
Extracellular Vesicles of Probiotics: Shedding Light on the Biological Activity and Future Applications




Abstract
:1. Introduction
2. Review Strategy and Literature Included
3. Discussion
3.1. Physicochemical Properties of EVs Produced by Probiotics

3.2. Proteomic Profile of EVs Produced by Probiotics

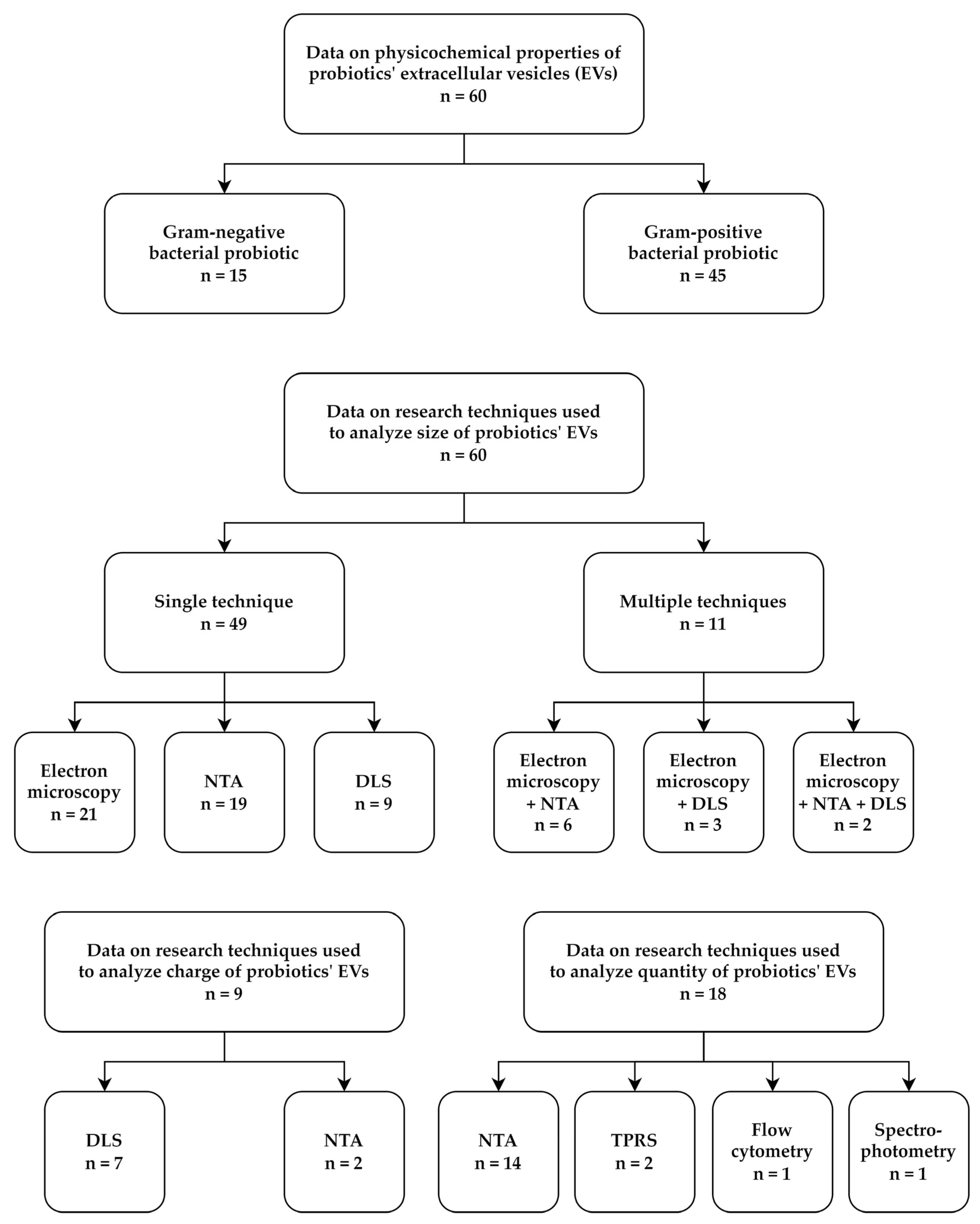{kind=link}
{kind=link}
| Bacterial Producer | Methodology of Isolation/Determination | Proteomic Data | Reference | ||
|---|---|---|---|---|---|
| Total Number of Identified Proteins * | Cellular Localization of Proteins | The Most Abundant Representatives/ Proteins Highlighted by Authors | |||
| Escherichia coli Nissle 1917 | Ultracentrifugation (vesicles) Electrophoresis + LC-MS/MS (proteome) | 192 | Outer membrane ~ 40% Cytoplasm 40% Periplasm 15% Inner membrane < 5 % |
| [52] |
| Ultracentrifugation + DGC (vesicles) Electrophoresis + LC-MS/MS (proteome) | 189 | Outer membrane 28% Cytoplasm 36.5% Periplasm 20% Inner membrane 9.5% Secretory 6% |
| [29] | |
| Ultracentrifugation (vesicles) Electrophoresis + LC-MS/MS (proteome) | 295 | ND |
| [51] | |
| Ultracentrifugation + DGC or SEC (vesicles) Chemical precipitation + LC-MS/MS (proteome) | 189 | Membrane ~ 60% Cytoplasm ~ 40% |
| [72] | |
| Lactiplantibacillus plantarum BGAN8 | Ultracentrifugation (vesicles) Electrophoresis + MALDI-TOF (proteome) | 1149 | Membrane ~ 45% Cytoplasm + ribosomes ~ 52% Secretory < 1% |
| [54] |
| Lactiplantibacillus plantarum WCFS1 | Chemical precipitation (vesicles) Electrophoresis + LC-MS/MS (proteome) | 31 | Membrane 42% Cytoplasm 13% Secretory 16% |
| [32] |
| Lacticaseibacillus casei ATCC 393 | Ultracentrifugation (vesicles) Chemical precipitation + LC-MS/MS (proteome) | 43 | Membrane ~ 20% Cytoplasm 65% Secretory 14% |
| [38] |
| Lacticaseibacillus casei BL23 | Ultracentrifugation (vesicles) Electrophoresis + LC-MS/MS (proteome) | 103 | Membrane + secretory 43% Cytoplasm 57% |
| [36] |
| Limosilactobacillus reuteri ATCC 23272 | Ultracentrifugation (vesicles) Chemical precipitation + LC-MS/MS (proteome) | 17 | Membrane 18% Cytoplasm 82% |
| [38] |
| Limosilactobacillus reuteri BBC3 | Ultracentrifugation + DGC (vesicles) Chemical precipitation + LC-MS/MS (proteome) | 92 | Membrane 27% Cytoplasm 56.5% Secretory 16% |
| [34] |
| Ligilactobacillus animalis ATCC 35046 | Ultracentrifugation + DGC (vesicles) Chemical precipitation + nLC-MS/MS (proteome) | 340 | From the top 74 proteins: Membrane 25.7% Cytoplasm 25.7% Secretory 1.3% Unknown 47.3% |
| [76] |
| Lactobacillus acidophilus ATCC 53544 | Ultracentrifugation (vesicles) Chemical precipitation + LC-MS/MS (proteome) | 26 | Membrane ~ 30% Cytoplasm 62% Secretory 12% |
| [38] |
| Lactobacillus gasseri BC12 | Ultracentrifugation (vesicles) Electrophoresis + LC-ESI-MS/MS (proteome) | 15 | Membrane ~ 44% Cytoplasm ~ 44% Secretory ~ 11% |
| [74] |
| Lactobacillus crispatus BC5 | Ultracentrifugation (vesicles) Electrophoresis + LC-ESI-MS/MS (proteome) | 11 | Membrane ~ 45% Cytoplasm ~ 45% Secretory ~ 9% |
| [74] |
| Lactobacillus johnsonii N6.2 | Ultracentrifugation (vesicles) Electrophoresis + LC-MS/MS (proteome) | 366 | Cytoplasm + ribosomes 86% Secretory 14% |
| [33] |
| Bifidobacterium longum NCC 2705 | Ultracentrifugation (vesicles) Electrophoresis + LC-MS/MS (proteome) | 24 | Membrane 21% Cytoplasm 75% |
| [64] |
| Propionibacterium freudenreichii CIRM-BIA 129 | SEC (vesicles) Electrophoresis + LC-ESI-MS/MS (proteome) | 319 | Membrane 16% Cytoplasm 75% Secretory 9% |
| [78] |
| SEC (vesicles) Electrophoresis + LC-MS/MS (proteome) | 391 (medium-dependent; 358 common for all) | Membrane 16.5% Cytoplasm 74% Secretory 9.5% |
| [79] | |
| Lactococcus lactis FM-YL11 | Ultracentrifugation (vesicles) Magnetic precipitation + LC-MS/MS (proteome) | 1283 | From the top 320 proteins: Membrane 16.5% Cytoplasm 74% Secretory 9.5% |
| [62] |
| Pediococcus pentosaceus ** | Ultracentrifugation (vesicles) Electrophoresis + LC-MS/MS (proteome) | 103 | Membrane 9% Cytoplasm + ribosomes ~ 83% Secretory 5.5% |
| [42] |
3.3. Biological Activities and Properties of EVs Produced by Probiotics
| Bacterial Producer | In Vitro Model EVs Treatment (Duration) | In Vivo Model EVs Treatment (Duration) | Observations on EVs Activity | EVs Properties | Reference |
|---|---|---|---|---|---|
| Escherichia coli Nissle 1917 | Caco-2 and T-84 cells (EPEC-infected) 0.1 mg/mL (24 h) | ND |
|
| [131] |
| RAW264.7 murine macrophages1 µg/mL (16 h) | ND |
|
| [29] | |
| OVA-Escherichia coli Nissle 1917, Escherichia coli BL21 ΔompA | ND | Tumor in C57BL/6 and BALB/c female 4–8-week-old mice (administration: oral gavage) 10 µg (3–5 times) |
|
| [122] |
| Escherichia coli serotype O6:K5:H1 | Caco-2 and HT-29 cells 10 mg/mL (8 h) | ND |
|
| [132] |
| Caco-2 and PMBCs cells 50 µg/mL (5–24 h) | Colon organ culture (ex vivo model) 50 µg/mL (5 h) |
|
| [63] | |
| Human Monocyte-Derived DCs 10 µg/mL (24 h) | ND |
|
| [133] | |
| Akkermansia muciniphila ATCC BAA-835 | Caco-2 cells 0.1, 0.5, and 5 μg (24 h) | ND |
|
| [68] |
| Caco-2 cells 10 μg (24 h) | HFD induced and ND thirty male C57BL/6 mice (administration: oral gavage) 10 μg (5 weeks) |
|
| [67] | |
| Akkermansia muciniphila ATCC BAA-835 | ND | NFD induced and NF 8-week-old male C57BL/6 mice (administration: oral gavage) 10 μg protein/200 μL (5 weeks) |
|
| [69] |
| Caco-2 cells (inflammation model) 0.1, 1, and 10 μg (4–8 h) | HFD in 6–8-week-old male C57BL/6 mice (administration: oral gavage) 10 μg (14 days) |
|
| [91] | |
| THP-1 and RAW264.7 10 μg/mL (24 h) | Prostate cancer RM-1 mice model (administration: injection) 40 μg per mouse (13 days) |
|
| [92] | |
| LX-2 cells (inflammation model) 1, 10, 50 µg/mL (24 h) | Chronic liver injury in 7–8-week-old male C57BL/6 mice (administration: intraperitoneal injection) 50 µg protein/200 µL (4 weeks) |
|
| [70] | |
| Caco-2 and Hep-G2 cells 50, 100 µg/mL (24 h) | ND |
|
| [53] | |
| Akkermansia muciniphila ATCC BAA-835 | LX-2 cells (inflammation model) 1, 10, 50 µg/mL (24 h) | Livery injury in 8-week-old male C57BL/6 mice (administration: oral gavage) 50 µg protein/200 µL (4 weeks) |
|
| [134] |
| A. muciniphila ATCC BAA-835, Faecalibacterium prausnitzii A2-165c | Caco-2 cells 1 and 50 μg/mL (24 h) | ND |
|
| [15] |
| A. muciniphila ATCC BAA-835, L. plantarum KCTC 11401BP, Bacillus subtilis * | HT22 cells (stress model) 20 μg (24 h) | Chronic stress in 7-week-old male C57BL6 mice (administration: intraperitoneal injection) 6 μg/100 μL mouse per day (14 days) |
|
| [130] |
| Lactiplantibacillus plantarum KCTC 11401BP | HT22 cells (stress model) 20 μg/mL (24 h) | Depression in 7-week-old male C57BL/6J mice (administration: intraperitoneal injection) 0.1, 0.18, and 0.27 μg/kg (1–35 days) |
|
| [129] |
