% of acute cases. These micro-communities strongly limit the entry of antimicrobial agents to the site of infection, resulting in prolonged hospitalization [53]. Aerobic microbes such as Pseudomonas and Staphylococcus are generally frequently isolated from chronic infection sites. These microbes also produce strong biofilms in the polymicrobial load that influence the presence of other pathogens [54]. A study conducted on Pseudomonas and Staphylococcus biofilms showed the production of virulence factors such as exopolysaccharide (EPS), showcasing the prolonged chronic nature of the wound. The increased microbial burden also showed the accumulation of inflammatory responses that affected the normal healing process [55]. The persistent infection response involves the movement of innate immune cells such as neutrophils, macrophages, and leukocytes. This ultimately increases the reactive oxygen species (ROS) production and release of proinflammatory markers at the site of infection, leading to a non-healing situation of the ulcer [56].
7. Innate Immune and Proinflammatory Mediators in Non-Healing DFUs
Chronic diabetic wounds have four distinct phases of healing: (i) the inflammatory response, (ii) the proliferative stage, (iii) migration, and (iv) revascularization [57]. In acute conditions, motor and sensory neuropathy are generally addressed by proper management and wound assessment, which involve evaluation of subcutaneous hemorrhage and deformity. The chronic condition involves immune outburst at the site of infection and an increased expression of innate immune cells such as mast cells, lymphocytes, neutrophils, macrophages, C-reactive proteins, proinflammatory cytokines, and ROS, which cause a delay in re-epithelization and angiogenesis [58].
7.1. Innate Immune Cells
Innate immune cells including neutrophils and macrophages are primary cells involved in the wound-healing process and early vascularization of a wound [59]. However, during a non-healing diabetic wound, there is a slow chemotaxis movement of cells that results in decreased adherence and proliferation, leading to an inadequate phagocytic elimination [60]. Apart from neutrophils, macrophages have also shown an impaired role during the wound-healing process. Aberrant stimulation of macrophages during diabetic wounds involves restricted switching from M1 to M2 macrophages, resulting in a constant outburst of IL-1β and TNF-α [61]. A study conducted on diabetes-induced mice showed an elevated level of a proinflammatory cytokine (IL-1β) that was exhibited by the proinflammatory responses of altered M1 and M2 macrophages at the site of infection. This differential expression of transforming growth factors (TGFs), an insulin-like growth factor (IGF-1), and MMP-9 highlighted the impaired response of macrophage switching in delaying the wound-healing process [62]. Macrophages and monocytes also play important roles in wound healing and collagen deposition during epithelization. Impaired regulation of either of the two leads to the aberrant transition of proinflammatory cytokines, resulting in delayed wound healing [63].
7.2. Proinflammatory Cytokines
ILs and TNF generally constitute the major classes of proinflammatory cytokines that promote insulin resistance and impair healing. Kolumam et al. [64] showed that IL-6, -18, -22, and -24 are highly expressed in diabetic wounds compared to non-diabetic wounds, highlighting the differential expression of pro- and anti-inflammatory responses during DFUs. Similarly, TNF-β is generated by monocytes and macrophages and helps to induce T- and B-cell responses to regulate the vascularization of an endothelial cell. During DFU dysregulation, an immunomodulatory burst (IL-1β and TNF-α) results in an impaired insulin metabolism, altering the humoral expression (B-cell regulation) [65]. A study conducted by Trengove et al. [66] on patients from Western Australian hospitals with chronic non-healing venous ulcers highlighted the direct relationship of an altered innate response and altered wound healing. The bioassay showed higher expressions of IL-1 and -6. Furthermore, levels of IL-1α, IL-β, and TNF-α showed a marked increase in the non-healing wound. Similarly, growth factors such as epidermal growth factor (EGF), platelet-derived growth factor (PDGF), and TGF-β1 were also altered in chronic venous ulcers.
Apart from innate immune cells, TLRs are one set of immune cells that link innate immunity with adaptive immunity and play a role against microbial invasion by targeting the lipopolysaccharide layer of pathogens [67]. The significant expression of TLRs during infection results in the production of proinflammatory responses; normally, during the healing of a wound, the expression of TLRs (TLR-2 and -4) decreases with tissue vascularization. A study by Pukstad et al. [68] on non-healing chronic wound ulcers showed a significant increase in the expression of TLR-2 and -4, which constitutively led to the expression of proinflammatory markers (IL-1α, IL-1β, and MIP-1δ). Hence, reducing the levels of anti-inflammatory cytokines (IL-8 and MIP-1α) and dysregulating TLRs can modulate the healing procedure in chronic wound infections.
7.3. Matrix Metalloproteinase
Apart from proinflammatory cytokines, MMPs also exert inflammatory responses in chronic diabetic wounds. In recent years, MMPs have gained a higher value in studies related to non-healing diabetic wounds and have been linked to the impaired activity of IL-1β and TNF-α [69]. A study conducted on acute and chronic surgical wounds showed a significantly higher expression of MMP-9 in chronic as compared to acute wounds. The increased expression of MMP-9 (from 10- to 25-fold) also highlighted the reduced expression of elastase, which was directly correlated with the delay in the endothelial vascularization [70]. MMPs play a vital role in tissue remodeling, and their differential expression (MMP-1, -2, -8, and -9) regulates poor wound healing [71].
7.4. Toll-Like Receptors (TLRs)
Impaired wound healing during DFUs involves the activation of TLRs, which recognize inflammatory factors such as pathogen-associated molecular patterns (PAMPs) of bacteria during chronic DFUs [72]. PAMPs generally participate in the activation of TLR signaling and have an important role in infection-mediated immune responses in DFUs [73]. Alteration in the microbial bioburden results in the expression of differential TLRs (TLR-1, -2, -4, -6, and -8); these factors have also been used as inflammatory biomarkers in DFUs. A study by Dasu et al. [74] on streptozotocin-induced mice showed increased expression of TLR-2 and TLR-4 with a high glycemic index, highlighting the T-cell-mediated responses in the inflammatory phase of DFUs. Creely et al. [75] also showed that the hyperglycemic state in DFUs results in inflammation in adipose tissues that increased the expression of TLR-2 and TLR-4. This in turn increased the expression of proinflammatory cytokines (TNF-α, IL-6, and NF-kB), impairing the normal healing process. EPS formation and the biofilm matrix also stimulate the production of proinflammatory cytokines (TLRs and TNF-α). A study on biofilm-producing S. aureus-induced wounds on a type 2 diabetic rat model showed an increased outburst of TLR-2, 4, and TNF-α, supporting a direct correlation with biofilm-mediated inflammatory responses [76]. DNA methylation also has a significant role in altering the normal wound-healing procedure. A study by Singh et al. [77] on the epigenetic role of TLR-2 showed a significant downregulation of TLR-2 in type 2 diabetes, highlighting the partial methylation of the CpG islands in the chronic diabetic wound.
8. Novel Therapies Targeting Inflammatory Modulators during Non-Healing DFUs
Based on our review, we understand that the multidisciplinary approaches of different specialties are required to decrease wound bioburdens and the chances of lower limb amputations. Since the management of DFU wounds varies from stage to stage [78], providing adequate care to the foot region is important. Conventional therapies such as debridement [79], hyperbaric oxygen therapy [80], shock wave therapy [81], offloading [82], larval therapy [83], laser therapy [84], and empirical antibiotic treatment [85] are promising as efficient treatments, but they are limited in terms of scope with respect to tissue vascularization and the healing process of ischemic ulcers. Therefore, the cost-to-benefit ratios are lower, and retrospective studies conducted on these therapies require more evidence from a larger demographic area. The major advantages and disadvantages of the above therapies are summarized in Table 1.

Table 1.
Advantages and disadvantages of conventional therapies.
During diabetic wounds, it is imperative to reduce the production of proinflammatory cells and ROS. The delay in apoptosis results in tissue granularization and long-term chronic inflammation, which further contributes to the poor healing of the wound site, resulting in a loss of epithelization [93]. The previous alternatives shown in Table 1 have limitations in being used for a longer period; therefore, some novel therapies are being investigated that involve fibroblast growth factors, macrophage-regulating drug therapy, stem cell therapy, neuropeptides, MMP inhibitors, monoclonal antibodies, and plant-derived bioactive compounds.
8.1. Fibroblast Growth Factor (FGF)-Associated Therapy
Basic fibroblast growth factors (bFGFs) have a prominent role in diabetic wound healing. They were found to activate signaling pathways of MAPK and JAK-STAT, which affect the basic movement of fibroblasts and keratinocytes in endothelial vascularization and collagen formation [94]. Therapies including fibroblasts have been used since the early 1990s because fibroblasts were regarded as a prominent factor in the formation of granular tissues during epithelization. A study on FGF and a keratinocyte-based hydrogel against diabetic rates showed a rapid healing rate and granularization of endothelial tissue. This study also showed that the fibroblast-associated hydrogel was effective against proinflammatory cytokines. It decreased IL-6, TNF-α, and IL-10 production and increased the expression of pro-healing factors such as TGF-β and collagen III after 1 to 2 weeks of therapy [42].
8.2. Macrophage-Regulating Therapy
The role of macrophages in acute and chronic injury involves phagocytosis and the removal of damaged cells from the site of infection. Characteristically, M1 macrophages activate antigen-presenting cells (APCs), toll-like receptors (TLRs), and MHC-II (major histocompatibility complex) [95]. Meanwhile, M2 macrophages produce elevated levels of TGF-β and IL-10. M1 and M2 macrophages work in tandem in the migration and proliferation of fibroblasts and keratinocytes to the endothelial lining [96]. However, during impaired healing, the production of M2 macrophages is sustained, resulting in the higher expression of IL-6 and TNF-α. Herbs such as Plectranthus amboinicus (PA), which have been studied for their anti-inflammatory and antibacterial activities, have known activity against proinflammatory responses and macrophage maturation [97]. A study conducted by Lin et al. [98] on a PA-F4 extract showed homeostasis between M1 and M2 expression, which increased collagen synthesis and fibroblast proliferation. Additionally, the expression of proinflammatory cytokines (IL-1β and TNF-α) was reduced, which further manifested the macrophage and monocyte chemotaxis.
8.3. Stem Cell Therapy
Mesenchymal stem cells (MSCs) are one of the prominent cells that portray various benefits in graft therapies such as better healing and angiogenesis properties [99]. A study on the intradermal injection of bone-marrow-derived MSC stem cells in diabetic rats showed an increased expression of IL-10 and M2 macrophages. We highlighted the pro-healing efficiency of MSC-derived stem cells in initiating the wound healing of chronic wound ulcers by directly regulating the expression of anti-inflammatory cells such as IL-10 and M2 macrophages. The MSC-derived stem cells also showed a decreased expression of IFN-γ and TNF-α, showcasing the clearance of proinflammatory markers during the wound-healing process [100].
8.4. Neuropeptides
Diabetic neuropathy or acute cutaneous infection has been found to correlate with neuropeptides, which facilitate the activation of the immune system and cell proliferation. Few neuropeptides, such as calcitonin gene-related peptide (CGRP), neuropeptide Y (NPY), neurotensin (NT), and α-melanocorticotropin-releasing hormone (α-MSH) were found to be potential biomarkers in diabetic wound healing. These neuropeptides promote the expression of interferon (IFN-β), TGF-β, and the macrophage inflammatory protein (MIP-1α) [101]. A study conducted on the CGRP protein in adult epidermal keratinocytes from breast cancer showed a marked increase in the production of proinflammatory cytokines (IL-1α, IL-8, and TNF-α). This induced an increased expression of pro-nerve growth factor (NGF) and fibroblast and keratinocyte proliferation, highlighting the direct role of the neural system in skin homeostasis [102].
8.5. Matrix Metalloproteinase (MMP) Inhibitors
DFU infections involve the production of MMPs; these proteases are present in acute and chronic phases of infection, but their production regulates the normal healing of the wound. Normally, MMP-8 and -9 are the major inflammatory markers that are present in a DFU wound. However, MMP-8 acts as a pro-healing factor, whereas MMP-9 overexpression results in delayed wound healing [103]. A study conducted on MMP-8 knocked-out rats showed a decreased wound healing ability and lower keratinocyte movement, which ultimately increased MMP-9 expression, showcasing increased proteinase activity. This study also showed a lower expression of the anti-inflammatory cytokine TGF-β that was affected by increased MMP-9 activity [104]. Recent technologies and therapies have been developed to overcome MMP-9 overexpression. Small toxic molecules such as enantiomer racemic ND-336 [105] and ND-332 [106] have been identified as selective inhibitors of MMP-9 that result in the downregulation of proinflammatory cytokines. RNA-based therapies have also been used that involved small non-coding RNAs. A study conducted by Wang et al. [107] on microRNA (miRNA-129) showed the downregulation of MMP-9 proteinase activity, making it a possible therapeutic agent in diabetic wound healing. This study showed that miRNA-129 acts as a direct inhibitor of the MMP-9 regulatory protein (specificity protein-1; Sp-1), which in turn increases the fibroblast production and keratinocyte activation in DFU healing.
8.6. Monoclonal Antibodies (mAbs)
Antibody therapy or mAb therapy against infectious agents involves the determination of epitopes on the microbial flora, resulting in toxin neutralization that limits pathogen invasion. The use of mAbs against multiple surface proteins and polysaccharide regions invaded by opportunistic pathogens helps to fight various bacterial pathogens such as methicillin-resistant S. aureus (MRSA) and polymicrobial biofilm-forming microbes [108]. A recent report included the use of AZD6389 mAb ((anti-alpha toxin (AT), anti-clumping factor, and cross-neutralizing leukotoxin) and tyrosine kinase met (C-mAb)) against Staphylococcus and Pseudomonas infection in a diabetic rat model. The study on the AZD6389 mAb showed a decreased expression of proinflammatory cytokines (IFN-γ and IFN-α) and MMP-9 metalloproteinase with neutrophil elastase. This study also highlighted a decrease in systemic inflammation and rapid wound closure [109]. Choi et al. [110] showed that C-mAb lowers inflammatory outbursts by confirming the decreases in MMP-9 and TGF-β, resulting in fibroblast proliferation and increased vascular endothelial growth factor (VEGF) production, which improved diabetic wound healing.
8.7. Bioactive Molecules
The role of bioactive molecules is to speed up the angiogenesis process with beneficial endothelial vascularization, thus improving the inflammatory and antimicrobial activities in DFU healing [111]. A polyphenol-like compound; i.e., propolis, showed increased antioxidant and anti-inflammatory properties in wound healing. The topical administration of propolis showed enhanced activity against diabetic wound closure with re-epithelization via enhanced production of collagen and TGF-β signaling factors. On the contrary, there was a decrease in the expression of proinflammatory cytokines (IL-6 and TNF-α) and MMP-9 [112]. Another potent bioactive agent is naturally produced epoxy-tiglianes from bush wood trees. Epoxy-tiglianes showed an effective degradation of bacterial biofilms by inducing the production of antimicrobial genes (DEFB4 and DEF103A) and leukocyte survival factors (IL-1β, IL-17, CXCL-8, and CCL-20). It also induced protein-kinase-dependent ROS generation, which resulted in enhanced fibroblast and keratinocyte production [113].
