
Sex Differences in Intestinal P-Glycoprotein Expression in Wistar versus Sprague Dawley Rats


 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animals
2.3. Intestinal Tissue Collection
2.4. Initial Total Protein Quantification
2.5. Measurement of P-glycoprotein (P-gp) Protein Levels by Quantitative Enzyme-Linked Immunosorbent Assay (ELISA) and Quantitative Liquid Chromatography–Tandem Mass Spectrometry (LC-MS/MS)
2.6. Statistical Analysis
3. Results
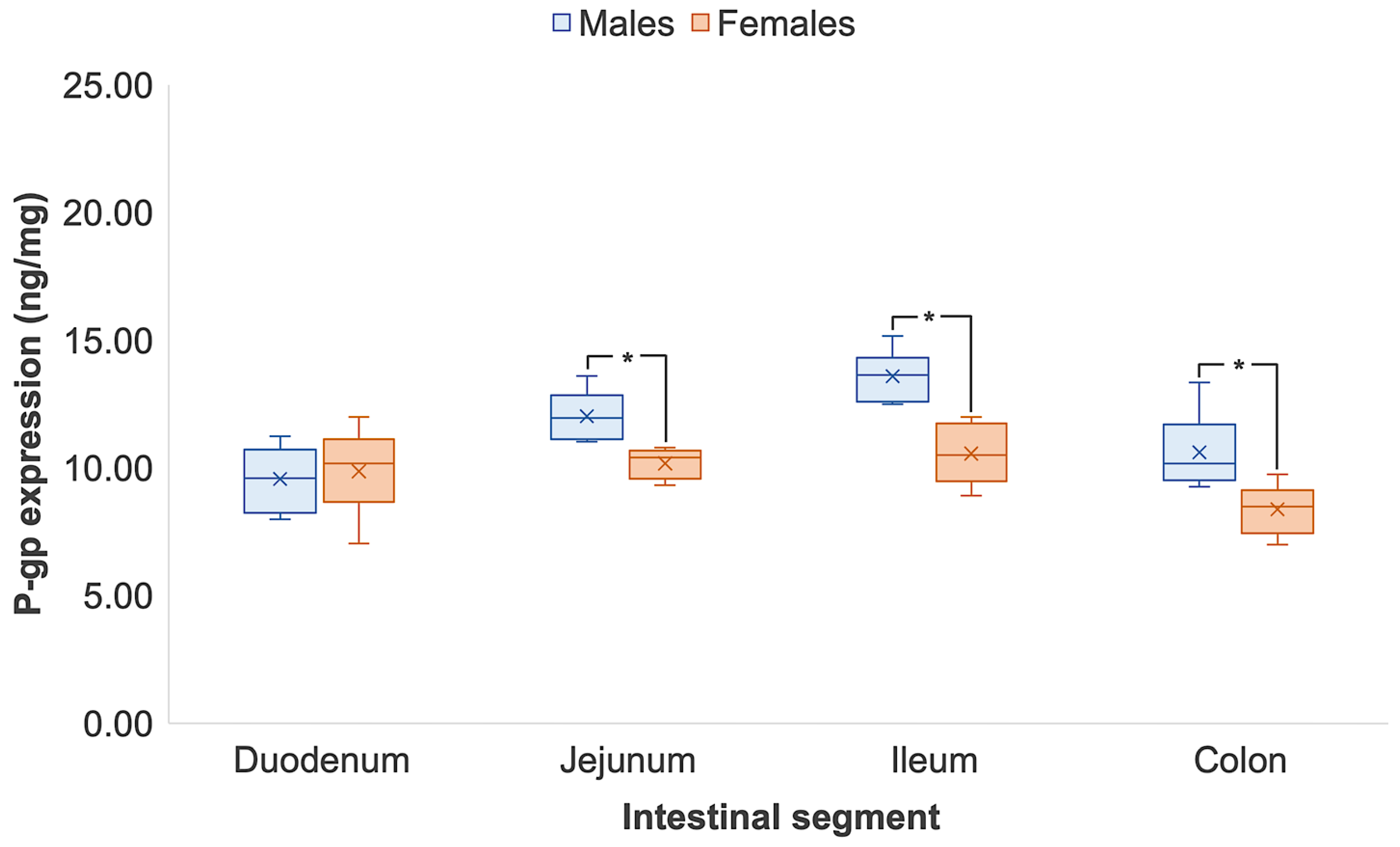
3.1. Intestinal P-glycoprotein Quantification in Wistar Rats
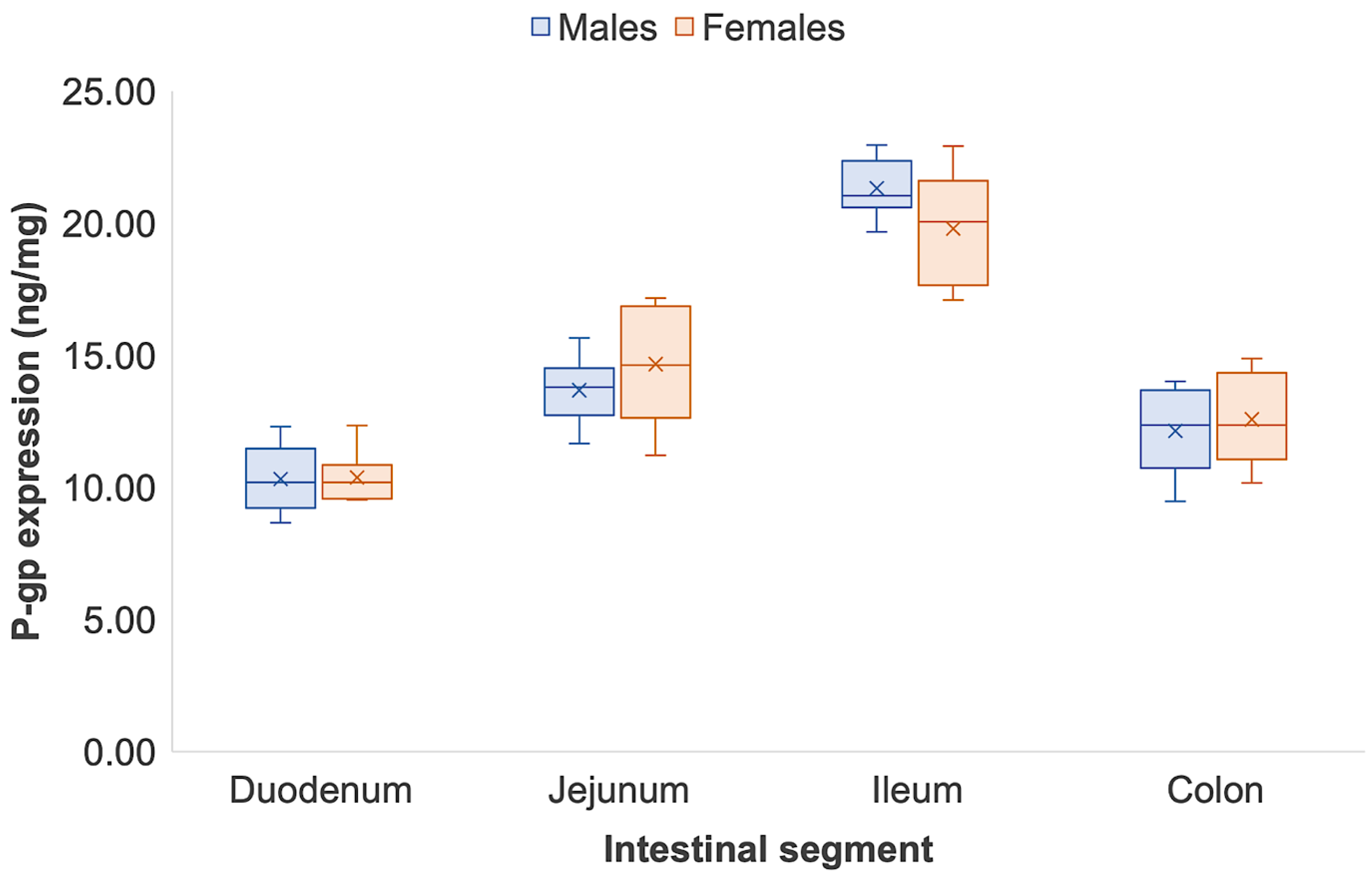
3.2. Intestinal P-glycoprotein Quantification in Sprague Dawley Rats
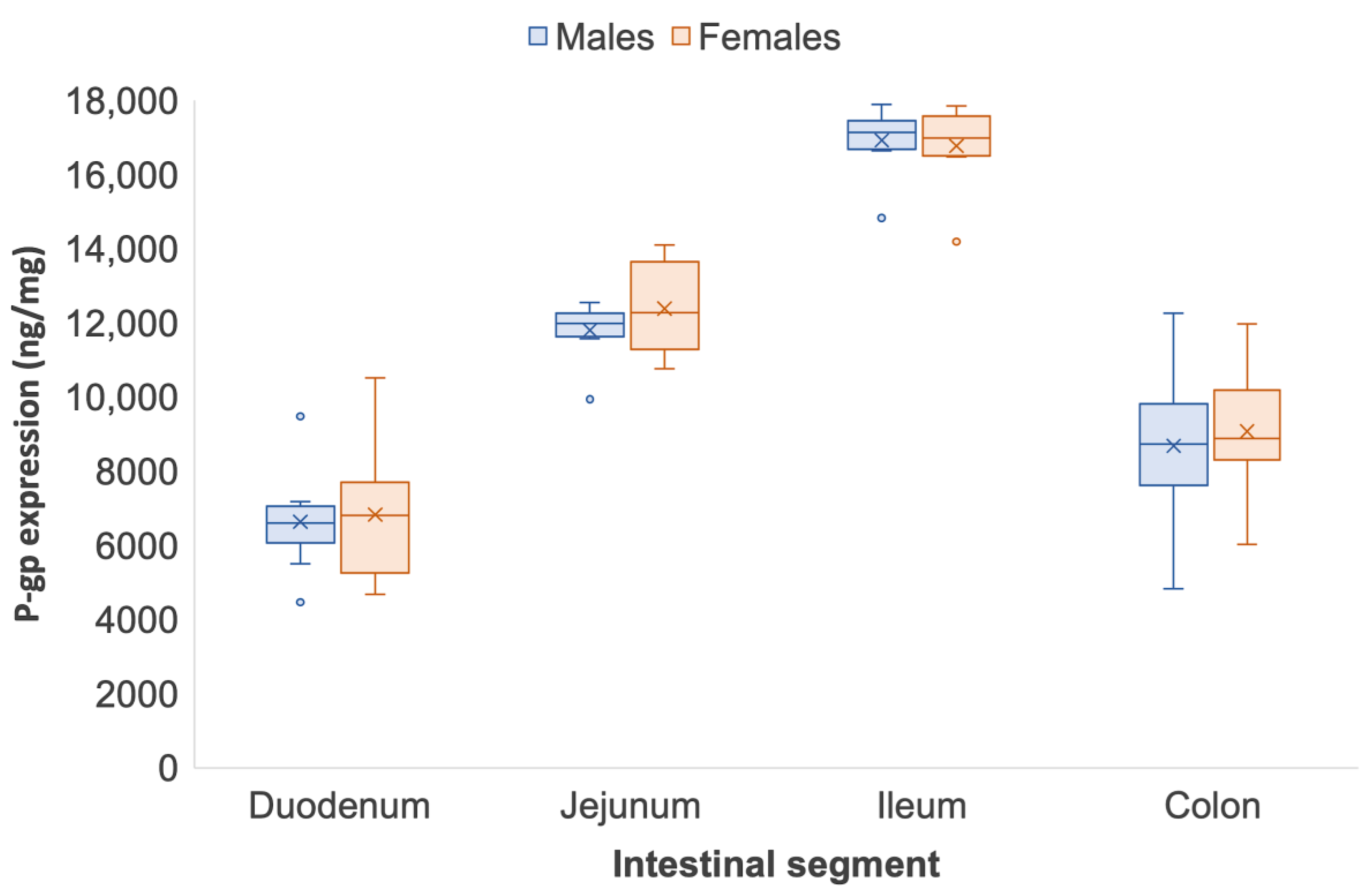
3.3. Strain Differences in Intestinal P-glycoprotein between Wistar and Sprague Dawley Rats
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Singh, V.K.; Seed, T.M. How necessary are animal models for modern drug discovery? Expert Opin. Drug Discov. 2021, 16, 1391–1397. [Google Scholar] [CrossRef] [PubMed]
- Vinarov, Z.; Abrahamsson, B.; Artursson, P.; Batchelor, H.; Berben, P.; Bernkop-Schnürch, A.; Butler, J.; Ceulemans, J.; Davies, N.; Dupont, D.; et al. Current challenges and future perspectives in oral absorption research: An opinion of the UNGAP network. Adv. Drug Deliv. Rev. 2021, 171, 289–331. [Google Scholar] [CrossRef] [PubMed]
- Szpirer, C. Rat models of human diseases and related phenotypes: A systematic inventory of the causative genes. J. Biomed. Sci. 2020, 27, 84. [Google Scholar] [CrossRef] [PubMed]
- Berkenhout, J. Outlines of the Natural History of Great Britain and Ireland, Containing a Systematic Arrangement and Concise Description of All the Animals, Vegetables and Fossiles Which Have Hitherto Been Discovered in These Kingdoms; Printed for P. Elmsly (Successor to Mr. Vaillant): London, UK, 1769. [Google Scholar]
- Tocher, C.B. The Wistar Rat as a Right Choice: Establishing Mammalian Standards and the Ideal of a Standardized Mammal. J. Hist. Biol. 1993, 26, 329–349. [Google Scholar]
- White, W.J.; Cham, S. The Development and Maintenance of the Crl: CH!! J (SD) IGSBR Rat Breeding System. Biol. Ref. Data CD (SD) IGS Rats 1998, 8–14. [Google Scholar]
- Alemáan, C.L.; Más, R.M.; Rodeiro, I.; Noa, M.; Hernández, C.; Menéndez, R.; Gámez, R. Reference database of the main physiological parameters in Sprague-Dawley rats from 6 to 32 months. Lab. Anim. 1998, 32, 457–466. [Google Scholar] [CrossRef] [Green Version]
