

Molecular Insights to the Structure-Interaction Relationships of Human Proton-Coupled Oligopeptide Transporters (PepTs)
 ,
, 
 ,
, 
Abstract
:1. Introduction
1.1. Substrate Specificities of PepT1 and PepT2
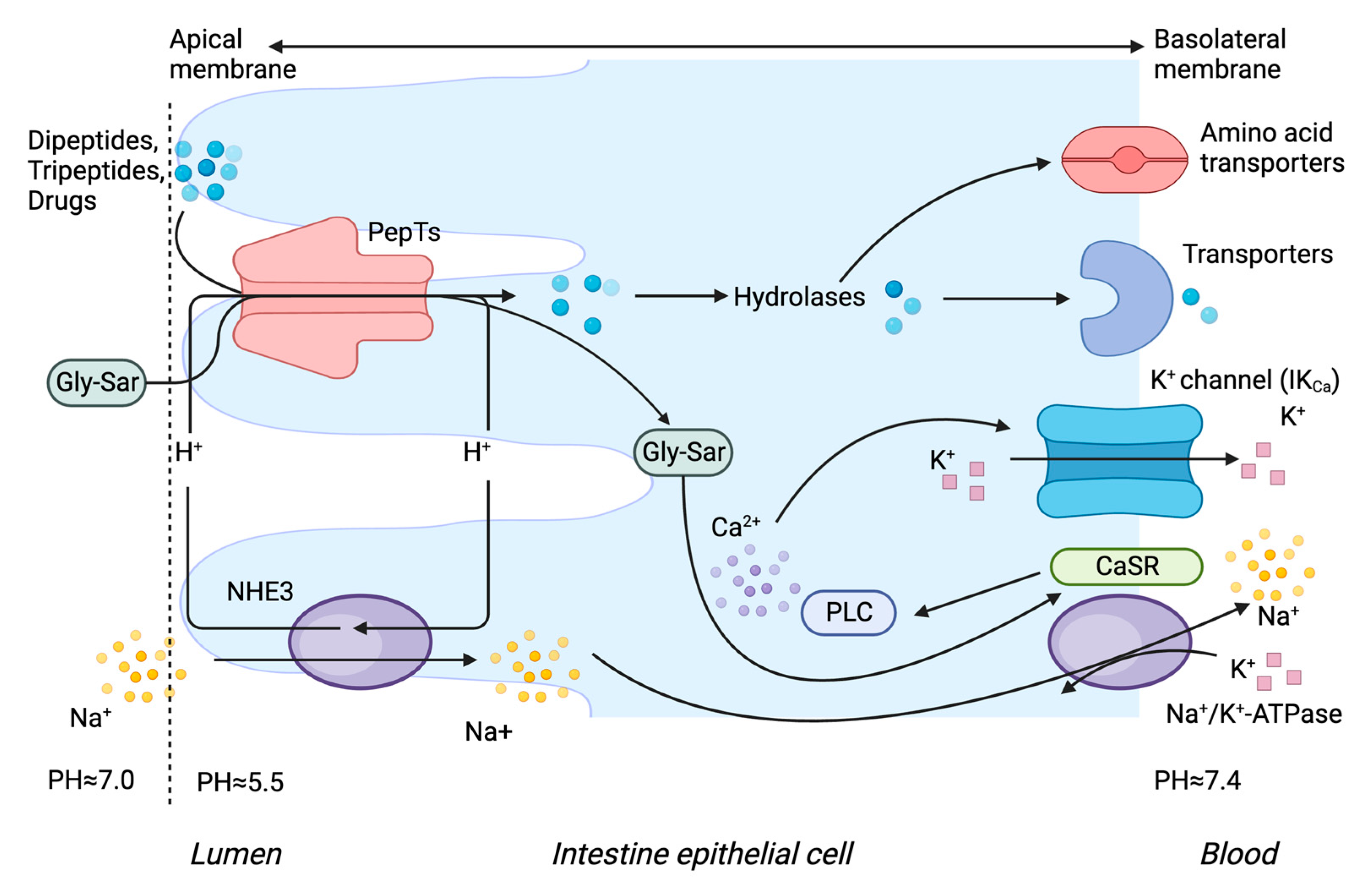
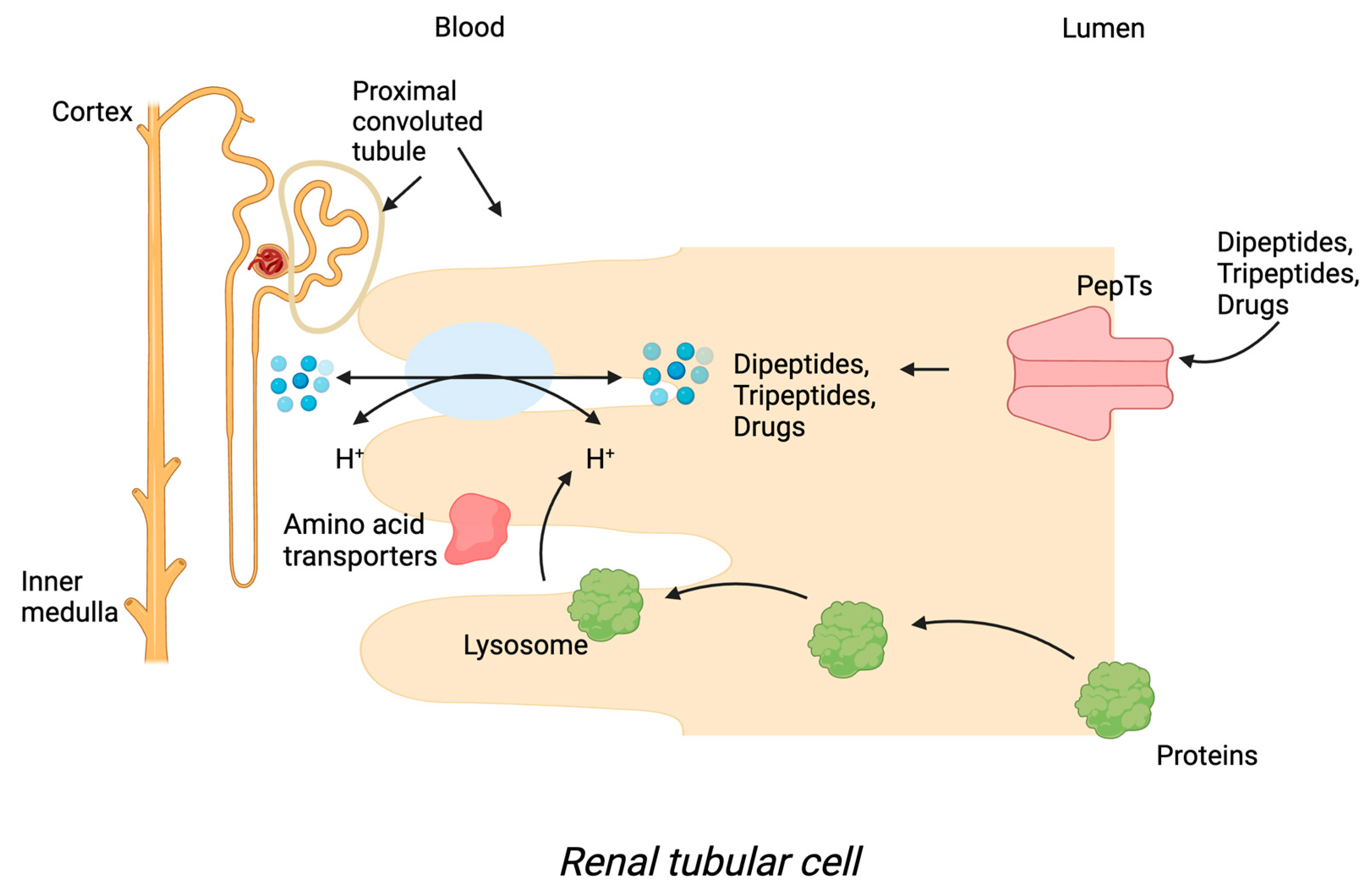
1.2. Tissue Localisation of PepT1 and PepT2
1.3. Transport Mechanism of PepT1 and PepT2
1.4. Molecular Regulation of PepT1 and PepT2
1.5. Regulation of PepT1 and PepT2 in Disease
1.6. Genetic Polymorphisms of PepT1 and PepT2
2. Structure–Function Relationships of PepT1 and PepT2
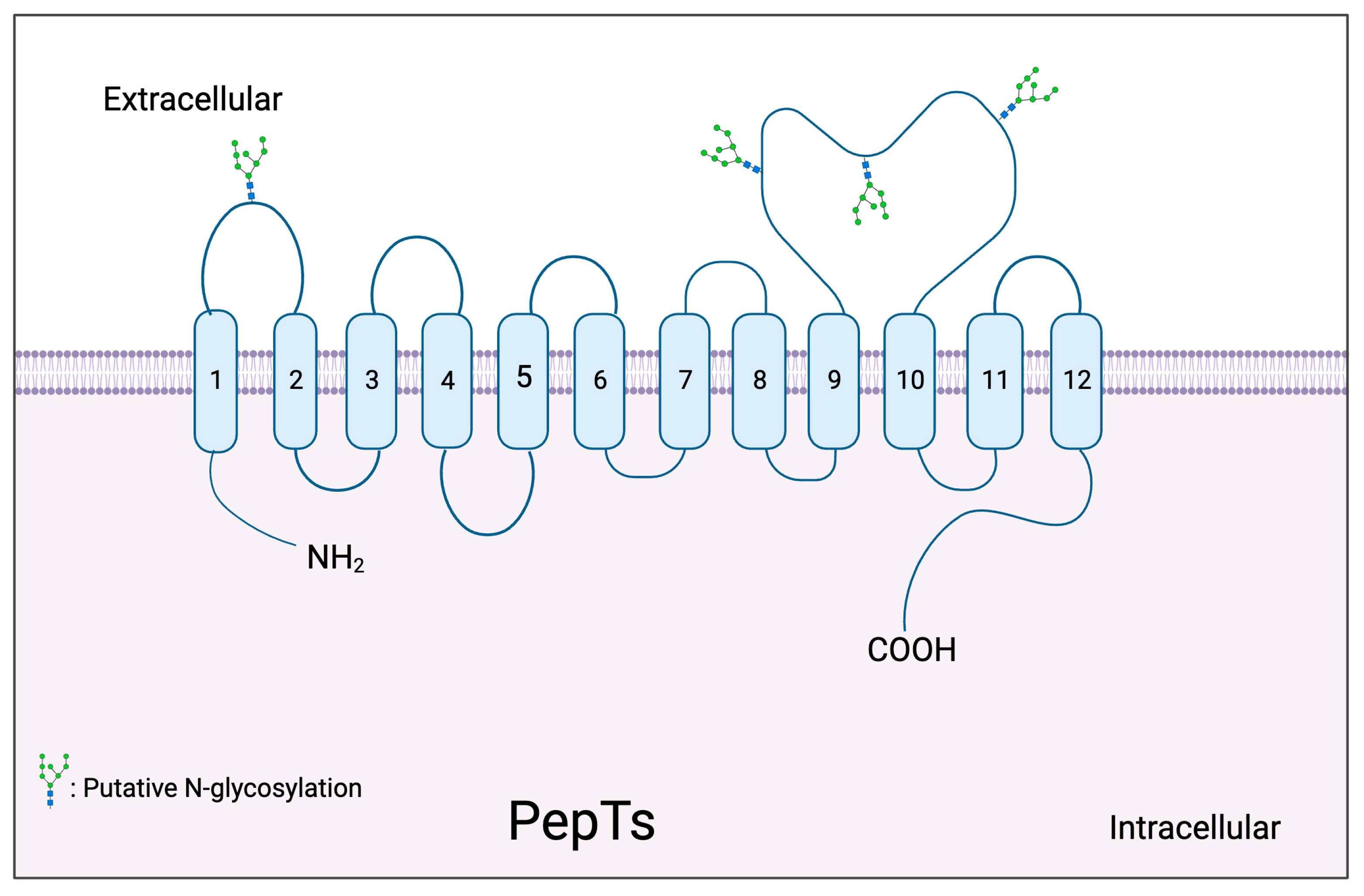
2.1. Topology Model of PepTs
2.2. D Structure of PepT1 and PepT2
3. Structure-Based Drug Design and Optimisation in Relation to PepTs
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Apostolopoulos, V.; Bojarska, J.; Chai, T.T.; Elnagdy, S.; Kaczmarek, K.; Matsoukas, J.; New, R.; Parang, K.; Lopez, O.P.; Parhiz, H.; et al. A Global Review on Short Peptides: Frontiers and Perspectives. Molecules 2021, 26, 430. [Google Scholar] [CrossRef] [PubMed]
- Daniel, H.; Rubio-Aliaga, I. An update on renal peptide transporters. Am. J. Physiol. Renal Physiol. 2003, 284, F885–F892. [Google Scholar] [CrossRef] [PubMed]
- Ganapathy, V.; Leibach, F.H. Peptide transport in intestinal and renal brush border membrane vesicles. Life Sci. 1982, 30, 2137–2146. [Google Scholar] [CrossRef]
- Ganapathy; Leibach, F. H. Is intestinal peptide transport energized by a proton gradient? Am. J. Physiol. 1985, 249, G153–G160. [Google Scholar] [CrossRef]
- Ganapathy, V.; Leibach, F.H. Carrier-mediated reabsorption of small peptides in renal proximal tubule. Am. J. Physiol. 1986, 251, F945–F953. [Google Scholar] [CrossRef] [PubMed]
- Fei, Y.J.; Kanai, Y.; Nussberger, S.; Ganapathy, V.; Leibach, F.H.; Romero, M.F.; Singh, S.K.; Boron, W.F.; Hediger, M.A. Expression cloning of a mammalian proton-coupled oligopeptide transporter. Nature 1994, 368, 563–566. [Google Scholar] [CrossRef]
- Martinez Molledo, M.; Quistgaard, E.M.; Flayhan, A.; Pieprzyk, J.; Low, C. Multispecific Substrate Recognition in a Proton-Dependent Oligopeptide Transporter. Structure 2018, 26, 467–476.e464. [Google Scholar] [CrossRef]
- Khavinson, V.; Linkova, N.; Kozhevnikova, E.; Dyatlova, A.; Petukhov, M. Transport of Biologically Active Ultrashort Peptides Using POT and LAT Carriers. Int. J. Mol. Sci. 2022, 23, 7733. [Google Scholar] [CrossRef]
- Kramer, W.; Girbig, F.; Gutjahr, U.; Kleemann, H.W.; Leipe, I.; Urbach, H.; Wagner, A. Interaction of renin inhibitors with the intestinal uptake system for oligopeptides and beta-lactam antibiotics. Biochim. Biophys. Acta 1990, 1027, 25–30. [Google Scholar] [CrossRef]
- Friedman, D.I.; Amidon, G.L. Passive and carrier-mediated intestinal absorption components of two angiotensin converting enzyme (ACE) inhibitor prodrugs in rats: Enalapril and fosinopril. Pharm. Res. 1989, 6, 1043–1047. [Google Scholar] [CrossRef]
- Friedman, D.I.; Amidon, G.L. Intestinal absorption mechanism of dipeptide angiotensin converting enzyme inhibitors of the lysyl-proline type: Lisinopril and SQ 29,852. J. Pharm. Sci. 1989, 78, 995–998. [Google Scholar] [CrossRef] [PubMed]
- Bretschneider, B.; Brandsch, M.; Neubert, R. Intestinal transport of beta-lactam antibiotics: Analysis of the affinity at the H+/peptide symporter (PEPT1), the uptake into Caco-2 cell monolayers and the transepithelial flux. Pharm. Res. 1999, 16, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Walter, E.; Kissel, T.; Reers, M.; Dickneite, G.; Hoffmann, D.; Stuber, W. Transepithelial transport properties of peptidomimetic thrombin inhibitors in monolayers of a human intestinal cell line (Caco-2) and their correlation to in vivo data. Pharm. Res. 1995, 12, 360–365. [Google Scholar] [CrossRef]
- de Vrueh, R.L.; Smith, P.L.; Lee, C.P. Transport of L-valine-acyclovir via the oligopeptide transporter in the human intestinal cell line, Caco-2. J. Pharmacol. Exp. Ther. 1998, 286, 1166–1170. [Google Scholar]
- Sugawara, M.; Huang, W.; Fei, Y.J.; Leibach, F.H.; Ganapathy, V.; Ganapathy, M.E. Transport of valganciclovir, a ganciclovir prodrug, via peptide transporters PEPT1 and PEPT2. J. Pharm. Sci. 2000, 89, 781–789. [Google Scholar] [CrossRef]