| HaCaT cells and keratinocytes 0.1, 1, and 10 μg/mL (12 h) | S. aureus atopic dermatitis-induced mouse model (administration: oral gavage) |
|
| [55] | |
| Lactiplantibacillus plantarum APsulloc 331261 | THP1 cells 10 μg/mL (48 h) | Human skin organ culture (ex vivo) 50 μg/mL (2–4 days) |
|
| [50] |
| Lactiplantibacillus plantarum WCFS1 | Caco-2 cells 500 µL (24 h) | C. elegans Bristol N2 EVs isolated from 109 CFU (1–15 days) |
|
| [32] |
| Lactiplantibacillus plantarum Q7 | ND | Colitis in 4–5-week-old SPF male C57BL/6J mice(administration: gavage) 10/20 mg Q7-EVs group (0.5/1 mg/kg body weight) (1–18 days) |
|
| [93] |
| Lactiplantibacillus plantarum * | CCD-986Sk cells 0.625%, 1.25%, 5%, and 10% EVs (24 h) | Korean women in their 50s (administration: topically on the skin) (twice a day, 4 weeks) |
|
| [87] |
| Lactiplantibacillus plantarum YW11 | Primary cortical neurons from C57BL/6 mice (OGD model) Co-culturing with EVs (24 h) | tMCAO (ischemic stroke model) in 10–12-week-old male C57BL/6 mice (administration: injection through the tail vein) 100 μg/day (3 days) |
|
| [86] |
| Lacticaseibacillus casei BL23 | T84 and HT-29 cells 20 ng/mL to 10 μg/mL (24 h) | ND |
|
| [35] |
| Lacticaseibacillus casei ATCC 393 | Caco-2 cells 100 and 150 μg/mL (24 h) | ND |
|
| [37] |
| L. casei DSMZ 20011, L. plantarum NCIMB 8826 | Caco-2 and THP-1 cells (inflammation model) 5 × 1011–5 × 1012 EVs/mL (24 h) | ND |
|
| [83] |
| THP-1 cells (inflammation model) 1:2 EVs per well (48 h) | ND |
|
| [31] | |
| Lacticaseibacillus paracasei * | RAW 264.7 cells (inflammation model) 0.1, 1, 10, 50 μg/mL (12 h) HT 29 cells (inflammation model) 500 ng/mL (12 h) | Acute colitis-induced 7-week-old male C57BL/6 mice (administration: oral gavage) 5 mg/day (12 days) |
|
| [58] |
| Lacticaseibacillus paracasei PC-H1 | Colorectal cancer cell line, HCT116, SW1116, and SW620 cells 200 μg/mL (24 h) | 4-week-old female BALB/c nude mice (administration with HCT116 and EVs through subcutaneous injection) 200 μg/mL (30 days) |
|
| [90] |
| Lactobacillus crispatus BC3, BC5; Lactobacillus gasseri BC12, BC13 | Human T-lymphocyte MT-4 and Jurkat-tat cell lines 50 μL (1–72 h) | Human tissue cultures (ex vivo model) 108 EVs/mL (12 days) |
|
| [74] |
| Limosilactobacillus reuteri BBC3 | HD11 cells 10 µg/mL (6 h or 12 h) Splenic lymphocytes 10 μg/mL (12 h) | Broiler chicks (inflammation model) (administration: oral gavage) 200 μg/bird (21 days) Jejunum explant culture (ex vivo model) (inflammation model) 10 µg/mL (6 h) |
|
| [34] |
| Lacticaseibacillus rhamnosus GG | HepG2 cells 50, 100, 150, and 200 μg (24 h) | ND |
|
| [56] |
| SW480 and HT 29 cells (human colon cancer cell lines) 5–200 µg/mL (24 h) | ND |
|
| [57] | |
| ND | Colitis-induced 4–5-week-old C57BL/6J male mice (administration: oral gavage) 1.2 mg/kg of body weight (14 days) |
|
| [94] | |
| Lacticaseibacillus rhamnosus JB-1 | HT-29 and MODE-K cells 3 × 1010 EVs (2 h) | 8- to 10-week-old SPF BALB/c male mice (administration: oral gavage) 3 × 1010 EVs (2 h) |
|
| [88] |
| L. rhamnosus GG, L. reuteri DSM 17938 | PBMCs cells 500:1, 100:1, and 20:1 (48 h) | ND |
|
| [135] |
| Lentilactobacillus kefirgranum PRCC-1301 | Caco-2 and HCT116 cells 0, 10, and 100 µg/mL (6–48 h) | Colitis-induced 6-week-old male C57BL/6 mice (administration: oral gavage) 3 mg/kg (3–14 days) |
|
| [118] |
| Lentilactobacillus kefir KCTC 3611, Lentilactobacillus kefiranofaciens KCTC 5075, Lentilactobacillus kefirgranum KCTC 5086 | Caco-2 cells (inflammation model) 1 × 109 EVs/mL (24 h) | IBD-induced 8-week-old male BALB mice (administration: oral gavage) (3 × 108 or 3 × 1010 EVs/head) |
|
| [60] |
| Latilactobacillus sakei NBRC 15893 | PP and BMDCs from BALB/c mice (female, 7–14 weeks old) 30 μg protein/mL (4 days) | ND |
|
| [125] |
| PP cells 37 μg/mL EVs (1–4 days) | ND |
|
| [61] | |
| Ligilactobacillus animalis ATCC 35046 | HMECs, MLO-Y4, MC3T3-E1, and BMSCs(MPS-treated) 10 μg/mL (6–24 h) | GC-induced ONFH male C57BL/6J mice (administration: oral gavage) 30 μg/200 μL (once a week) |
|
| [76] |
| Lactobacillus johnsonii N6.2 | Pancreatic cell line βlox5, Caco-2, Jurkat, and THP-1 cells 108 or 1010 EVs/mL (2–8 h) | Pancreatic islets isolated from human donors(ex vivo model) 6 × 109 EVs/mL (5 h) |
|
| [136] |
| Lactococcus lactis * | Dendritic cells isolated from asthmatic patients 10 μg/mL (24 h) | Allergic asthma-induced 6-week-old female BALB/c mice (administration: intranasally) 10 μg/20 μL PBS (5 days) |
|
| [95] |
| Bifidobacterium longum KACC 91563 | PP, T cells, B cells, eosinophils, and BMCCs from mice 2 µg/mL (2 h) | Food allergy-induced 6- to 8-week-old BALB/c mice (administration: oral gavage) EVs from 109 CFU/mouse (2 weeks) |
|
| [137] |
| Bifidobacterium bifidum LMG 13195 | Monocyte-derived DCs and naïve T cells 0.1 μg/mL (48 h) | ND |
|
| [138] |
| B. longum *, L. plantarum WCFS1 | DC2.4 and RAW264.7 cells 0.01 or 0.1 μg/well (6–24 h) | ND |
|
| [85] |
| DC2.4 and RAW264.7 cells 0.5 μg/well (6 h) | ND |
|
| [84] | |
| Propionibacterium freudenreichii CIRM-BIA 129 | HT-29 cells (inflammation model) 109 EVs/mL (1 h) | ND |
|
| [78] |
| HT-29 cells (inflammation model) 109 EVs/mL (24 h) | ND |
|
| [79] | |
| Bacillus subtilis 168 | Caco-2 cells 1.3 × 109 EVs (0–4 h) | ND |
|
| [80] |
| Clostridium butyricum MIYAIRI 588 | ND | Ulcerative colitis in 40–60-day-old male C57BL/6 mice (administration: intragastrically) 15 μg/200 μL (once a day, 5 days) |
|
| [82] |
| ND | Ulcerative colitis-induced male C57BL6J mice (administration: oral gavage) 50 μg/day (11 days) |
|
| [81] | |
| Leuconostoc holzapfelii GFC1203H, L. plantarum *, B. longum *, B. animalis *, L. acidophilus * | Human HFDP cells 1, 2.5, 5, and 10 μg/mL (6 h, 12 h, 24 h) | ND |
|
| [77] |
| Pediococcus pentosaceus *, Ligilactobacillus salivarius * | E.G7–EL4 and HEK-BLUE hTLR2, BMDCsMouse splenocytes (inflammation model) 0.2, 1, and 5 mg/mL (24 h) | Liver-fibrosis in 6- to 8-week-old male C57BL/6 mice (administration: injection) 10 μg/mouse (14 days) |
|
| [42] |
3.4. Future Application and Perspectives on EVs Produced by Probiotics
3.5. Challenges and Limitations of Articles Focusing on EVs Produced by Probiotics
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- You, S.; Ma, Y.; Yan, B.; Pei, W.; Wu, Q.; Ding, C.; Huang, C. The Promotion Mechanism of Prebiotics for Probiotics: A Review. Front. Nutr. 2022, 9, 1000517. [Google Scholar] [CrossRef]
- Ogunrinola, G.A.; Oyewale, J.O.; Oshamika, O.O.; Olasehinde, G.I. The Human Microbiome and Its Impacts on Health. Int. J. Microbiol. 2020, 2020, 8045646. [Google Scholar] [CrossRef]
- Hou, K.; Wu, Z.X.; Chen, X.Y.; Wang, J.Q.; Zhang, D.; Xiao, C.; Zhu, D.; Koya, J.B.; Wei, L.; Li, J.; et al. Microbiota in Health and Diseases. Signal Transduct. Target. Ther. 2022, 7, 135. [Google Scholar] [CrossRef]