The above-mentioned therapies have shown prominent effects in both acute and chronic conditions; however, major drawbacks such as limited cell survival, a shorter half-life, and adverse effects (oral ulcer and high cell death) constantly exert pressure on pharmaceutical companies to generate more sustainable therapies. Constant efforts are being made to develop more efficient therapies against DFU infections; these involve few readily available therapies such as Leucopatch [114] and Apligraph [115] as graft therapies against DFU infection. However, more drugs are still required to achieve commercialization. Most of the drugs that are in clinical trials either fail or take a long time to become commercialized; thus, the failure of most of the novel therapies is creating chaos in the current healthcare market.
9. Everything Starts in the Gut: Gut–Skin Axis in Non-Healing Wounds
DFUs primarily involve the impaired regulation of the epidermal and endothelial layer of the peripheral foot region. The commensal or gut microbiome constantly controls the expression of growth factors (VEFG, EGF, and fibroblasts) and major skin remodeling factors (inflammatory cytokines). The epidermal keratinocytes, neutrophils, and macrophages stimulate wound healing in a coordinated manner, which ensures proper wound closure and restoring of the skin barrier [116]. The dietary pattern of an individual modulates the production of key regulators, which leads to homeostasis between the gut and the skin microbiota [117]. The efficient role of the gut is directly related to the enhanced production of innate immunity during the breakage of skin. This constant correlation between the gut and skin microbiome governs the immune-protective response that allows positive feedback for epithelial vascularization and angiogenesis [118] (Figure 2).
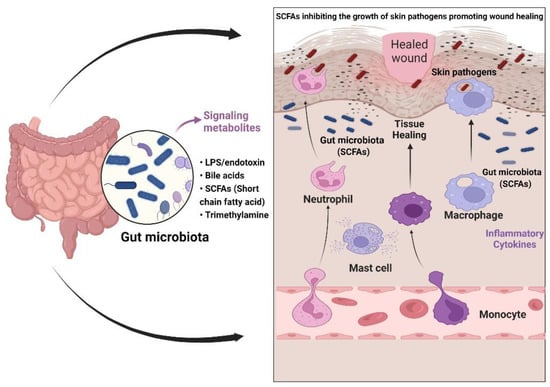
Figure 2.
Gut–skin axis in wound healing. The gut microbiota stimulates the production of short-chain fatty acids (SCFAs) and carbohydrate metabolites that regulate the skin’s commensal bacteria. Secondary metabolites such as SCFAs regulate anti-inflammatory properties (the production of innate immune cells and cytokines), causing skin commensal bacteria to mediate effective antibacterial action against skin pathogens.
9.1. Staphylococcus epidermidis and Propionibacterium spp.: Potential Skin Commensal Probiotics
A study conducted on fiber fermentation; i.e., propionic acid and butyrate in intestinal mucosa, influenced the metabolic role of skin microbiota. For instance, Propionibacterium spp., which has antibacterial activity against MRSA, supported coordinated behavior between gut and skin commensal bacteria [119]. A few other microbes such as Cutibacterium acne and S. epidermidis have also shown a beneficial effect in regulating the gut–skin axis. These microbes, through their bacteriocins and antimicrobial peptides, reduce skin inflammation by inducing innate immunity and keratinocyte movement, which encourages wound healing [120]. S. epidermidis enhanced the expression of IFN-γ, IL-17, and the CD8+-associated T-cell response, enhancing the innate barrier and inhibiting the pathogen invasion [121]. Another study found a direct correlation between the gut–skin crosstalk, supporting that S. epidermidis is involved in fine-tuning the T-cell response via IL-1 dependent downstream signaling for maintaining tissue health during skin homeostasis [122]. Similar to the Propionibacterium effect, S. epidermidis also utilizes short-fatty acid chains (SCFAs) in demonstrating its antibacterial effect against S. aureus [123].
9.2. Lactobacillus Species: A Superior Microbiota Regulating Gut–Skin Homeostasis
Lactobacillus is generally one set of microbes that have shown benefits against many skin infections; it is still being used as an alternative therapy against chronic infections [124]. Lactobacillus has a long-known activity against chronic infections such as burn wounds and has been used effectively against wound infections. Burn wounds are a case of chronic infection that occasionally involves graft rejection, which leads to high inflammation involving the secretion of proinflammatory cytokines and plasma proteins, thereby paving the way to a high bacterial burden and chronic biofilm formation. A classic example of L. plantarum on third-degree-burn wound patients also showed promising results in terms of parameters such as the local allergic reaction, smoothness of the skin, and non-recovery of Lactobacillus after 48 h of treatment. While considering burn wounds as severe skin infections, this study also showed a differential pattern of cytokine production compared to the proinflammatory cytokines that were produced by Gram-negative and Gram-positive pathogens. The acidic pH contributed to the fibroblast migration and tissue repair, which conclusively showed the cost-effective role of L. plantarum [125]. A study by Poutahidis et al. on L. reuteri [126] showed a marked increase in the wound-healing properties via upregulation of neuropeptide oxytocin, which has a potential role in vascularization via the gut–brain axis. These studies on Lactobacillus highlighted the role of good gut health flora in lowering the immunomodulatory action during wound closure.
10. Probiotics: Current Value in Host–Microbe Interactions
10.1. Probiotics: A Primitive Class of Microbes
There has been a constant debate regarding the defense mechanism of the human gut flora because bacterial colonization is too complex when it comes to beneficial or invasive microbes. A slight deviation in the microbiome equilibrium can cause serious detrimental effects on human health [127]. The relationship between gut microbial dysbiosis and diabetes is a long-standing debate that emphasizes the significant relationship between gut microbiota and glucose resistance during T2DM. According to a recent review, some gut microbes such as Ruminococcus, Fusobacterium, and Blautia are positively related to T2DM, while others such as Bifidobacterium and Lactobacillus are negatively related [128]. The crosstalk among various bacterial species during medical complications have always opened up the concept of “probiotics”, as they are the safest and most sustainable group of microbes that modulate positive effects in the gut–brain [129], gut–skin [130], and gut–bone [131] axes. Probiotics show a substantial positive effect on gastrointestinal metabolism and also contribute significantly to improvements in the host immune system [132]. Human studies have always defined probiotics as a strong modulator of the immune system that confer protection against numerous chronic diseases [133]. Universally, the major probiotic species are Lactobacillus and Bifidobacterium, which have numerous roles in human metabolic pathways. A study by Lim et al. [134] showed that treatment of high-fat-induced (HFD) hyperglycemic mice with L. sakei OK67 decreased the LPS levels in blood that was enhanced by HFD. The treatment decreased epididymal fat and suppressed the release of proinflammatory cytokines (TNF-α and IL-1) as well as the activation of NF-κB in the colon, highlighting the elevated expression of colon tight junction proteins and anti-inflammatory reactions in HFD mice. Another study on a diabetic mice model [135] showed that treatment with L. plantarum MTCC5690 and L. fermentum MTCC5689 improved insulin sensitivity and decreased the expression of the intestinal tight junction’s proteins (occludin and ZO-1), which in turn elevated the circulatory levels of LPS and proinflammatory markers (TNF-α, IL-6, and monocyte chemoattractant proteins (MCP-1)), demonstrating the antidiabetic property of Lactobacillus.
Streptococcus, Pedi coccus, Enterococcus, Lactococcus, and yeasts such as Saccharomyces boulardii, in addition to Lactobacillus spp., are important in regulating gut flora homeostasis [136]. Similarly, diabetes-related non-healing-wound studies have highlighted the role of commensal Lactobacillus spp. in the differential expression of cellular and metabolic genes that induced production of pro-healing cytokines (TNF-α, IL-12, and other non-inflammatory responses) [137]. Transcriptome profiling of L. acidophilus, L. casei, and L. rhamnosus showed the production of beneficial bacteriocins and secondary metabolites that can influence host metabolic regulation against various diseases [138]. In the current market, probiotics have a high yield, stability, and low-cost substrate requirement while posing no risk to the larger population [139]. Probiotics, as an effective immunomodulator, also hold great value in regulating the glycemic index of type 1 diabetes mellitus (T1DM) patients. T1DM is characterized by pancreatic beta cell damage, which results in abnormal insulin production in the regulation of blood glucose levels [140]. Continuous administration of L. johnsonii MH-68, Bifidobacterium animalis subsp. lactis CP-9, and L. salivarius subsp. salicinius AP-32 to patients with T1DM for 6 months resulted in decreased fasting blood glucose levels and HbA1c expression, as well as altered gut microbiota flora. The combination of insulin therapy and probiotic supplements in T1DM patients reduced the production of proinflammatory cytokines (IL-17, IL-18, and TNF-α), emphasizing the strong response of probiotics [141]. Another study on T2DM patients with continuous supplementation of multistrain probiotics for 12 weeks showed a decrease in glycated hemoglobin and improved the quality of life of the patients with a reduced body weight [142].
10.2. Basic Immunomodulatory Action of Probiotics
Probiotics have been categorized as the main factor in regulating the primary immune system and acquired immunity [143]. Probiotics generally activate dendritic cells and macrophages, which enhance the production of TGF-β, IL-4, IL-5, IL-12, and T-cell-mediated responses against pathogens [144]. The GI tract and intestinal flora are very specific for maintaining epithelial innate immunity. The presence of probiotics significantly activates humoral immune responses such as B-cell mediated immunoglobulin (Ig) production and enhances cytokine regulation. This triggers the production of anti-inflammatory cytokines (IL-10 and -12) and chemotaxis of innate immune cells (monocyte, NK cells, and dendritic cells), which induce immune homeostasis [145]. Probiotics also have a prominent role against viral or bacterial infections by modulating the NK-cell-associated T-cell response to establish antipathogenic activity [146]. A study conducted on the Gram-negative probiotic strain E. coli Nissle 1917 showed the production of NF-kB-mediated antibacterial peptide human β-defensin 2 (hBD-2), which promotes epithelial adhesion and activates innate cells (leukocytes and macrophages) [147]. Another study on Bifidobacterium showed enhanced cytokine responses and activation and maturation of dendritic cells against DSS-induced colitis. This highlighted the production of proinflammatory cytokines (IL-1β and TNF-α) that were further involved in CD4+ production via T-helper cells (TH), conferring an adaptive immune response [148].
11. Probiotic Effect in Acute and Chronic Biomarkers of Non-Healing DFUs
11.1. C-Reactive Proteins (CRPs)
CRPs are basic acute-phase proteins that play a role in various diabetic complications and are associated with high proinflammatory cytokines [149] (Figure 3). Currently, probiotic formulations that incorporate Bacillus coagulans (probiotic honey) [150] have a beneficial effect in reducing the CRP level in blood serum. Clinical trials have shown promising results of probiotics in reducing diabetic-related inflammation. For instance, we considered randomized trials on 523 participants with type 2 diabetes in which the participants were fed daily with probiotic yogurt from 2 to 12 weeks. Their inflammatory markers (CRPs, TNF-α, and IL-6) were monitored and significant decreases in the CRP level and IL-6 were found, suggesting the beneficial role of probiotics in lowering the acute phase proteins [151]. Similarly, another study conducted on patients suffering from diabetic nephropathy showed an increased production of creatinine and CRPs. These patients, who were supplemented with probiotic Lactobacillus and Bifidobacterium for 12 weeks, showed a significant reduction in the levels of CRPs, creatine, and IL-10, which highlighted the beneficial effects of probiotic supplements in reducing acute inflammation during diabetes [152].
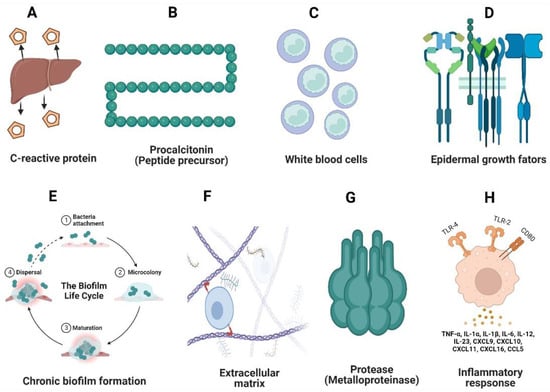
Figure 3.
Acute (A–D) and chronic (E–H) biomarkers during DFU infections. C-reactive proteins and procalcitonin are primary proteins whose differential expression indicates the onset of DFU infections (A,B). White blood cells (WBCs) typically express information about bacterial infections (C). Epidermal growth factors highlight lower neutrophil expression (D), indicating decreased re-epithelization during wound healing. Biofilm formation by opportunistic pathogens (E) and reductions in collagen and the extracellular matrix (F) due to the decreased keratinocyte movement (F) occur. Matrix metalloproteinase inhibits the production of anti-inflammatory cytokines, thereby impairing wound healing (G). The overproduction of proinflammatory cytokines leads to a decrease in macrophage-associated proteins and T-cell-mediated immune responses (H).
11.2. Procalcitonin
Procalcitonin is a hormonal peptide precursor of calcitonin that shows a higher expression during the acute phase of DFUs. The level of procalcitonin rises with the increased production of proinflammatory stimuli. Generally, these peptides are associated with bacterial infections and are used as a biomarker during sepsis conditions [153]. Studies conducted on hospital-acquired DFU infections have shown an increased production of procalcitonin biomarkers. The increased biomarker level highlighted other inflammatory markers (IL-3, IL-6, and IL-8), which showed a correlation between CRPs during DFU infections [154]. Constant research has found an effective role of probiotics in lowering the production of procalcitonin with a significant decrease in IL production. A study conducted on critically ill patients with sepsis showed a high production of IL-6, protein C, and procalcitonin as major inflammatory markers during acute infections. A constant supplementation of probiotics (Streptococcus, Bifidobacterium, and L. paracasei) resulted in a drastic reduction in the serum IL-6, protein C, and procalcitonin levels after 1 week, which also highlighted the beneficial effect of probiotics against acute-phase proteins [155].
11.3. White Blood Cell (WBC) Count
The major risk factor apart from CRPs, procalcitonin, and serum creatine involves the WBC count during the acute stage of diabetic foot infections. An elevated level of WBCs has been significantly associated with DFUs and is significantly affected by elevated levels of blood glucose [156]. The hematological and biochemical markers during diabetic-induced osteomyelitis also include the WBC count because variation in the WBC levels estimates the degree of a diabetic foot infection [154]. As previously described, probiotics have a major effect in reducing the acute-phase proteins and inflammatory marker levels. A recent study of probiotics on non-healing distal limb infections in equine species showed a significant decrease in the WBC count after topical administration of probiotics. This significant decrease in inflammation was accompanied by a 50% decrease in the WBC count in the wound area [157]. Another study conducted on healthy individuals who were supplemented with Lactobacillus and Bifidobacterium probiotics for 6 weeks showed a decreased production of leukocytes (WBCs) and proinflammatory cytokines (IL-6 and TNF-α), showing the important role of probiotics in maintaining healthy intestinal homeostasis [158].