- Doring, F.; Walter, J.; Will, J.; Focking, M.; Boll, M.; Amasheh, S.; Clauss, W.; Daniel, H. Delta-aminolevulinic acid transport by intestinal and renal peptide transporters and its physiological and clinical implications. J. Clin. Investig. 1998, 101, 2761–2767. [Google Scholar] [CrossRef] [PubMed]
- Balimane, P.V.; Tamai, I.; Guo, A.; Nakanishi, T.; Kitada, H.; Leibach, F.H.; Tsuji, A.; Sinko, P.J. Direct evidence for peptide transporter (PepT1)-mediated uptake of a nonpeptide prodrug, valacyclovir. Biochem. Biophys. Res. Commun. 1998, 250, 246–251. [Google Scholar] [CrossRef]
- Balvinder, S.; Vig, T.R.S.; Julita, K.; Timoszyk, J.K.; Quan, Y.; Wall, D.A.; Smith, R.L.; Faria, T.N. Human PEPT1 Pharmacophore Distinguishes between Dipeptide Transport and Binding. J. Med. Chem. 2005, 49, 3636–3644. [Google Scholar] [CrossRef]
- Wenzel, U.; Thwaites, D.T.; Daniel, H. Stereoselective uptake of beta-lactam antibiotics by the intestinal peptide transporter. Br. J. Pharmacol. 1995, 116, 3021–3027. [Google Scholar] [CrossRef]
- Brandsch, M.; Thunecke, F.; Kullertz, G.; Schutkowski, M.; Fischer, G.; Neubert, K. Evidence for the absolute conformational specificity of the intestinal H+/peptide symporter, PEPT1. J. Biol. Chem. 1998, 273, 3861–3864. [Google Scholar] [CrossRef]
- Meredith, D.; Temple, C.S.; Guha, N.; Sword, C.J.; Boyd, C.A.; Collier, I.D.; Morgan, K.M.; Bailey, P.D. Modified amino acids and peptides as substrates for the intestinal peptide transporter PepT1. Eur. J. Biochem. 2000, 267, 3723–3728. [Google Scholar] [CrossRef] [PubMed]
- Abe, H.; Satoh, M.; Miyauchi, S.; Shuto, S.; Matsuda, A.; Kamo, N. Conjugation of dipeptide to fluorescent dyes enhances its affinity for a dipeptide transporter (PEPT1) in human intestinal Caco-2 cells. Bioconjug. Chem. 1999, 10, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Knütter, I.; Theis, S.; Hartrodt, B.; Born, I.; Brandsch, M.; Daniel, H.; Neubert, K. A novel inhibitor of the mammalian peptide transporter PEPT1. Biochemistry 2001, 40, 4454–4458. [Google Scholar] [CrossRef] [PubMed]
- Niida, A.; Tomita, K.; Mizumoto, M.; Tanigaki, H.; Terada, T.; Oishi, S.; Otaka, A.; Inui, K.; Fujii, N. Unequivocal synthesis of (Z)-alkene and (E)-fluoroalkene dipeptide isosteres to probe structural requirements of the peptide transporter PEPT1. Org. Lett. 2006, 8, 613–616. [Google Scholar] [CrossRef] [PubMed]
- Brandsch, M. Transport of L-proline, L-proline-containing peptides and related drugs at mammalian epithelial cell membranes. Amino Acids 2006, 31, 119–136. [Google Scholar] [CrossRef] [PubMed]
- Bajraktari-Sylejmani, G.; von Linde, T.; Burhenne, J.; Haefeli, W.E.; Sauter, M.; Weiss, J. Evaluation of PepT1 (SLC15A1) Substrate Characteristics of Therapeutic Cyclic Peptides. Pharmaceutics 2022, 14, 1610. [Google Scholar] [CrossRef]
- Scherrmann, J.-M. Comprehensive Medicinal Chemistry II. In Chemistry, Molecular Sciences and Chemical Engineering; Elsevier: Amsterdam, The Netherlands, 2007; p. 5. [Google Scholar]
- Wang, M.; Zhang, X.; Zhao, H.; Wang, Q.; Pan, Y. Comparative analysis of vertebrate PEPT1 and PEPT2 genes. Genetica 2010, 138, 587–599. [Google Scholar] [CrossRef]
- Inui, K.; Terada, T. Dipeptide transporters. Pharm. Biotechnol. 1999, 12, 269–288. [Google Scholar] [CrossRef]
- Daniel, H.; Adibi, S.A. Transport of beta-lactam antibiotics in kidney brush border membrane. Determinants of their affinity for the oligopeptide/H+ symporter. J. Clin. Investig. 1993, 92, 2215–2223. [Google Scholar] [CrossRef]
- Akarawut, W.; Lin, C.J.; Smith, D.E. Noncompetitive inhibition of glycylsarcosine transport by quinapril in rabbit renal brush border membrane vesicles: Effect on high-affinity peptide transporter. J. Pharmacol. Exp. Ther. 1998, 287, 684–690. [Google Scholar]
- Terada, T.; Saito, H.; Inui, K. Interaction of beta-lactam antibiotics with histidine residue of rat H+/peptide cotransporters, PEPT1 and PEPT2. J. Biol. Chem. 1998, 273, 5582–5585. [Google Scholar] [CrossRef] [PubMed]
- Geissler, S.; Zwarg, M.; Knütter, I.; Markwardt, F.; Brandsch, M. The bioactive dipeptide anserine is transported by human proton-coupled peptide transporters. FEBS J. 2010, 277, 790–795. [Google Scholar] [CrossRef] [PubMed]
