
Antibacterial Activity from Momordica charantia L. Leaves and Flavones Enriched Phase

 ,
,  , , ,
, , ,
Abstract
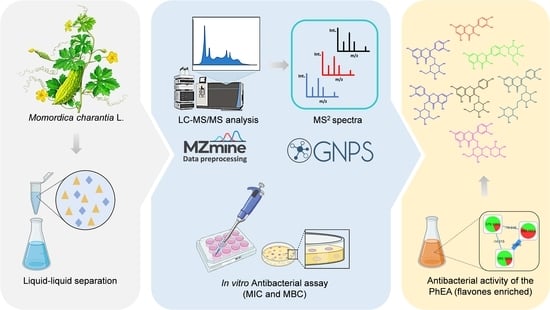
:
1. Introduction
2. Materials and Methods
2.1. Botanical Material and Extraction
2.2. Liquid Chromatography-High Resolution Mass Spectrometry Analysis
2.3. Mass Spectrum Data Treatment
2.4. Molecular Networks
2.5. In Vitro Antibacterial Assay
2.6. Determination of Minimum Inhibitory Concentration (MIC) and Minimum Bactericidal Concentration (MBC)
3. Results and Discussion
3.1. Antibacterial Activity
3.2. Identification of Chemical Constituents
3.3. Molecular Networking (GNPS Annotation)
3.4. Bioactivity and Structure
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Who No Time to Wait: Securing the Future from Drug-Resistant Infections. Available online: https://www.who.int/antimicrobial-resistance/interagency-coordination-group/final-report/en/ (accessed on 31 December 2020).
- O’Neill, J. Tackling Drug-Resistant Infections Globally: Final Report and Recommendations; The Government of the United Kingdom: London, UK, 2016. [Google Scholar]
- Ali Mirza, S.; Afzaal, M.; Begum, S.; Arooj, T.; Almas, M.; Ahmed, S.; Younus, M. Chapter 11-Uptake Mechanism of Antibiotics in Plants. In Antibiotics and Antimicrobial Resistance Genes in the Environment; Hashmi, M.Z., Ed.; Elsevier: Amsterdam, The Netherlands, 2020; Volume 1, pp. 183–188. ISBN 9780128188828. [Google Scholar]
- Michelin, D.C.; Moreschi, P.E.; Lima, A.C.; Nascimento, G.G.F.; Paganelli, M.O.; Chaud, M.V. Avaliação da atividade antimicrobiana de extratos vegetais. Rev. Bras. Farmacogn. 2005, 15, 316–320. [Google Scholar] [CrossRef]
- Guarniz, W.A.S.; Canuto, K.M.; Ribeiro, P.R.V.; Dodou, H.V.; Magalhaes, K.N.; Sá, K.; do Nascimento, P.G.G.; Silva, K.L.; Sales, G.W.P.; Monteiro, M.P.; et al. Momordica Charantia L. Variety from Northeastern Brazil: Analysis of Antimicrobial Activity and Phytochemical Components. Pharmacogn. J. 2019, 11, 1312–1324. [Google Scholar] [CrossRef]
- Lorenzi, H. Plantas Daninhas Do Brasil: Terrestres, Aquáticas, Parasitas e Tóxicas. 3\textordfeminine Edição. Inst. Plant. Bras. 2000, 309, 640. [Google Scholar]
- Jia, S.; Shen, M.; Zhang, F.; Xie, J. Recent Advances in Momordica Charantia: Functional Components and Biological Activities. Int. J. Mol. Sci. 2017, 18, 2555. [Google Scholar] [CrossRef]
- Bortolotti, M.; Mercatelli, D.; Polito, L. Momordica Charantia, a Nutraceutical Approach for Inflammatory Related Diseases. Front. Pharmacol. 2019, 10, 486. [Google Scholar] [CrossRef]
- Gürdal, B.; Kültür, Ş. An Ethnobotanical Study of Medicinal Plants in Marmaris (Muğla, Turkey). J. Ethnopharmacol. 2013, 146, 113–126. [Google Scholar] [CrossRef] [PubMed]
- Polito, L.; Bortolotti, M.; Maiello, S.; Battelli, M.; Bolognesi, A. Plants Producing Ribosome-Inactivating Proteins in Traditional Medicine. Molecules 2016, 21, 1560. [Google Scholar] [CrossRef]
