1. Introduction
In the past two decades of clinical testing, multiple attempts to develop viruses as oncolytic agents have yielded mixed results; while some oncolytic viruses (OVs) have made it to the market, there have also been notable failures in other developments [
1,
2]. Today, OVs continue to garner interest because of their mechanisms of action; the multimodality of OV therapy includes the lytic infection activity that exposes tumor neoantigens while simultaneously delivering transgenes for tumor-specific expression to enhance host anti-tumor immune response [
3,
4]. On top of that, recent studies have shown that OVs demonstrated remarkable synergy with the efficacious immune checkpoint inhibitors that has led some to believe that this combination therapy may become a new paradigm in treating cancers [
5,
6,
7]. Of the many OVs that have been studied, oncolytic vaccinia viruses (oVVs) have emerged as a popular choice for their ability to quickly induce oncolysis in cancer cells, and its potential to carry multiple transgenes in its viral DNA, and its short life cycle that takes place entirely in the cytoplasm lowers the risk of genomic integration [
8].
However, some obstacles remain; oVVs, being obligate intracellular parasites, are readily and rapidly cleared by a variety of host defenses [
9]. This phenomenon represents a key challenge that is especially difficult for intravenous (IV) administration; the IV administration of OVs would be highly desirable to allow these viruses to also target metastatic secondary tumors. Additionally, while oVVs are often thought of as enveloped viruses, their manufacturing process in larger scales often leads to a product that contains a mixed population of enveloped or unenveloped oVVs of which the exact proportion of each in the mix is typically not characterized. To add to that, while both enveloped and unenveloped oVV exhibit infectiousness in cancer cells, they have different physicochemical characteristics and biomolecular signatures that lend to their rapid clearance in vivo.
To address this issue, multiple OV coating/stealthing strategies have been tested including cellular, polymer, and nanoparticle modifications. Cellular stealthing has included variously sourced stem cells to deliver OVs systemically to tumors, often taking advantage of the inflammation-homing activity of mesenchymal stem cells and has been tested with adenovirus [
10,
11,
12], herpes virus [
13,
14], and vaccinia virus [
15]. Similarly, polymers such as polyethylene glycol or polyethyleneimine were evaluated for their ability to shield OVs including adenovirus [
16], measles virus [
17], vesicular stomatitis virus [
18], and vaccinia virus [
19]. Finally, nanotechnology inspired viral modifications have also been tested for the ability to protect OVs including adenovirus [
16,
20]. While some benefit is gained, the widespread adoption of these modifications has not been realized, as it is still unsure whether any of the modifications may themselves elicit immune responses that are not always immediately clear to the medical community [
21,
22,
23].
More recently, studies have shown that cell membrane-based encapsulations of adenoviruses can confer them a pseudo-envelope, even though adenoviruses are not naturally enveloped viruses. This pseudo-enveloped virus has exhibited meaningful benefits, such as prolonged circulating half-life, immune evasion, and tumor specificity. Here, we report that this can also be applied onto the conventionally enveloped oVV by coating the oVV with erythrocyte-derived membrane (EDM). By using the nonimmunogenic erythrocyte membrane as the oVV encapsulant, we can take advantage of the privileges conferred by its receptor expression, many of which allow the erythrocyte to remain in circulation for up to 3–4 months [
24,
25]. Additionally, apart from the improved circulatory retention, we observed pH responsiveness in the EDM-coated oVVs, which showed potential for the targeted and improved infectivity of the virus into cancer cells within the acidic tumor microenvironment.
2. Materials and Methods
2.1. The Cells, Viruses, and Reagents
The human small cell adenocarcinoma line A549, human breast adenocarcinoma line MCF-7, and African green monkey kidney cell line VERO E6 were obtained from the American Type Culture Collection (Manassas, VA, USA). We maintained these cell lines in Dulbecco modified Eagle’s media (DMEM) containing L-glutamine (Gibco, Waltham, MA, USA) amended with 10% fetal bovine serum (Gibco, Waltham, MA, USA) and 1X antibiotic–antimycotic (Gibco) in a humidified cell culture incubator at 37 °C and 5% CO
2. oVV derived from the Western Reserve strain with preferential tumor replication by the disruption of the thymidine kinase and vaccinia growth factor genes, with an inserted gene encoding an enhanced green fluorescent protein (eGFP) was kindly provided by Dr. Richard Koya (University of Chicago) and its construction is described in detail elsewhere [
26]. In brief, the constructed oVV-eGFP virus was then amplified on HeLa cells, lysed to collect the product, purified over the sucrose gradient to remove cellular debris, and titered. The expression of eGFP in oVV-eGFP–infected 4T1 cells was confirmed by immunofluorescence microscopy. The oVV-eGFP manufactured this way is hereafter referred to in this manuscript as “oVV”.
2.2. Derivation of EDM from Human/Mouse Whole Blood
Studies conducted in vitro and in the xenograft model were performed with erythrocyte-derived membrane (EDM) derived from human donor erythrocytes, while studies conducted in vivo with wild-type mice were performed with EDM derived from mouse erythrocytes. Both EDM materials were generated as previously described for the enrichment of cellular membrane by others [
27]. Briefly, for human erythrocytes, we procured donor Type O Rh-negative whole human blood from BioIVT (Westbury, NY, USA). The red blood cells were separated, osmotically lysed, and the membranous fraction was recovered by tangential flow filtration. In contrast, for mouse erythrocytes, the recovery of the membranous fraction was accomplished using centrifugation at 15,000 rcf. We characterized the recovered membrane for the total protein content by BCA assay following the manufacturers’ protocol (Pierce BCA Protein Assay, Thermo) and stored frozen at or below −20 °C until use.