- Groneberg, D.A.; Fischer, A.; Chung, K.F.; Daniel, H. Molecular mechanisms of pulmonary peptidomimetic drug and peptide transport. Am. J. Respir. Cell Mol. Biol. 2004, 30, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Knütter, I.; Hartrodt, B.; Tóth, G.; Keresztes, A.; Kottra, G.; Mrestani-Klaus, C.; Born, I.; Daniel, H.; Neubert, K.; Brandsch, M. Synthesis and characterization of a new and radiolabeled high-affinity substrate for H+/peptide cotransporters. FEBS J. 2007, 274, 5905–5914. [Google Scholar] [CrossRef]
- Rubio-Aliaga, I.; Daniel, H. Mammalian peptide transporters as targets for drug delivery. Trends Pharmacol. Sci. 2002, 23, 434–440. [Google Scholar] [CrossRef]
- Drozdzik, M.; Busch, D.; Lapczuk, J.; Muller, J.; Ostrowski, M.; Kurzawski, M.; Oswald, S. Protein Abundance of Clinically Relevant Drug Transporters in the Human Liver and Intestine: A Comparative Analysis in Paired Tissue Specimens. Clin. Pharmacol. Ther. 2019, 105, 1204–1212. [Google Scholar] [CrossRef]
- Rühl, A.; Hoppe, S.; Frey, I.; Daniel, H.; Schemann, M. Functional expression of the peptide transporter PEPT2 in the mammalian enteric nervous system. J. Comp. Neurol. 2005, 490, 1–11. [Google Scholar] [CrossRef]
- Smith, D.E.; Pavlova, A.; Berger, U.V.; Hediger, M.A.; Yang, T.; Huang, Y.G.; Schnermann, J.B. Tubular localization and tissue distribution of peptide transporters in rat kidney. Pharm. Res. 1998, 15, 1244–1249. [Google Scholar] [CrossRef]
- Shen, H.; Smith, D.E.; Keep, R.F.; Brosius, F.C., 3rd. Immunolocalization of the proton-coupled oligopeptide transporter PEPT2 in developing rat brain. Mol. Pharm. 2004, 1, 248–256. [Google Scholar] [CrossRef]
- Xu, J.; Zeug, A.; Riederer, B.; Yeruva, S.; Griesbeck, O.; Daniel, H.; Tuo, B.; Ponimaskin, E.; Dong, H.; Seidler, U. Calcium-sensing receptor regulates intestinal dipeptide absorption via Ca(2+) signaling and IK(Ca) activation. Physiol. Rep. 2020, 8, e14337. [Google Scholar] [CrossRef]
- Nielsen, C.U.; Amstrup, J.; Steffansen, B.; Frokjaer, S.; Brodin, B. Epidermal growth factor inhibits glycylsarcosine transport and hPepT1 expression in a human intestinal cell line. Am. J. Physiol. Gastrointest. Liver Physiol. 2001, 281, G191–G199. [Google Scholar] [CrossRef] [PubMed]
- Berlioz, F.; Maoret, J.J.; Paris, H.; Laburthe, M.; Farinotti, R.; Rozé, C. alpha(2)-adrenergic receptors stimulate oligopeptide transport in a human intestinal cell line. J. Pharmacol. Exp. Ther. 2000, 294, 466–472. [Google Scholar] [PubMed]
- Gangopadhyay, A.; Thamotharan, M.; Adibi, S.A. Regulation of oligopeptide transporter (Pept-1) in experimental diabetes. Am. J. Physiol. Gastrointest. Liver Physiol. 2002, 283, G133–G138. [Google Scholar] [CrossRef] [PubMed]
- Wenzel, U.; Kuntz, S.; Diestel, S.; Daniel, H. PEPT1-mediated cefixime uptake into human intestinal epithelial cells is increased by Ca2+ channel blockers. Antimicrob. Agents Chemother. 2002, 46, 1375–1380. [Google Scholar] [CrossRef]
- Pan, X.; Terada, T.; Irie, M.; Saito, H.; Inui, K. Diurnal rhythm of H+-peptide cotransporter in rat small intestine. Am. J. Physiol. Gastrointest. Liver Physiol. 2002, 283, G57–G64. [Google Scholar] [CrossRef]
- Pan, X.; Terada, T.; Okuda, M.; Inui, K. The diurnal rhythm of the intestinal transporters SGLT1 and PEPT1 is regulated by the feeding conditions in rats. J. Nutr. 2004, 134, 2211–2215. [Google Scholar] [CrossRef]
- Ma, K.; Hu, Y.; Smith, D.E. Influence of fed-fasted state on intestinal PEPT1 expression and in vivo pharmacokinetics of glycylsarcosine in wild-type and Pept1 knockout mice. Pharm. Res. 2012, 29, 535–545. [Google Scholar] [CrossRef]
- Thamotharan, M.; Bawani, S.Z.; Zhou, X.; Adibi, S.A. Functional and molecular expression of intestinal oligopeptide transporter (Pept-1) after a brief fast. Metabolism 1999, 48, 681–684. [Google Scholar] [CrossRef]
- Ogihara, H.; Suzuki, T.; Nagamachi, Y.; Inui, K.; Takata, K. Peptide transporter in the rat small intestine: Ultrastructural localization and the effect of starvation and administration of amino acids. Histochem. J. 1999, 31, 169–174. [Google Scholar] [CrossRef]
- McKenna, L.B.; Schug, J.; Vourekas, A.; McKenna, J.B.; Bramswig, N.C.; Friedman, J.R.; Kaestner, K.H. MicroRNAs control intestinal epithelial differentiation, architecture, and barrier function. Gastroenterology 2010, 139, 1654–1664.e1651. [Google Scholar] [CrossRef]
- Runtsch, M.C.; Round, J.L.; O’Connell, R.M. MicroRNAs and the regulation of intestinal homeostasis. Front. Genet. 2014, 5, 347. [Google Scholar] [CrossRef] [PubMed]