- Depoorter, L.; Vandenplas, Y. Probiotics in Pediatrics. A Review and Practical Guide. Nutrients 2021, 13, 2176. [Google Scholar] [CrossRef]
- Varela-Trinidad, G.U.; Domínguez-Díaz, C.; Solórzano-Castanedo, K.; Íñiguez-Gutiérrez, L.; Hernández-Flores, T.D.J.; Fafutis-Morris, M. Probiotics: Protecting Our Health from the Gut. Microorganisms 2022, 10, 1428. [Google Scholar] [CrossRef]
- Ranjha, M.M.A.N.; Shafique, B.; Batool, M.; Kowalczewski, P.Ł.; Shehzad, Q.; Usman, M.; Manzoor, M.F.; Zahra, S.M.; Yaqub, S.; Aadil, R.M. Nutritional and Health Potential of Probiotics: A Review. Appl. Sci. 2021, 11, 11204. [Google Scholar] [CrossRef]
- Wang, G.; Chen, Y.; Xia, Y.; Song, X.; Ai, L. Characteristics of Probiotic Preparations and Their Applications. Foods 2022, 11, 2472. [Google Scholar] [CrossRef]
- Thorakkattu, P.; Khanashyam, A.C.; Shah, K.; Babu, K.S.; Mundanat, A.S.; Deliephan, A.; Deokar, G.S.; Santivarangkna, C.; Nirmal, N.P. Postbiotics: Current Trends in Food and Pharmaceutical Industry. Foods 2022, 11, 3094. [Google Scholar] [CrossRef]
- Maccelli, A.; Carradori, S.; Puca, V.; Sisto, F.; Lanuti, P.; Crestoni, M.E.; Lasalvia, A.; Muraro, R.; Bysell, H.; Sotto, A.D.; et al. Correlation between the Antimicrobial Activity and Metabolic Profiles of Cell Free Supernatants and Membrane Vesicles Produced by Lactobacillus reuteri DSM 17938. Microorganisms 2020, 8, 1653. [Google Scholar] [CrossRef]
- Sabahi, S.; Homayouni Rad, A.; Aghebati-Maleki, L.; Sangtarash, N.; Ozma, M.A.; Karimi, A.; Hosseini, H.; Abbasi, A. Postbiotics as the New Frontier in Food and Pharmaceutical Research. Crit. Rev. Food Sci. Nutr. 2022, 1–28. [Google Scholar] [CrossRef]
- Srivastava, P.; Kim, K. Membrane Vesicles Derived from Gut Microbiota and Probiotics: Cutting-Edge Therapeutic Approaches for Multidrug-Resistant Superbugs Linked to Neurological Anomalies. Pharmaceutics 2022, 14, 2370. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, S.; Sabharwal, V.; Kaushik, P.; Joshi, A.; Aayushi, A.; Suri, M. Postbiotics: From Emerging Concept to Application. Front. Sustain. Food Syst. 2022, 6, 887642. [Google Scholar] [CrossRef]
- Viswanathan, K.; Muthusamy, S. Review on the Current Trends and Future Perspectives of Postbiotics for Developing Healtheir Foods. eFood 2022, 3, e47. [Google Scholar] [CrossRef]
- Salminen, S.; Collado, M.C.; Endo, A.; Hill, C.; Lebeer, S.; Quigley, E.M.M.; Sanders, M.E.; Shamir, R.; Swann, J.R.; Szajewska, H.; et al. The International Scientific Association of Probiotics and Prebiotics (ISAPP) Consensus Statement on the Definition and Scope of Postbiotics. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 649–667. [Google Scholar] [CrossRef]
- Zou, C.; Zhang, Y.; Liu, H.; Wu, Y.; Zhou, X. Extracellular Vesicles: Recent Insights Into the Interaction Between Host and Pathogenic Bacteria. Front. Immunol. 2022, 13, 840550. [Google Scholar] [CrossRef]
- Combo, S.; Mendes, S.; Nielsen, K.M.; da Silva, G.J.; Domingues, S. The Discovery of the Role of Outer Membrane Vesicles against Bacteria. Biomedicines 2022, 10, 2399. [Google Scholar] [CrossRef]
- Srivatsav, A.T.; Kapoor, S. The Emerging World of Membrane Vesicles: Functional Relevance, Theranostic Avenues and Tools for Investigating Membrane Function. Front. Mol. Biosci. 2021, 8, 59. [Google Scholar] [CrossRef]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal Information for Studies of Extracellular Vesicles 2018 (MISEV2018): A Position Statement of the International Society for Extracellular Vesicles and Update of the MISEV2014 Guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, Q.; Wang, S.; Weng, W.; Jing, Y.; Su, J. Bacterial Extracellular Vesicles as Bioactive Nanocarriers for Drug Delivery: Advances and Perspectives. Bioact. Mater. 2022, 14, 169–181. [Google Scholar] [CrossRef]
- Huang, Y.; Nieh, M.P.; Chen, W.; Lei, Y. Outer Membrane Vesicles (OMVs) Enabled Bio-applications: A Critical Review. Biotechnol. Bioeng. 2022, 119, 34–47. [Google Scholar] [CrossRef]
- Macion, A.; Wyszyńska, A.; Godlewska, R. Delivery of Toxins and Effectors by Bacterial Membrane Vesicles. Toxins 2021, 13, 845. [Google Scholar] [CrossRef]
- Wang, W.; Chanda, W.; Zhong, M. The Relationship between Biofilm and Outer Membrane Vesicles: A Novel Therapy Overview. FEMS Microbiol. Lett. 2015, 362, fnv117. [Google Scholar] [CrossRef]
- Reid, G.; Gadir, A.A.; Dhir, R. Probiotics: Reiterating What They Are and What They Are Not. Front. Microbiol. 2019, 10, 424. [Google Scholar] [CrossRef] [PubMed]
- Quinto, E.J.; Jiménez, P.; Caro, I.; Tejero, J.; Mateo, J.; Girbés, T.; Quinto, E.J.; Jiménez, P.; Caro, I.; Tejero, J.; et al. Probiotic Lactic Acid Bacteria: A Review. Food Nutr. Sci. 2014, 5, 1765–1775. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, J.; Lv, M.; Shao, Z.; Hungwe, M.; Wang, J.; Bai, X.; Xie, J.; Wang, Y.; Geng, W. Metabolism Characteristics of Lactic Acid Bacteria and the Expanding Applications in Food Industry. Front. Bioeng. Biotechnol. 2021, 9, 612285. [Google Scholar] [CrossRef] [PubMed]
- Nataraj, B.H.; Shivanna, S.K.; Rao, P.; Nagpal, R.; Behare, P.V. Evolutionary Concepts in the Functional Biotics Arena: A Mini-review. Food Sci. Biotechnol. 2021, 30, 487–496. [Google Scholar] [CrossRef]
- Derrien, M.; Vaughan, E.E.; Plugge, C.M.; de Vos, W.M. Akkermansia muciniphila gen. nov., sp. nov., a Human Intestinal Mucin-Degrading Bacterium. Int. J. Syst. Evol. Microbiol. 2004, 54, 1469–1476. [Google Scholar] [CrossRef]
- Si, J.; Kang, H.; You, H.J.; Ko, G.P. Revisiting the Role of Akkermansia muciniphila as a Therapeutic Bacterium. Gut Microbes 2022, 14, 2078619. [Google Scholar] [CrossRef] [PubMed]
- Hu, R.; Lin, H.; Li, J.; Zhao, Y.; Wang, M.; Sun, X.; Min, Y.; Gao, Y.; Yang, M. Probiotic Escherichia coli Nissle 1917-Derived Outer Membrane Vesicles Enhance Immunomodulation and Antimicrobial Activity in RAW264.7 Macrophages. BMC Microbiol. 2020, 20, 268. [Google Scholar] [CrossRef]
- Liu, H.; Zhao, F.; Zhang, K.; Zhao, J.; Wang, Y. Investigating the Growth Performance, Meat Quality, Immune Function and Proteomic Profiles of Plasmal Exosomes in Lactobacillus plantarum-Treated Broilers with Immunological Stress. Food Funct. 2021, 12, 11790–11807. [Google Scholar] [CrossRef]
- Müller, L.; Kuhn, T.; Koch, M.; Fuhrmann, G. Stimulation of Probiotic Bacteria Induces Release of Membrane Vesicles with Augmented Anti-inflammatory Activity. ACS Appl. Bio Mater. 2021, 4, 3739–3748. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Lee, K.; Hsu, M.; Nau, G.; Mylonakis, E.; Ramratnam, B. Lactobacillus-Derived Extracellular Vesicles Enhance Host Immune Responses against Vancomycin-Resistant Enterococci. BMC Microbiol. 2017, 17, 66. [Google Scholar] [CrossRef]