- Chen, J.; Tian, R.; Qiu, M.; Lu, L.; Zheng, Y.; Zhang, Z. Trinorcucurbitane and Cucurbitane Triterpenoids from the Roots of Momordica Charantia. Phytochemistry 2008, 69, 1043–1048. [Google Scholar] [CrossRef]
- Polito, L.; Djemil, A.; Bortolotti, M. Plant Toxin-Based Immunotoxins for Cancer Therapy: A Short Overview. Biomedicines 2016, 4, 12. [Google Scholar] [CrossRef]
- Grover, J.K.; Yadav, S.P. Pharmacological Actions and Potential Uses of Momordica Charantia: A Review. J. Ethnopharmacol. 2004, 93, 123–132. [Google Scholar] [CrossRef]
- Raman, A.; Lau, C. Anti-Diabetic Properties and Phytochemistry of Momordica Charantia L. (Cucurbitaceae). Phytomedicine 1996, 2, 349–362. [Google Scholar] [CrossRef]
- Bailey, C.J.; Day, C.; Leatherdale, B.A. Traditional Treatments for Diabetes from Asia and the West Indies. Pract. Diabetes Int. 1986, 3, 190–192. [Google Scholar] [CrossRef]
- Dans, A.M.L.; Villarruz, M.V.C.; Jimeno, C.A.; Javelosa, M.A.U.; Chua, J.; Bautista, R.; Velez, G.G.B. The Effect of Momordica Charantia Capsule Preparation on Glycemic Control in Type 2 Diabetes Mellitus Needs Further Studies. J. Clin. Epidemiol. 2007, 60, 554–559. [Google Scholar] [CrossRef] [PubMed]
- Hussein, R.A.; El-Anssary, A.A. Plants Secondary Metabolites: The Key Drivers of the Pharmacological Actions of Medicinal Plants. In Herbal Medicine; IntechOpen: London, UK, 2019. [Google Scholar]
- Lei, Z.; Huhman, D.V.; Sumner, L.W. Mass Spectrometry Strategies in Metabolomics. J. Biol. Chem. 2011, 286, 25435–25442. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.W.; Choi, S.Y.; Jang, H.S.; Ryu, B.; Sung, S.H.; Yang, H. Exploring Novel Secondary Metabolites from Natural Products Using Pre-Processed Mass Spectral Data. Sci. Rep. 2019, 9, 17430. [Google Scholar] [CrossRef]
- Graça, G.; Cai, Y.; Lau, C.-H.E.; Vorkas, P.A.; Lewis, M.R.; Want, E.J.; Herrington, D.; Ebbels, T.M.D. Automated Annotation of Untargeted All-Ion Fragmentation LC—MS Metabolomics Data with MetaboAnnotatoR. Anal. Chem. 2022, 94, 3446–3455. [Google Scholar] [CrossRef]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and Community Curation of Mass Spectrometry Data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef]
- Gomes, P.W.P.; Barretto, H.; Reis, J.D.E.; Muribeca, A.; Veloso, A.; Albuquerque, C.; Teixeira, A.; Braamcamp, W.; Pamplona, S.; Silva, C.; et al. Chemical Composition of Leaves, Stem, and Roots of Peperomia Pellucida (L.) Kunth. Molecules 2022, 27, 1847. [Google Scholar] [CrossRef]
- Gomes, P.; Quirós-Guerrero, L.; Muribeca, A.; Reis, J.; Pamplona, S.; Lima, A.H.; Trindade, M.; Silva, C.; Souza, J.N.S.; Boutin, J.; et al. Constituents of Chamaecrista Diphylla (L.) Greene Leaves with Potent Antioxidant Capacity: A Feature-Based Molecular Network Dereplication Approach. Pharmaceutics 2021, 13, 681. [Google Scholar] [CrossRef] [PubMed]