- Dalmasso, G.; Nguyen, H.T.; Yan, Y.; Laroui, H.; Charania, M.A.; Obertone, T.S.; Sitaraman, S.V.; Merlin, D. MicroRNA-92b regulates expression of the oligopeptide transporter PepT1 in intestinal epithelial cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 300, G52–G59. [Google Scholar] [CrossRef] [PubMed]
- Hosseinzadeh, Z.; Dong, L.; Bhavsar, S.K.; Warsi, J.; Almilaji, A.; Lang, F. Upregulation of peptide transporters PEPT1 and PEPT2 by Janus kinase JAK2. Cell Physiol. Biochem. 2013, 31, 673–682. [Google Scholar] [CrossRef]
- Warsi, J.; Hosseinzadeh, Z.; Dong, L.; Pakladok, T.; Umbach, A.T.; Bhavsar, S.K.; Shumilina, E.; Lang, F. Effect of Janus kinase 3 on the peptide transporters PEPT1 and PEPT2. J. Membr. Biol. 2013, 246, 885–892. [Google Scholar] [CrossRef]
- Brandsch, M.; Miyamoto, Y.; Ganapathy, V.; Leibach, F.H. Expression and protein kinase C-dependent regulation of peptide/H+ co-transport system in the Caco-2 human colon carcinoma cell line. Biochem. J. 1994, 299 Pt 1, 253–260. [Google Scholar] [CrossRef]
- Chen, H.Q.; Shen, T.Y.; Zhou, Y.K.; Zhang, M.; Chu, Z.X.; Hang, X.M.; Qin, H.L. Lactobacillus plantarum consumption increases PepT1-mediated amino acid absorption by enhancing protein kinase C activity in spontaneously colitic mice. J. Nutr. 2010, 140, 2201–2206. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.; Yoshida, K.; Watanabe, C.; Sai, Y.; Tsuji, A. Screening of the interaction between xenobiotic transporters and PDZ proteins. Pharm. Res. 2004, 21, 1886–1894. [Google Scholar] [CrossRef]
- Noshiro, R.; Anzai, N.; Sakata, T.; Miyazaki, H.; Terada, T.; Shin, H.J.; He, X.; Miura, D.; Inui, K.; Kanai, Y.; et al. The PDZ domain protein PDZK1 interacts with human peptide transporter PEPT2 and enhances its transport activity. Kidney Int. 2006, 70, 275–282. [Google Scholar] [CrossRef]
- Sugiura, T.; Kato, Y.; Kubo, Y.; Tsuji, A. Mutation in an adaptor protein PDZK1 affects transport activity of organic cation transporter OCTNs and oligopeptide transporter PEPT2. Drug Metab. Pharmacokinet. 2006, 21, 375–383. [Google Scholar] [CrossRef]
- Sugiura, T.; Kato, Y.; Wakayama, T.; Silver, D.L.; Kubo, Y.; Iseki, S.; Tsuji, A. PDZK1 regulates two intestinal solute carriers (Slc15a1 and Slc22a5) in mice. Drug Metab. Dispos. 2008, 36, 1181–1188. [Google Scholar] [CrossRef]
- Boehmer, C.; Palmada, M.; Klaus, F.; Jeyaraj, S.; Lindner, R.; Laufer, J.; Daniel, H.; Lang, F. The peptide transporter PEPT2 is targeted by the protein kinase SGK1 and the scaffold protein NHERF2. Cell Physiol. Biochem. 2008, 22, 705–714. [Google Scholar] [CrossRef] [PubMed]
- Viennois, E.; Pujada, A.; Zen, J.; Merlin, D. Function, Regulation, and Pathophysiological Relevance of the POT Superfamily, Specifically PepT1 in Inflammatory Bowel Disease. Compr. Physiol. 2018, 8, 731–760. [Google Scholar] [CrossRef] [PubMed]
- Alghamdi, O.; King, N.; Jones, G.L.; Moens, P.D.J. Effect of ageing and hypertension on the expression and activity of PEPT2 in normal and hypertrophic hearts. Amino Acids 2021, 53, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Xavier, R.J.; Podolsky, D.K. Unravelling the pathogenesis of inflammatory bowel disease. Nature 2007, 448, 427–434. [Google Scholar] [CrossRef]
- Charrier, L.; Merlin, D. The oligopeptide transporter hPepT1: Gateway to the innate immune response. Lab. Investig. 2006, 86, 538–546. [Google Scholar] [CrossRef]
- Ingersoll, S.A.; Ayyadurai, S.; Charania, M.A.; Laroui, H.; Yan, Y.; Merlin, D. The role and pathophysiological relevance of membrane transporter PepT1 in intestinal inflammation and inflammatory bowel disease. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 302, G484–G492. [Google Scholar] [CrossRef]
- Merlin, D.; Si-Tahar, M.; Sitaraman, S.V.; Eastburn, K.; Williams, I.; Liu, X.; Hediger, M.A.; Madara, J.L. Colonic epithelial hPepT1 expression occurs in inflammatory bowel disease: Transport of bacterial peptides influences expression of MHC class 1 molecules. Gastroenterology 2001, 120, 1666–1679. [Google Scholar] [CrossRef]
- Dalmasso, G.; Nguyen, H.T.; Ingersoll, S.A.; Ayyadurai, S.; Laroui, H.; Charania, M.A.; Yan, Y.; Sitaraman, S.V.; Merlin, D. The PepT1-NOD2 signaling pathway aggravates induced colitis in mice. Gastroenterology 2011, 141, 1334–1345. [Google Scholar] [CrossRef]