11.4. Growth Factors
The process of inflammation during diabetic neuropathy leads to an alteration in growth factors, which leads to a decrease in the chemotaxis movement of neutrophils, thus reducing the angiogenesis and endothelial growth regeneration and causing increased susceptibility to foot infections [159]. Probiotics have been shown to have a significant role in increasing VEFF-mediated cell regeneration and a major role in enhancing keratinocyte production. Studies conducted on L. plantarum showed low cytotoxicity and increased production of VEGF in skin regeneration and fibroblast production, demonstrating the prominent role of probiotics in the enhancement of growth factors [160]. Lactobacillus has also shown effects on chemokine activation and movement; these chemokines have a major role in angiogenesis and help in the recruitment of inflammatory cells [161].
11.5. Chronic Biofilm Formation
In contrast to acute wounds, chronic DFU wounds consist of 60% biofilm-forming cells, clinical growth of aerobic microbes (P. aeruginosa and S. aureus), and anaerobic microbes in the deep part of the wound; i.e., osteomyelitis, and significantly resist the entry of antimicrobial agents, which causes a delay in wound healing [162]. Conventional therapies for controlling biofilms are not very effective, hence alternatives are required, including probiotics, which have been used since the early 1990s and have shown a potential role against cutaneous skin infections. Recently, probiotic strains (Bacillus subtilis KATMIRA and B. amyloliquefaciens) showed efficient activity against wound infections [163]. Similarly, a study conducted on L. fermentum against potent opportunistic pathogens (Pseudomonas and Staphylococcus) showed reductions in the biofilm-forming cells and in the production of virulence factors (pyocyanin and pyoverdine) [164].
11.6. Extracellular Matrix (ECM)
Normal wound healing involves collagen remodeling and degradation of fibril proteins by MMPs, but a DFU condition tends to overexpress the fibrils, resulting in distortions in ECM remodeling. Fluctuations in the levels of collagen and fibrinogen in blood plasma decreases elastin and fibronectin production in the tissue remodeling [165]. The damaged host ECM results in the movement of opportunistic pathogens by lowering the ECM binding and promoting virulence. Some probiotic species have shown an effective role in reinitiating production of collagen I and fibronectin [166]. A study conducted on L. casei showed its significant effects in ECM remodeling and fibronectin production by revealing the efficient movement of fibroblasts and keratinocytes for re-epithelization [167].
11.7. Matrix Metalloproteinase (MMP)
During delays in the wound-healing process, MMP-9 plays a role in the production of acute-phase proteins and proinflammatory cytokines. The imbalance between the ECM formation and high levels of MMP-2 and MMP-9 in macrophages inhibits the expression of anti-inflammatory factors such as TimP-1 and TimP-2, leading to healing dysbiosis [168]. Most studies have not shown the regulation of probiotic species in these MMP proteins during DFUs, but some studies on sepsis showed a significant decrease in MMP when Lactobacillus spp. was administered. A study conducted by Maghsood et al. [169] on cell-free supernatants (CFS) of L. acidophilus and L. rhamnosus showed a decreased expression of MMP-9 in human monocyte cells. This highlighted a significant increase in the cell-surface expression of CD147 and TimP-1 and TimP-2, which promoted anti-inflammatory properties. Another study on an ethanol extract from L. plantarum showed a significant decrease in MMP-9, IL-6, and TNF-α expression in streptozotocin-induced diabetic rats. The study showed the production of collagen fiber and extracellular matrix, showcasing wound healing and the positive effect of Lactobacillus in lowering the inflammatory response [170].
11.8. Proinflammatory Cytokine Response
The infiltration of cytokines is the major factor related to increased inflammation during a chronic DFU condition. The major process of healing is retarded by excessive proinflammatory cytokine secretion, macrophage polarization, tissue destruction, loss of angiogenesis, and bacteria-induced inflammation [171]. To overcome the immunomodulatory burst, probiotics have been constantly recognized as an efficient therapy. Various studies showed the downregulation of inflammatory pathways. For instance, L. reuteri extracts increased the expression of the inflammatory protein MMP-1 with downregulation of TGF-β signaling. This enhanced the proliferation of skin fibroblasts. In addition, L. reuteri extracts increased the expression of P13-AKT pathways, which play a prominent role in cell differentiation and apoptosis [172]. Another clinical study of DFU patients that used topical administration of L. plantarum showed a remarkable difference in terms of angiogenesis and vascular area of the wounds. In addition, the study showed M1 and M2 macrophage polarization in fibroblast proliferation and tissue remodeling [173].
12. Results
The search yielded 17 potentially relevant articles related to the role of probiotics in modulating acute and chronic markers of DFUs, 4 of which were randomized control trials, placebo trials, and single blinded multicenter studies. The randomized trial studies used probiotic strains such as L. acidophilus, Bifidobacterium, L. reuteri, and Streptococcus spp. to lower the expression of acute-phase markers such as C-reactive proteins and procalcitonin. In a clinical trial study, L. plantarum and Bifidobacterium reduced the white blood cell (WBC) count and proinflammatory markers, highlighting the efficacy of probiotic supplements. One systematic meta-analysis review also highlighted the role of yogurt consumption in lowering the acute-phase proteins by reducing the expression of IL-6, a proinflammatory marker. The remaining 11 articles involved in vivo and in vitro observational studies. The major probiotic studies related to acute and chronic conditions are listed in Table 2.
Table 2.
Features of probiotic studies associated with acute and chronic conditions.
13. Novel Prospects of Probiotic-Related Therapies against DFUs
Currently, few probiotic therapies have shown an efficient response against non-healing wound ulcers (diabetic and non-diabetic). Non-healing diabetic wounds showed a decreased expression of vascularization due to reduced angiogenesis and insulin metabolism. The few probiotics mentioned in Table 3 highlight the efficient response against intrinsic mediators of non-healing diabetes-associated foot ulcers.
Table 3.
Probiotic responses against DFU infections.
Apart from the above-mentioned probiotic supplements (Table 2), few novel avenues can be utilized to increase the efficiency pf probiotic strains against DFU infections. Some of the novel prospects are highlighted below.
13.1. Probiotic Encapsulation
To attain high efficiency at the wound site, probiotics need to be biocompatible and non-toxic. The heterogeneous nature of chronic wounds involves polymicrobial flora that constantly trigger the immunogenic responses. The fine biocompatibility and non-toxicity of probiotics can be maintained by using various encapsulating agents such as polymeric substances (e.g., chitosan, alginate, and hydrogels). These agents have proven non-toxicity and an environmentally friendly nature that could protect probiotics from a harmful pathogenic environment [179]. A recent study conducted on probiotic encapsulation with antibiotics in an alginate-mediated encapsulating agent showed an increased potency of the tobramycin antibiotic in the eradication of MRSA strains [180]. Another study on delivering probiotics found that a potent self-assembling coated system composed of tannic acid and poloxamer showed strong intestinal adhesion, ROS production, and higher deleterious effects against DSS-induced-colitis mice [181]. Similarly, heparin-based hydrogels also showed a potent effect against inflammatory markers of wound healing by regulating M1 and M2 macrophage polarization [182] and reducing the expression of proinflammatory markers (IL-1β, TNF-α, and NF-kB). These potent regenerative concepts can be applied to the chronic DFU situation via co-administration of empirical antibiotics with a probiotic in an enclosed shell, which will limit the effects of harmful bacteria on probiotics and will also provide the benefit of targeted therapy against polymicrobial infection.
13.2. Prebiotics and Synbiotics: Nanoformulations
Prebiotics is a well-known concept in which the intake of dietary fiber invokes gut microbes to produce beneficial factors, while synbiotics is a concept in which both a probiotic and prebiotic are combined [183]. Both concepts involve the positive regulation of the host microbiota in that they showed beneficial effects in regulating intestinal homeostasis. A study with Lycium barbarum (herb)-derived polysaccharides on mice fecal microbiota showed a significant increase in the growth of Lactobacillus and Bifidobacterium, thereby modulating the intestinal microbial communities. Moreover, the inflammatory response was also varied significantly by changing IL-6 and TNF-β production [184]. Synbiotic studies involving the use of probiotic Bacillus coagulans [185] and a prebiotic grape extract with a Lactobacillus mixture [186] showed prominently decreased inflammatory markers in caco2 cells and decreased inflammation in diabetic patients. Non-digestible carbohydrate pullulan used as an NP-like prebiotic supplement with L. plantarum showed increased production of antimicrobial peptide that could eradicate Gram-negative pathogens [187]. The potency of nanotherapies with prebiotics and synbiotics can be applied to DFU infections because the gut microbiota play a major role in modulating skin commensal bacteria to produce antimicrobial peptides against pathogens. Therefore, various non-digested fibers should be screened and used as sources for nanoformulations with probiotic supplements to modulate the production of the inflammatory response during chronic conditions.
13.3. Probiotic-Derived Biogenic Nanoparticles (NPs)
Biogenic materials involve plants, microbes, and fungi; currently, these microbes are being used as a potent source to produce NPs. These NPs provide an edge over conventional NP synthesis due to their lower toxicity and high durability against various medical treatments [188]. Microbes are considered to be capable producers of NPs using elements such as silver (Ag), selenium (Se), gold (Au), and copper (Cu), which have enhanced therapeutic values. To this end, environmentally friendly and economic probiotics such as L. paraplantarum and L. crustorum, L. delbrukii, and L. plantarum are extensively being used to treat various pathological conditions (antibacterial, antioxidant, or even chronic biofilm conditions) [189]. The use of biogenic NPs using probiotic microbes showed a beneficial effect on chronic DFU infections because the above NPs could strongly produce ROS that efficiently degraded cell walls, could be synergistically used with other antimicrobial compounds to enhance immune cell activation, and could act as an anti-biofilm agent. NPs can overcome the limits of chronic ulcer biofilm, which makes it difficult for antimicrobial compounds to enter the wound site and results in a delay in wound healing (Figure 4).
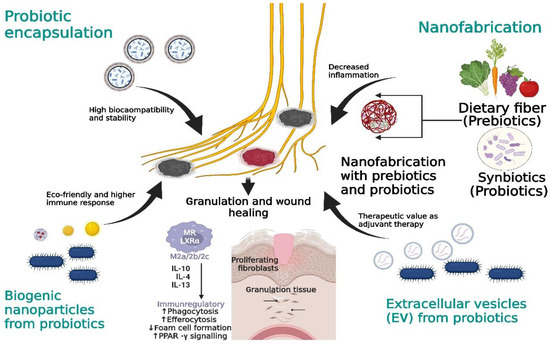
Figure 4.
Novel approaches to target DFU prognosis. Probiotic encapsulation ensures the probiotics’ long-term stability against chronic DFUs. Dietary fiber will be used in nanofabrication with prebiotics and probiotics for efficient probiotic regulation. Biogenic NPs boost immune activation; probiotic-derived EVs can be used as an alternative therapeutic delivery agent.
13.4. Probiotic-Derived Extracellular Vesicles (EVs)
Another class of nanotherapeutics involves extracellular vesicles (EVs), which are the membrane vesicles that can carry biogenic compounds including lipids, proteins, and nucleic acids. These vesicles have recently gained much value due to their intrinsic antimicrobial activity and their use as a successful delivery agent against various chronic illnesses [190]. A study using EVs derived from Propionibacterium freundenreichii showed enhanced immunogenic activity in HT-29 human epithelial cells. In this study, NF-kB and IL-8 were drastically reduced as compared to the non-treated cells with EVs, which supported the anti-inflammatory response of EVs against chronic illness [191]. Other studies of probiotic-derived EVs showed their potential role as adjuvants [192] in modulating the proliferation of epithelial growth factors during DSS-induced colitis [193]. Based on the above results, it is obvious that probiotic-derived vesicles have a potential role in reducing acute and chronic inflammation, since most of the inflammatory cytokines and biomarkers of IL-8, NF-kB, and TNF-α are also the prominent markers in DFU infections. Therefore, EVs derived from probiotics can be utilized as a futuristic therapeutic material in the betterment of the healing prognosis of DFU wounds.
14. Management of Acute and Chronic DFU Conditions via Probiotic Remodeling
The fundamental issue behind the global glycemic index is the lifelong risk of acquiring lower-limb DFUs. The current situation is disastrous because lower limbs are either amputated or patients suffer from emotional and physical stress as a result of exorbitant healthcare costs. Foot deformity due to a DFU is a progressive disorder that transforms the disorder into a life-threatening risk in due course. The ongoing quest for efficient technologies or therapies to assess DFU wounds has shown a chronicity of such wounds that encompasses a variety of pathological problems that require effective care [194]. Loss of sensation or persistent pressure in the limb region requires adequate foot care management as well as effective therapies to combat the resulting issues with a non-healing wound. The margin between the acute and chronic situations in a DFU is so narrow that even a subcutaneous hemorrhage or simple crack in the foot might result in a chronic gangrene situation. The complexity of DFU prognosis often requires proper medical practices such as ulcer classification or grading. The evaluation system provides both sufficient information to clinicians and comprehensive knowledge regarding the entire history of the foot complications of patients [195]. Sufficient data from the wound morphology provide evidence that may be utilized when prescribing medical facilities to the patient.
Acute and chronic diseases are generally discrete phases that entail the control of numerous parameters such as the blood serum level, biomarker presence, and inflammatory cytokines. The acute stage is characterized by the normal degradation of growth factors, which deregulate the vascularization process in normal healing. This causes systemic inflammatory responses that involve macrophages and neutrophils, resulting in decreased production of anti-inflammatory cytokines and constant tissue hypoxia in the peripheral wound region. Normal or acute healing takes 2 to 3 months and includes cell mitosis and tissue granularization, but constant and recurrent foot trauma results in a chronic condition of the foot region that includes a high inflammatory response involving an innate immune response and stimulation of proinflammatory cytokines. This makes the foot region more susceptible to infections. The high glycemic index also causes vascular neuropathy, which in turn causes a lack of angiogenesis, making the foot region more susceptible to persistent infections, which due to the formation of biofilm micro-communities in later stages, slows down the wound-healing process by more than 8 to 10 months. Out of all the factors mentioned here, the immune response plays a vital role in a significant delay of the wound-healing process because it is a host’s immune response that generally activates the proinflammatory modulators that cause immunogenic bursts at the site of infection.