- Gomes, P.W.P.; Pamplona, T.C.D.L.; Navegantes-Lima, K.C.; Quadros, L.B.G.; Oliveira, A.L.B.; Santos, S.M.; e Silva, C.Y.Y.; Silva, M.J.C.; Souza, J.N.S.; Quirós-Guerrero, L.M.; et al. Chemical Composition and Antibacterial Action of Stryphnodendron Pulcherrimum Bark Extract, “Barbatimão” Species: Evaluation of Its Use as a Topical Agent. Arab. J. Chem. 2021, 14, 103183. [Google Scholar] [CrossRef]
- Gomes, P.; Quirós-Guerrero, L.; Silva, C.; Pamplona, S.; Boutin, J.A.; Eberlin, M.; Wolfender, J.-L.; Silva, M. Feature-Based Molecular Network-Guided Dereplication of Natural Bioactive Products from Leaves of Stryphnodendron Pulcherrimum (Willd.) Hochr. Metabolites 2021, 11, 281. [Google Scholar] [CrossRef] [PubMed]
- Santiago, J.C.C.; Albuquerque, C.A.B.; Muribeca, A.d.J.B.; Sá, P.R.C.; Pamplona, S.d.G.S.R.; Silva, C.Y.Y.e.; Ribera, P.C.; Fontes-Júnior, E.d.A.; da Silva, M.N. Margaritaria Nobilis L.f. (Phyllanthaceae): Ethnopharmacology and Application of Computational Tools in the Annotation of Bioactive Molecules. Metabolites 2022, 12, 681. [Google Scholar] [CrossRef] [PubMed]
- Holman, J.D.; Tabb, D.L.; Mallick, P. Employing ProteoWizard to Convert Raw Mass Spectrometry Data. Curr. Protoc. Bioinform. 2014, 46, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Oresic, M. MZmine 2: Modular Framework for Processing, Visualizing, and Analyzing Mass Spectrometry-Based Molecular Profile Data. BMC Bioinform. 2010, 11, 395. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Lee, S.; van Santen, J.A.; Farzaneh, N.; Liu, D.Y.; Pye, C.R.; Baumeister, T.U.H.; Wong, W.R.; Linington, R.G. NP Analyst: An Open Online Platform for Compound Activity Mapping. ACS Cent. Sci. 2022, 8, 223–234. [Google Scholar] [CrossRef] [PubMed]
- CLSI M100-S11. Performance Standards for Antimicrobial Susceptibility Testing. Clin. Microbiol. Newsl. 2001, 23, 49. [Google Scholar] [CrossRef]
- Tortora, G.J.; Funke, B.R.; Case, C.L.; Weber, D.; Bair, W. Microbiology, 13th ed.; Pearson: London, UK, 2018; ISBN 9780134605180. [Google Scholar]
- Osonga, F.J.; Akgul, A.; Miller, R.M.; Eshun, G.B.; Yazgan, I.; Akgul, A.; Sadik, O.A. Antimicrobial Activity of a New Class of Phosphorylated and Modified Flavonoids. ACS Omega 2019, 4, 12865–12871. [Google Scholar] [CrossRef]
- Kuete, V. Potential of Cameroonian Plants and Derived Products against Microbial Infections: A Review. Planta Med. 2010, 76, 1479–1491. [Google Scholar] [CrossRef]
- Jabeen, U.; Khanum, A. Isolation and Characterization of Potential Food Preservative Peptide from Momordica Charantia L. Arab. J. Chem. 2017, 10, S3982–S3989. [Google Scholar] [CrossRef]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.-M.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed Minimum Reporting Standards for Chemical Analysis. Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amaral, J.G.; Bauermeister, A.; Pilon, A.C.; Gouvea, D.R.; Sakamoto, H.T.; Gobbo-Neto, L.; Lopes, J.L.C.; Lopes, N.P. Fragmentation Pathway and Structural Characterization of New Glycosylated Phenolic Derivatives from Eremanthus Glomerulatus Less (Asteraceae) by Electrospray Ionization Tandem Mass Spectrometry. J. Mass Spectrom. 2017, 52, 783–787. [Google Scholar] [CrossRef] [PubMed]