- Viennois, E.; Ingersoll, S.A.; Ayyadurai, S.; Zhao, Y.; Wang, L.; Zhang, M.; Han, M.K.; Garg, P.; Xiao, B.; Merlin, D. Critical role of PepT1 in promoting colitis-associated cancer and therapeutic benefits of the anti-inflammatory PepT1-mediated tripeptide KPV in a murine model. Cell Mol. Gastroenterol. Hepatol. 2016, 2, 340–357. [Google Scholar] [CrossRef]
- Cekaite, L.; Eide, P.W.; Lind, G.E.; Skotheim, R.I.; Lothe, R.A. MicroRNAs as growth regulators, their function and biomarker status in colorectal cancer. Oncotarget 2016, 7, 6476–6505. [Google Scholar] [CrossRef]
- Ayyadurai, S.; Charania, M.A.; Xiao, B.; Viennois, E.; Zhang, Y.; Merlin, D. Colonic miRNA expression/secretion, regulated by intestinal epithelial PepT1, plays an important role in cell-to-cell communication during colitis. PLoS ONE 2014, 9, e87614. [Google Scholar] [CrossRef] [PubMed]
- Bikhazi, A.B.; Skoury, M.M.; Zwainy, D.S.; Jurjus, A.R.; Kreydiyyeh, S.I.; Smith, D.E.; Audette, K.; Jacques, D. Effect of diabetes mellitus and insulin on the regulation of the PepT 1 symporter in rat jejunum. Mol. Pharm. 2004, 1, 300–308. [Google Scholar] [CrossRef] [PubMed]
- Sawada, K.; Terada, T.; Saito, H.; Hashimoto, Y.; Inui, K. Effects of glibenclamide on glycylsarcosine transport by the rat peptide transporters PEPT1 and PEPT2. Br. J. Pharmacol. 1999, 128, 1159–1164. [Google Scholar] [CrossRef] [PubMed]
- Terada, T.; Sawada, K.; Saito, H.; Hashimoto, Y.; Inui, K. Inhibitory effect of novel oral hypoglycemic agent nateglinide (AY4166) on peptide transporters PEPT1 and PEPT2. Eur. J. Pharmacol. 2000, 392, 11–17. [Google Scholar] [CrossRef]
- Alghamdi, O.A.; King, N.; Andronicos, N.M.; Jones, G.L.; Chami, B.; Witting, P.K.; Moens, P.D.J. Hypertension alters the function and expression profile of the peptide cotransporters PEPT1 and PEPT2 in the rodent renal proximal tubule. Amino Acids 2022, 54, 1001–1011. [Google Scholar] [CrossRef]
- Alghamdi, O.A.; King, N.; Andronicos, N.M.; Jones, G.L.; Chami, B.; Witting, P.K.; Moens, P.D.J. Molecular changes to the rat renal cotransporters PEPT1 and PEPT2 due to ageing. Mol. Cell Biochem. 2019, 452, 71–82. [Google Scholar] [CrossRef]
- Anderle, P.; Nielsen, C.U.; Pinsonneault, J.; Krog, P.L.; Brodin, B.; Sadee, W. Genetic variants of the human dipeptide transporter PEPT1. J. Pharmacol. Exp. Ther. 2006, 316, 636–646. [Google Scholar] [CrossRef]
- Zhang, E.Y.; Fu, D.J.; Pak, Y.A.; Stewart, T.; Mukhopadhyay, N.; Wrighton, S.A.; Hillgren, K.M. Genetic polymorphisms in human proton-dependent dipeptide transporter PEPT1: Implications for the functional role of Pro586. J. Pharmacol. Exp. Ther. 2004, 310, 437–445. [Google Scholar] [CrossRef]
- Fei, Y.J.; Liu, J.C.; Fujita, T.; Liang, R.; Ganapathy, V.; Leibach, F.H. Identification of a potential substrate binding domain in the mammalian peptide transporters PEPT1 and PEPT2 using PEPT1-PEPT2 and PEPT2-PEPT1 chimeras. Biochem. Biophys. Res. Commun. 1998, 246, 39–44. [Google Scholar] [CrossRef]
- Sala-Rabanal, M.; Loo, D.D.; Hirayama, B.A.; Turk, E.; Wright, E.M. Molecular interactions between dipeptides, drugs and the human intestinal H+ -oligopeptide cotransporter hPEPT1. J. Physiol. 2006, 574, 149–166. [Google Scholar] [CrossRef]
- Bolger, M.B.; Haworth, I.S.; Yeung, A.K.; Ann, D.; von Grafenstein, H.; Hamm-Alvarez, S.; Okamoto, C.T.; Kim, K.J.; Basu, S.K.; Wu, S.; et al. Structure, function, and molecular modeling approaches to the study of the intestinal dipeptide transporter PepT1. J. Pharm. Sci. 1998, 87, 1286–1291. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.Z.; Steel, A.; Hediger, M.A. Functional roles of histidine and tyrosine residues in the H(+)-peptide transporter PepT1. Biochem. Biophys. Res. Commun. 2000, 272, 726–730. [Google Scholar] [CrossRef] [PubMed]
- Fei, Y.J.; Liu, W.; Prasad, P.D.; Kekuda, R.; Oblak, T.G.; Ganapathy, V.; Leibach, F.H. Identification of the histidyl residue obligatory for the catalytic activity of the human H+/peptide cotransporters PEPT1 and PEPT2. Biochemistry 1997, 36, 452–460. [Google Scholar] [CrossRef] [PubMed]