- Harrison, N.A.; Gardner, C.L.; da Silva, D.R.; Gonzalez, C.F.; Lorca, G.L. Identification of Biomarkers for Systemic Distribution of Nanovesicles from Lactobacillus johnsonii N6.2. Front. Immunol. 2021, 12, 3491. [Google Scholar] [CrossRef] [PubMed]
- Hu, R.; Lin, H.; Wang, M.; Zhao, Y.; Liu, H.; Min, Y.; Yang, X.; Gao, Y.; Yang, M. Lactobacillus reuteri-Derived Extracellular Vesicles Maintain Intestinal Immune Homeostasis against Lipopolysaccharide-Induced Inflammatory Responses in Broilers. J. Anim. Sci. Biotechnol. 2021, 12, 25. [Google Scholar] [CrossRef] [PubMed]
- Bäuerl, C.; Coll-Marqués, J.M.; Tarazona-González, C.; Pérez-Martínez, G. Lactobacillus casei Extracellular Vesicles Stimulate EGFR Pathway Likely Due to the Presence of Proteins P40 and P75 Bound to Their Surface. Sci. Rep. 2020, 10, 19237. [Google Scholar] [CrossRef] [PubMed]
- Rubio, A.P.D.; Martínez, J.H.; Casillas, D.C.M.; Leskow, F.C.; Piuri, M.; Pérez, O.E. Lactobacillus casei BL23 Produces Microvesicles Carrying Proteins That Have Been Associated with Its Probiotic Effect. Front. Microbiol. 2017, 8, 1783. [Google Scholar] [CrossRef]
- Vargoorani, M.E.; Modarressi, M.H.; Vaziri, F.; Motevaseli, E.; Siadat, S.D. Stimulatory Effects of Lactobacillus casei Derived Extracellular Vesicles on Toll-Like Receptor 9 Gene Expression and Cytokine Profile in Human Intestinal Epithelial Cells. J. Diabetes Metab. Disord. 2020, 19, 223–231. [Google Scholar] [CrossRef]
- Dean, S.N.; Leary, D.H.; Sullivan, C.J.; Oh, E.; Walper, S.A. Isolation and Characterization of Lactobacillus-Derived Membrane Vesicles. Sci. Rep. 2019, 9, 877. [Google Scholar] [CrossRef]
- Grande, R.; Celia, C.; Mincione, G.; Stringaro, A.; Di Marzio, L.; Colone, M.; Di Marcantonio, M.C.; Savino, L.; Puca, V.; Santoliquido, R.; et al. Detection and Physicochemical Characterization of Membrane Vesicles (MVs) of Lactobacillus reuteri DSM 17938. Front. Microbiol. 2017, 8, 1040. [Google Scholar] [CrossRef]
- Shao, H.; Im, H.; Castro, C.M.; Breakefield, X.; Weissleder, R.; Lee, H. New Technologies for Analysis of Extracellular Vesicles. Chem. Rev. 2018, 118, 1917–1950. [Google Scholar] [CrossRef]
- Hilton, S.H.; White, I.M. Advances in the Analysis of Single Extracellular Vesicles: A Critical Review. Sens. Actuators Rep. 2021, 3, 100052. [Google Scholar] [CrossRef]
- Alpdundar Bulut, E.; Bayyurt Kocabas, B.; Yazar, V.; Aykut, G.; Guler, U.; Salih, B.; Surucu Yilmaz, N.; Ayanoglu, I.C.; Polat, M.M.; Akcali, K.C.; et al. Human Gut Commensal Membrane Vesicles Modulate Inflammation by Generating M2-like Macrophages and Myeloid-Derived Suppressor Cells. J. Immunol. 2020, 205, 2707–2718. [Google Scholar] [CrossRef] [PubMed]
- Campoccia, D.; Montanaro, L.; Arciola, C.R. Extracellular DNA (eDNA). A Major Ubiquitous Element of the Bacterial Biofilm Architecture. Int. J. Mol. Sci. 2021, 22, 9100. [Google Scholar] [CrossRef] [PubMed]
- Okshevsky, M.; Meyer, R.L. The Role of Extracellular DNA in the Establishment, Maintenance and Perpetuation of Bacterial Biofilms. Crit. Rev. Microbiol. 2015, 41, 341–352. [Google Scholar] [CrossRef] [PubMed]
- Puca, V.; Ercolino, E.; Celia, C.; Bologna, G.; Di Marzio, L.; Mincione, G.; Marchisio, M.; Miscia, S.; Muraro, R.; Lanuti, P.; et al. Detection and Quantification of eDNA-Associated Bacterial Membrane Vesicles by Flow Cytometry. Int. J. Mol. Sci. 2019, 20, 5307. [Google Scholar] [CrossRef]
- Zingl, F.G.; Leitner, D.R.; Thapa, H.B.; Schild, S. Outer Membrane Vesicles as Versatile Tools for Therapeutic Approaches. microLife 2021, 2, uqab006. [Google Scholar] [CrossRef]
- Caruana, J.C.; Walper, S.A. Bacterial Membrane Vesicles as Mediators of Microbe—Microbe and Microbe—Host Community Interactions. Front. Microbiol. 2020, 11, 432. [Google Scholar] [CrossRef]
- Díaz-Garrido, N.; Badia, J.; Baldomà, L. Microbiota-Derived Extracellular Vesicles in Interkingdom Communication in the Gut. J. Extracell. Vesicles 2021, 10, e12161. [Google Scholar] [CrossRef]
- Kim, H.; Kim, M.; Myoung, K.; Kim, W.; Ko, J.; Kim, K.P.; Cho, E.G. Comparative Lipidomic Analysis of Extracellular Vesicles Derived from Lactobacillus plantarum APsulloc 331261 Living in Green Tea Leaves Using Liquid Chromatography-Mass Spectrometry. Int. J. Mol. Sci. 2020, 21, 8076. [Google Scholar] [CrossRef]
- Kim, W.; Lee, E.J.; Bae, I.H.; Myoung, K.; Kim, S.T.; Park, P.J.; Lee, K.H.; Pham, A.V.Q.; Ko, J.; Oh, S.H.; et al. Lactobacillus plantarum-Derived Extracellular Vesicles Induce Anti-Inflammatory M2 Macrophage Polarization In Vitro. J. Extracell. Vesicles 2020, 9, 1793514. [Google Scholar] [CrossRef]
- Pérez-Cruz, C.; Cañas, M.A.; Giménez, R.; Badia, J.; Mercade, E.; Baldomà, L.; Aguilera, L. Membrane Vesicles Released by a Hypervesiculating Escherichia coli Nissle 1917 tolR Mutant Are Highly Heterogeneous and Show Reduced Capacity for Epithelial Cell Interaction and Entry. PLoS ONE 2016, 11, e0169186. [Google Scholar] [CrossRef]
- Aguilera, L.; Toloza, L.; Giménez, R.; Odena, A.; Oliveira, E.; Aguilar, J.; Badia, J.; Baldomà, L. Proteomic Analysis of Outer Membrane Vesicles from the Probiotic Strain Escherichia coli Nissle 1917. Proteomics 2014, 14, 222–229. [Google Scholar] [CrossRef]
- Ghaderi, F.; Sotoodehnejadnematalahi, F.; Hajebrahimi, Z.; Fateh, A.; Siadat, S.D. Effects of Active, Inactive, and Derivatives of Akkermansia muciniphila on The Expression of The Endocannabinoid System and PPARs Genes. Sci. Rep. 2022, 12, 10031. [Google Scholar] [CrossRef]
- Bajic, S.S.; Cañas, M.A.; Tolinacki, M.; Badia, J.; Sánchez, B.; Golic, N.; Margolles, A.; Baldomá, L.; Ruas-Madiedo, P. Proteomic Profile of Extracellular Vesicles Released by Lactiplantibacillus plantarum BGAN8 and Their Internalization by Non-Polarized HT29 Cell Line. Sci. Rep. 2020, 10, 21829. [Google Scholar] [CrossRef]
- Kim, M.H.; Choi, S.J.; Choi, H., II; Choi, J.P.; Park, H.K.; Kim, E.K.; Kim, M.J.; Moon, B.S.; Min, T.K.; Rho, M.; et al. Lactobacillus plantarum-Derived Extracellular Vesicles Protect Atopic Dermatitis Induced by Staphylococcus aureus-Derived Extracellular Vesicles. Allergy. Asthma Immunol. Res. 2018, 10, 516–532. [Google Scholar] [CrossRef]
- Behzadi, E.; Mahmoodzadeh Hosseini, H.; Imani Fooladi, A.A. The Inhibitory Impacts of Lactobacillus rhamnosus GG-Derived Extracellular Vesicles on the Growth of Hepatic Cancer Cells. Microb. Pathog. 2017, 110, 1–6. [Google Scholar] [CrossRef]
- Keyhani, G.; Hosseini, H.M.; Salimi, A. Effect of Extracellular Vesicles of Lactobacillus rhamnosus GG on The Expression of CEA Gene and Protein Released by Colorectal Cancer Cells. Iran. J. Microbiol. 2022, 14, 90–96. [Google Scholar] [CrossRef]
- Choi, J.H.; Moon, C.M.; Shin, T.S.; Kim, E.K.; McDowell, A.; Jo, M.K.; Joo, Y.H.; Kim, S.E.; Jung, H.K.; Shim, K.N.; et al. Lactobacillus paracasei-Derived Extracellular Vesicles Attenuate the Intestinal Inflammatory Response by Augmenting the Endoplasmic Reticulum Stress Pathway. Exp. Mol. Med. 2020, 52, 423–437. [Google Scholar] [CrossRef]