- Bittremieux, W.; Avalon, N.E.; Thomas, S.P.; Kakhkhorov, S.A.; Aksenov, A.A.; Gomes, P.W.P.; Aceves, C.M.; Rodríguez, A.M.C.; Gauglitz, J.M.; Gerwick, W.H.; et al. Open Access Repository-Scale Propagated Nearest Neighbor Suspect Spectral Library for Untargeted Metabolomics. bioRxiv 2022. [Google Scholar] [CrossRef]
- Li, A.; Hou, X.; Wei, Y. Fast Screening of Flavonoids from Switchgrass and Mikania Micrantha by Liquid Chromatography Hybrid-Ion Trap Time-of-Flight Mass Spectrometry. Anal. Methods 2018, 10, 109–122. [Google Scholar] [CrossRef]
- Colombo, R.; Yariwake, J.H.; McCullagh, M. Study of C- and O-Glycosylflavones in Sugarcane Extracts Using Liquid Chromatography: Exact Mass Measurement Mass Spectrometry. J. Braz. Chem. Soc. 2008, 19, 483–490. [Google Scholar] [CrossRef]
- March, R.; Brodbelt, J. Analysis of Flavonoids: Tandem Mass Spectrometry, Computational Methods, and NMR. J. Mass Spectrom. 2008, 43, 1581–1617. [Google Scholar] [CrossRef]
- Kumar, S.; Singh, A.; Kumar, B. Identification and Characterization of Phenolics and Terpenoids from Ethanolic Extracts of Phyllanthus Species by HPLC-ESI-QTOF-MS/MS. J. Pharm. Anal. 2017, 7, 214–222. [Google Scholar] [CrossRef]
- Du, L.-Y.; Zhao, M.; Xu, J.; Qian, D.-W.; Jiang, S.; Shang, E.-X.; Guo, J.-M.; Duan, J.-A. Analysis of the Metabolites of Isorhamnetin 3-O-Glucoside Produced by Human Intestinal Flora in Vitro by Applying Ultraperformance Liquid Chromatography/Quadrupole Time-of-Flight Mass Spectrometry. J. Agric. Food Chem. 2014, 62, 2489–2495. [Google Scholar] [CrossRef]
- Chen, Y.; Yu, H.; Wu, H.; Pan, Y.; Wang, K.; Jin, Y.; Zhang, C. Characterization and Quantification by LC-MS/MS of the Chemical Components of the Heating Products of the Flavonoids Extract in Pollen Typhae for Transformation Rule Exploration. Molecules 2015, 20, 18352–18366. [Google Scholar] [CrossRef]
- Nagy, R.; Grob, H.; Weder, B.; Green, P.; Klein, M.; Frelet-Barrand, A.; Schjoerring, J.K.; Brearley, C.; Martinoia, E. The Arabidopsis ATP-Binding Cassette Protein AtMRP5/AtABCC5 Is a High Affinity Inositol Hexakisphosphate Transporter Involved in Guard Cell Signaling and Phytate Storage. J. Biol. Chem. 2009, 284, 33614–33622. [Google Scholar] [CrossRef]
- Desai, M.; Rangarajan, P.; Donahue, J.L.; Williams, S.P.; Land, E.S.; Mandal, M.K.; Phillippy, B.Q.; Perera, I.Y.; Raboy, V.; Gillaspy, G.E. Two Inositol Hexakisphosphate Kinases Drive Inositol Pyrophosphate Synthesis in Plants. Plant J. 2014, 80, 642–653. [Google Scholar] [CrossRef] [PubMed]
- Raboy, V. Seed Total Phosphate and Phytic Acid. In Molecular Genetic Approaches to Maize Improvement; Kriz, A.L., Larkins, B.A., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 41–53. ISBN 9783540689225. [Google Scholar]
- Freed, C.; Adepoju, O.; Gillaspy, G. Can Inositol Pyrophosphates Inform Strategies for Developing Low Phytate Crops? Plants 2020, 9, 115. [Google Scholar] [CrossRef]