2.3. Encapsulation Procedure
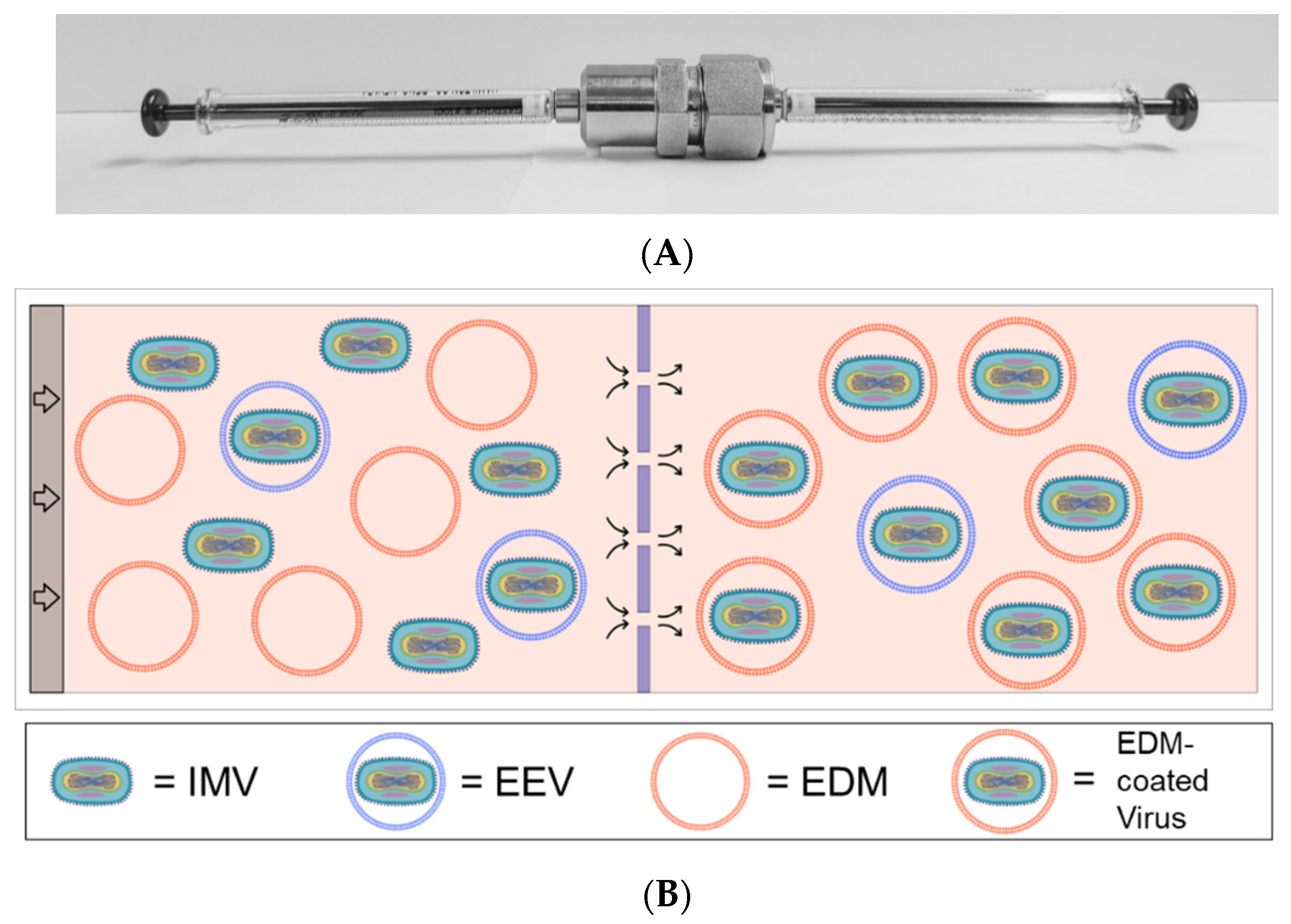
Encapsulation of oVV was achieved by fine membrane extrusion as has been previously described by others [
28,
29]. Briefly, we sanitized an extrusion apparatus consisting of two glass syringes, a membrane support housing (Avanti, Alabaster, AL, USA), and 1.0 µm-pore polycarbonate membrane (Whatman, Maidstone, UK) by soaking each component in 70% isopropanol followed by a rinse with sterile DI water (
Figure 1A). The pre-extrusion volume contains oVV obtained from the manufacturers (“oVV”) in EDM at approximately a 1:100 mass ratio based on the whole protein content. This mix of EDM plus oVV was then processed by repeatedly passing back and forth through the membrane, propelled by syringe pumps. The process is illustrated in the schematics shown in
Figure 1B and
1C. Coated oVV (“EDM-coated oVV”) were deposited in sterile 1.5 mL tubes and used immediately. We use qPCR, as detailed in the protocol below, to quantify the number of copies of oVV DNA and EDM-coated oVV DNA, to ensure that no oVV is lost during the encapsulation process.
2.4. ELISA
Enzyme-linked immunosorbent assay (ELISA) detecting oVV was performed using the protocol that follows. We procured Rabbit anti-vaccinia HRP-conjugated polyclonal antibody as a detector (Abcam#87387), which was generated in rabbits against whole vaccinia virus. Serving both as a positive control and as a standard curve, freshly thawed oVV is first diluted to the same effective concentration as the extruded sample used for infecting cells; which is 3.00 × 106 pfu/mL. From this, we further diluted the sample to yield concentrations of: 1.50 × 106 pfu/mL, 3.00 × 105 pfu/mL, and 3.00 × 104 pfu/mL. A negative control comprising only EDM without any oVV was used and labelled as 0 pfu/mL. These samples are then used for coating an ELISA plate (n = 3). After incubation, we decanted all samples from the plate and washed 4 times with PBS. We blocked the plate with a solution of PBS plus 5% BSA (0.05 g/mL) and allowed the plate to incubate for 1 hour at room temperature. The block was decanted, and the plates were washed 4 times with PBS. We added polyclonal anti-Vaccinia HRP-conjugated antibody diluted to 4 µg/mL in blocking buffer to the plate and again incubated the plate for 1 hour at room temperature. After, the antibody was decanted. The plate was again washed 4 times with PBS. Finally, 100 µL of TMB solution (BioLegend, San Diego, CA, USA) was added to each well. We monitored each assay well for development and stopped the TMB reaction with 1.0 M hydrochloric acid. Absorbances of each well were read with a BioTek plate reader at 450 nm.
This ELISA is also used to determine the coating efficiency using the following steps that are performed alongside the concentration curve protocol. The extrusion product, which we conservatively assumed contains a mix of EDM-coated oVV and untouched oVV, is directly used to coat the bottom of other wells on the same ELISA plate, and then subjected to the same protocol. We hypothesize that of the mixture, only oVV will contain any reactivity against the antibody, and produce a positive HRP signal, while the EDM-coated oVV will not.
2.5. DLS/Zeta Potential
Dynamic light scattering (DLS) particle sizing and zeta potential measurements were both measured using a Malvern Panalytical Zetasizer Nano ZS in accordance with the manufacturer’s user manual. For particle size measurements, we gently pipette-mixed the sample of interest and aliquoted 300 µL of the sample into a disposable semi-micro-Vis cuvette (EP0030079353). We measured the particle size using the optimized particle size protocol from the Zetasizer program. To measure the zeta potential, we gently pipette-mixed the sample of interest, diluted the sample 1:10 in DI water, aliquoted 800 uL of the diluted sample into a folded capillary cuvette (DTS1070), and finally measured the zeta potential using the optimized particle size protocol from the Zetasizer program. This was repeated across samples at different pHs. We adjusted the pH of the sample using hydrochloric acid. We again repeated the measurements at different concentrations of EDM and allowed the sample to incubate at room temperature for different amounts of time after dilution to ensure that the samples reached equilibrium.
2.6. Calculating Relative Interparticle Forces at Different pHs
In order to determine the relative interparticle forces at different pHs, we first obtained a relative interparticle distance. Others previously devised a method to approximate the interparticle distance in colloidal mixtures utilizing particle concentration (n), which can be derived from the average relative particle volume (V
avg) [
30]. We determined the EDM’s interparticle distance using the following:
where d
z-avg is the average diameter of the particles. We obtained d
z-avg using the DLS measurements, and that diameter scales cubically with volume. Then, we obtained the number concentration of particles in the system (n) by dividing the mass concentration of particles (C) by the relative average particle volume (V
avg):
where n represents the number concentration. Using this, as others have done, we then very coarsely estimated the relative interparticle distance (a) as the cube root of the number concentration of particles (n),
We assume that the zeta-potential of the suspension (ζ) is proportional to the relative magnitude of charge of the particles’ surface (q
i), given by:
Together, we approximate the relative repulsive force using the scalar form of Coulombic repulsion, given by:
where F is the relative repulsive force between particles [
30]. As every instance of q
i originates from the same species of particle, for the purpose of estimation, we reduce the equation to
Using this framework for analysis, we determine the relative interparticle distance and its corresponding relative interparticle force for every measured dz-avg and ζ quantity obtained across its various pHs and equilibrium times.