- Shiraishi, T.; Yokota, S.; Sato, Y.; Ito, T.; Fukiya, S.; Yamamoto, S.; Sato, T.; Yokota, A. Lipoteichoic Acids are Embedded in Cell Walls during Logarithmic Phase, but Exposed on Membrane Vesicles in Lactobacillus gasseri JCM 1131 T. Benef. Microbes 2018, 9, 653–662. [Google Scholar] [CrossRef]
- Seo, M.K.; Park, E.J.; Ko, S.Y.; Choi, E.W.; Kim, S. Therapeutic Effects of Kefir Grain Lactobacillus-Derived Extracellular Vesicles in Mice with 2,4,6-Trinitrobenzene Sulfonic Acid-Induced Inflammatory Bowel Disease. J. Dairy Sci. 2018, 101, 8662–8671. [Google Scholar] [CrossRef] [Green Version]
- Yamasaki-Yashiki, S.; Miyoshi, Y.; Nakayama, T.; Kunisawa, J.; Katakura, Y. IgA-Enhancing Effects of Membrane Vesicles Derived from Lactobacillus sakei subsp. sakei NBRC15893. Biosci. Microbiota, Food Health 2019, 38, 29. [Google Scholar]
- Liu, Y.; Tempelaars, M.H.; Boeren, S.; Alexeeva, S.; Smid, E.J.; Abee, T. Extracellular Vesicle Formation in Lactococcus lactis is Stimulated by Prophage-Encoded Holin–Lysin System. Microb. Biotechnol. 2022, 15, 1281–1295. [Google Scholar] [CrossRef] [PubMed]
- Fábrega, M.J.; Aguilera, L.; Giménez, R.; Varela, E.; Cañas, M.A.; Antolín, M.; Badía, J.; Baldomà, L. Activation of Immune and Defense Responses in the Intestinal Mucosa by Outer Membrane Vesicles of Commensal and Probiotic Escherichia coli Strains. Front. Microbiol. 2016, 7, 705. [Google Scholar] [CrossRef]
- Nishiyama, K.; Takaki, T.; Sugiyama, M.; Fukuda, I.; Aiso, M.; Mukai, T.; Odamaki, T.; Xiao, J.Z.; Osawa, R.; Okada, N. Extracellular Vesicles Produced by Bifidobacterium longum Export Mucin-Binding Proteins. Appl. Environ. Microbiol. 2020, 86, e01464-20. [Google Scholar] [CrossRef] [PubMed]
- Fábrega, M.J.; Rodríguez-Nogales, A.; Garrido-Mesa, J.; Algieri, F.; Badía, J.; Giménez, R.; Gálvez, J.; Baldomà, L. Intestinal Anti-inflammatory Effects of Outer Membrane Vesicles from Escherichia coli Nissle 1917 in DSS-Experimental Colitis in Mice. Front. Microbiol. 2017, 8, 1274. [Google Scholar] [CrossRef] [PubMed]
- Hirayama, S.; Nakao, R. Glycine Significantly Enhances Bacterial Membrane Vesicle Production: A Powerful Approach for Isolation of LPS-Reduced Membrane Vesicles of Probiotic Escherichia coli. Microb. Biotechnol. 2020, 13, 1162–1178. [Google Scholar] [CrossRef]
- Ashrafian, F.; Shahriary, A.; Behrouzi, A.; Moradi, H.R.; Keshavarz Azizi Raftar, S.; Lari, A.; Hadifar, S.; Yaghoubfar, R.; Ahmadi Badi, S.; Khatami, S.; et al. Akkermansia muciniphila-Derived Extracellular Vesicles as a Mucosal Delivery Vector for Amelioration of Obesity in Mice. Front. Microbiol. 2019, 10, 2155. [Google Scholar] [CrossRef]
- Ashrafian, F.; Behrouzi, A.; Shahriary, A.; Badi, S.A.; Davari, M.; Khatami, S.; Jamnani, F.R.; Fateh, A.; Vaziri, F.; Siadat, S.D. Comparative Study of Effect of Akkermansia muciniphila and Its Extracellular Vesicles on Toll-Like Receptors and Tight Junction. Gastroenterol. Hepatol. Bed Bench 2019, 12, 168. [Google Scholar]
- Ashrafian, F.; Keshavarz Azizi Raftar, S.; Lari, A.; Shahryari, A.; Abdollahiyan, S.; Moradi, H.R.; Masoumi, M.; Davari, M.; khatami, S.; Omrani, M.D.; et al. Extracellular Vesicles and Pasteurized Cells Derived from Akkermansia muciniphila Protect against High-Fat Induced Obesity in Mice. Microb. Cell Fact. 2021, 20, 219. [Google Scholar] [CrossRef]
- Keshavarz Azizi Raftar, S.; Ashrafian, F.; Yadegar, A.; Lari, A.; Moradi, H.R.; Shahriary, A.; Azimirad, M.; Alavifard, H.; Mohsenifar, Z.; Davari, M.; et al. The Protective Effects of Live and Pasteurized Akkermansia muciniphila and Its Extracellular Vesicles against HFD/CCl4-Induced Liver Injury. Microbiol. Spectr. 2021, 9, e0048421. [Google Scholar] [CrossRef]
- Yaghoubfar, R.; Behrouzi, A.; Zare Banadkoki, E.; Ashrafian, F.; Lari, A.; Vaziri, F.; Nojoumi, S.A.; Fateh, A.; Khatami, S.; Siadat, S.D. Effect of Akkermansia muciniphila, Faecalibacterium prausnitzii, and Their Extracellular Vesicles on the Serotonin System in Intestinal Epithelial Cells. Probiotics Antimicrob. Proteins 2021, 13, 1546–1556. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.; Dauros-Singorenko, P.; Whitcombe, A.; Payne, L.; Blenkiron, C.; Phillips, A.; Swift, S. Analysis of the Escherichia coli Extracellular Vesicle Proteome Identifies Markers of Purity and Culture Conditions. J. Extracell. Vesicles 2019, 8, 1632099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, B.H.; Wu, S.C.; Shen, T.L.; Hsu, Y.Y.; Chen, C.H.; Hsu, W.H. The Applications of Lactobacillus plantarum-Derived Extracellular Vesicles as a Novel Natural Antibacterial Agent for Improving Quality and Safety in Tuna Fish. Food Chem. 2021, 340, 128104. [Google Scholar] [CrossRef]
- Ñahui Palomino, R.A.; Vanpouille, C.; Laghi, L.; Parolin, C.; Melikov, K.; Backlund, P.; Vitali, B.; Margolis, L. Extracellular Vesicles from Symbiotic Vaginal Lactobacilli Inhibit HIV-1 Infection of Human Tissues. Nat. Commun. 2019, 10, 5656. [Google Scholar] [CrossRef] [PubMed]
- West, C.L.; Stanisz, A.M.; Mao, Y.K.; Champagne-Jorgensen, K.; Bienenstock, J.; Kunze, W.A. Microvesicles from Lactobacillus reuteri (DSM-17938) Completely Reproduce Modulation of Gut Motility by Bacteria in Mice. PLoS ONE 2020, 15, e0225481. [Google Scholar] [CrossRef]
- Chen, C.Y.; Rao, S.S.; Yue, T.; Tan, Y.J.; Yin, H.; Chen, L.J.; Luo, M.J.; Wang, Z.; Wang, Y.Y.; Hong, C.G.; et al. Glucocorticoid-Induced Loss of Beneficial Gut Bacterial Extracellular Vesicles Is Associated with The Pathogenesis of Osteonecrosis. Sci. Adv. 2022, 8, eabg8335. [Google Scholar] [CrossRef]
- Yoon, Y.C.; Ahn, B.H.; Min, J.W.; Lee, K.R.; Park, S.H.; Kang, H.C. Stimulatory Effects of Extracellular Vesicles Derived from Leuconostoc holzapfelii That Exists in Human Scalp on Hair Growth in Human Follicle Dermal Papilla Cells. Curr. Issues Mol. Biol. 2022, 44, 845–866. [Google Scholar] [CrossRef]
- de Rodovalho, V.R.; da Luz, B.S.R.; Rabah, H.; do Carmo, F.L.R.; Folador, E.L.; Nicolas, A.; Jardin, J.; Briard-Bion, V.; Blottière, H.; Lapaque, N.; et al. Extracellular Vesicles Produced by the Probiotic Propionibacterium freudenreichii CIRM-BIA 129 Mitigate Inflammation by Modulating the NF-κB Pathway. Front. Microbiol. 2020, 11, 1544. [Google Scholar] [CrossRef]
- de Rezende Rodovalho, V.; da Luz, B.S.R.; Nicolas, A.; do Carmo, F.L.R.; Jardin, J.; Briard-Bion, V.; Jan, G.; Loir, Y.L.; de Carvalho Azevedo, V.A.; Guédon, E. Environmental Conditions Modulate the Protein Content and Immunomodulatory Activity of Extracellular Vesicles Produced by the Probiotic Propionibacterium freudenreichii. Appl. Environ. Microbiol. 2020, 87, e02263-20. [Google Scholar]