- Musther, H.; Olivares-Morales, A.; Hatley, O.J.; Liu, B.; Hodjegan, A.R. Animal versus human oral drug bioavailability: Do they correlate? Eur. J. Pharm. Sci. 2014, 57, 280–291. [Google Scholar] [CrossRef]
- Seelig, A. P-Glycoprotein: One Mechanism, Many Tasks and the Consequences for Pharmacotherapy of Cancers. Front. Oncol. 2020, 10, 576559. [Google Scholar] [CrossRef]
- Terada, T.; Hira, D. Intestinal and hepatic drug transporters: Pharmacokinetic, pathophysiological, and pharmacogenetic roles. J. Gastroenterol. 2015, 50, 508–519. [Google Scholar] [CrossRef] [Green Version]
- Goebel, J.; Chmielewski, J.; Hrycyna, C.A. The roles of the human ATP-binding cassette transporters P-glycoprotein and ABCG2 in multidrug resistance in cancer and at endogenous sites: Future opportunities for structure-based drug design of inhibitors. Cancer Drug Resist 2021, 4, 784–804. [Google Scholar] [CrossRef]
- Nguyen, T.-T.; Duong, V.-A.; Maeng, H.-J. Pharmaceutical Formulations with P-Glycoprotein Inhibitory Effect as Promising Approaches for Enhancing Oral Drug Absorption and Bioavailability. Pharmaceutics 2021, 13, 1103. [Google Scholar] [CrossRef] [PubMed]
- FDA. Drug Development and Drug Interactions/Table of Substrates, Inhibitors and Inducers. 2020. Available online: https://www.fda.gov/drugs/drug-interactions-labeling/drug-development-and-drug-interactions-table-substrates-inhibitors-and-inducers (accessed on 27 March 2022).
- FDA. Clinical Drug Interaction Studies—Cytochrome P450 Enzyme- and Transporter-Mediated Drug Interactions Guidance for Industry; FDA: Silver Spring, MD, USA, 2020. [Google Scholar]
- Dou, L.; Gavins, F.K.H.; Mai, Y.; Madla, C.M.; Taherali, F.; Orlu, M.; Murdan, S.; Basit, A.W. Effect of Food and an Animal’s Sex on P-Glycoprotein Expression and Luminal Fluids in the Gastrointestinal Tract of Wistar Rats. Pharmaceutics 2020, 12, 296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dou, L.; Mai, Y.; Madla, C.M.; Orlu, M.; Basit, A.W. P-glycoprotein expression in the gastrointestinal tract of male and female rats is influenced differently by food. Eur. J. Pharm. Sci. 2018, 123, 569–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacLean, C.; Moenning, U.; Reichel, A.; Fricker, G. Closing the Gaps: A Full Scan of the Intestinal Expression of P-Glycoprotein, Breast Cancer Resistance Protein, and Multidrug Resistance-Associated Protein 2 in Male and Female Rats. Drug Metab. Dispos. 