The dysregulated response during DFUs involves the production of acute-phase proteins, WBCs, and other serum proteins that provide the primary signs of infection. Then, inflammatory cytokines come into the picture that involve the significant upregulation and downregulation of immunoregulators. Chronic DFUs primarily involve the dysregulation of neutrophils and macrophages, which initiates the downregulation of dendritic cells (MCP-1 and IFN-γ) and MMP-9. The imbalance in macrophages causes a decrease in the growth factors (VEGF and EGF) and tissue remodeling of collagen and fibronectin. This regulates the movement of proinflammatory cytokines (IL-1, IL-1β, and TGF-α), which increases the infection rate of the region and dysregulates the T-cells’ maturation. The cell-mediated immunity is hampered by a decreased, restricted movement of T-regulatory cells (Treg) and TH to the site of the infection. Constant production of chemokines and cytokines at the site of the wound lowers the epithelial remodeling and diminishes the vascularization property of keratinocytes, thereby rupturing the endothelial lining, which certainly paves the way for pathogens to enter. These pathogens secrete virulence factors that inhibit the normal healing process of the wound and involve the prognosis of tissue necrosis and a low oxygen state at the site of infection, causing it to reach the gangrenous state or the amputation stage.
Regarding the management of wound ulcer situations, various novel therapies that directly target the immune responses during DFU infections, such as therapies using fibroblasts, macrophage activation, stem cell or MMP inhibitors, and mAbs, have shown a drastic reduction in the proinflammatory mediators but have not been efficient in terms of longevity and static efficient responses. The major drawbacks involve a shorter half-life, repeated administration, degradation via a proteolytic environment of the wound, high cell death, expensiveness, hydrolysis at a low pH, limited downstream processing, renal clearance, and a high production cost. Still, there are FDA-approved drugs that have shown potent activity in lowering inflammatory responses, but there should be a continuous search for more efficient and better therapies that can regulate the host metabolism by lowering chronic malignancies. In the quest to find efficient therapies, researchers found a constant relationship between the microbe and the host cell response. However, a decade has passed and there have been a substantial amount of data that showed the influence of environmental microflora on human health; several microbes have been identified as pathogens, including basic categories such as Staphylococcus, Streptococcus, Bacillus, Pseudomonas, Klebsiella, and other fungal bodies. There is one class that has always had a beneficial role in host–microbe interactions; i.e., “probiotics.” The constant regulation between the host gut microbiota and skin has shown a consistent homeostasis relationship between the dermal and mucosal microbiota. The regulation of various metabolites (short-chain fatty acids, exopolysaccharides, lipids, and other metabolites) directly helps to maintain the protective microbiota in cutaneous and subcutaneous layers against pathogenic interactions.
Probiotics have been known to provide a protective barrier against various pathogenic species. These microbes constantly regulate the gut–skin metabolism to provide essential metabolites such as bacteriocins and anti-inflammatory molecules, which modulate the innate immune response against DFUs. Most probiotics degrade the proinflammatory chemokines (CXCL10), which have a role in stimulating inflammatory cells. Some species have been found to regulate the epithelial lining by regulating protein kinase activities. L. plantarum provided a maximum benefit when used in a probiotic cocktail with Bifidobacterium because it regulated immunoglobulin activity by establishing a sustainable relationship with the tissue repair [196]. In the current scenario, probiotic drinks have shown numerous benefits in regulating all three primary immune metabolisms; i.e., innate, adaptive, and acquired immunity, by constantly regulating the leukocyte, macrophage, and neutrophil chemotaxis [197]. A delay in wound healing is a chronic condition that is affected not only by neuropathy or vasculopathy, but also by the extent of infection. The high bacterial density at the DFU site plays an important role in forming biofilm communities. Constant intercommunicating signals (quorum sensing and siderophore production) increase the chances of forming compact microbiota, which often results in hypoxia situations and reduced blood flow, ultimately forming gangrene. These pathogens act as natural barriers against antimicrobial compounds and evade the host’s natural immune mechanisms by eradicating the surrounding tissue, which leads to inflammation and a delay in the wound-healing process. Therefore, these tandem roles of the host innate response and chronic stimulation increase the chances of amputations or long-term infections. Therapies have shown effectivity to a great extent, but consistent deterioration of therapies might complicate the treatment efficiency; therefore, probiotics or sustainable dietary intake will not only moderately affect the glycemia index (diabetes), but also will regulate the innate immune response by limiting other risk factors such as neuropathy, nephropathy, or retinopathy. Because most of the inflammatory cytokines or proinflammatory markers have a prominent role in maintaining the gut flora metabolism in normal wound-healing procedures, the same immunomodulators can have a detrimental effect during chronic dysbiosis.
15. Limitations
The use of entire microbes as probiotics has beneficial effects in reducing systemic inflammation by maintaining gut–skin–brain homeostasis. However, effectively defining the microbiome as a “probiotic” still requires an extensive number of supporting explanations in terms of its use in a larger population and clinical outcomes. Currently, trial-and-error-based strategies are being explored; therefore, medical interventions, more detailed analyses, and proper clinical trials are required to generate more user-friendly data with conclusive knowledge related to dosage values. Normally, the source of probiotics varies from Gram-positive to Gram-negative bacteria, and environmental microbes acclimatize themselves according to the respective niches. Therefore, in some instances, probiotic ingestion might allow probiotic species to reshuffle their entire genetic makeup, which can cause detrimental effects on patients; the impact on critically immunocompromised patients would be especially severe. Apart from this, in the era of antibiotic resistance, parameters such as drug usage, the potentiality of drugs, and the prescription of drugs ultimately influence the microbial flora of the human system. The major problems of these probiotic species from friend to foe involve horizontal gene transfer mechanisms; such mechanisms would limit the use of probiotic products. Certain examples such as S. epidermidis and Propionibacterium spp. have shown numerous benefits in terms of the production of secondary metabolites and fatty acids, which constantly limit the growth of other dermal pathogens. However, a slight imbalance in the gut–skin axis may cause them to become detrimental to the host’s microflora.
16. Conclusions
16.1. Current Progress
Prebiotics and synbiotics are important in regulating an individual’s intestinal mucosa; products combining dietary fibers with probiotics have been shown to modulate intestinal gut flora, which in turn can reduce the glycemic index and lipid metabolism [198]. The commercialized probiotic product, which includes a cocktail of Lactobacillus spp. and Bifidobacterium spp. in a capsule, provides nearly all of the benefits in modulating an effective immune response against chronic DFU infections. A novel concept known as “postbiotics” is currently being investigated as a replacement for probiotics. Postbiotics are metabolic products released by probiotic organisms that have several advantages, including a high scale-up, prolonged storage viability, and potent immunomodulatory properties. Similarly, a concept known as “paraprobiotics”, which involves dead or inactive probiotic cells, has demonstrated beneficial anti-inflammatory and anti-biofilm properties, as well as an improved gut barrier function and immunomodulatory properties, suggesting their use in reducing chronic wound infections [199]. These concepts may lead to effective gut–skin homeostasis. However, large-scale clinical trials are required before they can be effectively rolled out in the market.
16.2. Future Perspectives
There is a need for metagenomic approaches to identify as many probiotics or beneficial microbes as possible because such microbes have the potency to induce our intestinal homeostasis. The DFU is a severe metabolic disorder that is combined with dysbiosis of the neural system (motor neuropathy and vascular neuropathy); its subsequent progression leads to a chronic situation of the wound. Probiotics generally have much potential in regulating wounds at the acute and chronic stages. Therefore, commercially available or newly discovered beneficial microbes can be used as resources for bioactive materials such as biogenic molecules or EVs to be combined with DFU-efficient NPs, which can be alternatively used with hydrogel dressings or can be used as a synergistic molecule with antibiotics in biofilm-related DFUs. Finally, this review on probiotics represents an interesting area of research in treating the acute and chronic conditions of DFUs and provides an accurate assessment of the manner in which they regulate gut–skin homeostasis and how current advancements in prebiotics and synbiotics can increase the potency of probiotics against DFU infections.
Author Contributions
P.S.: conceptualization, writing—original draft; T.S.: writing—original draft; K.S.: writing—original draft; K.-s.K.: conceptualization, supervision, funding acquisition, writing—reviewing and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This research was financially supported by the 2022 Post-Doc. Development Program of Pusan National University and a National Research Foundation of Korea (NRF) grant funded by the Korean government (MSIT) (grant number: NRF-2021R1A2C1007413).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
All figures in this manuscript were prepared by utilizing BioRender (www.biorender.com) (accessed on 7 November 2022).
Conflicts of Interest
The authors declare that there are no conflict of interest.
References
- Feldman, E.L.; Callaghan, B.C.; Pop-Busui, R.; Zochodne, D.W.; Wright, D.E.; Bennett, D.L.; Bril, V.; Russell, J.W.; Viswanathan, V. Diabetic neuropathy. Nat. Rev. Dis. Primers 2019, 5, 41. [Google Scholar] [CrossRef] [PubMed]
- Ince, P.; Abbas, Z.G.; Lutale, J.K.; Basit, A.; Ali, S.M.; Chohan, F.; Morbach, S.; Mollenberg, J.; Game, F.L.; Jeffcoate, W.J. Use of the SINBAD classification system and score in comparing outcome of foot ulcer management on three continents. Diabetes Care 2008, 31, 964–967. [Google Scholar] [CrossRef]
- Shi, L.; Wei, H.; Zhang, T.; Li, Z.; Chi, X.; Liu, D.; Chang, D.; Zhang, Y.; Wang, X.; Zhao, Q. A potent weighted risk model for evaluating the occurrence and severity of diabetic foot ulcers. Diabetol. Metab. Syndr 2021, 13, 92. [Google Scholar] [CrossRef]
- Karri, V.V.; Kuppusamy, G.; Talluri, S.V.; Yamjala, K.; Mannemala, S.S.; Malayandi, R. Current and emerging therapies in the management of diabetic foot ulcers. Curr. Med. Res. Opin. 2016, 32, 519–542. [Google Scholar] [CrossRef]
- Jodheea-Jutton, A.; Hindocha, S.; Bhaw-Luximon, A. Health economics of diabetic foot ulcer and recent trends to accelerate treatment. Foot 2022, 52, 101909. [Google Scholar] [CrossRef] [PubMed]
- Coates, M.; Lee, M.J.; Norton, D.; MacLeod, A.S. The Skin and Intestinal Microbiota and Their Specific Innate Immune Systems. Front. Immunol. 2019, 10, 2950. [Google Scholar] [CrossRef]
- Kawai, K.; Kamochi, R.; Oiki, S.; Murata, K.; Hashimoto, W. Probiotics in human gut microbiota can degrade host glycosaminoglycans. Sci. Rep. 2018, 8, 10674. [Google Scholar] [CrossRef]
- Zuo, F.; Marcotte, H. Advancing mechanistic understanding and bioengineering of probiotic lactobacilli and bifidobacteria by genome editing. Curr. Opin. Biotechnol. 2021, 70, 75–82. [Google Scholar] [CrossRef]
- Sun, H.; Saeedi, P.; Karuranga, S.; Pinkepank, M.; Ogurtsova, K.; Duncan, B.B.; Stein, C.; Basit, A.; Chan, J.C.N.; Mbanya, J.C.; et al. IDF Diabetes Atlas: Global, regional and country-level diabetes prevalence estimates for 2021 and projections for 2045. Diabetes Res. Clin. Pr. 2022, 183, 109119. [Google Scholar] [CrossRef]
- Magliano, D.J.; Boyko, E.J. IDF Diabetes Atlas, 10th ed.; International Diabetes Federation: Brussels, Belgium, 2021; Available online: https://www.idf.org/e-library/epidemiology-research/diabetes-atlas.html (accessed on 18 October 2022).
- Sato, J.; Kanazawa, A.; Ikeda, F.; Yoshihara, T.; Goto, H.; Abe, H.; Komiya, K.; Kawaguchi, M.; Shimizu, T.; Ogihara, T.; et al. Gut dysbiosis and detection of “live gut bacteria” in blood of Japanese patients with type 2 diabetes. Diabetes Care 2014, 37, 2343–2350. [Google Scholar] [CrossRef] [PubMed]
- Gerard, C.; Vidal, H. Impact of Gut Microbiota on Host Glycemic Control. Front. Endocrinol. 2019, 10, 29. [Google Scholar] [CrossRef]
- He, X.; Sun, J.; Liu, C.; Yu, X.; Li, H.; Zhang, W.; Li, Y.; Geng, Y.; Wang, Z. Compositional Alterations of Gut Microbiota in Patients with Diabetic Kidney Disease and Type 2 Diabetes Mellitus. Diabetes Metab. Syndr. Obes. 2022, 15, 755–765. [Google Scholar] [CrossRef] [PubMed]
- Global Report on Diabetes; World Health Organization: Geneva, Switzerland, 2016.