2.7. pH-Mediated Infectivity Assay
For in vitro studies, A549, Vero E6, and MCF-7 cells were plated into 24-well plates at a density of 100,000 cells per well and incubated overnight for adhesion. The day following plating, we used DMEM (sans serum) “as is” for the neutral pH condition. For the acidified condition, to best mimic the pH conditions of a tumor microenvironment, we acidified DMEM with hydrochloric acid to a pH of ~5.2. Subsequently, we reconstituted the bare and coated viruses into their respective conditions, neutral or acidified, to an equivalent MOI of 0.5 and added to the cells for our infection phase, comprising 1 hour of incubation in a humidified incubator at 37 °C, 5% CO2. Following the infection phase, we aspirated the virus preps and washed cells with growth media 3 times to rinse residual virus preps and prevent further infection. For A549, Vero E6, and MCF-7 cells, two separate well plates were prepared to measure (1) the viability at 24 hours via a neutral red reuptake viability assay; and (2) total virus titer at 24 hours using a qPCR against viral DNA.
2.8. Neutral Red Reuptake Viability Assay
The neutral red reuptake assay quantitatively measures the number of viable cells. This assay was performed on infected A549, Vero E6, and MCF-7 cells 24 hours post-infection. First, we aspirated the virus-containing media from the wells and washed the cells once with PBS. Subsequently, we diluted 0.3% Neutral Red dye (Sigma-Aldrich, St. Louis, MO, USA) 100-fold in DMEM, added the diluted dye into the wells, and allowed it to incubate in a humidified incubator at 37 °C, 5% CO2 for one hour and a half, during which living cells will take in neutral red. After incubation, we removed the neutral red stain from the cells and then washed cells with PBS. To obtain a representative viability, we released the neutral red from live cells by first adding 50 µL of Sorenson’s citrate buffer and then adding 50 µL of 200 proof ethanol. The amount of neutral red dye present is directly correlated with the number of live cells in culture. We measured the amount of neutral red in each well using a BioTek plate reader by measuring the absorbance of the solution in the 540 nm wavelength.
2.9. qPCR
Quantitative polymerase chain reaction (qPCR) was used for the quantification of the number of oVVs or EDM-coated oVVs present. Here, in this study, it is used in several instances. (1) To measure the concentration of EDM-coated oVVs in the extrusion product to ensure that the encapsulation did not exclude any oVVs; (2) To measure the total number of copies of oVVs that would be present in A549, Vero E6, and MCF-7 cells 24 hours post infection with their respective experimental sample; (3) Concentration of oVVs or EDM-coated oVVs present in mouse blood, organs, or tumor in their respective in vivo experiments.
We extracted oVV DNA following the manufacturers protocol (Zymo Viral DNA kit). Briefly, we added a volume of vDNA extraction buffer equivalent to 4 times the plated volume to each well of a 24-well plate and incubated for 10 minutes. The samples were transferred to IC column tubes, centrifuged, and washed two times with DNA wash buffer. Then, we eluted the extracted oVV DNA with 20 µL of elution buffer. Once the oVV DNA was extracted from the column tubes, we added 2 uL of the oVV DNA to 18 µL of a SYBR green containing “one step” Mastermix (Applied Biosystems, Waltham, MA, USA) with 1 µmol forward and reverse primer against the Vaccinia E3L gene. The sequences used were: forward-primer 5′-TCCGTCGATGTCTACACAGG-3′ and reverse-primer 5′-ATGTATCCCGCGAAAAATCA-3′ [
31]. A standard curve of known amounts of oVV DNA correlated to the infectious units of the input DNA (pfu/mL) was included in each qPCR analysis. For the qPCR, we used an Applied Biosystems Stratagene MX3005P detecting SYBR green signal with a program of 1 cycle of 10 minutes at 95 °C, 40 cycles of 30 seconds at 95 °C, 1 minute at 55 °C, 1 minute at 72 °C.
2.10. In Vivo Study: Acute Pharmacokinetics and Biodistribution of oVV and EDM-Coated oVV in Wild-Type Mice upon Intravenous Administration
oVV and EDM-coated oVV inoculums were prepared as described using mouse EDM and injected intravenously into 24 healthy 6-week-old female C57/BL6 mice per experimental group. Six mice from each experimental group were then sacrificed in groups at the following timepoints: 15 minutes, 1 hour, 2 hours, and 4 hours, and the maximum amount of blood was withdrawn via aortic puncture. Whole blood was then used for qPCR in accordance with the protocol outlined in 2.8. All animal experiments were conducted at Anticancer Inc. (San Diego, CA, USA). The sacrificed mice were then dissected, and its various organs (liver, lung, kidney, spleen, and heart) extracted. The organs were then frozen at −80 °C until they were ready for tissue digestion, upon which viral DNA was then quantified.
2.11. Tissue Digestion for oVV DNA Extraction
To prepare the mouse tissue for DNA extraction and subsequently qPCR, we first cut up to 100 mg of organ tissue into small pieces, placed organ pieces into a 1.5 mL microfuge tubes, and recorded the weight of each sample. To begin the digestion, the sliced tissue is then submerged in 400 µL a proteinase-k buffer containing 190 µL DNase free water, 190 µL 2X proteinase K buffer, and 20 µL of proteinase K. We then incubated the tissue in a water bath set at 55 °C for 2 hours. Then, we centrifuged the 1.5 microfuge tubes to pellet the tissue debris and extracted the supernatant to collect vDNA for qPCR. We ran qPCR using the same primers, program, and procedure as previously described.
2.12. In Vivo Xenograft Study: Surgical Orthotopic Implantation
Tumor xenografts were generated orthotopically using surgical protocols as described elsewhere. In brief, 6 female NOG mice (5–6 weeks old) were injected with a single dose of 5 × 106 MDA-MB-231 human breast cancer cells that have been genetically engineered to stably express red fluorescent protein (RFP) into their second left mammary fat pad and allowed to grow. Once the tumor reached ~50 mm3, they were ready for dosing. Mice were sorted into 3 equal groups to receive EDM, oVV, or EDM-coated oVV; the concentration of viral inoculums was equivalent to 1 × 107 pfu in 100 uL PBS of oVV-GFP. Human EDM was used for the EDM-only control, and EDM-coated oVV. Tumors were monitored for size by 3 direction caliper measurements to compute total volume. Fluorescent imaging for green fluorescent protein (GFP) and RFP was captured, and each mouse was monitored for weight for a total of 2 weeks. All animal experiments were conducted at Anticancer Inc. (San Diego, CA, USA). The fluorescent imaging was digitally processed to remove autofluorescent signal from the skin and fur of the mice. A digital binary mask is then generated using signal in the red fluorescent channel gating for presence of cancer cells, and then used to look for co-localizations of GFP.
4. Discussion
Our work here builds on previous studies showing that the encapsulation of oncolytic viruses with cell membranes improves the infectivity of oncolytic viruses both in vitro and in vivo [
26,
27]. Past studies have shown that cancer cell-derived membranes can be used to coat oncolytic adenoviruses, creating a non-viral enveloped adenovirus, that exhibits improved cancer cell killing capabilities that look to surpass that of even viral protein-mediated infection through viral envelopes. This has led us to believe that artificial, non-viral envelopes may possess more than one avenue of infection, and that these routes can work synergistically to facilitate viral entry to cells more quickly compared to native viral envelopes. Here, we highlight the possibility that one such avenue could be electrochemical, mediated through pH stimuli, which allows the virus to behave like a “smart” nanoparticle.
To generate these non-viral enveloped OVs, we used erythrocyte membrane as the non-virus envelope. Erythrocyte membrane derivation has been fine-tuned over decades since it was discovered and is well characterized. Erythrocyte membranes also bear many hallmarks of its original cell, including the expression of cell surface markers that allow it to remain in systemic circulation for longer periods of time. This could be important for the systemic administration of OVs.