2008, 36, 1249–1254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mai, Y.; Dou, L.; Yao, Z.; Madla, C.M.; Gavins, F.K.H.; Taherali, F.; Yin, H.; Orlu, M.; Murdan, S.; Basit, A.W. Quantification of P-Glycoprotein in the Gastrointestinal Tract of Humans and Rodents: Methodology, Gut Region, Sex, and Species Matter. Mol. Pharm. 2021, 18, 1249–1254. [Google Scholar] [CrossRef]
- Pu, J.; An, B.; Vazvaei, F.; Qu, J. Enrichment of protein therapeutics and biomarkers for LC–MS quantification. Bioanalysis 2018, 10, 979–982. [Google Scholar] [CrossRef]
- Gandhi, A.S.; Budac, D.; Khayrullina, T.; Staal, R.; Chandrasena, G. Quantitative analysis of lipids: A higher-throughput LC–MS/MS-based method and its comparison to ELISA. Futur. Sci. OA 2017, 3, FSO157. [Google Scholar] [CrossRef] [Green Version]
- Genovese, I.; Ilari, A.; Assaraf, Y.G.; Fazi, F.; Colotti, G. Not only P-glycoprotein: Amplification of the ABCB1- containing chromosome region 7q21 confers multidrug resistance upon cancer cells by coordinated overexpression of an assortment of resistance-related proteins. Drug Resist. Updat. 2017, 32, 23–46. [Google Scholar] [CrossRef]
- Ferreira, G.S.; Veening-Griffioen, D.H.; Boon, W.P.C.; Moors, E.H.M.; Wied, C.C.G.; Schellekens, H.; van Meer, P.J.K. A standardised framework to identify optimal animal models for efficacy assessment in drug development. PLoS ONE 2019, 14, e0218014. [Google Scholar] [CrossRef] [Green Version]
- Clarke, L.L. A guide to Ussing chamber studies of mouse intestine. Am. J. Physiol. Gastrointest Liver Physiol. 2009, 296, G1151–G1166. [Google Scholar] [CrossRef] [Green Version]
- Gröer, C.; Brück, S.; Lai, Y.; Paulick, A.; Busemann, A.; Heidecke, C.; Siegmund, W.; Oswald, S. LC–MS/MS-based quantification of clinically relevant intestinal uptake and efflux transporter proteins. J. Pharm. Biomed. Anal. 2013, 85, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Uchida, Y.; Tachikawa, M.; Obuchi, W.; Hoshi, Y.; Tomioka, Y.; Ohtsuki, S.; Terasaki, T. A study protocol for quantitative targeted absolute proteomics (QTAP) by LC-MS/MS: Application for inter-strain differences in protein expression levels of