- Chan, L.Y.; Wang, C.K.L.; Major, J.M.; Greenwood, K.P.; Lewis, R.J.; Craik, D.J.; Daly, N.L. Isolation and Characterization of Peptides from Momordica Cochinchinensis Seeds. J. Nat. Prod. 2009, 72, 1453–1458. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Tu, Z.; Wang, H.; Zhang, L. Ultrasound-Assisted Extraction Optimization of α-Glucosidase Inhibitors from Ceratophyllum Demersum L. and Identification of Phytochemical Profiling by HPLC-QTOF-MS/MS. Molecules 2020, 25, 4507. [Google Scholar] [CrossRef]
- Kai, H.; Baba, M.; Okuyama, T. Two New Megastigmanes from the Leaves of Cucumis Sativus. Chem. Pharm. Bull. 2007, 55, 133–136. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Krynitsky, A.J.; Grundel, E.; Rader, J.I. Quantitative Determination of Cucurbitane-Type Triterpenes and Triterpene Glycosides in Dietary Supplements Containing Bitter Melon (Momordica Charantia) by HPLC-MS/MS. J. AOAC Int. 2012, 95, 1597–1608. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, M.-J.U.; Duarte, N.; Reis, M.; Madureira, A.M.; Molnár, J. Euphorbia and Momordica Metabolites for Overcoming Multidrug Resistance. Phytochem. Rev. 2014, 13, 915–935. [Google Scholar] [CrossRef]
- Ramalhete, C.; Mansoor, T.A.; Mulhovo, S.; Molnár, J.; Ferreira, M.-J.U. Cucurbitane-Type Triterpenoids from the African Plant Momordica Balsamina. J. Nat. Prod. 2009, 72, 2009–2013. [Google Scholar] [CrossRef]
- Dixon, R.A.; Howles, P.A.; Lamb, C.; He, X.Z.; Reddy, J.T. Prospects for the Metabolic Engineering of Bioactive Flavonoids and Related Phenylpropanoid Compounds. Adv. Exp. Med. Biol. 1998, 439, 55–66. [Google Scholar] [CrossRef]
- Alcaráz, L.E.; Blanco, S.E.; Puig, O.N.; Tomás, F.; Ferretti, F.H. Antibacterial Activity of Flavonoids Against Methicillin-Resistant Staphylococcus Aureus Strains. J. Theor. Biol. 2000, 205, 231–240. [Google Scholar] [CrossRef]
- Mirzoeva, O.K.; Grishanin, R.N.; Calder, P.C. Antimicrobial Action of Propolis and Some of Its Components: The Effects on Growth, Membrane Potential and Motility of Bacteria. Microbiol. Res. 1997, 152, 239–246. [Google Scholar] [CrossRef]
- Kołpa, M.; Wałaszek, M.; Gniadek, A.; Wolak, Z.; Dobroś, W. Incidence, Microbiological Profile and Risk Factors of Healthcare-Associated Infections in Intensive Care Units: A 10 Year Observation in a Provincial Hospital in Southern Poland. Int. J. Environ. Res. Public Health 2018, 15, 112. [Google Scholar] [CrossRef] [PubMed]
- Hodek, P. Flavonoids. In Metabolism of Drugs and Other Xenobiotics; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2012; pp. 543–582. ISBN 9783527630905. [Google Scholar]