- Pourkazemi, A.; Ghanbari, A.; Khojamli, M.; Balo, H.; Hemmati, H.; Jafaryparvar, Z.; Motamed, B. Diabetic foot care: Knowledge and practice. BMC Endocr. Disord. 2020, 20, 40. [Google Scholar] [CrossRef]
- Jiang, M.; Gan, F.; Gan, M.; Deng, H.; Chen, X.; Yuan, X.; Huang, D.; Liu, S.; Qin, B.; Wei, Y.; et al. Predicting the Risk of Diabetic Foot Ulcers From Diabetics With Dysmetabolism: A Retrospective Clinical Trial. Front Endocrinol. 2022, 13, 929864. [Google Scholar] [CrossRef]
- Armstrong, D.G.; Boulton, A.J.M.; Bus, S.A. Diabetic Foot Ulcers and Their Recurrence. N. Engl. J. Med. 2017, 376, 2367–2375. [Google Scholar] [CrossRef] [PubMed]
- Okonkwo, U.A.; Chen, L.; Ma, D.; Haywood, V.A.; Barakat, M.; Urao, N.; DiPietro, L.A. Compromised angiogenesis and vascular Integrity in impaired diabetic wound healing. PLoS ONE 2020, 15, e0231962. [Google Scholar] [CrossRef] [PubMed]
- Morbach, S.; Furchert, H.; Groblinghoff, U.; Hoffmeier, H.; Kersten, K.; Klauke, G.T.; Klemp, U.; Roden, T.; Icks, A.; Haastert, B.; et al. Long-term prognosis of diabetic foot patients and their limbs: Amputation and death over the course of a decade. Diabetes Care 2012, 35, 2021–2027. [Google Scholar] [CrossRef]
- Ndosi, M.; Wright-Hughes, A.; Brown, S.; Backhouse, M.; Lipsky, B.A.; Bhogal, M.; Reynolds, C.; Vowden, P.; Jude, E.B.; Nixon, J.; et al. Prognosis of the infected diabetic foot ulcer: A 12-month prospective observational study. Diabet. Med. 2018, 35, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Song, J.F.; Zhang, L.; Li, X. Analysis of risk factors for multidrug-resistant organisms in diabetic foot infection. BMC Endocr. Disord. 2022, 22, 46. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, A.; Berkache, M.; Morency-Potvin, P.; Juneau, D.; Koenig, M.; Bourduas, K.; Freire, V. Diabetic foot infections: How to investigate more efficiently? A retrospective study in a quaternary university center. Insights Imaging 2022, 13, 88. [Google Scholar] [CrossRef] [PubMed]
- Game, F. Classification of diabetic foot ulcers. Diabetes Metab. Res. Rev. 2016, 32 (Suppl. 1), 186–194. [Google Scholar] [CrossRef]
- Santema, T.B.; Lenselink, E.A.; Balm, R.; Ubbink, D.T. Comparing the Meggitt-Wagner and the University of Texas wound classification systems for diabetic foot ulcers: Inter-observer analyses. Int. Wound J. 2016, 13, 1137–1141. [Google Scholar] [CrossRef] [PubMed]
- Karthikesalingam, A.; Holt, P.J.; Moxey, P.; Jones, K.G.; Thompson, M.M.; Hinchliffe, R.J. A systematic review of scoring systems for diabetic foot ulcers. Diabet. Med. 2010, 27, 544–549. [Google Scholar] [CrossRef] [PubMed]
- Oyibo, S.O.; Jude, E.B.; Tarawneh, I.; Nguyen, H.C.; Armstrong, D.G.; Harkless, L.B.; Boulton, A.J.M. The effects of ulcer size and site, patient’s age, sex and type and duration of diabetes on the outcome of diabetic foot ulcers. Diabet. Med. 2001, 18, 8. [Google Scholar] [CrossRef] [PubMed]
- Bravo-Molina, A.; Linares-Palomino, J.P.; Vera-Arroyo, B.; Salmeron-Febres, L.M.; Ros-Die, E. Inter-observer agreement of the Wagner, University of Texas and PEDIS classification systems for the diabetic foot syndrome. Foot Ankle Surg. 2018, 24, 60–64. [Google Scholar] [CrossRef]
- Forsythe, R.O.; Ozdemir, B.A.; Chemla, E.S.; Jones, K.G.; Hinchliffe, R.J. Interobserver Reliability of Three Validated Scoring Systems in the Assessment of Diabetic Foot Ulcers. Int. J. Low Extrem. Wounds 2016, 15, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Xie, T.; Cao, Y.; Wu, M.; Yu, L.; Lu, S.; Xu, G.; Hu, J.; Ruan, H. Comparison of two classification systems in predicting the outcome of diabetic foot ulcers: The Wagner grade and the Saint Elian Wound score systems. Wound Repair Regen 2015, 23, 379–385. [Google Scholar] [CrossRef]
- Monteiro-Soares, M.; Russell, D.; Boyko, E.J.; Jeffcoate, W.J.; Mills, J.L.; Morbach, S.; Game, F. IWGDF Guidelines on the Classification of Diabetic Foot Ulcers. Diabetes Metab. Res. Rev. 2019, 36 (Suppl. 1), e3273. [Google Scholar]
- Pastar, I.; Stojadinovic, O.; Yin, N.C.; Ramirez, H.; Nusbaum, A.G.; Sawaya, A.; Patel, S.B.; Khalid, L.; Isseroff, R.R.; Tomic-Canic, M. Epithelialization in Wound Healing: A Comprehensive Review. Adv. Wound Care 2014, 3, 445–464. [Google Scholar] [CrossRef]
- Volmer-Thole, M.; Lobmann, R. Neuropathy and Diabetic Foot Syndrome. Int. J. Mol. Sci. 2016, 17, 917. [Google Scholar] [CrossRef]
- Amin, N.; Doupis, J. Diabetic foot disease: From the evaluation of the "foot at risk" to the novel diabetic ulcer treatment modalities. World J. Diabetes 2016, 7, 153–164. [Google Scholar] [CrossRef]
- Nishide, K.; Nagase, T.; Oba, M.; Oe, M.; Ohashi, Y.; Iizaka, S.; Nakagami, G.; Kadowaki, T.; Sanada, H. Ultrasonographic and thermographic screening for latent inflammation in diabetic foot callus. Diabetes Res. Clin. Pr. 2009, 85, 304–309. [Google Scholar] [CrossRef]
- Cheng, Y.; Zu, P.; Zhao, J.; Shi, L.; Shi, H.; Zhang, M.; Wang, A. Differences in initial versus recurrent diabetic foot ulcers at a specialized tertiary diabetic foot care center in China. J. Int. Med. Res. 2021, 49, 300060520987398. [Google Scholar] [CrossRef] [PubMed]
- Petrova, N.L.; Edmonds, M.E. Acute Charcot neuro-osteoarthropathy. Diabetes Metab. Res. Rev. 2016, 32 (Suppl. 1), 281–286. [Google Scholar] [CrossRef]
- Jansen, R.B.; Jorgensen, B.; Holstein, P.E.; Moller, K.K.; Svendsen, O.L. Mortality and complications after treatment of acute diabetic Charcot foot. J. Diabetes Complicat. 2018, 32, 1141–1147. [Google Scholar] [CrossRef] [PubMed]
- Weigelt, C.; Rose, B.; Poschen, U.; Ziegler, D.; Friese, G.; Kempf, K.; Koenig, W.; Martin, S.; Herder, C. Immune mediators in patients with acute diabetic foot syndrome. Diabetes Care 2009, 32, 1491–1496. [Google Scholar] [CrossRef] [PubMed]
- Korkmaz, P.; Kocak, H.; Onbasi, K.; Bicici, P.; Ozmen, A.; Uyar, C.; Ozatag, D.M. The Role of Serum Procalcitonin, Interleukin-6, and Fibrinogen Levels in Differential Diagnosis of Diabetic Foot Ulcer Infection. J. Diabetes Res. 2018, 2018, 7104352. [Google Scholar] [CrossRef] [PubMed]
- Muller, S.; Martin, S.; Koenig, W.; Hanifi-Moghaddam, P.; Rathmann, W.; Haastert, B.; Giani, G.; Illig, T.; Thorand, B.; Kolb, H. Impaired glucose tolerance is associated with increased serum concentrations of interleukin 6 and co-regulated acute-phase proteins but not TNF-alpha or its receptors. Diabetologia 2002, 45, 805–812. [Google Scholar] [CrossRef] [PubMed]
- Holl, J.; Kowalewski, C.; Zimek, Z.; Fiedor, P.; Kaminski, A.; Oldak, T.; Moniuszko, M.; Eljaszewicz, A. Chronic Diabetic Wounds and Their Treatment with Skin Substitutes. Cells 2021, 10, 655. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, Y.; Deng, J.; Li, W.; Nie, X. Fibroblast Growth Factor in Diabetic Foot Ulcer: Progress and Therapeutic Prospects. Front Endocrinol. 2021, 12, 744868. [Google Scholar] [CrossRef]
- Ayuk, S.M.; Abrahamse, H.; Houreld, N.N. The Role of Matrix Metalloproteinases in Diabetic Wound Healing in relation to Photobiomodulation. J. Diabetes Res. 2016, 2016, 2897656. [Google Scholar] [CrossRef] [PubMed]
- Jouhar, L.; Jaafar, R.F.; Nasreddine, R.; Itani, O.; Haddad, F.; Rizk, N.; Hoballah, J.J. Microbiological profile and antimicrobial resistance among diabetic foot infections in Lebanon. Int. Wound J. 2020, 17, 1764–1773. [Google Scholar] [CrossRef]
- Baig, M.S.; Banu, A.; Zehravi, M.; Rana, R.; Burle, S.S.; Khan, S.L.; Islam, F.; Siddiqui, F.A.; Massoud, E.E.S.; Rahman, M.H.; et al. An Overview of Diabetic Foot Ulcers and Associated Problems with Special Emphasis on Treatments with Antimicrobials. Life 2022, 12, 1054. [Google Scholar] [CrossRef] [PubMed]
- Chu, Y.; Wang, C.; Zhang, J.; Wang, P.; Xu, J.; Ding, M.; Li, X.; Hou, X.; Feng, S.; Li, X. Can We Stop Antibiotic Therapy When Signs and Symptoms Have Resolved in Diabetic Foot Infection Patients? Int. J. Low Extrem. Wounds 2015, 14, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Aragon-Sanchez, J.; Lazaro-Martinez, J.L.; Pulido-Duque, J.; Maynar, M. From the diabetic foot ulcer and beyond: How do foot infections spread in patients with diabetes? Diabet. Foot Ankle 2012, 3, 18693. [Google Scholar] [CrossRef]
- Zhang, X.; Yang, X.; Chen, Y.; Wang, G.; Ding, P.; Zhao, Z.; Bi, H. Clinical study on orthopaedic treatment of chronic osteomyelitis with soft tissue defect in adults. Int. Wound J. 2022, 19, 1349–1356. [Google Scholar] [CrossRef]
- Sadeghpour Heravi, F.; Zakrzewski, M.; Vickery, K.D.; Armstrong, D.; Hu, H. Bacterial Diversity of Diabetic Foot Ulcers: Current Status and Future Prospectives. J. Clin. Med. 2019, 8, 1935. [Google Scholar] [CrossRef]
- Buch, P.J.; Chai, Y.; Goluch, E.D. Bacterial chatter in chronic wound infections. Wound Repair Regen. 2021, 29, 106–116. [Google Scholar] [CrossRef]
- Buch, P.J.; Chai, Y.; Goluch, E.D. Treating Polymicrobial Infections in Chronic Diabetic Wounds. Clin. Microbiol. Rev. 2019, 32, e00091-18. [Google Scholar] [CrossRef]
- Percival, S.L.; Malone, M.; Mayer, D.; Salisbury, A.M.; Schultz, G. Role of anaerobes in polymicrobial communities and biofilms complicating diabetic foot ulcers. Int. Wound J. 2018, 15, 776–782. [Google Scholar] [CrossRef]
- Nussbaum, S.R.; Carter, M.J.; Fife, C.E.; DaVanzo, J.; Haught, R.; Nusgart, M.; Cartwright, D. An Economic Evaluation of the Impact, Cost, and Medicare Policy Implications of Chronic Nonhealing Wounds. Value Health 2018, 21, 27–32. [Google Scholar] [CrossRef]
- Srivastava, P.; Sivashanmugam, K. Combinatorial Drug Therapy for Controlling Pseudomonas aeruginosa and Its Association with Chronic Condition of Diabetic Foot Ulcer. Int. J. Low Extrem. Wounds 2020, 19, 7–20. [Google Scholar] [CrossRef] [PubMed]
- Johani, K.; Malone, M.; Jensen, S.; Gosbell, I.; Dickson, H.; Hu, H.; Vickery, K. Microscopy visualisation confirms multi-species biofilms are ubiquitous in diabetic foot ulcers. Int. Wound J. 2017, 14, 1160–1169. [Google Scholar] [CrossRef] [PubMed]
- Dunnill, C.; Patton, T.; Brennan, J.; Barrett, J.; Dryden, M.; Cooke, J.; Leaper, D.; Georgopoulos, N.T. Reactive oxygen species (ROS) and wound healing: The functional role of ROS and emerging ROS-modulating technologies for augmentation of the healing process. Int. Wound J. 2017, 14, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.; Srivastava, S.; Singh, M.R.; Singh, D. Mechanistic insight into diabetic wounds: Pathogenesis, molecular targets and treatment strategies to pace wound healing. Biomed. Pharm. 2019, 112, 108615. [Google Scholar] [CrossRef] [PubMed]
- Mouritzen, M.V.; Petkovic, M.; Qvist, K.; Poulsen, S.S.; Alarico, S.; Leal, E.C.; Dalgaard, L.T.; Empadinhas, N.; Carvalho, E.; Jenssen, H. Improved diabetic wound healing by LFcinB is associated with relevant changes in the skin immune response and microbiota. Mol. Methods Clin. Dev. 2021, 20, 726–739. [Google Scholar] [CrossRef] [PubMed]
- Suh, W.; Kim, K.L.; Kim, J.M.; Shin, I.S.; Lee, Y.S.; Lee, J.Y.; Jang, H.S.; Lee, J.S.; Byun, J.; Choi, J.H.; et al. Transplantation of endothelial progenitor cells accelerates dermal wound healing with increased recruitment of monocytes/macrophages and neovascularization. Stem Cells 2005, 23, 1571–1578. [Google Scholar] [CrossRef] [PubMed]
- Bjarnsholt, T.; Kirketerp-Moller, K.; Jensen, P.O.; Madsen, K.G.; Phipps, R.; Krogfelt, K.; Hoiby, N.; Givskov, M. Why chronic wounds will not heal: A novel hypothesis. Wound Repair. Regen 2008, 16, 2–10. [Google Scholar] [CrossRef]
- Li, M.; Hou, Q.; Zhong, L.; Zhao, Y.; Fu, X. Macrophage Related Chronic Inflammation in Non-Healing Wounds. Front. Immunol. 2021, 12, 681710. [Google Scholar] [CrossRef]
- Mirza, R.E.; Fang, M.M.; Ennis, W.J.; Koh, T.J. Blocking interleukin-1beta induces a healing-associated wound macrophage phenotype and improves healing in type 2 diabetes. Diabetes 2013, 62, 2579–2587. [Google Scholar] [CrossRef]
- Pang, J.; Maienschein-Cline, M.; Koh, T.J. Enhanced Proliferation of Ly6C(+) Monocytes/Macrophages Contributes to Chronic Inflammation in Skin Wounds of Diabetic Mice. J. Immunol. 2021, 206, 621–630. [Google Scholar] [CrossRef]