transporters, receptors, claudin-5, and marker proteins at the blood–brain barrier in ddY, FVB, and C57BL/6J mice. Fluids Barriers CNS 2013, 10, 21–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musther, H.; Harwood, M.; Yang, J.; Turner, D.B.; Rostami-Hodjegan, A.; Jamei, M. The Constraints, Construction, and Verification of a Strain-Specific Physiologically Based Pharmacokinetic Rat Model. J. Pharm. Sci. 2017, 106, 2826–2838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brower, M.; Grace, M.; Kotz, C.M.; Koya, V. Comparative analysis of growth characteristics of Sprague Dawley rats obtained from different sources. Lab. Anim. Res. 2015, 31, 166–173. [Google Scholar] [CrossRef] [Green Version]
- Rybnikova, E.A.; Vetrovoi, O.V.; Zenko, M.Y. Comparative Characterization of Rat Strains (Wistar, Wistar–Kyoto, Sprague Dawley, Long Evans, LT, SHR, BD-IX) by Their Behavior, Hormonal Level and Antioxidant Status. J. Evol. Biochem. Physiol. 2018, 54, 374–382. [Google Scholar] [CrossRef]
- Oltra-Noguera, D.; Mangas-Sanjuan, V.; González-Álvarez, I.; Colon-Useche, S.; González-Álvarez, M.; Bermejo, M. Drug gastrointestinal absorption in rat: Strain and gender differences. Eur. J. Pharm. Sci. 2015, 78, 198–203. [Google Scholar] [CrossRef]
- Liu, H.; Sun, H.; Wu, Z.; Zhang, X.; Wu, B. P-glycoprotein (P-gp)-mediated efflux limits intestinal absorption of the Hsp90 inhibitor SNX-2112 in rats. Xenobiotica 2014, 44, 763–768. [Google Scholar] [CrossRef]
- Madla, C.M.; Gavins, F.K.; Merchant, H.A.; Orlu, M.; Murdan, S.; Basit, A.W. Let’s talk about sex: Differences in drug therapy in males and females. Adv. Drug Deliv. Rev. 2021, 175, 113804. [Google Scholar] [CrossRef]
- Beery, A.; Zucker, I. Sex bias in neuroscience and biomedical research. Neurosci. Biobehav. Rev. 2011, 35, 565–572. [Google Scholar] [CrossRef] [Green Version]
- Woitowich, N.C.; Beery, A.; Woodruff, T. A 10-year follow-up study of sex inclusion in the biological sciences. eLife 2020, 9, e56344. [Google Scholar] [CrossRef]
- Clark, J.M. The 3Rs in research: A contemporary approach to replacement, reduction and refinement. Br. J. Nutr. 2017, 120, S1–S7. [Google Scholar] [CrossRef] [PubMed]
- Afonso-Pereira, F.; Murdan, S.; Sousa, J.; Veiga, F.; Basit, A.W. Sex differences in excipient effects: Enhancement in ranitidine bioavailability in the presence of polyethylene glycol in male, but not female, rats. Int. J. Pharm. 2016, 506, 237–241. [Google Scholar] [CrossRef] [PubMed]