- Yeung, A.K.; Basu, S.K.; Wu, S.K.; Chu, C.; Okamoto, C.T.; Hamm-Alvarez, S.F.; von Grafenstein, H.; Shen, W.C.; Kim, K.J.; Bolger, M.B.; et al. Molecular identification of a role for tyrosine 167 in the function of the human intestinal proton- coupled dipeptide transporter (hPepT1). Biochem. Biophys. Res. Commun. 1998, 250, 103–107. [Google Scholar] [CrossRef]
- Meredith, D.; Price, R.A. Molecular Modeling of PepT1—Towards a Structure. J. Membr. Biol. 2006, 213, 79–88. [Google Scholar] [CrossRef]
- Terada, T.; Irie, M.; Okuda, M.; Inui, K. Genetic variant Arg57His in human H+/peptide cotransporter 2 causes a complete loss of transport function. Biochem. Biophys. Res. Commun. 2004, 316, 416–420. [Google Scholar] [CrossRef]
- Terada, T.; Saito, H.; Sawada, K.; Hashimoto, Y.; Inui, K.I. N-terminal halves of rat H+/peptide transporters are responsible for their substrate recognition. Pharm. Res. 2000, 17, 15–20. [Google Scholar] [CrossRef]
- Döring, F.; Martini, C.; Walter, J.; Daniel, H. Importance of a small N-terminal region in mammalian peptide transporters for substrate affinity and function. J. Membr. Biol. 2002, 186, 55–62. [Google Scholar] [CrossRef]
- Sala-Rabanal, M.; Loo, D.D.; Hirayama, B.A.; Wright, E.M. Molecular mechanism of dipeptide and drug transport by the human renal H+/oligopeptide cotransporter hPEPT2. Am. J. Physiol. Renal Physiol. 2008, 294, F1422–F1432. [Google Scholar] [CrossRef]
- Doring, F.; Dorn, D.; Bachfischer, U.; Amasheh, S.; Herget, M.; Daniel, H. Functional analysis of a chimeric mammalian peptide transporter derived from the intestinal and renal isoforms. J. Physiol. 1996, 497 Pt 3, 773–779. [Google Scholar] [CrossRef]
- Klapper, M.; Daniel, H.; Doring, F. Cytosolic COOH terminus of the peptide transporter PEPT2 is involved in apical membrane localization of the protein. Am. J. Physiol. Cell Physiol. 2006, 290, C472–C483. [Google Scholar] [CrossRef] [PubMed]
- Kukuruzinska, M.A.; Lennon, K. Protein N-glycosylation: Molecular genetics and functional significance. Crit. Rev. Oral. Biol. Med. 1998, 9, 415–448. [Google Scholar] [CrossRef] [PubMed]
- Dorn, M.; Jaehme, M.; Weiwad, M.; Markwardt, F.; Rudolph, R.; Brandsch, M.; Bosse-Doenecke, E. The role of N-glycosylation in transport function and surface targeting of the human solute carrier PAT1. FEBS Lett. 2009, 583, 1631–1636. [Google Scholar] [CrossRef] [PubMed]
- Hoover, R.S.; Poch, E.; Monroy, A.; Vázquez, N.; Nishio, T.; Gamba, G.; Hebert, S.C. N-Glycosylation at two sites critically alters thiazide binding and activity of the rat thiazide-sensitive Na(+):Cl(−) cotransporter. J. Am. Soc. Nephrol. 2003, 14, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Stelzl, T.; Baranov, T.; Geillinger, K.E.; Kottra, G.; Daniel, H. Effect of N-glycosylation on the transport activity of the peptide transporter PEPT1. Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 310, G128–G141. [Google Scholar] [CrossRef]
- Chan, T.; Lu, X.; Shams, T.; Zhu, L.; Murray, M.; Zhou, F. The Role of N-Glycosylation in Maintaining the Transporter Activity and Expression of Human Oligopeptide Transporter 1. Mol. Pharm. 2016, 13, 3449–3456. [Google Scholar] [CrossRef]
- Stelzl, T.; Geillinger-Kastle, K.E.; Stolz, J.; Daniel, H. Glycans in the intestinal peptide transporter PEPT1 contribute to function and protect from proteolysis. Am. J. Physiol. Gastrointest. Liver Physiol. 2017, 312, G580–G591. [Google Scholar] [CrossRef]
- Newstead, S.; Drew, D.; Cameron, A.D.; Postis, V.L.; Xia, X.; Fowler, P.W.; Ingram, J.C.; Carpenter, E.P.; Sansom, M.S.; McPherson, M.J.; et al. Crystal structure of a prokaryotic homologue of the mammalian oligopeptide-proton symporters, PepT1 and PepT2. Embo J. 2011, 30, 417–426. [Google Scholar] [CrossRef]
- Solcan, N.; Kwok, J.; Fowler, P.W.; Cameron, A.D.; Drew, D.; Iwata, S.; Newstead, S. Alternating access mechanism in the POT family of oligopeptide transporters. EMBO J. 2012, 31, 3411–3421. [Google Scholar] [CrossRef]
- Doki, S.; Kato, H.E.; Solcan, N.; Iwaki, M.; Koyama, M.; Hattori, M.; Iwase, N.; Tsukazaki, T.; Sugita, Y.; Kandori, H.; et al. Structural basis for dynamic mechanism of proton-coupled symport by the peptide transporter POT. Proc. Natl. Acad. Sci. USA 2013, 110, 11343–11348. [Google Scholar] [CrossRef]
- Guettou, F.; Quistgaard, E.M.; Trésaugues, L.; Moberg, P.; Jegerschöld, C.; Zhu, L.; Jong, A.J.; Nordlund, P.; Löw, C. Structural insights into substrate recognition in proton-dependent oligopeptide transporters. EMBO Rep. 2013, 14, 804–810. [Google Scholar] [CrossRef] [PubMed]