- Kolumam, G.; Wu, X.; Lee, W.P.; Hackney, J.A.; Zavala-Solorio, J.; Gandham, V.; Danilenko, D.M.; Arora, P.; Wang, X.; Ouyang, W. IL-22R Ligands IL-20, IL-22, and IL-24 Promote Wound Healing in Diabetic db/db Mice. PLoS ONE 2017, 12, e0170639. [Google Scholar] [CrossRef] [PubMed]
- Lampropoulou, I.T.; Stangou, M.; Papagianni, A.; Didangelos, T.; Iliadis, F.; Efstratiadis, G. TNF-alpha and microalbuminuria in patients with type 2 diabetes mellitus. J. Diabetes Res. 2014, 2014, 394206. [Google Scholar] [CrossRef] [PubMed]
- Trengove, N.J.; Bielefeldt-Ohmann, H.; Stacey, M.C. Mitogenic activity and cytokine levels in non-healing and healing chronic leg ulcers. Wound Repair. Regen 2000, 8, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Medzhitov, R. TOLL-LIKE RECEPTORS AND INNATE IMMUNITY. Nat. Rev. Neurosci. 2001, 1, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Pukstad, B.S.; Ryan, L.; Flo, T.H.; Stenvik, J.; Moseley, R.; Harding, K.; Thomas, D.W.; Espevik, T. Non-healing is associated with persistent stimulation of the innate immune response in chronic venous leg ulcers. J. Derm. Sci. 2010, 59, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Shen, J.; Chai, Y.; Chen, H. IL-1beta Impaired Diabetic Wound Healing by Regulating MMP-2 and MMP-9 through the p38 Pathway. Mediat. Inflamm. 2021, 2021, 6645766. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Yang, C.; Zhang, S.; Ke, Z.X.; Chen, D.X.; Li, Y.Q.; Li, Q. The Imbalance of MMP-2/TIMP-2 and MMP-9/TIMP-1 Contributes to Collagen Deposition Disorder in Diabetic Non-Injured Skin. Front. Endocrinol. 2021, 12, 734485. [Google Scholar] [CrossRef]
- Lobmann, R.; Ambrosch, A.; Schultz, G.; Waldmann, K.; Schiweck, S.; Lehnert, H. Expression of matrix-metalloproteinases and their inhibitors in the wounds of diabetic and non-diabetic patients. Diabetologia 2002, 45, 1011–1016. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Akira, S. The roles of TLRs, RLRs and NLRs in pathogen recognition. Int. Immunol. 2009, 21, 317–337. [Google Scholar] [CrossRef]
- Sepehri, Z.; Kiani, Z.; Nasiri, A.A.; Kohan, F. Toll-like receptor 2 and type 2 diabetes. Cell Mol. Biol. Lett. 2016, 21, 2. [Google Scholar] [CrossRef]
- Dasu, M.R.; Thangappan, R.K.; Bourgette, A.; DiPietro, L.A.; Isseroff, R.; Jialal, I. TLR2 expression and signaling-dependent inflammation impair wound healing in diabetic mice. Lab Investig. 2010, 90, 1628–1636. [Google Scholar] [CrossRef]
- Creely, S.J.; McTernan, P.G.; Kusminski, C.M.; Fisher, F.M.; Da Silva, N.F.; Khanolkar, M.; Evans, M.; Harte, A.L.; Kumar, S. Lipopolysaccharide activates an innate immune system response in human adipose tissue in obesity and type 2 diabetes. Am. J. Physiol. Endocrinol. Metab. 2007, 292, E740–E747. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, K.T.; Seth, A.K.; Hong, S.J.; Geringer, M.R.; Xie, P.; Leung, K.P.; Mustoe, T.A.; Galiano, R.D. Deficient cytokine expression and neutrophil oxidative burst contribute to impaired cutaneous wound healing in diabetic, biofilm-containing chronic wounds. Wound Repair. Regen. 2013, 21, 833–841. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.; Agrawal, N.K.; Gupta, S.K.; Mohan, G.; Chaturvedi, S.; Singh, K. Genetic and epigenetic alterations in Toll like receptor 2 and wound healing impairment in type 2 diabetes patients. J. Diabetes Complicat. 2015, 29, 222–229. [Google Scholar] [CrossRef]
- Blakely, M. The Use of Best Practice in the Treatment of a Complex Diabetic Foot Ulcer: A Case Report. Healthcare 2016, 4, 18. [Google Scholar] [CrossRef]
- Tettelbach, W.H.; Cazzell, S.M.; Hubbs, B.; Jong, J.L.D.; Forsyth, R.A.; Reyzelman, A.M. The influence of adequate debridement and placental-derived allografts on diabetic foot ulcers. J. Wound Care 2022, 31, S16–S29. [Google Scholar] [CrossRef] [PubMed]
- Brouwer, R.J.; Lalieu, R.C.; Hoencamp, R.; van Hulst, R.A.; Ubbink, D.T. A systematic review and meta-analysis of hyperbaric oxygen therapy for diabetic foot ulcers with arterial insufficiency. J. Vasc. Surg. 2020, 71, 682–692.e681. [Google Scholar] [CrossRef] [PubMed]
- Snyder, R.; Galiano, R.; Mayer, P.; Rogers, L.C.; Alvarez, O.; Sanuwave Trial Investigators. Diabetic foot ulcer treatment with focused shockwave therapy: Two multicentre, prospective, controlled, double-blinded, randomised phase III clinical trials. J. Wound Care 2018, 27, 822–836. [Google Scholar] [CrossRef] [PubMed]
- Racaru, S.; Bolton Saghdaoui, L.; Roy Choudhury, J.; Wells, M.; Davies, A.H. Offloading treatment in people with diabetic foot disease: A systematic scoping review on adherence to foot offloading. Diabetes Metab. Syndr. 2022, 16, 102493. [Google Scholar] [CrossRef]
- Siavash, M.; Najjarnezhad, A.; Mohseni, N.; Abtahi, S.M.; Karimy, A.; Sabzevari, M.H. Efficacy of Maggot Debridement Therapy on Refractory Atypical Diabetic Foot Ulcers: An Open-Label Study. Int. J. Low Extrem. Wounds 2021, 20, 315–320. [Google Scholar] [CrossRef]
- Huang, J.; Chen, J.; Xiong, S.; Huang, J.; Liu, Z. The effect of low-level laser therapy on diabetic foot ulcers: A meta-analysis of randomised controlled trials. Int. Wound J. 2021, 18, 763–776. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.; Nguyen, T.T. Strategy for Treatment of Infected Diabetic Foot Ulcers. Acc. Chem. Res. 2021, 54, 1080–1093. [Google Scholar] [CrossRef] [PubMed]
- Game, F. The advantages and disadvantages of non-surgical management of the diabetic foot. Diabetes Metab. Res. Rev. 2008, 24 (Suppl. 1), S72–S75. [Google Scholar] [CrossRef]
- Oley, M.H.; Oley, M.C.; Tjandra, D.E.; Sedu, S.W.; Sumarauw, E.R.N.; Aling, D.M.R.; Kalangi, J.A.; Islam, A.A.; Hatta, M.; Faruk, M. Hyperbaric oxygen therapy in the healing process of foot ulcers in diabetic type 2 patients marked by interleukin 6, vascular endothelial growth factor, and PEDIS score: A randomized controlled trial study. Int. J. Surg. Open 2020, 27, 154–161. [Google Scholar] [CrossRef]
- Jebril, W.; Nowak, M.; Palin, L.; Nordgren, M.; Bachar-Wikstrom, E.; Wikstrom, J.D. Topical oxygen treatment relieves pain from hard-to-heal leg ulcers and improves healing: A case series. J. Wound Care 2022, 31, 4–11. [Google Scholar] [CrossRef] [PubMed]
- Shendy, W.S.; Elsoghier, O.M.; El Semary, M.M.; Ahmed, A.A.; Ali, A.F.; Saber-Khalaf, M. Effect of low-intensity extracorporeal shock wave therapy on diabetic erectile dysfunction: Randomised control trial. Andrologia 2021, 53, e13997. [Google Scholar] [CrossRef]
- Lazzarini, P.A.; Jarl, G.; Gooday, C.; Viswanathan, V.; Caravaggi, C.F.; Armstrong, D.G.; Bus, S.A. Effectiveness of offloading interventions to heal foot ulcers in persons with diabetes: A systematic review. Diabetes Metab. Res. Rev. 2020, 36 (Suppl. 1), e3275. [Google Scholar] [CrossRef] [PubMed]
- Bazalinski, D.; Kozka, M.; Karnas, M.; Wiech, P. Effectiveness of Chronic Wound Debridement with the Use of Larvae of Lucilia Sericata. J. Clin. Med. 2019, 8, 1845. [Google Scholar] [CrossRef]
- Ramirez-Acuna, J.M.; Cardenas-Cadena, S.A.; Marquez-Salas, P.A.; Garza-Veloz, I.; Perez-Favila, A.; Cid-Baez, M.A.; Flores-Morales, V.; Martinez-Fierro, M.L. Diabetic Foot Ulcers: Current Advances in Antimicrobial Therapies and Emerging Treatments. Antibiotics 2019, 8, 193. [Google Scholar] [CrossRef]
- Shaikh-Kader, A.; Houreld, N.N.; Rajendran, N.K.; Abrahamse, H. The link between advanced glycation end products and apoptosis in delayed wound healing. Cell Biochem. Funct. 2019, 37, 432–442. [Google Scholar] [CrossRef]
- Futrega, K.; King, M.; Lott, W.B.; Doran, M.R. Treating the whole not the hole: Necessary coupling of technologies for diabetic foot ulcer treatment. Trends Mol. Med. 2014, 20, 137–142. [Google Scholar] [CrossRef]
- Sica, A.; Erreni, M.; Allavena, P.; Porta, C. Macrophage polarization in pathology. Cell Mol. Life Sci. 2015, 72, 4111–4126. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-W.; Hung, C.-M.; Chen, W.-J.; Chen, J.-C.; Huang, W.-Y.; Lu, C.-S.; Kuo, M.-L.; Chen, S.-G. New Horizons of Macrophage Immunomodulation in the Healing of Diabetic Foot Ulcers. Pharmaceutics 2022, 14, 2065. [Google Scholar] [CrossRef]
- Leu, W.J.; Chen, J.C.; Guh, J.H. Extract From Plectranthus amboinicus Inhibit Maturation and Release of Interleukin 1beta Through Inhibition of NF-kappaB Nuclear Translocation and NLRP3 Inflammasome Activation. Front. Pharm. 2019, 10, 573. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.W.; Chen, C.C.; Huang, W.Y.; Chen, Y.Y.; Chen, S.T.; Chou, H.W.; Hung, C.M.; Chen, W.J.; Lu, C.S.; Nian, S.X.; et al. Restoring Prohealing/Remodeling-Associated M2a/c Macrophages Using ON101 Accelerates Diabetic Wound Healing. JID Innov. 2022, 2, 100138. [Google Scholar] [CrossRef] [PubMed]
- Chiang, K.J.; Chiu, L.C.; Kang, Y.N.; Chen, C. Autologous Stem Cell Therapy for Chronic Lower Extremity Wounds: A Meta-Analysis of Randomized Controlled Trials. Cells 2021, 10, 3307. [Google Scholar] [CrossRef] [PubMed]
- Xie, F.; Teng, L.; Lu, J.; Xu, J.; Zhang, C.; Yang, L.; Ma, X.; Zhao, M. Interleukin-10-Modified Adipose-Derived Mesenchymal Stem Cells Prevent Hypertrophic Scar Formation via Regulating the Biological Characteristics of Fibroblasts and Inflammation. Mediat. Inflamm. 2022, 2022, 6368311. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, L.; Nabzdyk, C.; Andersen, N.D.; LoGerfo, F.W.; Veves, A. Inflammation and neuropeptides: The connection in diabetic wound healing. Expert. Rev. Mol. Med. 2009, 11, e2. [Google Scholar] [CrossRef]
- Dallos, A.; Kiss, M.; Polyanka, H.; Dobozy, A.; Kemeny, L.; Husz, S. Effects of the neuropeptides substance P, calcitonin gene-related peptide, vasoactive intestinal polypeptide and galanin on the production of nerve growth factor and inflammatory cytokines in cultured human keratinocytes. Neuropeptides 2006, 40, 251–263. [Google Scholar] [CrossRef] [PubMed]
- Gajendrareddy, P.K.; Engeland, C.G.; Junges, R.; Horan, M.P.; Rojas, I.G.; Marucha, P.T. MMP-8 overexpression and persistence of neutrophils relate to stress-impaired healing and poor collagen architecture in mice. Brain Behav. Immun. 2013, 28, 44–48. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Fernandez, A.; Inada, M.; Balbin, M.; Fueyo, A.; Pitiot, A.S.; Astudillo, A.; Hirose, K.; Hirata, M.; Shapiro, S.D.; Noel, A.; et al. Increased inflammation delays wound healing in mice deficient in collagenase-2 (MMP-8). FASEB J. 2007, 21, 2580–2591. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.T.; Ding, D.; Wolter, W.R.; Perez, R.L.; Champion, M.M.; Mahasenan, K.V.; Hesek, D.; Lee, M.; Schroeder, V.A.; Jones, J.I.; et al. Validation of Matrix Metalloproteinase-9 (MMP-9) as a Novel Target for Treatment of Diabetic Foot Ulcers in Humans and Discovery of a Potent and Selective Small-Molecule MMP-9 Inhibitor That Accelerates Healing. J. Med. Chem. 2018, 61, 8825–8837. [Google Scholar] [CrossRef] [PubMed]
- Gooyit, M.; Lee, M.; Schroeder, V.A.; Ikejiri, M.; Suckow, M.A.; Mobashery, S.; Chang, M. Selective water-soluble gelatinase inhibitor prodrugs. J. Med. Chem. 2011, 54, 6676–6690. [Google Scholar] [CrossRef][Green Version]
- Wang, W.; Yang, C.; Wang, X.Y.; Zhou, L.Y.; Lao, G.J.; Liu, D.; Wang, C.; Hu, M.D.; Zeng, T.T.; Yan, L.; et al. MicroRNA-129 and -335 Promote Diabetic Wound Healing by Inhibiting Sp1-Mediated MMP-9 Expression. Diabetes 2018, 67, 1627–1638. [Google Scholar] [CrossRef]
- Motley, M.P.; Banerjee, K.; Fries, B.C. Monoclonal antibody-based therapies for bacterial infections. Curr. Opin. Infect. Dis. 2019, 32, 210–216. [Google Scholar] [CrossRef]
- Tkaczyk, C.; Jones-Nelson, O.; Shi, Y.Y.; Tabor, D.E.; Cheng, L.; Zhang, T.; Sellman, B.R. Neutralizing Staphylococcus aureus Virulence with AZD6389, a Three mAb Combination, Accelerates Closure of a Diabetic Polymicrobial Wound. mSphere 2022, 7, e00130-00122. [Google Scholar] [CrossRef]