- Lin, R.-D.; Chin, Y.-P.; Lee, M.-H. Antimicrobial Activity of Antibiotics in Combination with Natural Flavonoids against Clinical Extended-Spectrum Beta-Lactamase (ESBL)-Producing Klebsiella Pneumoniae. Phytother. Res. 2005, 19, 612–617. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.; Li, Y.; Maguy, B.L.; Lou, Z.; Wang, H.; Zhao, W.; Chen, X. Separation and Enrichment of Phenolics Improved the Antibiofilm and Antibacterial Activity of the Fractions from Citrus Medica L. Var. Sarcodactylis in Vitro and in Tofu. Food Chem. 2019, 294, 533–538. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentration (µg·mL−1) | EE | PhHex | PhEA | PhWOH | EE | PhHex | PhEA | PhWOH | EE | PhHex | PhEA | PhWOH |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Klebsiella pneumoniae | Proteus mirabilis | Staphylococcus aureus | ||||||||||
| 2500 | = | + | = | + | = | + | − | + | − | + | − | + |
| 1250 | = | + | = | + | = | + | + | + | − | + | + | + |
| 625 | = | + | = | + | = | + | + | + | − | + | + | + |
| 312.5 | + | + | = | + | = | + | + | + | + | + | + | + |
| 156.2 | + | + | = | + | + | + | + | + | + | + | + | + |
| 78.1 | + | + | + | + | + | + | + | + | + | + | + | + |
| 39.0 | + | + | + | + | + | + | + | + | + | + | + | + |
| Peak | Rt (Min) | Molecular Formula | [M − H]− (m/z) | Main Product Ions (MS/MS) | Annotated Compound | EE | PhEA | ||
|---|---|---|---|---|---|---|---|---|---|
| Calculated | Accurate | Error (ppm) | |||||||
| 1 | 0.42 | C13H24O13 | 387.1139 | 387.1129 | 2.6 | 341, 278, 179 | melibiose | x | - |
| 2 | 1.63 | C12H14O8 | 285.0610 | 285.0602 | 2.8 | 153 | dihydrobenzoic acid pentose | x | x |
| 3 | 3.23 | C20H32O10 | a 431.1917 | a 431.1910 | 1.6 | 385.1856 [M − H]−, 223, 205, 163, 119, 113 | hydroxy-2,4,4-trimethyl-3-(3-oxobutyl)-2-cyclohexen-1-one glucoside | x | x |
| 4 | 3.71 | C26H28O16 | 595.1299 | 595.1306 | 1.2 | 445, 300, 272, 251, 191, 178 | quercetin-O-sambubioside | x | x |
| 5 | 3.97 | C27H30O16 | 609.1456 | 609.1458 | 0.3 | 463, 301 | rutin | x | x |
| 6 | 4.08 | C21H20O12 | 463.0877 | 463.0869 | 1.7 | 301, 271, 179 | quercetin-O-glucoside | x | x |
| 7 | 4.14 | C26H28O15 | 579.1350 | 579.1348 | 0.3 | 463, 399, 327, 285, 151, 109 | kaempferol-O-glucoside-O-pentoside | x | - |
| 8 | 4.39 | C27H30O15 | 593.1506 | 593.1511 | 0.8 | 547, 447, 357, 327, 285 | luteolin-O-rutinoside | x | x |
| 9 | 4.37 | C23H22O13 | 505.0982 | 505.0980 | 0.4 | 300, 271, 255, 243, 178, 151 | quercetin-O-glucosyl-6′′-acetate | x | x |
| 10 | 4.53 | C21H20O11 | 447.0927 | 447.0926 | 0.2 | 327, 284, 255, 227 | kaempferol-O-glucoside | x | x |
| 11 | 4.65 | C7H6O3 | 137.0239 | 137.0228 | 8.0 | 93 | 4-hydroxybenzoic acid | x | - |
| 12 | 4.67 | C22H22O12 | 477.1033 | 477.1035 | 0.4 | 431, 357, 315, 300, 285, 271, 151 | isorhamnetin-O-glucoside | x | - |
| 13 | 4.87 | C23H22O12 | 489.1038 | 489.1033 | 1.0 | 285, 255, 227 | quercetin-O-acetylpentoside | x | x |
| 14 | 4.98 | C20H34O9 | a 417.2125 | a 417.2113 | 2.9 | 371.2052 [M − H]−, 209, 161, | icariside B6 | x | x |
| 15 | 5.66 | C18H32O7 | 359.2070 | 359.2061 | 2.5 | 343, 305, 287, 239, 227, 209, 197, 171 | unknown | x | - |
| 16 | 5.77 | C15H10O7 | 301.0348 | 301.0341 | 2.3 | 273, 245, 193, 179, 151, 121 | quercetin | x | x |