- Mai, Y.; Dou, L.; Murdan, S.; Basit, A. An animal’s sex influences the effects of the excipient PEG 400 on the intestinal P-gp protein and mRNA levels, which has implications for oral drug absorption. Eur. J. Pharm. Sci. 2018, 120, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Mai, Y.; Dou, L.; Madla, C.M.; Murdan, S.; Basit, A.W. Sex-Dependence in the Effect of Pharmaceutical Excipients: Polyoxyethylated Solubilising Excipients Increase Oral Drug Bioavailability in Male but Not Female Rats. Pharmaceutics 2019, 11, 228. [Google Scholar] [CrossRef] [Green Version]
- Ashiru, D.A.I.; Patel, R.; Basit, A.W. Polyethylene Glycol 400 Enhances the Bioavailability of a BCS Class III Drug (Ranitidine) in Male Subjects but Not Females. Pharm. Res. 2008, 25, 2327–2333. [Google Scholar] [CrossRef]
- Mai, Y.; Ashiru-Oredope, D.A.; Yao, Z.; Dou, L.; Madla, C.M.; Taherali, F.; Murdan, S.; Basit, A.W. Boosting drug bioavailability in men but not women through the action of an excipient. Int. J. Pharm. 2020, 587, 119678. [Google Scholar] [CrossRef]
- Mariana, B.; Adrián, L.; Guillermo, V.; Juan, S.; Laura, M.; Carlos, L. Gender-related differences on P-glycoprotein-mediated drug intestinal transport in rats. J. Pharm. Pharmacol. 2011, 63, 619–626. [Google Scholar] [CrossRef]
- Yin, H.; Shao, H.; Liu, J.; Qin, Y.; Deng, W. Sex-specific and concentration-dependent influence of Cremophor RH 40 on ampicillin absorption via its effect on intestinal membrane transporters in rats. PLoS ONE 2022, 17, e0263692. [Google Scholar] [CrossRef]
- Hua, S. Advances in Oral Drug Delivery for Regional Targeting in the Gastrointestinal Tract—Influence of Physiological, Pathophysiological and Pharmaceutical Factors. Front. Pharmacol. 2020, 11, 524. [Google Scholar] [CrossRef]
- Mennen, S.M.; Alhambra, C.; Allen, C.L.; Barberis, M.; Berritt, S.; Brandt, T.A.; Campbell, A.D.; Castañón, J.; Cherney, A.H.; Christensen, M.; et al. The Evolution of High-Throughput Experimentation in Pharmaceutical Development and Perspectives on the Future. Org. Process Res. Dev. 2019, 23, 1213–1242. [Google Scholar] [CrossRef]
- Tighe, P.; Ryder, R.R.; Todd, I.; Fairclough, L.C. ELISA in the multiplex era: Potentials and pitfalls. Proteom. Clin. Appl. 2015, 9, 406–422. [Google Scholar] [CrossRef] [PubMed]