- Choi, M.H.; Jang, T.S.; Kim, H.; Ku, I.; Lee, J.; Jeong, J.G.; Kim, S.; Park, J.U. An Agonistic Monoclonal Antibody Targeting cMet Attenuates Inflammation and Up-Regulates Collagen Synthesis and Angiogenesis in Type 2 Diabetic Mice Wounds. Plast Reconstr. Surg. 2022, 150, 572e–583e. [Google Scholar] [CrossRef]
- Artem Ataide, J.; Caramori Cefali, L.; Machado Croisfelt, F.; Arruda Martins Shimojo, A.; Oliveira-Nascimento, L.; Gava Mazzola, P. Natural actives for wound healing: A review. Phytother Res. 2018, 32, 1664–1674. [Google Scholar] [CrossRef]
- Hozzein, W.N.; Badr, G.; Al Ghamdi, A.A.; Sayed, A.; Al-Waili, N.S.; Garraud, O. Topical application of propolis enhances cutaneous wound healing by promoting TGF-beta/Smad-mediated collagen production in a streptozotocin-induced type I diabetic mouse model. Cell Physiol. Biochem. 2015, 37, 940–954. [Google Scholar] [CrossRef]
- Powell, L.C.; Cullen, J.K.; Boyle, G.M.; De Ridder, T.; Yap, P.-Y.; Xue, W.; Pierce, C.J.; Pritchard, M.F.; Menzies, G.E.; Abdulkarim, M.; et al. Topical, immunomodulatory epoxy-tiglianes induce biofilm disruption and healing in acute and chronic skin wounds. Sci. Transl. Med. 2022, 14, eabn3758. [Google Scholar] [CrossRef] [PubMed]
- Game, F.; Jeffcoate, W.; Tarnow, L.; Jacobsen, J.L.; Whitham, D.J.; Harrison, E.F.; Ellender, S.J.; Fitzsimmons, D.; Löndahl, M.; Dhatariya, K.; et al. LeucoPatch system for the management of hard-to-heal diabetic foot ulcers in the UK, Denmark, and Sweden: An observer-masked, randomised controlled trial. Lancet Diabetes Endocrinol. 2018, 6, 870–878. [Google Scholar] [CrossRef]
- Yang, J.Y.; Chen, C.C.; Chang, S.C.; Yeh, J.T.; Huang, H.F.; Lin, H.C.; Lin, S.H.; Lin, Y.H.; Wei, L.G.; Liu, T.J.; et al. ENERGI-F703 gel, as a new topical treatment for diabetic foot and leg ulcers: A multicenter, randomized, double-blind, phase II trial. EClinicalMedicine 2022, 51, 101497. [Google Scholar] [CrossRef] [PubMed]
- Luqman, A.; Gotz, F. The Ambivalent Role of Skin Microbiota and Adrenaline in Wound Healing and the Interplay between Them. Int. J. Mol. Sci. 2021, 22, 4996. [Google Scholar] [CrossRef]
- Diotallevi, F.; Campanati, A.; Martina, E.; Radi, G.; Paolinelli, M.; Marani, A.; Molinelli, E.; Candelora, M.; Taus, M.; Galeazzi, T.; et al. The Role of Nutrition in Immune-Mediated, Inflammatory Skin Disease: A Narrative Review. Nutrients 2022, 14, 591. [Google Scholar] [CrossRef]
- Thye, A.Y.; Bah, Y.R.; Law, J.W.; Tan, L.T.; He, Y.W.; Wong, S.H.; Thurairajasingam, S.; Chan, K.G.; Lee, L.H.; Letchumanan, V. Gut-Skin Axis: Unravelling the Connection between the Gut Microbiome and Psoriasis. Biomedicines 2022, 10, 1037. [Google Scholar] [CrossRef]
- Salem, I.; Ramser, A.; Isham, N.; Ghannoum, M.A. The Gut Microbiome as a Major Regulator of the Gut-Skin Axis. Front. Microbiol. 2018, 9, 1459. [Google Scholar] [CrossRef]
- Szabo, K.; Erdei, L.; Bolla, B.S.; Tax, G.; Biro, T.; Kemeny, L. Factors shaping the composition of the cutaneous microbiota. Br. J. Derm. 2017, 176, 344–351. [Google Scholar] [CrossRef]
- Naik, S.; Bouladoux, N.; Linehan, J.L.; Han, S.J.; Harrison, O.J.; Wilhelm, C.; Conlan, S.; Himmelfarb, S.; Byrd, A.L.; Deming, C.; et al. Commensal-dendritic-cell interaction specifies a unique protective skin immune signature. Nature 2015, 520, 104–108. [Google Scholar] [CrossRef]
- Naik, S.; Bouladoux, N.; Wilhelm, C.; Molloy, M.J.; Salcedo, R.; Kastenmuller, W.; Deming, C.; Quinones, M.; Koo, L.; Conlan, S.; et al. Compartmentalized control of skin immunity by resident commensals. Sci. Transl. Med. 2012, 337, 1115–1119. [Google Scholar] [CrossRef]
- Kao, M.S.; Huang, S.; Chang, W.L.; Hsieh, M.F.; Huang, C.J.; Gallo, R.L.; Huang, C.M. Microbiome precision editing: Using PEG as a selective fermentation initiator against methicillin-resistant Staphylococcus aureus. Biotechnol. J. 2017, 12, 1600399. [Google Scholar] [CrossRef]
- Argenta, A.; Satish, L.; Gallo, P.; Liu, F.; Kathju, S. Local Application of Probiotic Bacteria Prophylaxes against Sepsis and Death Resulting from Burn Wound Infection. PLoS ONE 2016, 11, e0165294. [Google Scholar] [CrossRef] [PubMed]
- Peral, M.C.; Huaman Martinez, M.A.; Valdez, J.C. Bacteriotherapy with Lactobacillus plantarum in burns. Int. Wound J. 2009, 6, 73–81. [Google Scholar] [CrossRef]
- Poutahidis, T.; Kearney, S.M.; Levkovich, T.; Qi, P.; Varian, B.J.; Lakritz, J.R.; Ibrahim, Y.M.; Chatzigiagkos, A.; Alm, E.J.; Erdman, S.E. Microbial symbionts accelerate wound healing via the neuropeptide hormone oxytocin. PLoS ONE 2013, 8, e78898. [Google Scholar] [CrossRef] [PubMed]
- Tegegne, B.A.; Kebede, B. Probiotics, their prophylactic and therapeutic applications in human health development: A review of the literature. Heliyon 2022, 8, e09725. [Google Scholar] [CrossRef]
- Gurung, M.; Li, Z.; You, H.; Rodrigues, R.; Jump, D.B.; Morgun, A.; Shulzhenko, N. Role of gut microbiota in type 2 diabetes pathophysiology. EBioMedicine 2020, 51, 102590. [Google Scholar] [CrossRef] [PubMed]
- Dahiya, D.; Nigam, P.S. Probiotics, Prebiotics, Synbiotics, and Fermented Foods as Potential Biotics in Nutrition Improving Health via Microbiome-Gut-Brain Axis. Fermentation 2022, 8, 303. [Google Scholar] [CrossRef]
- McLoughlin, I.J.; Wright, E.M.; Tagg, J.R.; Jain, R.; Hale, J.D.F. Skin Microbiome-The Next Frontier for Probiotic Intervention. Probiotics Antimicrob. Proteins 2022, 14, 630–647. [Google Scholar] [CrossRef] [PubMed]
- de Sire, A.; de Sire, R.; Curci, C.; Castiglione, F.; Wahli, W. Role of Dietary Supplements and Probiotics in Modulating Microbiota and Bone Health: The Gut-Bone Axis. Cells 2022, 11, 743. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.; Veiga, A.S.; Tavares, L.; Castanho, M.; Oliveira, M. Bacterial Biofilms in Diabetic Foot Ulcers: Potential Alternative Therapeutics. In Microbial Biofilms—Importance and Applications; IntechOpen: Rijeka, Croatia, 2016; pp. 251–269. [Google Scholar]
- Klaenhammer, T.R.; Kleerebezem, M.; Kopp, M.V.; Rescigno, M. The impact of probiotics and prebiotics on the immune system. Nat. Rev. Immunol. 2012, 12, 728–734. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.M.; Jeong, J.J.; Woo, K.H.; Han, M.J.; Kim, D.H. Lactobacillus sakei OK67 ameliorates high-fat diet-induced blood glucose intolerance and obesity in mice by inhibiting gut microbiota lipopolysaccharide production and inducing colon tight junction protein expression. Nutr. Res. 2016, 36, 337–348. [Google Scholar] [CrossRef]
- Balakumar, M.; Prabhu, D.; Sathishkumar, C.; Prabu, P.; Rokana, N.; Kumar, R.; Raghavan, S.; Soundarajan, A.; Grover, S.; Batish, V.K.; et al. Improvement in glucose tolerance and insulin sensitivity by probiotic strains of Indian gut origin in high-fat diet-fed C57BL/6J mice. Eur. J. Nutr. 2018, 57, 279–295. [Google Scholar] [CrossRef]
- Maldonado Galdeano, C.; Cazorla, S.I.; Lemme Dumit, J.M.; Velez, E.; Perdigon, G. Beneficial Effects of Probiotic Consumption on the Immune System. Ann. Nutr. Metab. 2019, 74, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Van Baarlen, P.; Troost, F.J.; van Hemert, S.; van der Meer, C.; de Vos, W.M.; de Groot, P.J.; Hooiveld, G.J.; Brummer, R.J.M.; Kleerebezem, M. Differential NF-κB pathways induction by Lactobacillus plantarum in the duodenum of healthy humans correlating with immune tolerance. Proc. Natl. Acad. Sci. USA 2009, 106, 2371–2376. [Google Scholar] [CrossRef]
- van Baarlen, P.; Troost, F.; van der Meer, C.; Hooiveld, G.; Boekschoten, M.; Brummer, R.J.; Kleerebezem, M. Human mucosal in vivo transcriptome responses to three lactobacilli indicate how probiotics may modulate human cellular pathways. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. 1), 4562–4569. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Naik, B.; Kumar, A.; Khanduri, N.; Rustagi, S.; Kumar, S. Probiotics media: Significance, challenges, and future perspective—A mini review. Food Prod. Processing Nutr. 2022, 4, 17. [Google Scholar] [CrossRef]
- Han, H.; Li, Y.; Fang, J.; Liu, G.; Yin, J.; Li, T.; Yin, Y. Gut Microbiota and Type 1 Diabetes. Int. J. Mol. Sci. 2018, 19, 995. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.-H.; Yen, H.-R.; Lu, W.-L.; Ho, H.-H.; Lin, W.-Y.; Kuo, Y.-W.; Huang, Y.-Y.; Tsai, S.-Y.; Lin, H.-C. Adjuvant Probiotics of Lactobacillus salivarius subsp. salicinius AP-32, L. johnsonii MH-68, and Bifidobacterium animalis subsp. lactis CP-9 Attenuate Glycemic Levels and Inflammatory Cytokines in Patients With Type 1 Diabetes Mellitus. Front. Endocrinol. 2022, 13, 754401. [Google Scholar] [CrossRef]
- Madempudi, R.S.; Ahire, J.J.; Neelamraju, J.; Tripathi, A.; Nanal, S. Efficacy of UB0316, a multi-strain probiotic formulation in patients with type 2 diabetes mellitus: A double blind, randomized, placebo controlled study. PLoS ONE 2019, 14, e0225168. [Google Scholar] [CrossRef]
- Zamora-Pineda, J.; Kalinina, O.; Osborne, B.A.; Knight, K.L. Probiotic Molecules That Inhibit Inflammatory Diseases. Appl. Sci. 2022, 12, 1147. [Google Scholar] [CrossRef]
- Mantis, N.J.; Rol, N.; Corthesy, B. Secretory IgA’s complex roles in immunity and mucosal homeostasis in the gut. Mucosal. Immunol. 2011, 4, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Azad, M.A.K.; Sarker, M.; Wan, D. Immunomodulatory Effects of Probiotics on Cytokine Profiles. Biomed. Res. Int. 2018, 2018, 8063647. [Google Scholar] [CrossRef] [PubMed]
- Hickey, A.; Stamou, P.; Udayan, S.; Ramon-Vazquez, A.; Esteban-Torres, M.; Bottacini, F.; Woznicki, J.A.; Hughes, O.; Melgar, S.; Ventura, M.; et al. Bifidobacterium breve Exopolysaccharide Blocks Dendritic Cell Maturation and Activation of CD4(+) T Cells. Front. Microbiol. 2021, 12, 653587. [Google Scholar] [CrossRef] [PubMed]
- Schlee, M.; Wehkamp, J.; Altenhoefer, A.; Oelschlaeger, T.A.; Stange, E.F.; Fellermann, K. Induction of human beta-defensin 2 by the probiotic Escherichia coli Nissle 1917 is mediated through flagellin. Infect. Immun. 2007, 75, 2399–2407. [Google Scholar] [CrossRef]
- Yan, S.; Yang, B.; Zhao, J.; Zhao, J.; Stanton, C.; Ross, R.P.; Zhang, H.; Chen, W. A ropy exopolysaccharide producing strain Bifidobacterium longum subsp. longum YS108R alleviates DSS-induced colitis by maintenance of the mucosal barrier and gut microbiota modulation. Food Funct. 2019, 10, 1595–1608. [Google Scholar] [CrossRef] [PubMed]
- Sproston, N.R.; Ashworth, J.J. Role of C-Reactive Protein at Sites of Inflammation and Infection. Front. Immunol. 2018, 9, 754. [Google Scholar] [CrossRef] [PubMed]
- Mazruei Arani, N.; Emam-Djomeh, Z.; Tavakolipour, H.; Sharafati-Chaleshtori, R.; Soleimani, A.; Asemi, Z. The Effects of Probiotic Honey Consumption on Metabolic Status in Patients with Diabetic Nephropathy: A Randomized, Double-Blind, Controlled Trial. Probiotics Antimicrob. Proteins 2019, 11, 1195–1201. [Google Scholar] [CrossRef] [PubMed]
- Mousavi, S.N.; Saboori, S.; Asbaghi, O. Effect of daily probiotic yogurt consumption on inflammation: A systematic review and meta-analysis of randomized Controlled Clinical trials. Obes. Med. 2020, 18, 100221. [Google Scholar] [CrossRef]
- Mafi, A.; Namazi, G.; Soleimani, A.; Bahmani, F.; Aghadavod, E.; Asemi, Z. Metabolic and genetic response to probiotics supplementation in patients with diabetic nephropathy: A randomized, double-blind, placebo-controlled trial. Food Funct. 2018, 9, 4763–4770. [Google Scholar] [CrossRef] [PubMed]
- Sharma, H.; Sharma, S.; Krishnan, A.; Yuan, D.; Vangaveti, V.N.; Malabu, U.H.; Haleagrahara, N. The efficacy of inflammatory markers in diagnosing infected diabetic foot ulcers and diabetic foot osteomyelitis: Systematic review and meta-analysis. PLoS ONE 2022, 17, e0267412. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Suh, D.H.; Kim, H.J.; Lee, Y.I.; Kwak, I.H.; Choi, G.W. Role of procalcitonin in infected diabetic foot ulcer. Diabetes Res. Clin. Pr. 2017, 128, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Sanaie, S.; Ebrahimi-Mameghani, M.; Hamishehkar, H.; Mojtahedzadeh, M.; Mahmoodpoor, A. Effect of a multispecies probiotic on inflammatory markers in critically ill patients: A randomized, double-blind, placebo-controlled trial. J. Res. Med. Sci. 2014, 9, 827. [Google Scholar]