- Rubio, A.P.D.; Martínez, J.; Palavecino, M.; Fuentes, F.; López, C.M.S.; Marcilla, A.; Pérez, O.E.; Piuri, M. Transcytosis of Bacillus subtilis Extracellular Vesicles through an In Vitro Intestinal Epithelial Cell Model. Sci. Rep. 2020, 10, 3120. [Google Scholar] [CrossRef]
- Ma, L.; Shen, Q.; Lyu, W.; Lv, L.; Wang, W.; Yu, M.; Yang, H.; Tao, S.; Xiao, Y. Clostridium butyricum and Its Derived Extracellular Vesicles Modulate Gut Homeostasis and Ameliorate Acute Experimental Colitis. Microbiol. Spectr. 2022, 10, e0136822. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.; Yang, C.; Liu, L.; Mai, G.; Li, H.; Wu, L.; Jin, M.; Chen, Y. Commensal Bacteria-Derived Extracellular Vesicles Suppress Ulcerative Colitis through Regulating The Macrophages Polarization and Remodeling The Gut Microbiota. Microb. Cell Fact. 2022, 21, 88. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, T.; Koch, M.; Fuhrmann, G.; Kuhn, T.; Fuhrmann, G.; Koch, M. Probiomimetics—Novel Lactobacillus-Mimicking Microparticles Show Anti-Inflammatory and Barrier-Protecting Effects in Gastrointestinal Models. Small 2020, 16, 2003158. [Google Scholar] [CrossRef] [PubMed]
- Morishita, M.; Sagayama, R.; Yamawaki, Y.; Yamaguchi, M.; Katsumi, H.; Yamamoto, A. Activation of Host Immune Cells by Probiotic-Derived Extracellular Vesicles via TLR2-Mediated Signaling Pathways. Biol. Pharm. Bull. 2022, 45, 354–359. [Google Scholar] [CrossRef]
- Morishita, M.; Horita, M.; Higuchi, A.; Marui, M.; Katsumi, H.; Yamamoto, A. Characterizing Different Probiotic-Derived Extracellular Vesicles as a Novel Adjuvant for Immunotherapy. Mol. Pharm. 2021, 18, 1080–1092. [Google Scholar] [CrossRef]
- Yang, Z.; Gao, Z.; Yang, Z.; Zhang, Y.; Chen, H.; Yang, X.; Fang, X.; Zhu, Y.; Zhang, J.; Ouyang, F.; et al. Lactobacillus plantarum-Derived Extracellular Vesicles Protect against Ischemic Brain Injury via The MicroRNA-101a-3p/c-Fos/TGF-β Axis. Pharmacol. Res. 2022, 182, 106332. [Google Scholar] [CrossRef]
- Jo, C.S.; Myung, C.H.; Yoon, Y.C.; Ahn, B.H.; Min, J.W.; Seo, W.S.; Lee, D.H.; Kang, H.C.; Heo, Y.H.; Choi, H.; et al. The Effect of Lactobacillus plantarum Extracellular Vesicles from Korean Women in Their 20s on Skin Aging. Curr. Issues Mol. Biol. 2022, 44, 526–540. [Google Scholar] [CrossRef]
- Champagne-Jorgensen, K.; Mian, M.F.; McVey Neufeld, K.A.; Stanisz, A.M.; Bienenstock, J. Membrane Vesicles of Lacticaseibacillus rhamnosus JB-1 Contain Immunomodulatory Lipoteichoic Acid and Are Endocytosed by Intestinal Epithelial Cells. Sci. Rep. 2021, 11, 13756. [Google Scholar] [CrossRef]
- Champagne-Jorgensen, K.; Jose, T.A.; Stanisz, A.M.; Mian, M.F.; Hynes, A.P.; Bienenstock, J. Bacterial Membrane Vesicles and Phages in Blood After Consumption of Lacticaseibacillus rhamnosus JB-1. Gut Microbes 2021, 13, e1993583. [Google Scholar] [CrossRef]
- Shi, Y.; Meng, L.; Zhang, C.; Zhang, F.; Fang, Y. Extracellular Vesicles of Lacticaseibacillus paracasei PC-H1 Induce Colorectal Cancer Cells Apoptosis via PDK1/AKT/Bcl-2 Signaling Pathway. Microbiol. Res. 2021, 255, 126921. [Google Scholar] [CrossRef]
- Chelakkot, C.; Choi, Y.; Kim, D.K.; Park, H.T.; Ghim, J.; Kwon, Y.; Jeon, J.; Kim, M.S.; Jee, Y.K.; Gho, Y.S.; et al. Akkermansia muciniphila-Derived Extracellular Vesicles Influence Gut Permeability through the Regulation of Tight Junctions. Exp. Mol. Med. 2018, 50, e450. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.W.; Xia, K.; Liu, Y.W.; Liu, J.H.; Rao, S.S.; Hu, X.K.; Chen, C.Y.; Xu, R.; Wang, Z.X.; Xie, H. Extracellular Vesicles from Akkermansia muciniphila Elicit Antitumor Immunity Against Prostate Cancer via Modulation of CD8 + T Cells and Macrophages. Int. J. Nanomed. 2021, 16, 2949–2963. [Google Scholar] [CrossRef] [PubMed]
- Hao, H.; Zhang, X.; Tong, L.; Liu, Q.; Liang, X.; Bu, Y.; Gong, P.; Liu, T.; Zhang, L.; Xia, Y.; et al. Effect of Extracellular Vesicles Derived From Lactobacillus plantarum Q7 on Gut Microbiota and Ulcerative Colitis in Mice. Front. Immunol. 2021, 12, 5167. [Google Scholar] [CrossRef]
- Tong, L.; Zhang, X.; Hao, H.; Liu, Q.; Zhou, Z.; Liang, X.; Liu, T.; Gong, P.; Zhang, L.; Zhai, Z.; et al. Lactobacillus rhamnosus GG Derived Extracellular Vesicles Modulate Gut Microbiota and Attenuate Inflammatory in DSS-Induced Colitis Mice. Nutrients 2021, 13, 3319. [Google Scholar] [CrossRef]
- Lee, D.H.; Park, H.K.; Lee, H.R.; Sohn, H.; Sim, S.; Park, H.J.; Shin, Y.S.; Kim, Y.K.; Choi, Y.; Park, H.S. Immunoregulatory Effects of Lactococcus lactis-Derived Extracellular Vesicles in Allergic Asthma. Clin. Transl. Allergy 2022, 12, e12138. [Google Scholar] [CrossRef] [PubMed]
- Nagakubo, T.; Nomura, N.; Toyofuku, M. Cracking Open Bacterial Membrane Vesicles. Front. Microbiol. 2020, 10, 3026. [Google Scholar] [CrossRef]
- Lee, J.; Kim, O.Y.; Gho, Y.S. Proteomic Profiling of Gram-negative Bacterial Outer Membrane Vesicles: Current Perspectives. PROTEOMICS—Clin. Appl. 2016, 10, 897–909. [Google Scholar] [CrossRef]
- Pedersen, S.K.; Harry, J.L.; Sebastian, L.; Baker, J.; Traini, M.D.; McCarthy, J.T.; Manoharan, A.; Wilkins, M.R.; Gooley, A.A.; Righetti, P.G.; et al. Unseen Proteome: Mining Below the Tip of the Iceberg To Find Low Abundance and Membrane Proteins. J. Proteome Res. 2003, 2, 303–311. [Google Scholar] [CrossRef]
- Ahmed, N.; Rice, G.E. Strategies for Revealing Lower Abundance Proteins in Two-Dimensional Protein Maps. J. Chromatogr. B. Analyt. Technol. Biomed. Life Sci. 2005, 815, 39–50. [Google Scholar] [CrossRef]
- Klimentová, J.; Stulík, J. Methods of Isolation and Purification of Outer Membrane Vesicles from Gram-Negative Bacteria. Microbiol. Res. 2015, 170, 1–9. [Google Scholar] [CrossRef]
- Graf, A.C.; Leonard, A.; Schäuble, M.; Rieckmann, L.M.; Hoyer, J.; Maass, S.; Lalk, M.; Becher, D.; Pané-Farré, J.; Riedel, K. Virulence Factors Produced by Staphylococcus aureus Biofilms Have a Moonlighting Function Contributing to Biofilm Integrity. Mol. Cell. Proteomics 2019, 18, 1053. [Google Scholar] [CrossRef] [PubMed]
- Kesavan, D.; Vasudevan, A.; Wu, L.; Chen, J.; Su, Z.; Wang, S.; Xu, H. Integrative Analysis of Outer Membrane Vesicles Proteomics and Whole-Cell Transcriptome Analysis of Eravacycline Induced Acinetobacter baumannii Strains. BMC Microbiol. 2020, 20, 31. [Google Scholar] [CrossRef]
- Sung, K.; Chon, J.; Kweon, O.; Nho, S.; Kim, S.; Park, M.; Paredes, A.; Lim, J.H.; Khan, S.A.; Phillips, K.S.; et al. Dynamic Adaptive Response of Pseudomonas aeruginosa to Clindamycin/Rifampicin-Impregnated Catheters. Antibiotics 2021, 10, 752. [Google Scholar] [CrossRef]
- Rolhion, N.; Barnich, N.; Bringer, M.A.; Glasser, A.L.; Ranc, J.; Hébuterne, X.; Hofman, P.; Darfeuille-Michaud, A. Abnormally Expressed ER Stress Response Chaperone Gp96 in CD Favours Adherent-Invasive Escherichia coli Invasion. Gut 2010, 59, 1355–1362. [Google Scholar] [CrossRef]