- Myasein, F.; Kim, E.; Zhang, J.; Wu, H.; El-Shourbagy, T.A. Rapid, simultaneous determination of lopinavir and ritonavir in human plasma by stacking protein precipitations and salting-out assisted liquid/liquid extraction, and ultrafast LC-MS/MS. Anal. Chim. Acta 2009, 651, 112–116. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.S.; Ji, Q.; Zhang, J.; El-Shourbagy, T.A. Historical review of sample preparation for chromatographic bioanalysis: Pros and cons. Drug Dev. Res. 2007, 68, 107–133. [Google Scholar] [CrossRef]
- Shen, Y.; Prinyawiwatkul, W.; Xu, Z. Insulin: A review of analytical methods. Analyst 2019, 144, 4139–4148. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, L.; Yu, N.; Dai, L.; Stella, C.; Chang, V.; Kaur, S.; Xu, K.; Wakshull, E. Immunoaffinity LC–MS/MS is more suitable than ELISA to quantify a PEGylated molecule in cynomolgus monkey serum. Bioanalysis 2020, 12, 1061–1069. [Google Scholar] [CrossRef]
- Sauer, F.J.; Gerber, V.; Frei, S.; Bruckmaier, R.M.; Groessl, M. Salivary cortisol measurement in horses: Immunoassay or LC-MS/MS? Domest. Anim. Endocrinol. 2020, 72, 106445. [Google Scholar] [CrossRef]
- Kotapati, S.; Deshpande, M.; Jashnani, A.; Thakkar, D.; Xu, H.; Dollinger, G. The role of ligand-binding assay and LC–MS in the bioanalysis of complex protein and oligonucleotide therapeutics. Bioanalysis 2021, 13, 931–954. [Google Scholar] [CrossRef]
- Wang, S.J.; Wu, S.T.; Gokemeijer, J.; Fura, A.; Krishna, M.; Morin, P.; Chen, G.; Price, K.; Wang-Iverson, D.; Olah, T.; et al. Attribution of the discrepancy between ELISA and LC-MS/MS assay results of a PEGylated scaffold protein in post-dose monkey plasma samples due to the presence of anti-drug antibodies. Anal. Bioanal. Chem. 2012, 402, 1229–1239. [Google Scholar] [CrossRef]
- Chieli, E.; Romiti, N.; Cervelli, F.; Paolicchi, A.; Tongiani, R. Influence of rat strain on P-glycoprotein expression in cultured hepatocytes. Cell Biol. Toxicol. 1994, 10, 163–166. [Google Scholar] [CrossRef]
- Schiebinger, L.; Klinge, I. Gendered Innovations 2: How Inclusive Analysis Contributes to Research and Innovation. 2020. Available online: https://ec.europa.eu/info/sites/default/files/research_and_innovation/strategy_on_research_and_innovation/documents/ki0320108enn_final.pdf (accessed on 24 March 2022).
