1. Introduction
The
Flaviviridae family consist of 89 virus species divided into four genera. The genus
Flavivirus encompasses 53 enveloped positive-sense single RNA viruses. The virion consists of the three structural proteins (capsid (C), membrane (M), and envelope (E)) and seven nonstructural proteins presented in infected cells (NS1, NS2A, NS2B, NS3, NS4A, NS4B, and NS5) [
1,
2,
3].
Classified as mosquito-borne flaviviruses, Dengue virus (DENV), yellow fever virus (YFV), and Zika virus (ZIKV) are mainly transmitted by
Aedes spp. [
4] and West Nile virus (WNV) is transmitted by
Culex spp. [
5] DENV, YFV, and ZIKV are responsible for causing endemics or epidemics worldwide in tropical/subtropical regions [
6], and WNV is endo/epidemic in Europe [
7].
Mosquito-borne viruses are highly suited to the ‘one health’ concept for combating infectious diseases, demonstrating the interdependence of human, animal, and ecosystem health [
8]. Mosquito-borne viruses are categorized as zoonotic diseases, accounting for approximately 75% of emerging infectious diseases [
9], with more than 800 human pathogens [
10]. Wild animals are victims of those pathogens, but in some cases, animals only act as reservoirs. Transmission can occur through direct contact with infected animals, such as through food consumption or as pets.
Intense media coverage of SARS [
11], West Nile virus infection in the western hemisphere [
12], and other infectious diseases have brought public attention to the health risk of zoonoses and other emerging or re-emerging diseases.