- Boggavarapu, R.; Jeckelmann, J.M.; Harder, D.; Ucurum, Z.; Fotiadis, D. Role of electrostatic interactions for ligand recognition and specificity of peptide transporters. BMC Biol. 2015, 13, 58. [Google Scholar] [CrossRef] [PubMed]
- Stauffer, M.; Jeckelmann, J.M.; Ilgu, H.; Ucurum, Z.; Boggavarapu, R.; Fotiadis, D. Peptide transporter structure reveals binding and action mechanism of a potent PEPT1 and PEPT2 inhibitor. Commun. Chem. 2022, 5, 23. [Google Scholar] [CrossRef] [PubMed]
- Parker, J.L.; Li, C.; Brinth, A.; Wang, Z.; Vogeley, L.; Solcan, N.; Ledderboge-Vucinic, G.; Swanson, J.M.J.; Caffrey, M.; Voth, G.A.; et al. Proton movement and coupling in the POT family of peptide transporters. Proc. Natl. Acad. Sci. USA 2017, 114, 13182–13187. [Google Scholar] [CrossRef]
- Parker, J.L.; Deme, J.C.; Wu, Z.; Kuteyi, G.; Huo, J.; Owens, R.J.; Biggin, P.C.; Lea, S.M.; Newstead, S. Cryo-EM structure of PepT2 reveals structural basis for proton-coupled peptide and prodrug transport in mammals. Sci. Adv. 2021, 7. [Google Scholar] [CrossRef]
- Lyons, J.A.; Parker, J.L.; Solcan, N.; Brinth, A.; Li, D.; Shah, S.T.; Caffrey, M.; Newstead, S. Structural basis for polyspecificity in the POT family of proton-coupled oligopeptide transporters. EMBO Rep. 2014, 15, 886–893. [Google Scholar] [CrossRef]
- Martinez Molledo, M.; Quistgaard, E.M.; Low, C. Tripeptide binding in a proton-dependent oligopeptide transporter. FEBS Lett. 2018, 592, 3239–3247. [Google Scholar] [CrossRef]
- Beale, J.H.; Parker, J.L.; Samsudin, F.; Barrett, A.L.; Senan, A.; Bird, L.E.; Scott, D.; Owens, R.J.; Sansom, M.S.P.; Tucker, S.J.; et al. Crystal Structures of the Extracellular Domain from PepT1 and PepT2 Provide Novel Insights into Mammalian Peptide Transport. Structure 2015, 23, 1889–1899. [Google Scholar] [CrossRef]
- Killer, M.; Wald, J.; Pieprzyk, J.; Marlovits, T.C.; Löw, C. Structural snapshots of human PepT1 and PepT2 reveal mechanistic insights into substrate and drug transport across epithelial membranes. Sci. Adv. 2021, 7, eabk3259. [Google Scholar] [CrossRef]
- Quistgaard, E.M.; Martinez Molledo, M.; Löw, C. Structure determination of a major facilitator peptide transporter: Inward facing PepTSt from Streptococcus thermophilus crystallized in space group P3121. PLoS ONE 2017, 12, e0173126. [Google Scholar] [CrossRef]
- Foley, D.W.; Pathak, R.B.; Phillips, T.R.; Wilson, G.L.; Bailey, P.D.; Pieri, M.; Senan, A.; Meredith, D. Thiodipeptides targeting the intestinal oligopeptide transporter as a general approach to improving oral drug delivery. Eur. J. Med. Chem. 2018, 156, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Tian, C.; Wang, M.; Huang, D.; Wei, W.; Liu, Y.; Li, L.; Sun, B.; Kou, L.; Kan, Q.; et al. Dipeptide-modified nanoparticles to facilitate oral docetaxel delivery: New insights into PepT1-mediated targeting strategy. Drug Deliv. 2018, 25, 1403–1413. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Sun, M.; Wang, D.; Li, G.; Huang, J.; Tan, S.; Bao, L.; Li, Q.; Li, G.; Si, L. A PepT1 mediated medicinal nano-system for targeted delivery of cyclosporine A to alleviate acute severe ulcerative colitis. Biomater. Sci. 2019, 7, 4299–4309. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Zhang, J.; Tong, P.; Gao, Y.; Lv, Y.; Wang, C.; Luo, M.; Sun, M.; Wang, J.; Feng, Y.; et al. Bioactivatable Pseudotripeptidization of Cyclic Dipeptides To Increase the Affinity toward Oligopeptide Transporter 1 for Enhanced Oral Absorption: An Application to Cyclo(l-Hyp-l-Ser) (JBP485). J. Med. Chem. 2019, 62, 7708–7721. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Gan, W.; Lei, M.; Jiang, W.; Cheng, M.; He, J.; Sun, Q.; Liu, W.; Hu, L.; Jin, Y. PEPT1-mediated prodrug strategy for oral delivery of peramivir. Asian J. Pharm. Sci. 2018, 13, 555–565. [Google Scholar] [CrossRef]
- Yuri, T.; Kono, Y.; Okada, T.; Terada, T.; Miyauchi, S.; Fujita, T. Transport Characteristics of 5-Aminosalicylic Acid Derivatives Conjugated with Amino Acids via Human H(+)-Coupled Oligopeptide Transporter PEPT1. Biol. Pharm. Bull. 2020, 43, 697–706. [Google Scholar] [CrossRef] [PubMed]