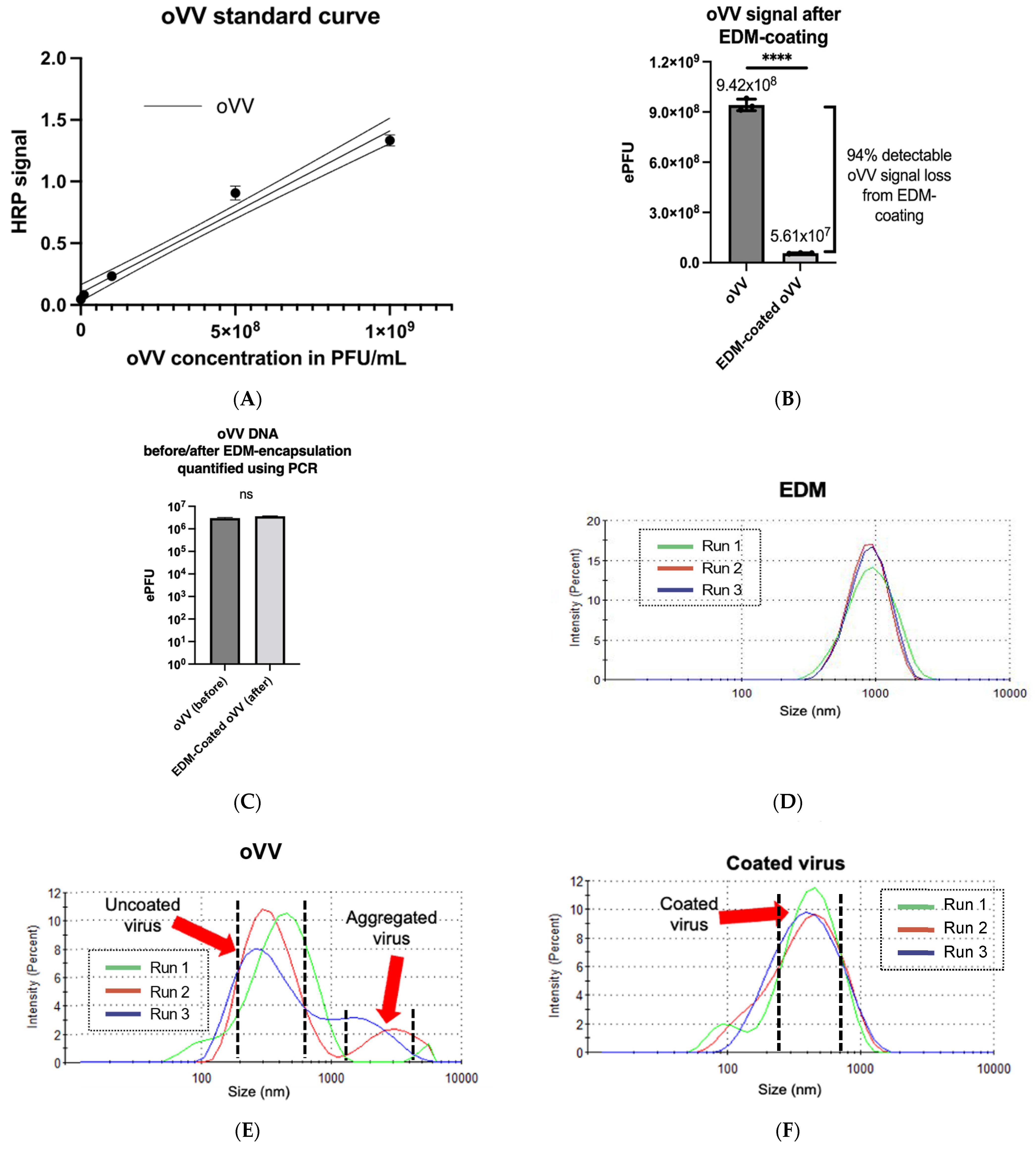
Our studies show that, after encapsulation, the EDM undergoes consistent reduction in size from 1000 nm to a very uniform distribution of a new EDM-coated oVV with an even distribution peaking at a size of 450 nm, with a low PDI of 0.274 (
Figure 2F). It is also worth noting that a particle size measurement of the oVV produced multi-modal distribution with larger particles peaking between 1000 nm and 2000 nm, suggesting that our starting oVV material, while purified of cellular debris in the sucrose gradient, is prone to aggregation (
Figure 2E). Coating oVV with EDM produced a much more homogenous product that is stable and resists aggregation.
It is important to note here that at the last step of oVV production, lysis of the cells used to grow the oVVs, releases both IMVs and EEVs and the all the oVVs are collected after purification. Studies have shown that these IMVs often comprise the majority of the entire viral population, with EEVs constituting the remainder [
32]. The composition of the product is often not characterized, as separating the IMVs from EEVs is difficult to accomplish on a commercial scale and may not be economically feasible given the larger proportion of IMVs in the product. Here, we conservatively choose not to make any assumptions about the envelope state of the oVV, and as such, for our ELISA, we chose a polyclonal antibody which would in theory bind to both the IMV and EEV in our oVV. We also clearly see that the oVV signal diminishes significantly after coating, which shows that extrusion with EDM has led to lower detectable proportions of both IMV and EEV in the product. Hence, we conclude that we were able to perform the coating of our oVV consistently with >94% efficiency (
Figure 2B).
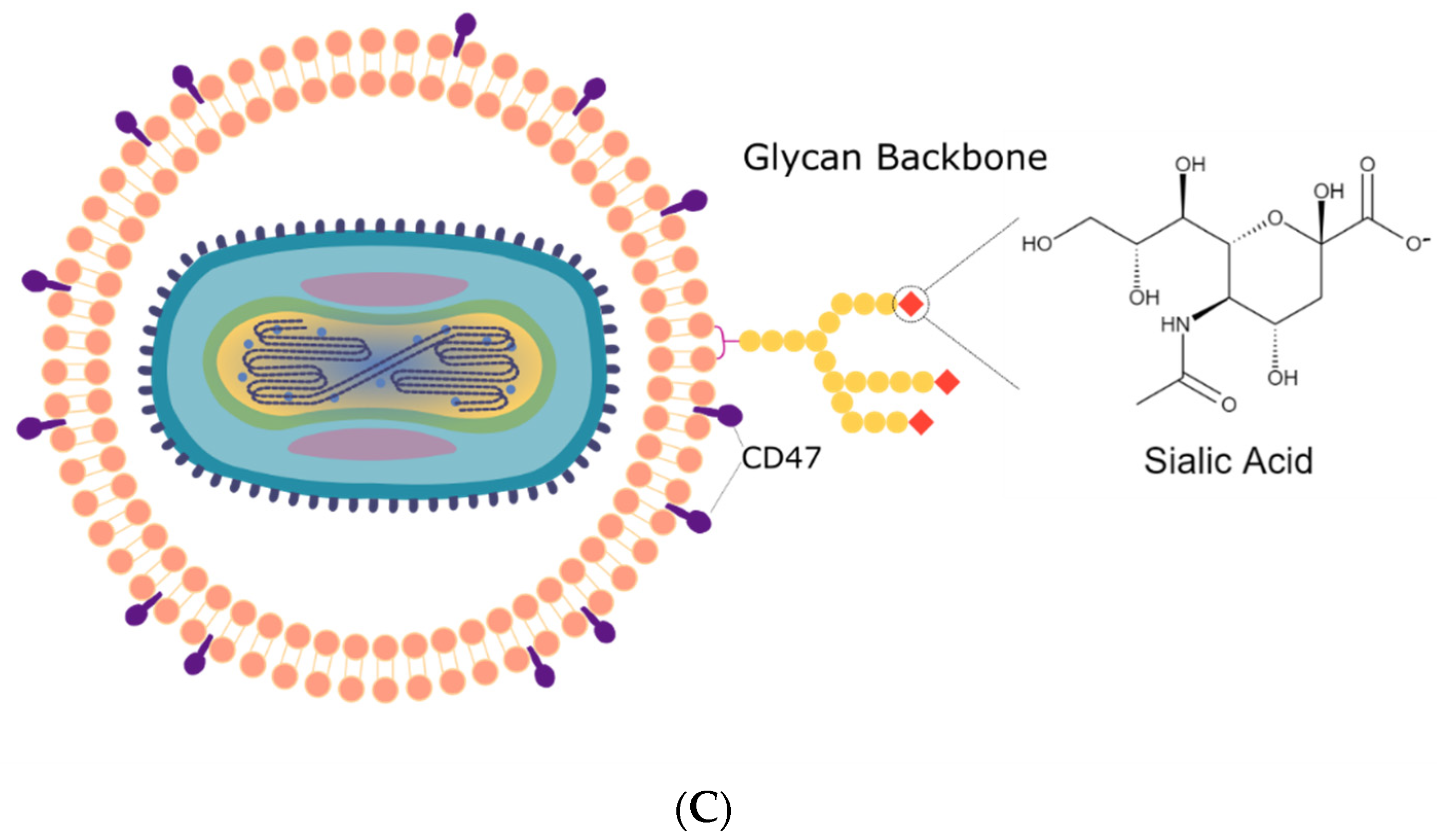