- Taheri, N.; Mahmud, A.K.M.F.; Sandblad, L.; Fällman, M.; Wai, S.N.; Fahlgren, A. Campylobacter jejuni Bile Exposure Influences Outer Membrane Vesicles Protein Content and Bacterial Interaction with Epithelial Cells. Sci. Rep. 2018, 8, 16996. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.I.; Kim, S.; Kim, E.; Hwang, S.Y.; Yoon, H. Secretion of Salmonella Pathogenicity Island 1-Encoded Type III Secretion System Effectors by Outer Membrane Vesicles in Salmonella enterica Serovar Typhimurium. Front. Microbiol. 2018, 9, 2810. [Google Scholar] [CrossRef]
- Avila-Calderón, E.D.; del Ruiz-Palma, M.S.; Aguilera-Arreola, M.G.; Velázquez-Guadarrama, N.; Ruiz, E.A.; Gomez-Lunar, Z.; Witonsky, S.; Contreras-Rodríguez, A. Outer Membrane Vesicles of Gram-Negative Bacteria: An Outlook on Biogenesis. Front. Microbiol. 2021, 12, 345. [Google Scholar] [CrossRef]
- Clarke, A.J. The “Hole” Story of Predatory Outer-Membrane Vesicles. Can. J. Microbiol. 2018, 64, 589–599. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.-P.; Nie, C.; Guan, W.-Y.; Qiao, L.-D.; Lu, L.; Cao, S.-J. Regulation of Probiotics on Metabolism of Dietary Protein in Intestine. Curr. Protein Pept. Sci. 2019, 21, 766–771. [Google Scholar] [CrossRef]
- Díez-Sainz, E.; Lorente-Cebrián, S.; Aranaz, P.; Riezu-Boj, J.I.; Martínez, J.A.; Milagro, F.I. Potential Mechanisms Linking Food-Derived MicroRNAs, Gut Microbiota and Intestinal Barrier Functions in the Context of Nutrition and Human Health. Front. Nutr. 2021, 8, 85. [Google Scholar] [CrossRef]
- Villard, A.; Boursier, J.; Andriantsitohaina, R. Microbiota-Derived Extracellular Vesicles and Metabolic Syndrome. Acta Physiol. 2021, 231, e13600. [Google Scholar] [CrossRef]
- McMillan, H.M.; Kuehn, M.J. The Extracellular Vesicle Generation Paradox: A Bacterial Point of View. EMBO J. 2021, 40, e108174. [Google Scholar] [CrossRef]
- Bitto, N.J.; Zavan, L.; Johnston, E.L.; Stinear, T.P.; Hill, A.F.; Kaparakis-Liaskos, M. Considerations for the Analysis of Bacterial Membrane Vesicles: Methods of Vesicle Production and Quantification Can Influence Biological and Experimental Outcomes. Microbiol. Spectr. 2021, 9, e01273-21. [Google Scholar] [CrossRef]
- Papadimitriou, K.; Zoumpopoulou, G.; Foligné, B.; Alexandraki, V.; Kazou, M.; Pot, B.; Tsakalidou, E. Discovering Probiotic Microorganisms: In Vitro, In Vivo, Genetic And Omics Approaches. Front. Microbiol. 2015, 6, 58. [Google Scholar] [CrossRef] [PubMed]
- Sagheddu, V.; Guidesi, E.; Galletti, S.; Elli, M. Selection and Characterization Criteria of Probiotics Intended for Human Use from the Past to the Future. Food Sci. Nutr. Stud. 2019, 3, p73. [Google Scholar] [CrossRef]
- Suzuki, T. Regulation of Intestinal Epithelial Permeability By Tight Junctions. Cell. Mol. Life Sci. 2013, 70, 631–659. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.R. Intestinal Mucosal Barrier Function in Health and Disease. Nat. Rev. Immunol. 2009, 9, 799–809. [Google Scholar] [CrossRef]
- Kang, E.A.; Choi, H.I.; Hong, S.W.; Kang, S.; Jegal, H.Y.; Choi, E.W.; Park, B.S.; Kim, J.S. Extracellular Vesicles Derived from Kefir Grain Lactobacillus Ameliorate Intestinal Inflammation via Regulation of Proinflammatory Pathway and Tight Junction Integrity. Biomedicines 2020, 8, 522. [Google Scholar] [CrossRef]
- Xu, X.; Lin, S.; Yang, Y.; Gong, X.; Tong, J.; Li, K.; Li, Y. Histological and Ultrastructural Changes of the Colon in Dextran Sodium Sulfate-Induced Mouse Colitis. Exp. Ther. Med. 2020, 20, 1987. [Google Scholar] [CrossRef]
- Hu, Y.; Ye, Z.; Wu, M.; She, Y.; Li, L.; Xu, Y.; Qin, K.; Hu, Z.; Yang, M.; Lu, F.; et al. The Communication Between Intestinal Microbiota and Ulcerative Colitis: An Exploration of Pathogenesis, Animal Models, and Potential Therapeutic Strategies. Front. Med. 2021, 8, 766126. [Google Scholar] [CrossRef]
- Petersson, J.; Schreiber, O.; Hansson, G.C.; Gendler, S.J.; Velcich, A.; Lundberg, J.O.; Roos, S.; Holm, L.; Phillipson, M. Importance and Regulation of the Colonic Mucus Barrier In A Mouse Model of Colitis. Am. J. Physiol. 2011, 300, G327. [Google Scholar] [CrossRef] [PubMed]
- Tomasi, M.; Caproni, E.; Benedet, M.; Zanella, I.; Giorgetta, S.; Dalsass, M.; König, E.; Gagliardi, A.; Fantappiè, L.; Berti, A.; et al. Outer Membrane Vesicles From The Gut Microbiome Contribute to Tumor Immunity by Eliciting Cross-Reactive T Cells. Front. Oncol. 2022, 12, 912639. [Google Scholar] [CrossRef]
- Hiam-Galvez, K.J.; Allen, B.M.; Spitzer, M.H. Systemic Immunity in Cancer. Nat. Rev. Cancer 2021, 21, 345–359. [Google Scholar] [CrossRef] [PubMed]
- Fessler, J.; Matson, V.; Gajewski, T.F. Exploring the Emerging Role of the Microbiome in Cancer Immunotherapy. J. Immunother. Cancer 2019, 7, 108. [Google Scholar] [CrossRef] [PubMed]
- Miyoshi, Y.; Saika, A.; Nagatake, T.; Matsunaga, A.; Kunisawa, J.; Katakura, Y.; Yamasaki-Yashiki, S. Mechanisms Underlying Enhanced IgA Production in Peyer’s Patch Cells by Membrane Vesicles Derived from Lactobacillus sakei. Biosci. Biotechnol. Biochem. 2021, 85, 1536–1545. [Google Scholar] [CrossRef] [PubMed]
- Geuking, M.B.; Köller, Y.; Rupp, S.; McCoy, K.D. The Interplay Between the Gut Microbiota and the Immune System. Gut Microbes 2014, 5, 411. [Google Scholar] [CrossRef]
- Kamada, N.; Seo, S.U.; Chen, G.Y.; Núñez, G. Role of The Gut Microbiota in Immunity and Inflammatory Disease. Nat. Rev. Immunol. 2013, 13, 321–335. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, P.; Zhang, X. Probiotics Regulate Gut Microbiota: An Effective Method to Improve Immunity. Molecules 2021, 26, 6076. [Google Scholar] [CrossRef]
- Choi, J.; Kim, Y.K.; Han, P.L. Extracellular Vesicles Derived from Lactobacillus plantarum Increase BDNF Expression in Cultured Hippocampal Neurons and Produce Antidepressant-Like Effects in Mice. Exp. Neurobiol. 2019, 28, 158–171. [Google Scholar] [CrossRef]
- Choi, J.; Kwon, H.; Kim, Y.K.; Han, P.L. Extracellular Vesicles from Gram-positive and Gram-negative Probiotics Remediate Stress-Induced Depressive Behavior in Mice. Mol. Neurobiol. 2022, 59, 2715–2728. [Google Scholar] [CrossRef]
- Alvarez, C.S.; Giménez, R.; Cañas, M.A.; Vera, R.; Díaz-Garrido, N.; Badia, J.; Baldomà, L. Extracellular Vesicles and Soluble Factors Secreted by Escherichia coli Nissle 1917 and ECOR63 Protect against Enteropathogenic E. coli-Induced Intestinal Epithelial Barrier Dysfunction. BMC Microbiol. 2019, 19, 166. [Google Scholar] [CrossRef]
- Cañas, M.A.; Fábrega, M.J.; Giménez, R.; Badia, J.; Baldomà, L. Outer Membrane Vesicles From Probiotic and Commensal Escherichia coli Activate NOD1-Mediated Immune Responses in Intestinal Epithelial Cells. Front. Microbiol. 2018, 9, 498. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Garrido, N.; Bonnin, S.; Riera, M.; Gíménez, R.; Badia, J.; Baldomà, L. Transcriptomic microRNA Profiling of Dendritic Cells in Response to Gut Microbiota-Secreted Vesicles. Cells 2020, 9, 1534. [Google Scholar] [CrossRef]