- Yunir, E.; Tahapary, D.L.; Tarigan, T.J.E.; Harbuwono, D.S.; Oktavianda, Y.D.; Kristanti, M.; Iswati, E.; Sarumpaet, A.; Soewondo, P. Non-vascular contributing factors of diabetic foot ulcer severity in national referral hospital of Indonesia. J. Diabetes Metab. Disord. 2021, 20, 805–813. [Google Scholar] [CrossRef] [PubMed]
- Wilmink, J.M.; Ladefoged, S.; Jongbloets, A.; Vernooij, J.C.M. The evaluation of the effect of probiotics on the healing of equine distal limb wounds. PLoS ONE 2020, 15, e0236761. [Google Scholar] [CrossRef] [PubMed]
- Stene, C.; Rome, A.; Palmquist, I.; Linninge, C.; Molin, G.; Ahrne, S.; Johnson, L.B.; Jeppsson, B. Administration of probiotics to healthy volunteers: Effects on reactivity of intestinal mucosa and systemic leukocytes. BMC Gastroenterol. 2022, 22, 100. [Google Scholar] [CrossRef] [PubMed]
- Maione, A.G.; Smith, A.; Kashpur, O.; Yanez, V.; Knight, E.; Mooney, D.J.; Veves, A.; Tomic-Canic, M.; Garlick, J.A. Altered ECM deposition by diabetic foot ulcer-derived fibroblasts implicates fibronectin in chronic wound repair. Wound Repair Regen. 2016, 24, 630–643. [Google Scholar] [CrossRef]
- Woo, Y.M.; Kim, O.J.; Jo, E.S.; Jo, M.Y.; Ahn, M.Y.; Lee, Y.-H.; Li, C.-r.; Lee, S.-H.; Choi, J.-S.; Ha, J.M.; et al. The effect of Lactobacillus plantarum hydrolysates promoting VEGF production on vascular growth and hair growth of C57BL/6 mice. J. Anal. Sci. Technol. 2019, 10, 1–9. [Google Scholar] [CrossRef]
- Ridiandries, A.; Tan, J.T.M.; Bursill, C.A. The Role of Chemokines in Wound Healing. Int. J. Mol. Sci. 2018, 19, 3217. [Google Scholar] [CrossRef]
- Watters, C.; DeLeon, K.; Trivedi, U.; Griswold, J.A.; Lyte, M.; Hampel, K.J.; Wargo, M.J.; Rumbaugh, K.P. Pseudomonas aeruginosa biofilms perturb wound resolution and antibiotic tolerance in diabetic mice. Med. Microbiol. Immunol. 2013, 202, 131–141. [Google Scholar] [CrossRef]
- Algburi, A.; Al-Hasani, H.M.; Ismael, T.K.; Abdelhameed, A.; Weeks, R.; Ermakov, A.M.; Chikindas, M.L. Antimicrobial Activity of Bacillus subtilis KATMIRA1933 and Bacillus amyloliquefaciens B-1895 Against Staphylococcus aureus Biofilms Isolated from Wound Infection. Probiotics Antimicrob. Proteins 2021, 13, 125–134. [Google Scholar] [CrossRef]
- Varma, P.; Nisha, N.; Dinesh, K.R.; Kumar, A.V.; Biswas, R. Anti-infective properties of Lactobacillus fermentum against Staphylococcus aureus and Pseudomonas aeruginosa. J. Mol. Microbiol. Biotechnol. 2011, 20, 137–143. [Google Scholar] [CrossRef]
- Huang, Y.; Kyriakides, T.R. The role of extracellular matrix in the pathophysiology of diabetic wounds. Matrix Biol. Plus 2020, 6–7, 100037. [Google Scholar] [CrossRef] [PubMed]
- Lorca, G.; Torino, M.I.; Font de Valdez, G.; Ljungh, Å. Lactobacilli express cell surface proteins which mediate binding of immobilized collagen and fibronectin. FEMS Microbiol. Lett. 2002, 206, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Styriak, I.; Nemcova, R.; Chang, Y.H.; Ljungh, A. Binding of extracellular matrix molecules by probiotic bacteria. Lett. Appl. Microbiol. 2003, 37, 329–333. [Google Scholar] [CrossRef] [PubMed]
- McCarty, S.M.; Percival, S.L. Proteases and Delayed Wound Healing. Adv. Wound Care 2013, 2, 438–447. [Google Scholar] [CrossRef]
- Maghsood, F.; Mirshafiey, A.; Farahani, M.M.; Modarressi, M.H.; Jafari, P.; Motevaseli, E. Dual Effects of Cell Free Supernatants from Lactobacillus acidophilus and Lactobacillus rhamnosus GG in Regulation of MMP-9 by Up-Regulating TIMP-1 and Down-Regulating CD147 in PMADifferentiated THP-1 Cells. Cell J. 2018, 19, 559–568. [Google Scholar] [CrossRef] [PubMed]
- Chuang, Y.C.; Cheng, M.C.; Lee, C.C.; Chiou, T.Y.; Tsai, T.Y. Effect of ethanol extract from Lactobacillus plantarum TWK10-fermented soymilk on wound healing in streptozotocin-induced diabetic rat. AMB Express 2019, 9, 163. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, K.; Asai, J.; Ii, M.; Thorne, T.; Losordo, D.W.; D’Amore, P.A. Decreased macrophage number and activation lead to reduced lymphatic vessel formation and contribute to impaired diabetic wound healing. Am. J. Pathol. 2007, 170, 1178–1191. [Google Scholar] [CrossRef] [PubMed]
- Han, N.; Jia, L.; Su, Y.; Du, J.; Guo, L.; Luo, Z.; Liu, Y. Lactobacillus reuteri extracts promoted wound healing via PI3K/AKT/beta-catenin/TGFbeta1 pathway. Stem Cell Res. 2019, 10, 243. [Google Scholar] [CrossRef]
- Arganaraz Aybar, J.N.; Ortiz Mayor, S.; Olea, L.; Garcia, J.J.; Nisoria, S.; Kolling, Y.; Melian, C.; Rachid, M.; Torres Dimani, R.; Werenitzky, C.; et al. Topical Administration of Lactiplantibacillus plantarum Accelerates the Healing of Chronic Diabetic Foot Ulcers through Modifications of Infection, Angiogenesis, Macrophage Phenotype and Neutrophil Response. Microorganisms 2022, 10, 634. [Google Scholar] [CrossRef] [PubMed]
- Mohammedsaeed, W.; Cruickshank, S.; McBain, A.J.; O’Neill, C.A. Lactobacillus rhamnosus GG Lysate Increases Re-Epithelialization of Keratinocyte Scratch Assays by Promoting Migration. Sci. Rep. 2015, 5, 16147. [Google Scholar] [CrossRef]
- Mohseni, S.; Bayani, M.; Bahmani, F.; Tajabadi-Ebrahimi, M.; Bayani, M.A.; Jafari, P.; Asemi, Z. The beneficial effects of probiotic administration on wound healing and metabolic status in patients with diabetic foot ulcer: A randomized, double-blind, placebo-controlled trial. Diabetes Metab. Res. Rev. 2018, 34, 2970. [Google Scholar] [CrossRef] [PubMed]
- Vagesjo, E.; Ohnstedt, E.; Mortier, A.; Lofton, H.; Huss, F.; Proost, P.; Roos, S.; Phillipson, M. Accelerated wound healing in mice by on-site production and delivery of CXCL12 by transformed lactic acid bacteria. Proc. Natl. Acad. Sci. USA 2018, 115, 1895–1900. [Google Scholar] [CrossRef] [PubMed]
- Campos, L.F.; Tagliari, E.; Casagrande, T.A.C.; Noronha, L.; Campos, A.C.L.; Matias, J.E.F. Effects of Probiotics Supplementation on Skin Wound Healing in Diabetic Rats. Arq Bras. Cir. Dig. 2020, 33, e1498. [Google Scholar] [CrossRef]
- Mohtashami, M.; Mohamadi, M.; Azimi-Nezhad, M.; Saeidi, J.; Nia, F.F.; Ghasemi, A. Lactobacillus bulgaricus and Lactobacillus plantarum improve diabetic wound healing through modulating inflammatory factors. Biotechnol. Appl. Biochem. 2021, 68, 1421–1431. [Google Scholar] [CrossRef]
- Zhu, Y.; Wang, Z.; Bai, L.; Deng, J.; Zhou, Q. Biomaterial-based encapsulated probiotics for biomedical applications: Current status and future perspectives. Mater. Des. 2021, 210, 110018. [Google Scholar] [CrossRef]
- Li, Z.; Behrens, A.M.; Ginat, N.; Tzeng, S.Y.; Lu, X.; Sivan, S.; Langer, R.; Jaklenec, A. Biofilm-Inspired Encapsulation of Probiotics for the Treatment of Complex Infections. Adv. Mater. 2018, 30, e1803925. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhang, G.; Yang, X.; Peng, M.; Ge, S.; Tan, S.; Wen, Z.; Wang, Y.; Wu, S.; Liang, Y.; et al. An oral “Super probiotics” with versatile self-assembly adventitia for enhanced intestinal colonization by autonomous regulating the pathological microenvironment. Chem. Eng. J. 2022, 446, 137204. [Google Scholar] [CrossRef]
- Peng, Y.; He, D.; Ge, X.; Lu, Y.; Chai, Y.; Zhang, Y.; Mao, Z.; Luo, G.; Deng, J.; Zhang, Y. Construction of heparin-based hydrogel incorporated with Cu5.4O ultrasmall nanozymes for wound healing and inflammation inhibition. Bioact. Mater. 2021, 6, 3109–3124. [Google Scholar] [CrossRef]
- Durazzo, A.; Nazhand, A.; Lucarini, M.; Atanasov, A.G.; Souto, E.B.; Novellino, E.; Capasso, R.; Santini, A. An Updated Overview on Nanonutraceuticals: Focus on Nanoprebiotics and Nanoprobiotics. Int. J. Mol. Sci. 2020, 21, 2285. [Google Scholar] [CrossRef]
- Zhu, W.; Zhou, S.; Liu, J.; McLean, R.J.C.; Chu, W. Prebiotic, immuno-stimulating and gut microbiota-modulating effects of Lycium barbarum polysaccharide. Biomed. Pharm. 2020, 121, 109591. [Google Scholar] [CrossRef]
- Ghafouri, A.; Zarrati, M.; Shidfar, F.; Heydari, I.; Shokouhi Shoormasti, R.; Eslami, O. Effect of synbiotic bread containing lactic acid on glycemic indicators, biomarkers of antioxidant status and inflammation in patients with type 2 diabetes: A randomized controlled trial. Diabetol. Metab. Syndr. 2019, 11, 103. [Google Scholar] [CrossRef]
- Pistol, G.C.; Marin, D.E.; Dragomir, C.; Taranu, I. Synbiotic combination of prebiotic grape pomace extract and probiotic Lactobacillus sp. reduced important intestinal inflammatory markers and in-depth signalling mediators in lipopolysaccharide-treated Caco-2 cells. Br. J. Nutr. 2019, 121, 291–305. [Google Scholar] [CrossRef] [PubMed]
- Hong, L.; Kim, W.S.; Lee, S.M.; Kang, S.K.; Choi, Y.J.; Cho, C.S. Pullulan Nanoparticles as Prebiotics Enhance the Antibacterial Properties of Lactobacillus plantarum Through the Induction of Mild Stress in Probiotics. Front. Microbiol. 2019, 10, 142. [Google Scholar] [CrossRef]
- Ghosh, S.; Ahmad, R.; Banerjee, K.; AlAjmi, M.F.; Rahman, S. Mechanistic Aspects of Microbe-Mediated Nanoparticle Synthesis. Front. Microbiol. 2021, 12, 638068. [Google Scholar] [CrossRef]
- Sharma, S.; Sharma, N.; Kaushal, N. Comparative Account of Biogenic Synthesis of Silver Nanoparticles Using Probiotics and Their Antimicrobial Activity Against Challenging Pathogens. BioNanoScience 2022, 12, 833–840. [Google Scholar] [CrossRef]
- Joshi, B.S.; Ortiz, D.; Zuhorn, I.S. Converting extracellular vesicles into nanomedicine: Loading and unloading of cargo. Mater. Today Nano 2021, 16, 100148. [Google Scholar] [CrossRef]
- Rodovalho, V.R.; da Luz, B.S.R.; Rabah, H.; do Carmo, F.L.R.; Folador, E.L.; Nicolas, A.; Jardin, J.; Briard-Bion, V.; Blottiere, H.; Lapaque, N.; et al. Extracellular Vesicles Produced by the Probiotic Propionibacterium freudenreichii CIRM-BIA 129 Mitigate Inflammation by Modulating the NF-kappaB Pathway. Front. Microbiol. 2020, 11, 1544. [Google Scholar] [CrossRef]
- Morishita, M.; Horita, M.; Higuchi, A.; Marui, M.; Katsumi, H.; Yamamoto, A. Characterizing Different Probiotic-Derived Extracellular Vesicles as a Novel Adjuvant for Immunotherapy. Mol. Pharm. 2021, 18, 1080–1092. [Google Scholar] [CrossRef] [PubMed]
- Bauerl, C.; Coll-Marques, J.M.; Tarazona-Gonzalez, C.; Perez-Martinez, G. Lactobacillus casei extracellular vesicles stimulate EGFR pathway likely due to the presence of proteins P40 and P75 bound to their surface. Sci. Rep. 2020, 10, 19237. [Google Scholar] [CrossRef]
- Yazdanpanah, L.; Nasiri, M.; Adarvishi, S. Literature review on the management of diabetic foot ulcer. World J. Diabetes 2015, 6, 37–53. [Google Scholar] [CrossRef] [PubMed]
- Goyal, M.; Reeves, N.D.; Rajbhandari, S.; Ahmad, N.; Wang, C.; Yap, M.H. Recognition of ischaemia and infection in diabetic foot ulcers: Dataset and techniques. Comput. Biol. Med. 2020, 117, 103616. [Google Scholar] [CrossRef] [PubMed]
- Abdelhamid, A.G.; El-Masry, S.S.; El-Dougdoug, N.K. Probiotic Lactobacillus and Bifidobacterium strains possess safety characteristics, antiviral activities and host adherence factors revealed by genome mining. EPMA J. 2019, 10, 337–350. [Google Scholar] [CrossRef] [PubMed]
- Martin Manuel, P.; Elena, B.; Carolina, M.G.; Gabriela, P. Oral probiotics supplementation can stimulate the immune system in a stress process. J. Nutr. Intermed. Metab. 2017, 8, 29–40. [Google Scholar] [CrossRef]
- Kilic Yildirim, G.; Dinleyici, M.; Vandenplas, Y.; Dinleyici, E.C. Effects of Multispecies Synbiotic Supplementation on Anthropometric Measurements, Glucose and Lipid Parameters in Children With Exogenous Obesity: A Randomized, Double Blind, Placebo-Controlled Clinical Trial (Probesity-2 Trial). Front. Nutr. 2022, 9, 898037. [Google Scholar] [CrossRef]
- Nataraj, B.H.; Ali, S.A.; Behare, P.V.; Yadav, H. Postbiotics-parabiotics: The new horizons in microbial biotherapy and functional foods. Microb. Cell Fact. 2020, 19, 168. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).