Increased human mobility, population growth, the exploitation of a previously undisturbed ecosystem and climate change constitute major risk factors leading them to contact new pathogens and their vectors, consequently altering the behavior, migration, and interaction of pathogens and their hosts. A glaring example is the significant spread and establishment of vector-borne zoonotic diseases worldwide, such as WNV, Crimean–Congo hemorrhagic fever (CCHF), Japanese encephalitis virus (JEV) [
13], and the appearance of Chikungunya virus in Italy in 2007 [
3].
Human infection caused by flavivirus ranges from mild illness (flu-like febrile illnesses with a headache, myalgia, arthralgia, and a rash without long-term consequences) to severe and life-threatening disease, which is classified into two categories, visceral and neurotropic [
14].
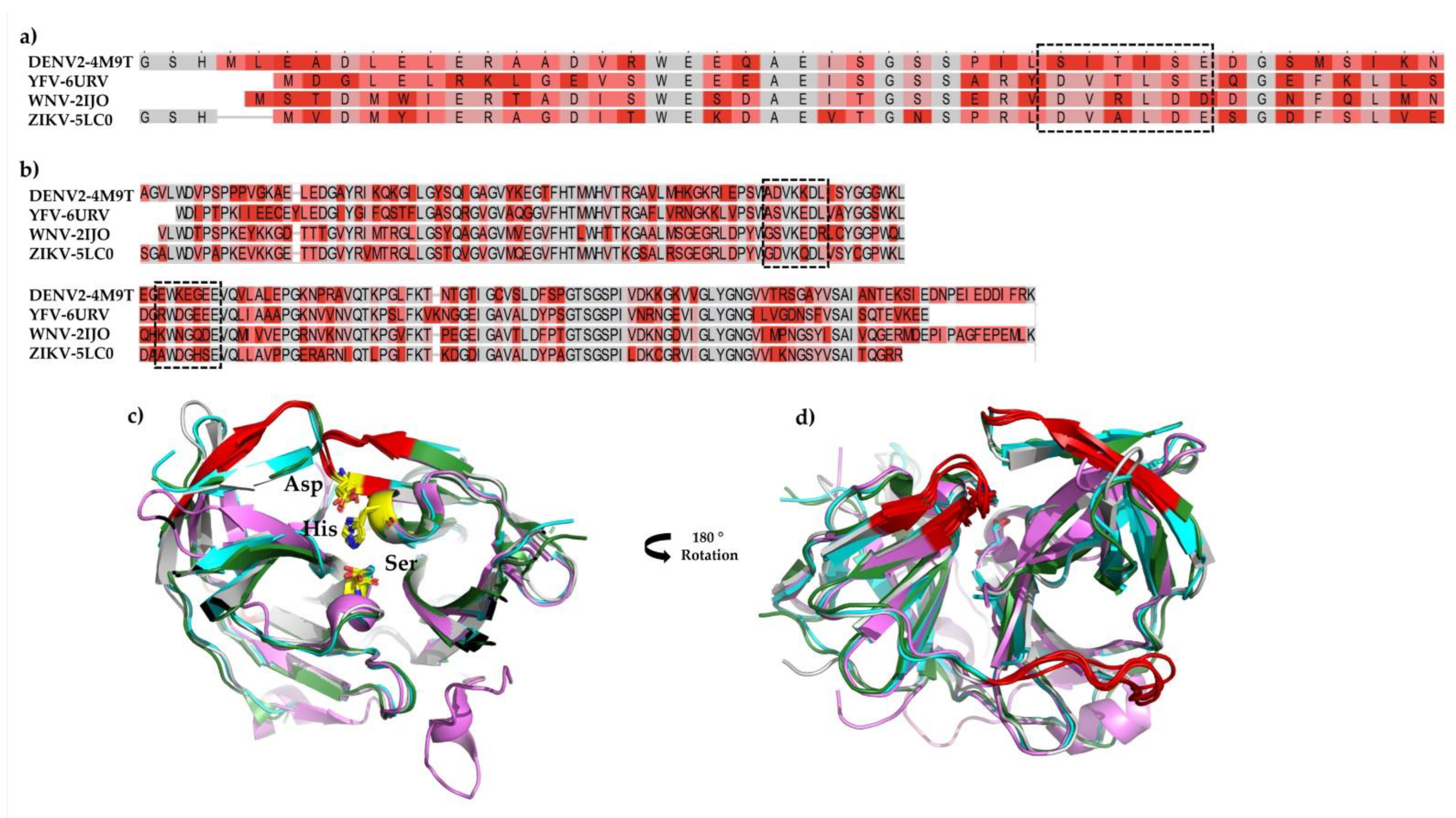
Among the viral proteins, NS3 is a multifunctional protein with N-terminal serine protease activity. However, the protease is a complex of two subunits, the already described Nter domain (~184 aa) and an essential cofactor (~40 aa) within the viral NS2B protein [
15,
16]. Highly conserved among flaviviruses, it is an essential enzyme in virus replication. Given its essential role, the NS2B/NS3 proteases (NS2B/NS3
pro) of flaviviruses are promising drug targets; for example, DENV2, YFV, WNV, and ZIKV NS2B/NS3
pros present high amino acids sequence conservation and all four have the same structural fold, which confers a similar substrate specificity profile [
16,
17]. Due to their importance, we selected all four viruses to study the effect of a bioactive plant derivative molecule against NS2B/NS3
pro.
To obtain much deeper insights and to identify a broad-spectrum molecule, we performed a combinatorial approach to identify how a small, potent molecule derived from green tea is able to inhibit the selected NS2B/NS3pro of DENV2, YFV, WNV, and ZIKV, which may serve as a lead compound to develop a potent antiviral drug.