By coating the oVV with EDM, we recapitulate the functionality granted to the virus by the envelope. The envelope is a key component for many enveloped viruses as viral surface proteins directs infection. While EDM-coating does not bear any viral proteins, it can disguise the virus as an erythrocyte, which is nonimmunogenic and, through the expression of CD47 expressed on its surface (otherwise known as the “don’t eat me” signal), prevents the virus from phagocytosis [
24,
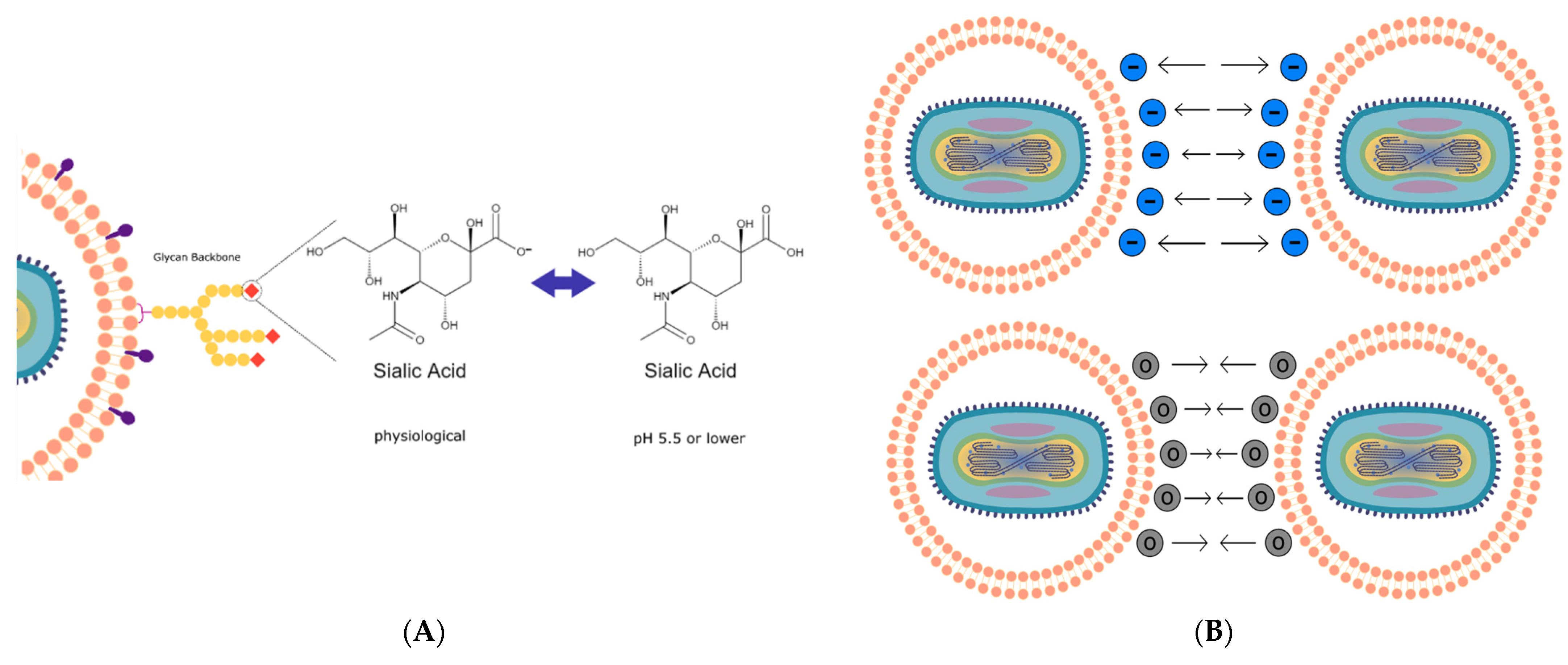
25]. We further discover that the sialic acid groups that decorate the EDM surface can lead to the particle becoming a pH-sensitive vehicle that can respond to changes in environmental pH (
Figure 3A,
3B,
4A and
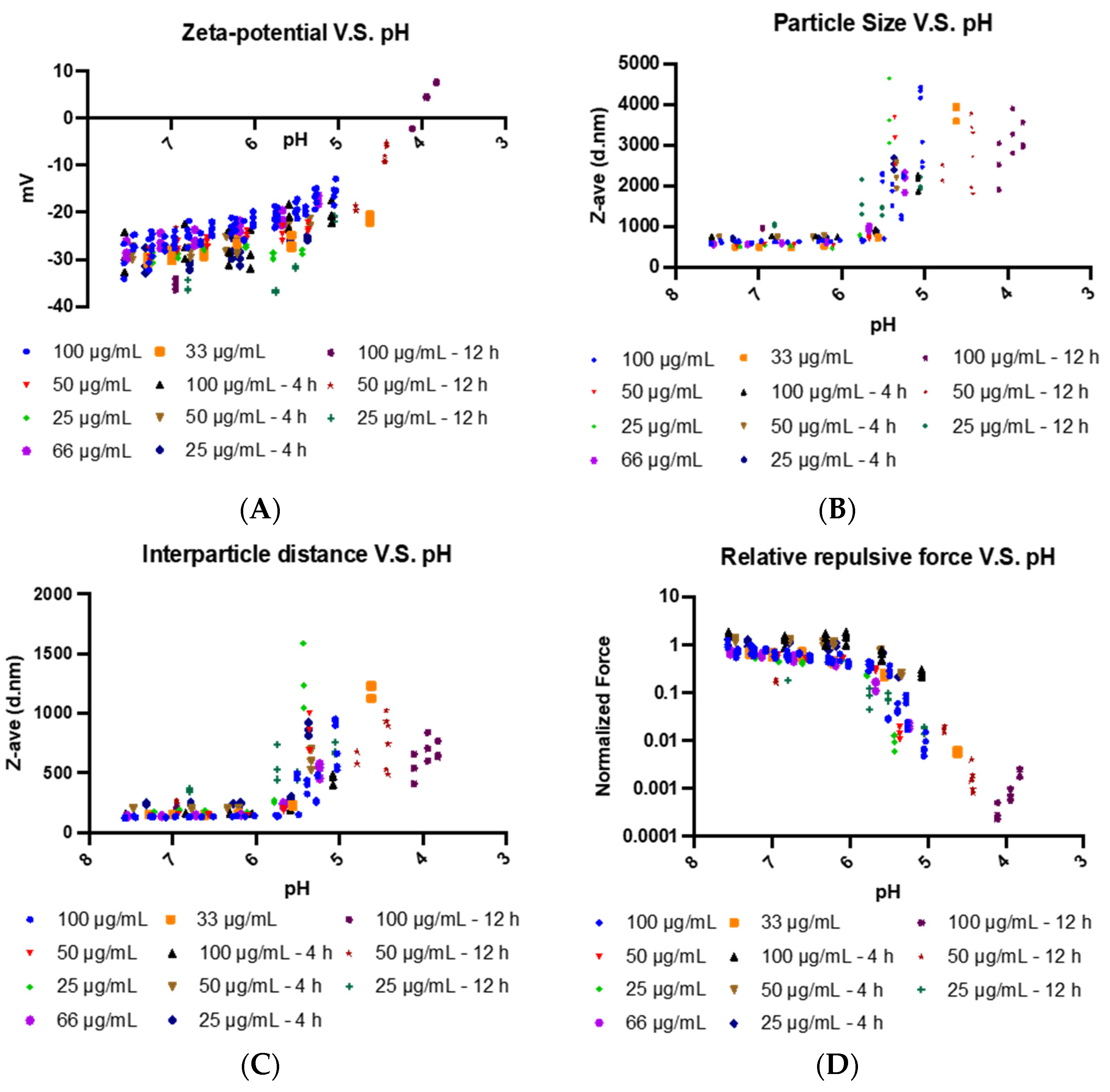
4B). To understand this pH sensitivity and its influence on the delivery of the EDM-coated oVV into the cancer cell, we built a mathematical model correlating differences in physical EDM behavior to its environmental pH. As the environmental pH decreased, these erythrocyte-derived membranes underwent neutralization in surface charge (
Figure 4A), suggesting that the sialic residues that decorate the EDM surface are being protonated in the increasingly acidic environment as illustrated in
Figure 3A and
3B. The correlating particle size measurements showed no change as the acidity increased, up to a pH of 6, below which the particles sizes started increasing with decreasing pH (
Figure 4B). Based on DLVO theory, we deduce that the repulsive force generated by the remaining negative charges has weakened to a state at which the particles can no longer repel each other [
33]. Conversely, the inherent Van der Waals forces between the particles become dominant at this pH, allowing the particles to become more physically drawn to each other. This leads to the aggregation and eventual precipitation of the membrane. This phenomenon is also observed in liposomal literature, where dispersion stability is also breached between ±20 mV, below which liposomes also tend to crash out of solution [
34,
35,