- Khavinson, V.K.; Linkova, N.S.; Rudskoy, A.I.; Petukhov, M.G. Feasibility of Transport of 26 Biologically Active Ultrashort Peptides via LAT and PEPT Family Transporters. Biomolecules 2023, 13, 552. [Google Scholar] [CrossRef]
- Tashima, T. Intriguing possibilities and beneficial aspects of transporter-conscious drug design. Bioorg Med. Chem. 2015, 23, 4119–4131. [Google Scholar] [CrossRef]
- Robison, S.W.; Li, J.; Viera, L.; Blackburn, J.P.; Patel, R.P.; Blalock, J.E.; Gaggar, A.; Xu, X. A mechanism for matrikine regulation in acute inflammatory lung injury. JCI Insight 2021, 6, e140750. [Google Scholar] [CrossRef]
- Matthay, M.A.; Zemans, R.L. The Acute Respiratory Distress Syndrome: Pathogenesis and Treatment. Annu. Rev. Pathol. Mech. Dis. 2011, 6, 147–163. [Google Scholar] [CrossRef]
- Li, S.; Jiang, L.; Li, X.; Lin, F.; Wang, Y.; Li, B.; Jiang, T.; An, W.; Liu, S.; Liu, H.; et al. Clinical and pathological investigation of patients with severe COVID-19. JCI Insight 2020, 5, e138070. [Google Scholar] [CrossRef] [PubMed]
- Gerloff, T. Impact of genetic polymorphisms in transmembrane carrier-systems on drug and xenobiotic distribution. Naunyn Schmiedebergs Arch. Pharmacol. 2004, 369, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Tang, A.M.; Tan, Y.L.; Limenta, L.M.; Lee, E.J. Effects of sodium bicarbonate and ammonium chloride pre-treatments on PEPT2 (SLC15A2) mediated renal clearance of cephalexin in healthy subjects. Drug Metab. Pharmacokinet. 2011, 26, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Zheng, J.; Zhu, L.; Jodal, A.; Cui, P.H.; Wong, M.; Gurney, H.; Church, W.B.; Murray, M. Functional analysis of novel polymorphisms in the human SLCO1A2 gene that encodes the transporter OATP1A2. AAPS J. 2013, 15, 1099–1108. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Mutant | Putative Location | Transport Function | References |
|---|---|---|---|
| Y12A | TMD1 | Gly-Sar uptake - | [82] |
| F28Y | Extracellular Loop 1 | Cephalexin uptake ↓ | [78] |
| Y56A | TMD2 | Gly-Sar uptake - | [83] |
| Y56F | TMD2 | Gly-Sar uptake ↓ | [83] |
| H57N | TMD2 | Gly-Sar uptake ↓↓ | [84] |
| H57Q | TMD2 | Gly-Sar uptake ↓↓ | [84] |
| H57R | TMD2 | Gly-Sar uptake - | [83] |
| Y64A | TMD2 | Gly-Sar uptake - | [83] |
| Y64F | TMD2 | Gly-Sar uptake ↓ | [83] |
| H111C | Extracellular Loop 2 | Gly-Sar uptake - | [83] |
| H111R | Extracellular Loop 2 | Gly-Sar uptake - | [83] |
| S117N | Extracellular Loop 2 | Gly-Sar uptake - | [81] |
| H121C | TMD4 | Gly-Sar uptake ↓ | [83] |
| H121N | TMD4 | Gly-Sar uptake - | [84] |
| H121Q | TMD4 | Gly-Sar uptake - | [84] |
| H121R | TMD4 | Gly-Sar uptake ↓ | [83] |
| Y167A | TMD5 | Gly-Sar uptake ↓↓ | [85] |
| Y167F | TMD5 | Gly-Sar uptake ↓↓ | [85] |
| Y167H | TMD5 | Gly-Sar uptake ↓↓ | [85] |
| Y167S | TMD5 | Gly-Sar uptake ↓↓ | [85] |
| H260N | Intracellular Loop 3 | Gly-Sar uptake - | [84] |
| H260Q | Intracellular Loop 3 | Gly-Sar uptake - | [84] |
| R282A | TMD7 | Gly-Sar uptake - | [82] |
| W294A | TMD7 | Gly-Sar uptake ↓↓ | [82,86] |
| G419A | Extracellular Loop 5 | Gly-Sar uptake- | [81] |
| P586L | TMD10 | Glycyl-sarcosine uptake ↓ | [78] |
| E595A | TMD10 | Gly-Sar uptake ↓↓ | [82] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, Y.; Gao, J.; Jiang, X.; Zhu, L.; Zhou, Q.T.; Murray, M.; Li, J.; Zhou, F. Molecular Insights to the Structure-Interaction Relationships of Human Proton-Coupled Oligopeptide Transporters (PepTs). Pharmaceutics 2023, 15, 2517. https://doi.org/10.3390/pharmaceutics15102517
Luo Y, Gao J, Jiang X, Zhu L, Zhou QT, Murray M, Li J, Zhou F. Molecular Insights to the Structure-Interaction Relationships of Human Proton-Coupled Oligopeptide Transporters (PepTs). Pharmaceutics. 2023; 15(10):2517. https://doi.org/10.3390/pharmaceutics15102517
Chicago/Turabian StyleLuo, Yining, Jingchun Gao, Xukai Jiang, Ling Zhu, Qi Tony Zhou, Michael Murray, Jian Li, and Fanfan Zhou. 2023. "Molecular Insights to the Structure-Interaction Relationships of Human Proton-Coupled Oligopeptide Transporters (PepTs)" Pharmaceutics 15, no. 10: 2517. https://doi.org/10.3390/pharmaceutics15102517
APA StyleLuo, Y., Gao, J., Jiang, X., Zhu, L., Zhou, Q. T., Murray, M., Li, J., & Zhou, F. (2023). Molecular Insights to the Structure-Interaction Relationships of Human Proton-Coupled Oligopeptide Transporters (PepTs). Pharmaceutics, 15(10), 2517. https://doi.org/10.3390/pharmaceutics15102517