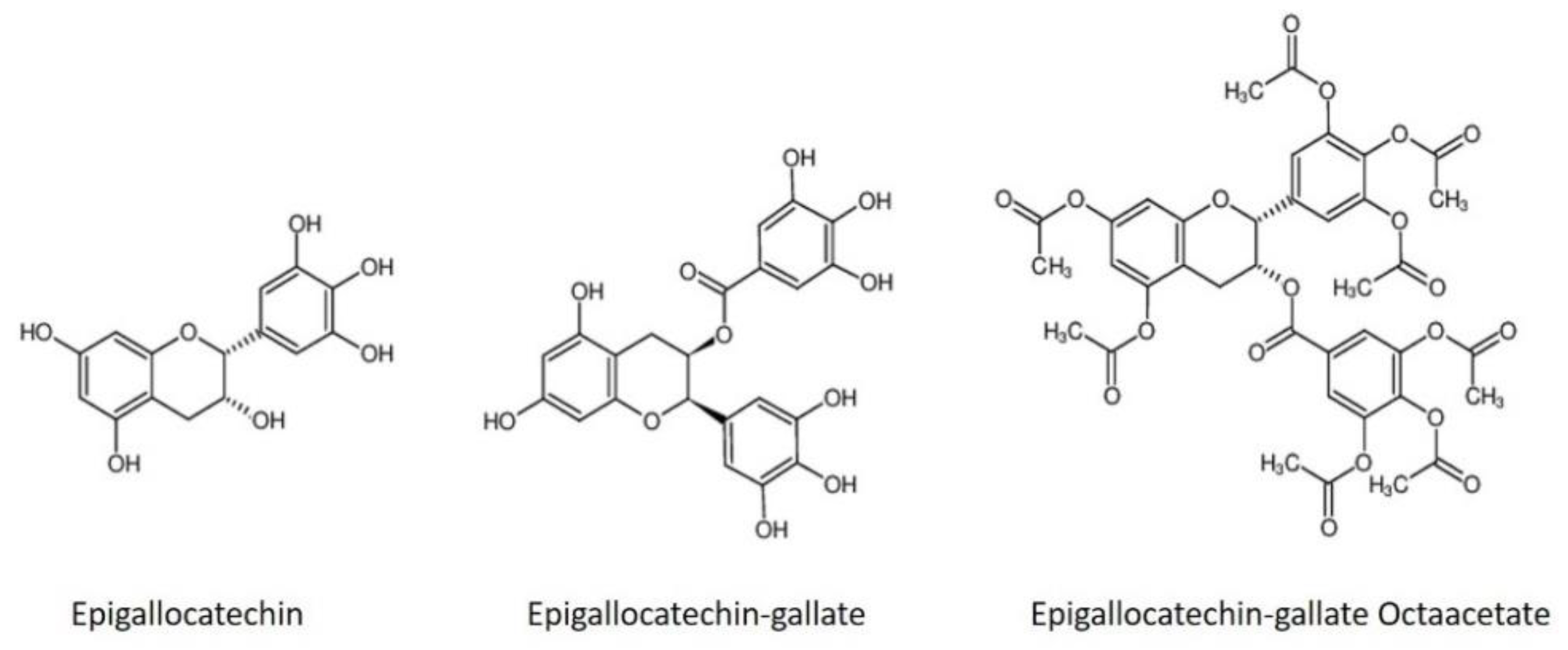
Epigallocatechin gallate (EGCG) and epigallocatechin (EGC) are polyphenols found in green tea; several studies have shown a broad-spectrum of antiviral activity against viral entry into host cells [
18,
19]. Li and collaborators [
16] showed the inhibition of ZIKV NS2B/NS3
pro by EGCG and ECG (epicatechin gallate) with an IC
50 value of 87 and 89 µM, respectively. It was also shown that EGCG inhibits ZIKV entry in Vero E6 cells [
20]. Recently, Liu and coll. demonstrated the effect of EGCG against SARS-CoV-2 inhibiting the spike protein from binding to the ACE2 receptor [
21] while reducing coronavirus replication in a mouse model [
22].
In this report, we have explored the in vitro and in silico inhibitory capacity of EGC, EGCG, and EGCG octaacetate (a modified form of EGCG) against NS2B/NS3 protease of DENV2, YFV, WNV, and ZIKV.
4. Discussion
Considering the concept of health, natural compounds that favor human and animal health would benefit, for example, in the defense against diseases transmitted by mosquitoes. Green tea catechins have a number of proven positive effects on human and animal health [
43,
44]. Natural molecules account for over one-third of all new, FDA-approved products, demonstrating the importance of plants and animals as a source of bioactive molecules [
30]. Epigallocatechin gallate (EGCG) and epigallocatechin (EGC) are catechins and represent 59 and 19% of the total polyphenols in green tea extract [
45].
This work reports the in vitro validation of promising epigallocatechin molecules against NS2B/NS3 proteases from four flaviviruses (DENV2, YFV, WNV, and ZIKV) and shows that EGC, EGCG, and its derivative EGCG octaacetate inhibit protease activity, with the EGCG showing the finest IC
50, in the range of 1.8 to 8 µM. It has already been described that EGCG showed antiviral activity during the early steps of infection, as well as a virucidal agent [
18,
20,
46,
47].