36]. To characterize these interactions more quantitatively, we developed a model that considers the surface charge and interparticle distance to estimate and study the trends in repulsive force across the pH of interest. As the interparticle distance starts increasing due to aggregation at a pH of 6, we see that repulsive force between particles start to drop much faster due to the inverse relationship between force and distance in the Coulombic equation (
Figure 4C and
4D). In healthy, cellular states, this proposed phenomenon cannot be realized to the dominant repulsive electrostatic forces that push cell membranes apart. However, this is not the same in the acidic tumor microenvironment. We believe that this aggregation of particles leads to an alternative mode of infection via non-endogenous membrane fusion, resembling that of the liposomal delivery of coated payloads. This allows the coated-oVV to be released into the cell.
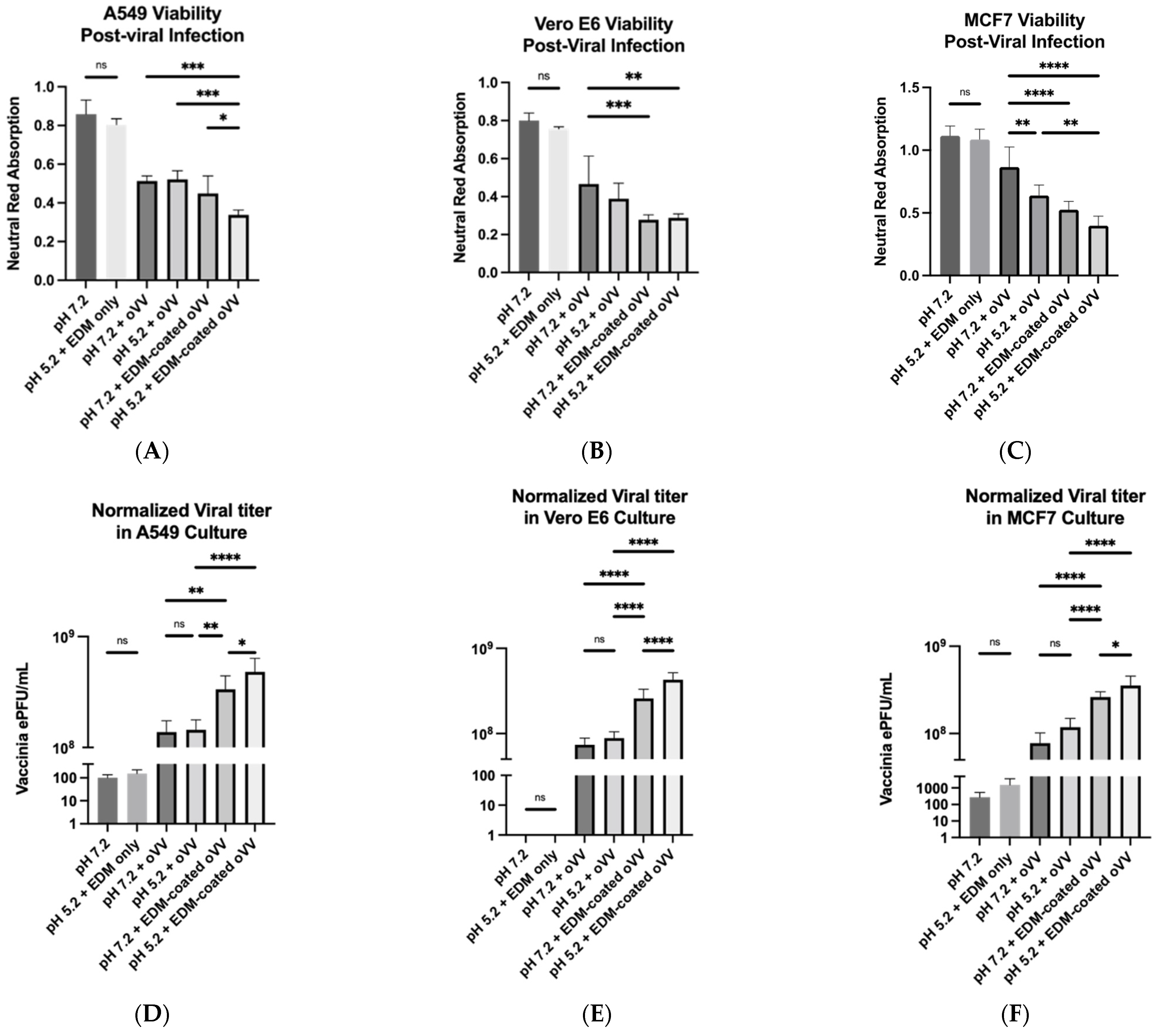
To test our hypothesis in a controlled environment, we treated several cell lines in vitro following the previously defined experimental groups of: (1) neutral pH + bare virus; (2) acidic pH + bare virus; (3) neutral pH + coated virus; and (4) acidic pH + coated virus. We also prepared negative controls with no treatment and environmental/vehicle controls where the cells are subject to pH perturbations in the presence of EDM but without any viruses. No significant differences were observed between the negative control and the environmental/vehicle control, which shows that neither the pH nor the presence of EDM were cytotoxic to the cells. Minimal differences were observed between experimental groups 1 and 2, with group 2 presenting as slightly more cytotoxic (
Figure 5A,
5B, and
5C). This suggests that the virus is not inherently pH sensitive. However, it is worth noting that past studies have shown that A25 and A26, which are mature vaccinia virion proteins that mediate its uptake into a host, are pH sensitive and perform better in acidic environments [
37,
38]. Since our results show that oVV performs equally in both groups regardless of acidity, this is suggestive that a bigger proportion of the oVV in our oVV samples consists of IMVs.