- Lai, J.-I.; Tseng, Y.-J.; Chen, M.-H.; Huang, C.-Y.F.; Chang, P.M.-H. Clinical Perspective of FDA Approved Drugs with P-Glycoprotein Inhibition Activities for Potential Cancer Therapeutics. Front. Oncol. 2020, 10, 561936. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Molecule | Peptide Sequence | Mass | Transition Number | Q1 m/z | Q1-CE | Q3 m/z | Q3-CE |
|---|---|---|---|---|---|---|---|
| ABCB1 (P-gp) | AGAVAEEVLAAIR (surrogate peptide) | 1268.7 | 1 | 635.3 | 30 | 771.3 | 30 |
| 2 | 635.3 | 30 | 900.5 | 30 | |||
| 3 | 635.3 | 30 | 971.6 | 30 | |||
| AGAVAEEVLAAIR a (internal standard) | 1278.6 | 1 | 640.3 | 30 | 781.4 | 30 | |
| 2 | 640.3 | 30 | 910.5 | 30 | |||
| 3 | 640.3 | 30 | 981.5 | 30 |
| Intestinal P-gp in Male Wistar Rats | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Quantified by ELISA (ng/mg) (n = 8) | ||||||||||
| Intestinal segments | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | Mean | S.D. |
| Duodenum | 10.78 | 7.98 | 8.08 | 10.55 | 9.27 | 9.91 | 11.23 | 8.71 | 9.56 | 1.25 |
| Jejunum | 12.59 | 11.03 | 11.03 | 12.07 | 12.13 | 11.43 | 11.83 | 13.08 | 12.02 | 0.92 |
| Ileum | 12.57 | 12.51 | 13.95 | 14.12 | 14.38 | 12.67 | 13.33 | 15.16 | 13.59 | 0.97 |
| Colon | 12.06 | 9.27 | 10.52 | 9.45 | 13.35 | 9.77 | 9.82 | 10.67 | 10.61 | 1.42 |
| Quantified by LC-MS/MS (ng/mg) (n = 8) | ||||||||||
| Intestinal segments | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | Mean | S.D. |
| Duodenum | 5894.88 | 3746.03 | 5030.55 | 5634.86 | 5152.56 | 6074.69 | 5399.90 | 5524.15 | 5307.20 | 720.63 |
| Jejunum | 11,206.95 | 7938.44 | 7624.05 | 8952.71 | 9580.88 | 8787.68 | 9770.73 | 10,273.60 | 9266.88 | 1190.55 |
| Ileum | 7838.09 | 9156.47 | 10,342.37 | 12,591.57 | 9395.06 | 10,494.22 | 11,978.40 | 10,176.80 | 10,258.72 | 1525.14 |
| Colon | 7447.57 | 5741.64 | 6694.71 | 5513.41 | 6896.77 | 6474.12 | 8805.73 | 6987.12 | 6820.13 | 1026.52 |
| Intestinal P-gp in Female Wistar Rats | ||||||||||
| Quantified by ELISA (ng/mg) (n = 8) | ||||||||||
| Intestinal segments | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | Mean | S.D. |
| Duodenum | 10.52 | 11.32 | 8.45 | 9.27 | 11.99 | 10.37 | 9.99 | 7.04 | 9.87 | 1.59 |
| Jejunum | 10.80 | 10.61 | 9.57 | 9.57 | 10.31 | 10.70 | 10.49 | 9.33 | 10.17 | 0.59 |
| Ileum | 8.91 | 10.87 | 9.28 | 12.00 | 10.04 | 11.42 | 10.16 | 11.85 | 10.57 | 1.16 |
| Colon | 7.90 | 7.01 | 8.91 | 9.74 | 8.94 | 9.19 | 7.29 | 8.09 | 8.39 | 0.96 |
| Quantified by LC-MS/MS (ng/mg) (n = 8) | ||||||||||
| Intestinal segments | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | Mean | S.D. |
| Duodenum | 5280.92 | 7759.25 | 4080.69 | 4081.68 | 5595.60 | 5900.99 | 6495.27 | 5138.45 | 5541.61 | 1222.15 |
| Jejunum | 5953.21 | 6276.65 | 5625.97 | 4700.24 | 7927.81 | 7582.72 | 6552.90 | 5402.36 | 6252.73 | 1087.68 |
| Ileum | 4810.97 | 6245.36 | 5978.92 | 8828.72 | 7429.65 | 7048.28 | 7631.05 | 7159.52 | 6672.04 | 1310.89 |
| Colon | 4131.86 | 3975.40 | 5079.73 | 4132.10 | 5341.54 | 4783.79 | 3752.69 | 4351.89 | 4443.62 | 563.90 |
| Intestinal P-gp in Male Sprague Dawley Rats | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Quantified by ELISA (ng/mg) (n = 8) | ||||||||||
| Intestinal segments | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | Mean | S.D. |
| Duodenum | 10.13 | 8.67 | 12.32 | 9.09 | 9.68 | 10.71 | 10.28 | 11.75 | 10.33 | 1.25 |