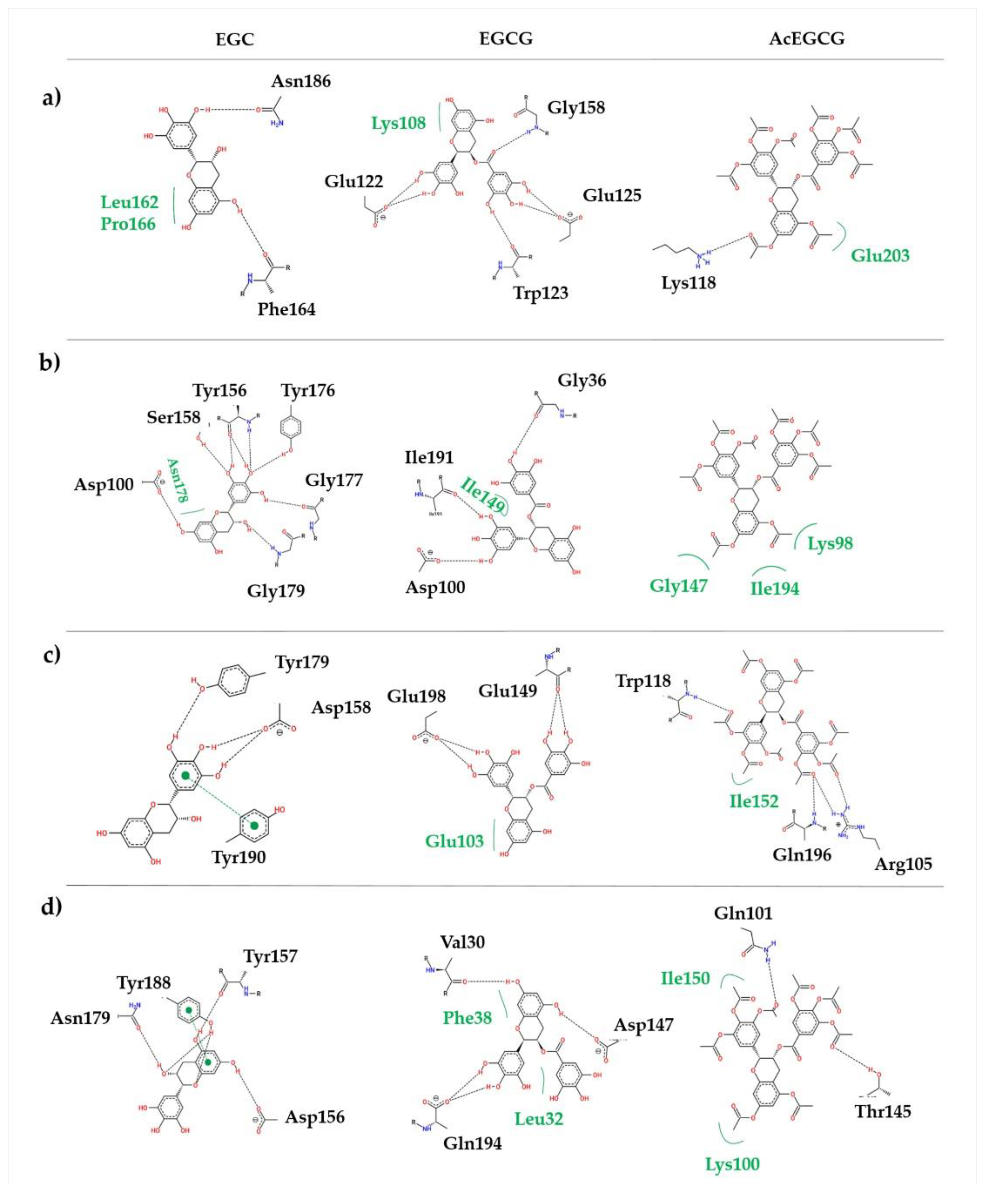
The mode of action of these molecules is not very well understood, but in this work, we can hypothesize that the molecules interact with the NS2B/NS3
pro; however, in cell infection studies, they might interact with multiple binding sites [
48]. In this sense, in vitro studies were performed, and the inhibition mode demonstrated the probable interaction site of the molecules on the NS2B/NS3
pro. Molecular docking and dynamics simulation studies predicted the epigallocatechin molecule’s binding pocket, showing that the EGC potentially forms interactions with the activity site of the studied proteins. On the other hand, EGCG and AcEGCG molecules interact with the viral protease allosterically.