While our results also yielded no significant differences in viability between the coated virus groups, we detected significant differences in the intracellular viral concentration whether the infection was achieved under acidic or neutral conditions (
Figure 5A–F). To quantify intracellular viral concentration, we not only extrapolated the amount of PFUs of oVV within the cells in culture, we also normalized it to the neutral red signal, which we use here to determine the number of living cells in the culture. We felt that this makes the results more meaningful in this context due to the rapid killing capabilities of oVVs in in vitro culture; the more effective the virus became (as a result of the coating), the fewer the number of cells remained in culture, which then prevented the replication of more viruses. In our experimental observations, the viability in both coated virus-treated samples were extremely poor regardless of whether it belonged to the acidic group or the neutral group; most of the well had copious amounts of cell death in the center of the well, leaving only a ring of live cells around the edges. This abundance in cell killing led to our difficulty in statistically distinguishing between the coated samples. However, despite the equivalence in cell death, our intracellular viral concentration assay yielded a significant difference, wherein the cells that were exposed to the coated virus under the acidic condition presented higher viral titers (
Figure 5D,
5E, and
5F). For this, we hypothesize that either or both of the following must occur under acidic conditions: (1) increased viral entry per cell; or (2) increased infectivity rate of the viruses into the cells such that viral replication occurs at an earlier time. Since an increase in infectivity of the oVV was not significant across the different pH conditions, we can conclude that the pH sensitivity is a manifestation of the coating and not an inherent property of the virus.
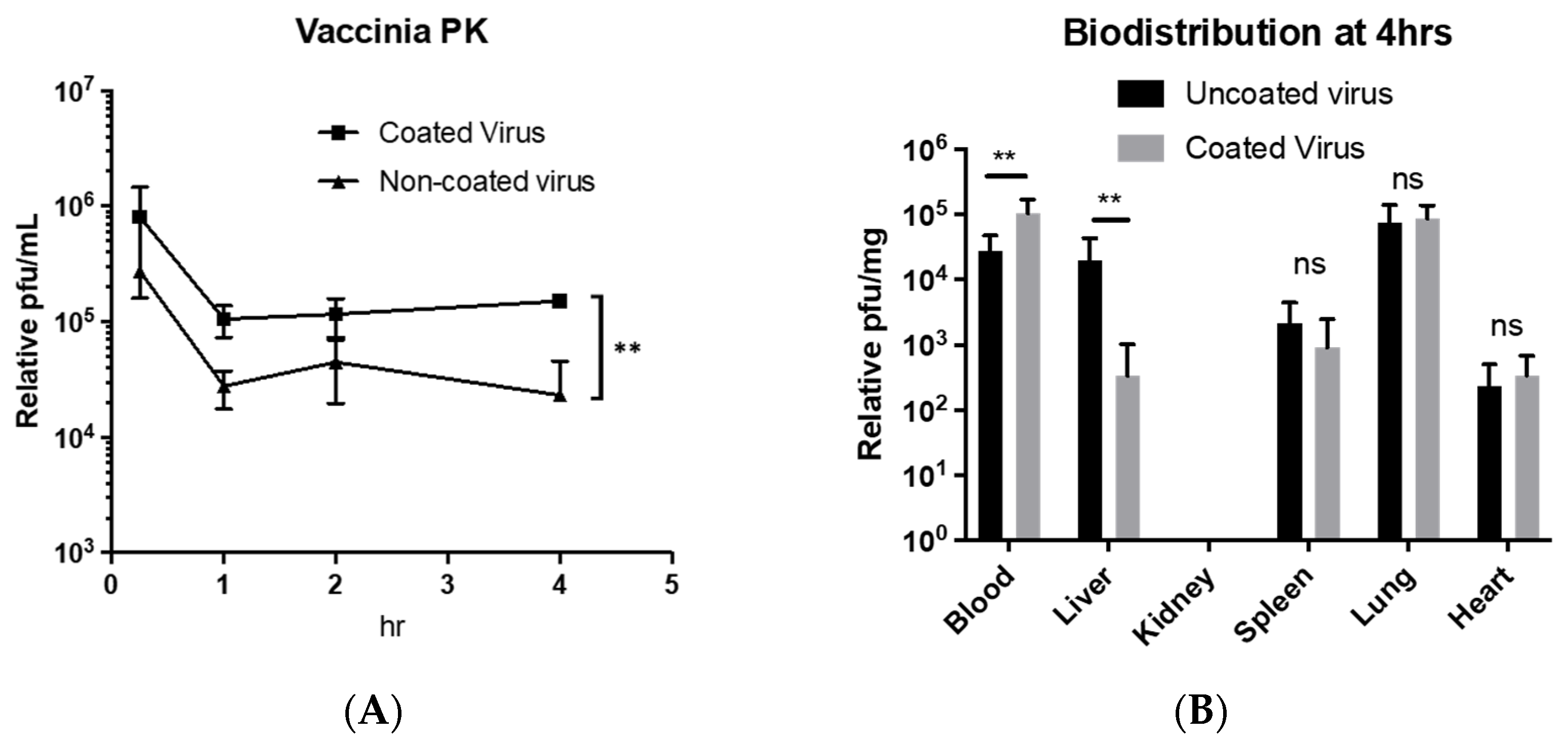
In the other arm of our study, we continue to explore the retention of circulating viruses that can be conveyed onto the oVV through encapsulation. Manufactured oVV can be cleared or deactivated very quickly by many different mechanisms, such as sequestration into the liver or neutralization by complement proteins [
39]. Our experiment comparing the circulation of EDM-coated oVV against oVV shows greater short-term persistence when the coating is applied in immune-competent wild-type mice. We chose wild-type mice here as we wanted to study circulatory retention in an immune competent model while remaining outside of the influence of sequestration within tumors. As such, we specially derived EDM from mice as a material for coating the oVVs to maintain the same physiologic relevance.
Given that the pharmacokinetics of the virus was altered by the coating (
Figure 6A), it is then just as important here to then understand that the biodistribution of the EDM-coated oVV could also differ. First, we hypothesized that, due to the erythrocyte coating, we will see an increased accumulation of the virus in the liver and spleen after coating (
Figure 6B). We base this hypothesis on the natural degradation sites of erythrocytes, where they can either be broken down in the spleen or the liver. However, our results here also show that apart from a lowered liver accumulation, the virus does not tend to sequester in any other organ that it would not have without the coating. Hence, not only do we see that the coating does not alter the biodistribution in any concerning way, but the converse effect of liver evasion also occurred. We also noted that even though the spleen is a major site of erythrocyte sequestration, this accumulation occurs primarily due to the aging of erythrocytes that leads to immunogenic conformational change in the band-3 protein on the surface of erythrocytes, which is tied to its immune-related clearance. Since there is no increased sequestration of EDM-coated viruses in the spleen at the time of sampling, we think that our process of EDM derivation and application of the coating onto the oVV does not lead to the immunogenic conformational change of band 3 protein within 4 hours, which then allows the EDM-coated oVV to side-step accumulation in the spleen. Additionally, the loss of viral signal in the liver when we compare the oVV to the EDM-coated oVV suggests that the coated viruses gained the ability to resist the physical filtration within the liver where the uncoated virus would not.