- Raftar, S.K.A.; Ashrafian, F.; Abdollahiyan, S.; Yadegar, A.; Moradi, H.R.; Masoumi, M.; Vaziri, F.; Moshiri, A.; Siadat, S.D.; Zali, M.R. The Anti-Inflammatory Effects of Akkermansia muciniphila and Its Derivates in HFD/CCL4-Induced Murine Model of Liver Injury. Sci. Rep. 2022, 12, 2453. [Google Scholar] [CrossRef] [PubMed]
- Mata Forsberg, M.; Björkander, S.; Pang, Y.; Lundqvist, L.; Ndi, M.; Ott, M.; Escribá, I.B.; Jaeger, M.C.; Roos, S.; Sverremark-Ekström, E. Extracellular Membrane Vesicles from Lactobacilli Dampen IFN-γ Responses in a Monocyte-Dependent Manner. Sci. Rep. 2019, 9, 17109. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, L.D.; Harrison, N.A.; da Silva, D.R.; Mathews, C.E.; Gonzalez, C.F.; Lorca, G.L. Nanovesicles From Lactobacillus johnsonii N6.2 Reduce Apoptosis in Human Beta Cells by Promoting AHR Translocation and IL10 Secretion. Front. Immunol. 2022, 13, 899413. [Google Scholar] [CrossRef]
- Kim, J.H.; Jeun, E.J.; Hong, C.P.; Kim, S.H.; Jang, M.S.; Lee, E.J.; Moon, S.J.; Yun, C.H.; Im, S.H.; Jeong, S.G.; et al. Extracellular Vesicle-Derived Protein from Bifidobacterium longum Alleviates Food Allergy through Mast Cell Suppression. J. Allergy Clin. Immunol. 2016, 137, 507–516.e8. [Google Scholar] [CrossRef]
- López, P.; González-Rodríguez, I.; Sánchez, B.; Gueimonde, M.; Margolles, A.; Suárez, A. Treg-Inducing Membrane Vesicles from Bifidobacterium bifidum LMG13195 as Potential Adjuvants in Immunotherapy. Vaccine 2012, 30, 825–829. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Han, S.-Y.; Lee, J.; Kim, N.-R.; Lee, B.R.; Kim, H.; Kwon, M.; Ahn, K.; Noh, Y.; Kim, S.J.; et al. Bifidobacterium longum and Galactooligosaccharide Improve Skin Barrier Dysfunction and Atopic Dermatitis-Like Skin. Allergy. Asthma Immunol. Res. 2022, 14, 564. [Google Scholar] [CrossRef]
- An, J.J.; Ha, E.M. Extracellular Vesicles Derived from Lactobacillus plantarum Restore Chemosensitivity through the PDK2-Mediated Glucose Metabolic Pathway in 5-FU-Resistant Colorectal Cancer Cells. J. Microbiol. 2022, 60, 735–745. [Google Scholar] [CrossRef]
- Pang, Y.; Ermann Lundberg, L.; Mata Forsberg, M.; Ahl, D.; Bysell, H.; Pallin, A.; Sverremark-Ekström, E.; Karlsson, R.; Jonsson, H.; Roos, S. Extracellular Membrane Vesicles from Limosilactobacillus reuteri Strengthen the Intestinal Epithelial Integrity, Modulate Cytokine Responses and Antagonize Activation of TRPV1. Front. Microbiol. 2022, 13, 1032202. [Google Scholar] [CrossRef]
- Kurata, A.; Kiyohara, S.; Imai, T.; Yamasaki-Yashiki, S.; Zaima, N.; Moriyama, T.; Kishimoto, N.; Uegaki, K. Characterization of Extracellular Vesicles from Lactiplantibacillus plantarum. Sci. Rep. 2022, 12, 13330. [Google Scholar] [CrossRef] [PubMed]
- Nakao, R.; Kobayashi, H.; Iwabuchi, Y.; Kawahara, K.; Hirayama, S.; Ramstedt, M.; Sasaki, Y.; Kataoka, M.; Akeda, Y.; Ohnishi, M. A Highly Immunogenic Vaccine Platform against Encapsulated Pathogens Using Chimeric Probiotic Escherichia coli Membrane Vesicles. NPJ Vaccines 2022, 7, 153. [Google Scholar] [CrossRef] [PubMed]
- da Silva Barreira, D.; Lapaquette, P.; Novion Ducassou, J.; Couté, Y.; Guzzo, J.; Rieu, A. Spontaneous Prophage Induction Contributes to the Production of Membrane Vesicles by the Gram-positive Bacterium Lacticaseibacillus casei BL23. MBio 2022, 13, e0237522. [Google Scholar] [CrossRef]
- Díez-Sainz, E.; Milagro, F.I.; Riezu-Boj, J.I.; Lorente-Cebrián, S. Effects of Gut Microbiota-Derived Extracellular Vesicles on Obesity and Diabetes and Their Potential Modulation through Diet. J. Physiol. Biochem. 2022, 78, 485–499. [Google Scholar] [CrossRef] [PubMed]
- Jahromi, L.P.; Fuhrmann, G. Bacterial Extracellular Vesicles: Understanding Biology Promotes Applications as Nanopharmaceuticals. Adv. Drug Deliv. Rev. 2021, 173, 125–140. [Google Scholar] [CrossRef] [PubMed]
- Domínguez Rubio, A.P.; D’Antoni, C.L.; Piuri, M.; Pérez, O.E. Probiotics, Their Extracellular Vesicles and Infectious Diseases. Front. Microbiol. 2022, 13, 864720. [Google Scholar] [CrossRef]
- González-Lozano, E.; García-García, J.; Gálvez, J.; Hidalgo-García, L.; Rodríguez-Nogales, A.; Rodríguez-Cabezas, M.E.; Sánchez, M. Novel Horizons in Postbiotics: Lactobacillaceae Extracellular Vesicles and Their Applications in Health and Disease. Nutrients 2022, 14, 5296. [Google Scholar] [CrossRef]
- Yuan, Y.; Sun, J.; You, T.; Shen, W.; Xu, W.; Dong, Q.; Cui, M. Extracellular Vesicle-Based Therapeutics in Neurological Disorders. Pharmaceutics 2022, 14, 2652. [Google Scholar] [CrossRef]
- Sun, K.; Zheng, X.; Jin, H.; Yu, F.; Zhao, W. Exosomes as CNS Drug Delivery Tools and Their Applications. Pharmaceutics 2022, 14, 2252. [Google Scholar] [CrossRef]
- Abid, R.; Waseem, H.; Ali, J.; Ghazanfar, S.; Ali, G.M.; Elasbali, A.M.; Alharethi, S.H. Probiotic Yeast Saccharomyces: Back to Nature to Improve Human Health. J. Fungi 2022, 8, 444. [Google Scholar] [CrossRef] [PubMed]
- Saarela, M.H. Safety Aspects of Next Generation Probiotics. Curr. Opin. Food Sci. 2019, 30, 8–13. [Google Scholar] [CrossRef]
- Cunningham, M.; Azcarate-Peril, M.A.; Barnard, A.; Benoit, V.; Grimaldi, R.; Guyonnet, D.; Holscher, H.D.; Hunter, K.; Manurung, S.; Obis, D.; et al. Shaping the Future of Probiotics and Prebiotics. Trends Microbiol. 2021, 29, 667–685. [Google Scholar] [CrossRef] [PubMed]
- Cieślik, M.; Nazimek, K.; Bryniarski, K. Extracellular Vesicles-Oral Therapeutics of the Future. Int. J. Mol. Sci. 2022, 23, 7554. [Google Scholar] [CrossRef]
- Chang, X.; Wang, S.L.; Zhao, S.B.; Shi, Y.H.; Pan, P.; Gu, L.; Yao, J.; Li, Z.S.; Bai, Y. Extracellular Vesicles with Possible Roles in Gut Intestinal Tract Homeostasis and IBD. Mediators Inflamm. 2020, 2020, 1945832. [Google Scholar] [CrossRef]
- Hendrix, A.; De Wever, O. Systemically Circulating Bacterial Extracellular Vesicles: Origin, Fate, and Function. Trends Microbiol. 2022, 30, 213–216. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krzyżek, P.; Marinacci, B.; Vitale, I.; Grande, R. Extracellular Vesicles of Probiotics: Shedding Light on the Biological Activity and Future Applications. Pharmaceutics 2023, 15, 522. https://doi.org/10.3390/pharmaceutics15020522
Krzyżek P, Marinacci B, Vitale I, Grande R. Extracellular Vesicles of Probiotics: Shedding Light on the Biological Activity and Future Applications. Pharmaceutics. 2023; 15(2):522. https://doi.org/10.3390/pharmaceutics15020522
Chicago/Turabian StyleKrzyżek, Paweł, Beatrice Marinacci, Irene Vitale, and Rossella Grande. 2023. "Extracellular Vesicles of Probiotics: Shedding Light on the Biological Activity and Future Applications" Pharmaceutics 15, no. 2: 522. https://doi.org/10.3390/pharmaceutics15020522