Despite the polymorphism identified in several viral proteins, this modification may not affect the catalytic efficiency of the enzyme [
49,
50]. Therefore, further studies are needed to confirm this; however, it seems reasonable to assume the inhibition and interaction properties of the molecules observed in this study.
The molecule EGC exhibited a competitive inhibition model suggesting that even if the molecule interacts with amino acid residues in the active/binding site, it still allows the interaction between the substrate and the protein, probably regarding the size of the molecule.
In our in vitro assay, the EGCG molecule interacts noncompetitively with the proteases, indicating that this molecule is an allosteric inhibitor. We suggest the trihydroxybenzoate ring prevents the molecule from interacting with the active site. Yadav and colleagues [
51] described the interaction of ZIKV NS2B/NS3
pro with the EGCG molecule through the active site using in silico studies, which disagrees with our experimental results, which show that this molecule is an allosteric inhibitor. In addition to their virucidal capacity, several studies have shown that EGCG can act by blocking the necessary receptors for viral attachment to the target cell, as in the case of HIV, where the drug binds to CD4 expressed on T cells and inhibits the virus binding to this site, blocking viral infection [
20,
52]. Carneiro and coll. [
20] described no influence of the molecule on viral infection, using a pre-treatment assay, which suggested that EGCG has no action on the expression or in blocking the cellular receptors used by ZIKV during the entry process in the host cell. Vázquez-Calvo and colleagues [
18] described that the EGCG molecule directly affects WNV particles exerting a virucidal effect and reducing the effectivity of ZIKV and DENV. The inhibitory capacity of green tea catechins has also been described for SARS-CoV-2 main protease [
18,
49], HasV endopeptidase [
50], HCV NS3/4A protease [
51], and HIV-1 protease [
52].
Here, we have demonstrated that the epigallocatechin molecules presented in this study, natural compounds found in abundance in many foods (EGC and EGCG) and its modified form (AcEGCG), inhibit in vitro the activity of the essential virus protease (NS2B/NS3). Despite these encouraging results, we have shown that combining the EGC and EGCG molecules, which displayed different interaction modes (competitive and noncompetitive), drastically increased the inhibition of the YFV, WNV, and ZIKV proteases, displaying an IC50 value in the nM range.
EGCG and its derivatives have a range of wide biological activity, but in the human body, these molecules can undergo multiple processes of metabolization [
53,
54,
55]. Temperature, pH, and oxidant concentration accelerate the degradation rate [
56]. Green tea catechins are very unstable in the saliva, stomach, and upper intestine [
57]. The intestinal microflora has the potential to dissociate and degrade EGCG in vitro and in vivo [
58]. The chemical instability of green tea catechins reduces intestinal absorption and affects bioavailability. However, several studies have demonstrated the improvement of bioaccessibility by nanoliposome encapsulation [
57], structural modification [
55], methylation [
59], cyclisation [
60], or glycosidation [
61]. Even though epigallocatechin molecules are derived from food plants, high doses have side effects on the human body; for example, green tea catechins can trigger anxiolytic reactions, hypoglycemic activity, hypochromic anemia, and liver and kidney failure. However, these effects are not related to the host serine proteases [
62,
63].
In conclusion, our results have revealed the antiviral activity of two different epigallocatechin, EGC and EGCG, and its derivative, AcEGCG, against the protease of four clinically relevant members of the Flavivirus genus (DENV2, YFV, WNV, and ZIKV) and point to EGC and EGCG as potential scaffolds for the prospect design of mixed antiviral compounds that affect the viral protein in two different binding regions. Optimization of these bioactive molecules is essential to achieve high oral bioavailability, including chemical modifications, absorption, and penetration enhancers, as well as formulation design. It is important to emphasize that oral delivery of new lead molecules can be facilitated by a thorough understanding of the interaction of the inhibitory molecule with its target.



 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}