To measure the translation of this increase in infectivity into a physiologically relevant in vivo model, we chose (1) the MDA-MB-231 breast cancer xenograft model; and (2) to administer the virus intratumorally. Breast cancer was chosen as studies have shown that it can develop acidic tumors [
40], and the NOG mice was used as it is the most immune-compromised mouse model that is currently being studied. These NOG mice have not only lost their adaptive immunity but also have reduced leukocytes and impaired phagocytosis mechanisms. This model was specifically used for two reasons: (1) to reduce the amount of immune clearance of the oVV; and (2) to allow us to deliver the EDM-coated oVV using human extracted membranes. Studies with blood transfusions into various mouse models have shown that the NOG mice would clear our human erythrocytes very slowly, which is most conducive for our experiment. In order to eliminate the differences between the tropism of oVV and EDM-coated oVV that may arise because of the differences in blood virus concentration, and homing to the tumor, we opted to directly inject the viruses into the tumor. Our PCR data from
Figure 2C show that both samples contain approximately the same number of viruses by genome copy, so we can be sure that tumors across groups receive the same viral dose. This allows us to remove other influences and study, in isolation, the differences in infectivity of oVV and EDM-coated oVV.
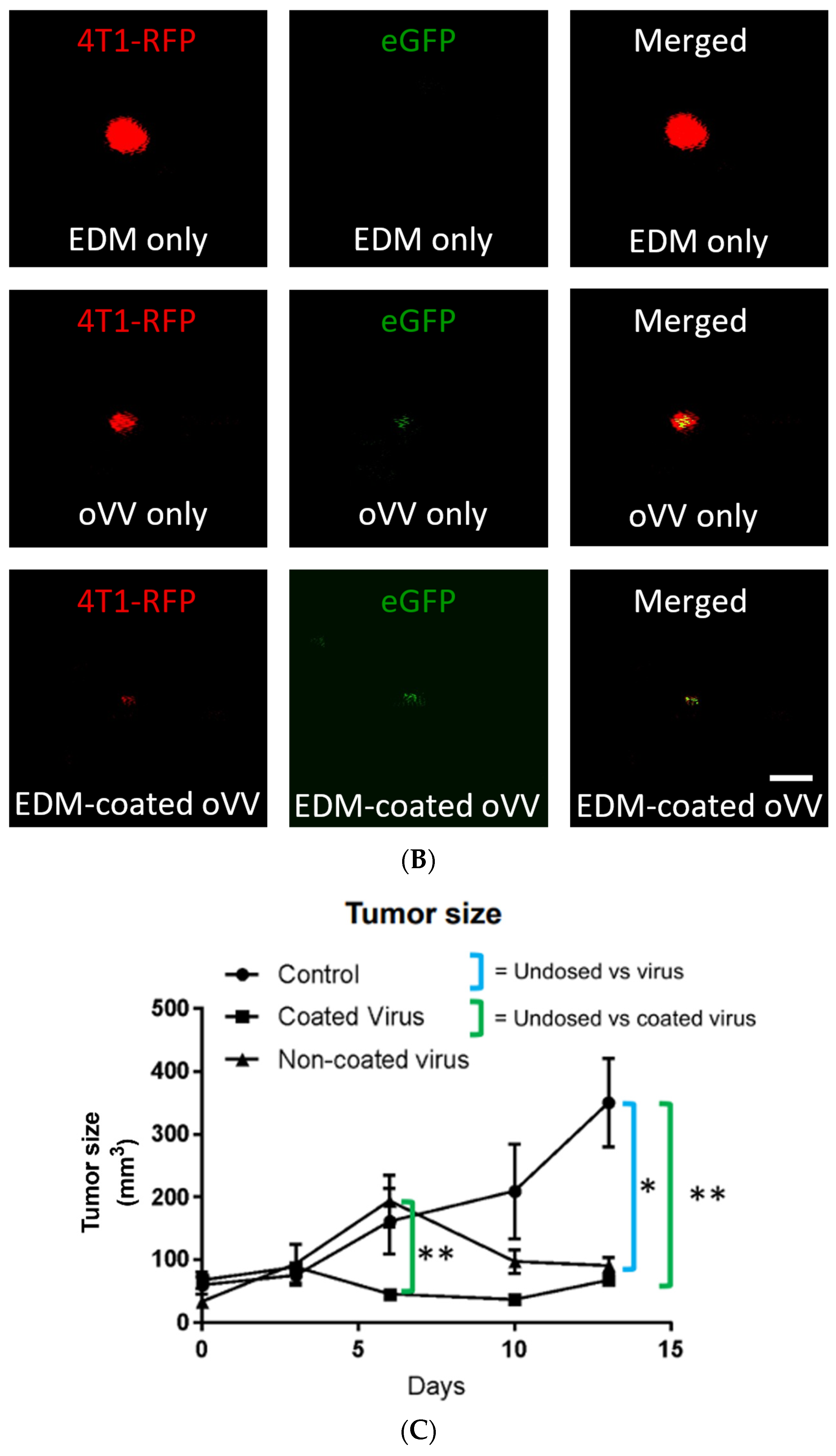
Our fluorescent imaging showed a decrease in RFP signal as we compare between the mice that only received EDM, the mice that received the oVV, and mice that received the EDM-coated oVV. This proves that the EDM-coated virus can trigger cancer cell death faster than the oVV itself. Both groups did not show much GFP, which is a direct result of the rapid cancer cell death upon oVV infection and its subsequent apoptosis. This would cause the transcribed GFP to leak out of the tumor, and it would then cease to be detected (
Figure 7B).
The tumors that received EDM-coated oVV demonstrated a much faster tumor shrinking response when compared to the tumors that received the oVV (
Figure 7C). This echoes our earlier in vitro conclusion that one or both of two things must have occurred: either the coated viruses were able to enter the cancer cells much faster such that the viral replication process was initiated much earlier, or more copies of the coated virus entered the cancer cells which led to a response of higher magnitude. As we eliminated other confounding in vivo factors that could have led to this difference in tumor response, discussions made from our in vitro experiment are more relevant and similar parallels can be drawn, which is that the biggest contributing factor to increased infectivity is the presence of the EDM coating itself, with further improvements due to acidity.
It is also important here to note that, at the end of the study, both the coated and uncoated viruses managed to achieve the same efficacy, noting that the virus is efficacious, albeit slower to act. Ultimately, our goal here was to demonstrate two things: (1) that biological membrane coatings that are not viral envelope can still assist oVV therapy overcome certain barriers, in this case achieving prolonged circulatory retention without sacrificing its infectivity, and (2) that these coating may present stimuli responsive properties that scientists and clinicians can take advantage of when considering the development of a therapeutic. We also would like to add that, since we utilized a xenograft mouse model, there would be a lack of clearance and neutralization of the native oVV. However, had this study been performed in an immune competent mouse, the differences could have been much bigger because the slower acting therapeutic would be more liable to immune clearance, even when injected intratumorally.



 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}