| Jejunum | 14.38 | 12.67 | 15.67 | 12.97 | 14.18 | 14.60 | 13.43 | 11.67 | 13.69 | 1.26 |
| Ileum | 21.07 | 19.67 | 20.58 | 20.71 | 22.43 | 21.03 | 22.97 | 22.19 | 21.33 | 1.10 |
| Colon | 9.49 | 10.66 | 13.51 | 14.02 | 12.21 | 11.02 | 12.52 | 13.75 | 12.15 | 1.63 |
| Quantified by LC-MS/MS (ng/mg) (n = 8) | ||||||||||
| Intestinal segments | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | Mean | S.D. |
| Duodenum | 5498.43 | 4463.30 | 9478.77 | 6244.87 | 7007.73 | 6393.02 | 6805.60 | 7172.54 | 6633.03 | 1452.23 |
| Jejunum | 12,154.04 | 9940.48 | 12,551.53 | 11,652.34 | 11,578.94 | 12,553.57 | 12,129.70 | 11,834.41 | 11,799.37 | 836.35 |
| Ileum | 17,073.66 | 14,931.03 | 17,894.74 | 16,642.60 | 17,684.77 | 17,182.61 | 17,375.64 | 16,694.31 | 16,314.93 | 2042.77 |
| Colon | 4821.69 | 6249.81 | 11,198.55 | 12,258.88 | 9356.98 | 8422.04 | 8074.38 | 8553.51 | 8679.00 | 2419.33 |
| Intestinal P-gp in Female Sprague Dawley Rats | ||||||||||
| Quantified by ELISA (ng/mg) (n = 8) | ||||||||||
| Intestinal segments | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | Mean | S.D. |
| Duodenum | 12.35 | 10.12 | 9.66 | 9.54 | 10.93 | 10.26 | 10.65 | 9.56 | 10.38 | 0.94 |
| Jejunum | 15.20 | 14.07 | 16.51 | 11.23 | 16.98 | 14.01 | 17.19 | 12.20 | 14.67 | 2.21 |
| Ileum | 21.12 | 19.17 | 17.63 | 17.09 | 17.78 | 22.94 | 20.97 | 21.78 | 19.81 | 2.19 |
| Colon | 12.08 | 10.84 | 14.53 | 11.74 | 10.18 | 13.78 | 12.64 | 14.89 | 12.59 | 1.71 |
| Quantified by LC-MS/MS (ng/mg) (n = 8) | ||||||||||
| Intestinal segments | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | Mean | S.D. |
| Duodenum | 10,509.45 | 6445.07 | 5168.71 | 4678.34 | 5483.31 | 7807.17 | 7162.20 | 7347.60 | 6825.23 | 1860.21 |
| Jejunum | 12,708.23 | 11,088.31 | 14,104.71 | 12,248.01 | 12,305.06 | 13,957.70 | 10,766.36 | 11,868.85 | 12,380.90 | 1202.86 |
| Ileum | 17,848.27 | 17,492.35 | 14,189.24 | 16,474.53 | 16,597.27 | 17,608.26 | 17,010.48 | 16,952.30 | 16,136.16 | 2071.22 |
| Colon | 8845.54 | 6019.95 | 11,973.16 | 8319.22 | 9703.36 | 10,345.87 | 8920.06 | 8553.51 | 9072.58 | 1724.76 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Madla, C.M.; Qin, Y.; Gavins, F.K.H.; Liu, J.; Dou, L.; Orlu, M.; Murdan, S.; Mai, Y.; Basit, A.W. Sex Differences in Intestinal P-Glycoprotein Expression in Wistar versus Sprague Dawley Rats. Pharmaceutics 2022, 14, 1030. https://doi.org/10.3390/pharmaceutics14051030
Madla CM, Qin Y, Gavins FKH, Liu J, Dou L, Orlu M, Murdan S, Mai Y, Basit AW. Sex Differences in Intestinal P-Glycoprotein Expression in Wistar versus Sprague Dawley Rats. Pharmaceutics. 2022; 14(5):1030. https://doi.org/10.3390/pharmaceutics14051030
Chicago/Turabian StyleMadla, Christine M., Yujia Qin, Francesca K. H. Gavins, Jing Liu, Liu Dou, Mine Orlu, Sudaxshina Murdan, Yang Mai, and Abdul W. Basit. 2022. "Sex Differences in Intestinal P-Glycoprotein Expression in Wistar versus Sprague Dawley Rats" Pharmaceutics 14, no. 5: 1030. https://doi.org/10.3390/pharmaceutics14051030
APA StyleMadla, C. M., Qin, Y., Gavins, F. K. H., Liu, J., Dou, L., Orlu, M., Murdan, S., Mai, Y., & Basit, A. W. (2022). Sex Differences in Intestinal P-Glycoprotein Expression in Wistar versus Sprague Dawley Rats. Pharmaceutics, 14(5), 1030. https://doi.org/10.3390/pharmaceutics14051030