| 17 | 7.14 | C18H32O5 | 327.2171 | 327.2162 | 2.7 | 291, 229, 171 | trihydroxy octadecadienoic acid isomer | x | x |
| 18 | 7.24 | C18H32O5 | 327.2171 | 327.2160 | 3.3 | 291, 229, 171 | trihydroxy octadecadienoic acid isomer | x | x |
| 19 | 7.64 | C18H34O5 | 329.2328 | 329.2320 | 2.4 | 211, 171 | trihydroxy octadecenoic acid | x | x |
| 20 | 8.38 | C37H60O11 | 679.4057 | a 679.4053 | 0.6 | 633.3994 [M − H]−, 285 | momordicoside L isomer | x | - |
| 21 | 8.58 | C18H28O4 | 307.1909 | 307.1903 | 2.0 | 289, 267, 235, 209, 185 | unknown | x | - |
| 22 | 9.06 | C39H60O13 | a 735.3956 | a 735.3963 | 0.9 | 689.3918 [M − H]−, 667, 599, 527, 339 | hederagenin base-2H + 1O, O-AcetylHex | x | - |
| 23 | 9.74 | C39H62O12 | 721.4163 | 721.4156 | 1.0 | 675 [M − H]−, 633, 513, 275, 193 | hederagenin-O-AcetylHex | x | - |
| 24 | 9.18 | C18H26O4 | 305.1753 | 305.1746 | 2.3 | 287, 249, 207 | unknown | x | - |
| 25 | 9.37 | C37H60O11 | a 679.4057 | a 679.4060 | 0.4 | 633.4015 [M − H]−, 575, 549, 471, 343 | momordicoside L isomer | x | - |
| 26 | 9.92 | C46H56O6 | 703.3999 | 703.4061 | 0.6 | 659, 633, 597, 482, 350 | unknown | x | - |
| 27 | 10.0 | C30H60O16 | a 675.3803 | a 675.3735 | 10 | 629.3677 [M − H]−, 569, 467, 447, 339, 297 | triterpene glycosides derivative | x | x |
| 28 | 10.4 | C37H60O11 | a 679.4057 | a 679.4052 | 0.7 | 633.4084 [M − H]−, 530, 339, 291, 137 | momordicoside L isomer | x | - |
| 29 | 11.51 | C18H29O3 | 293.2117 | 293.2109 | 2.7 | 275, 235, 183, 121 | unknown | x | - |
| 30 | 11.56 | C28H62O21 | 733.3705 | 733.3729 | 3.3 | 689, 554, 412, 364, 259, 175 | unknown | x | - |
| 31 | 12.0 | C36H54O10 | 645.3639 | 645.3639 | 0.0 | 601, 559, 513, 407, 339, 243, 168, 127 | unknown | x | - |
| 32 | 14.9 | C32H44O9 | 571.2907 | 571.2882 | 4.4 | 525, 481, 391, 325, 315, 255, 241, 153 | 1-Hexadecanoyl-sn-glycero-3-phospho-(1′-myo-inositol) isomer | x | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muribeca, A.d.J.B.; Gomes, P.W.P.; Paes, S.S.; da Costa, A.P.A.; Gomes, P.W.P.; Viana, J.d.S.; Reis, J.D.E.; Pamplona, S.d.G.S.R.; Silva, C.; Bauermeister, A.; et al. Antibacterial Activity from Momordica charantia L. Leaves and Flavones Enriched Phase. Pharmaceutics 2022, 14, 1796. https://doi.org/10.3390/pharmaceutics14091796
Muribeca AdJB, Gomes PWP, Paes SS, da Costa APA, Gomes PWP, Viana JdS, Reis JDE, Pamplona SdGSR, Silva C, Bauermeister A, et al. Antibacterial Activity from Momordica charantia L. Leaves and Flavones Enriched Phase. Pharmaceutics. 2022; 14(9):1796. https://doi.org/10.3390/pharmaceutics14091796
Chicago/Turabian StyleMuribeca, Abraão de Jesus B., Paulo Wender P. Gomes, Steven Souza Paes, Ana Paula Alves da Costa, Paulo Weslem Portal Gomes, Jéssica de Souza Viana, José Diogo E. Reis, Sônia das Graças Santa R. Pamplona, Consuelo Silva, Anelize Bauermeister, and et al. 2022. "Antibacterial Activity from Momordica charantia L. Leaves and Flavones Enriched Phase" Pharmaceutics 14, no. 9: 1796. https://doi.org/10.3390/pharmaceutics14091796