

Zinc(II)-Sterol Hydrazone Complex as a Potent Anti-Leishmania Agent: Synthesis, Characterization, and Insight into Its Mechanism of Antiparasitic Action

 ,
,
Abstract
:1. Introduction
2. Experimental Section
2.1. Materials and Methods
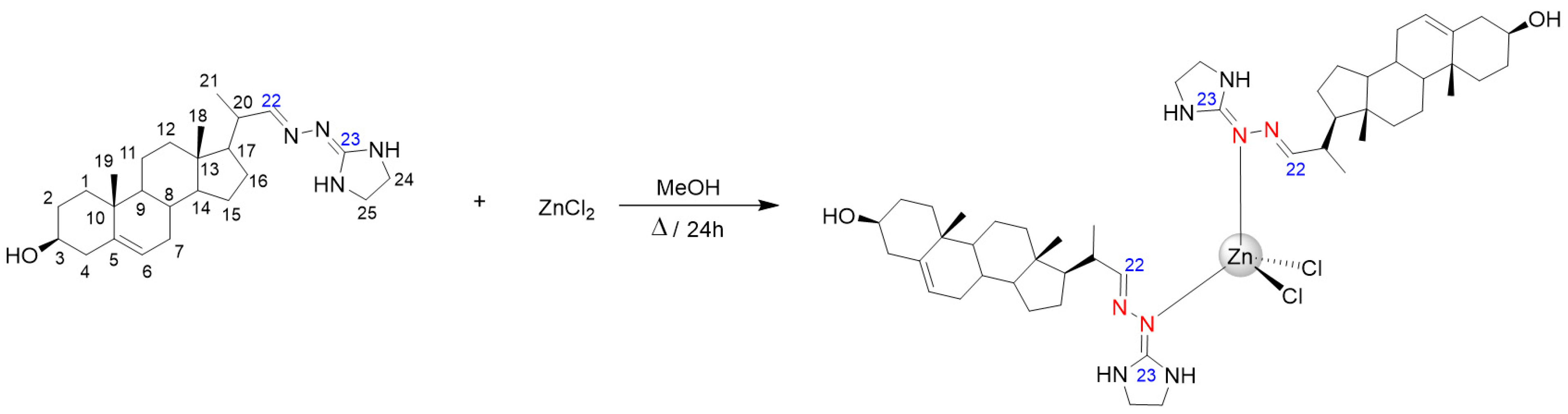
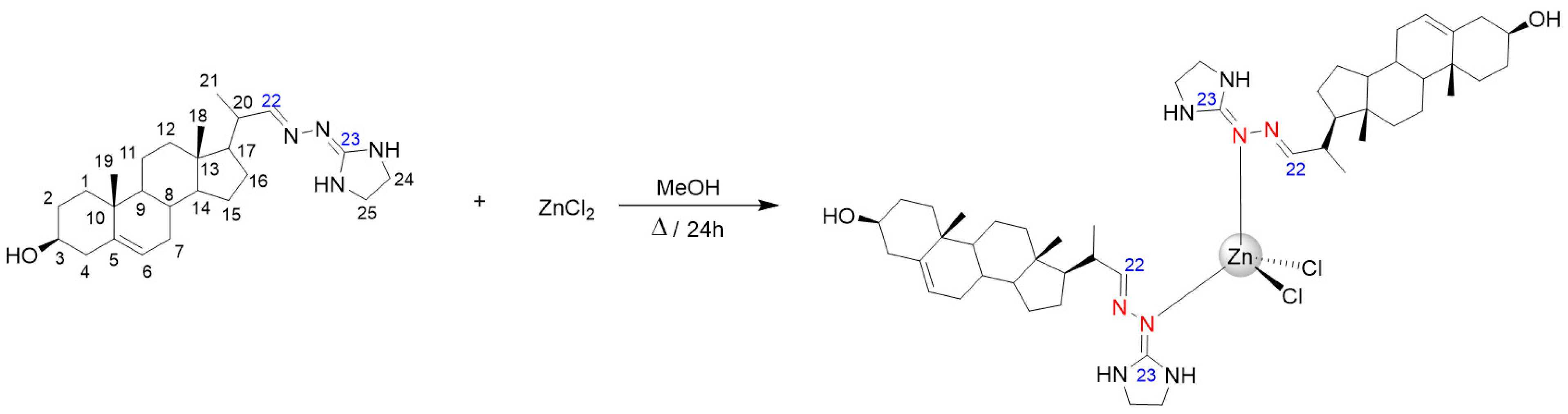
2.2. Synthesis of ZnCl2(H3)2
2.3. Biological Assays
3. Results and Discussions
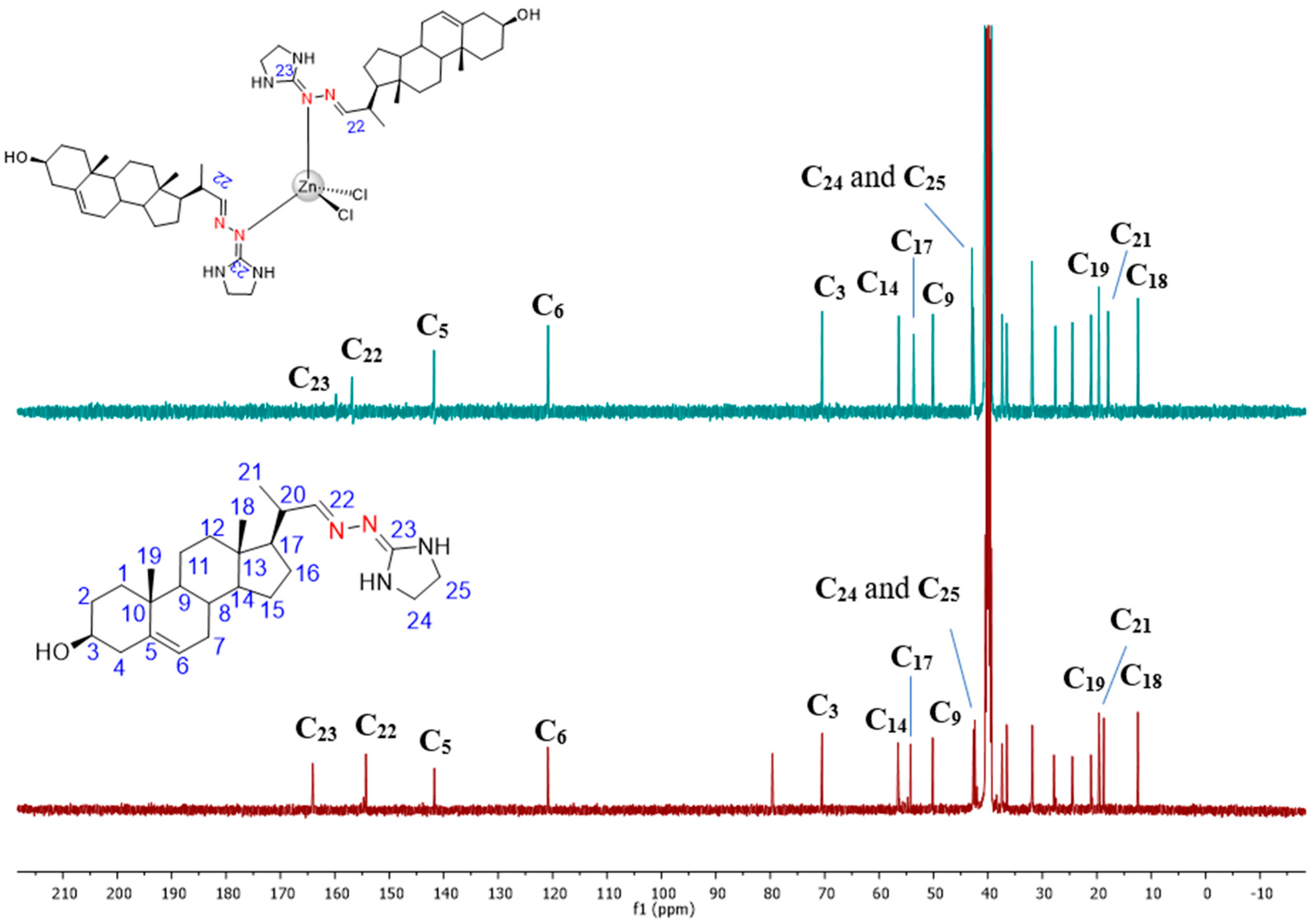
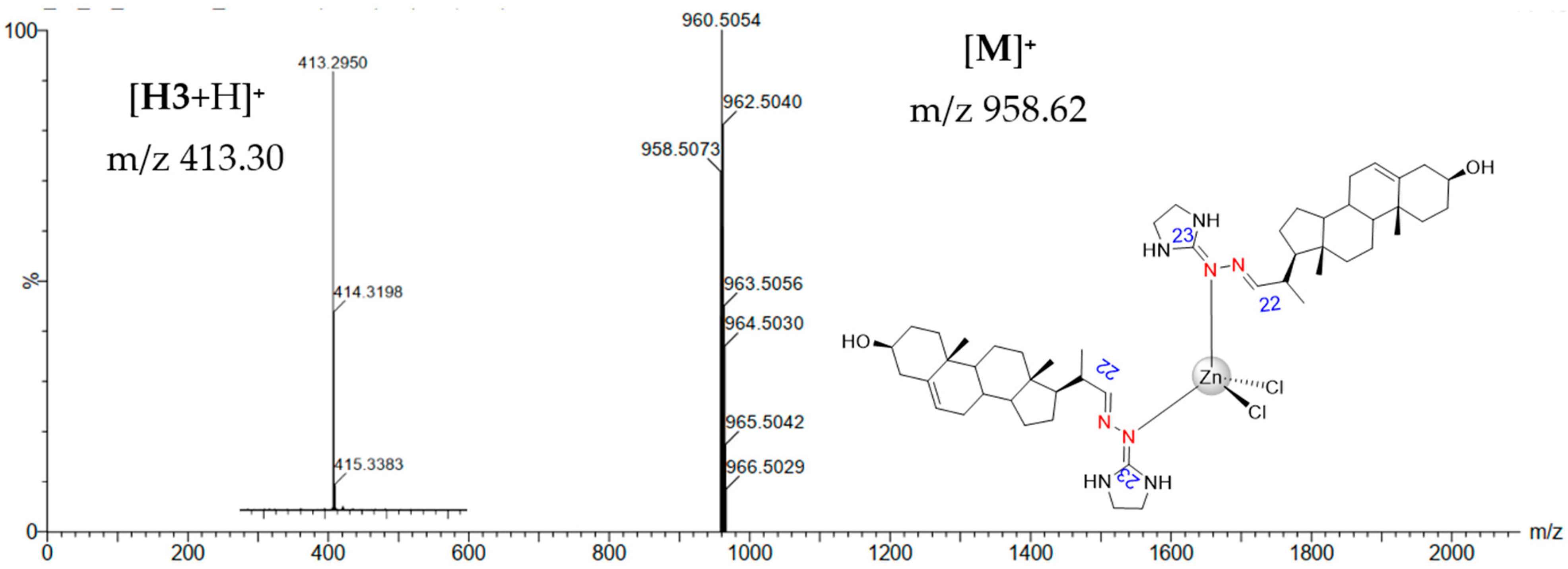
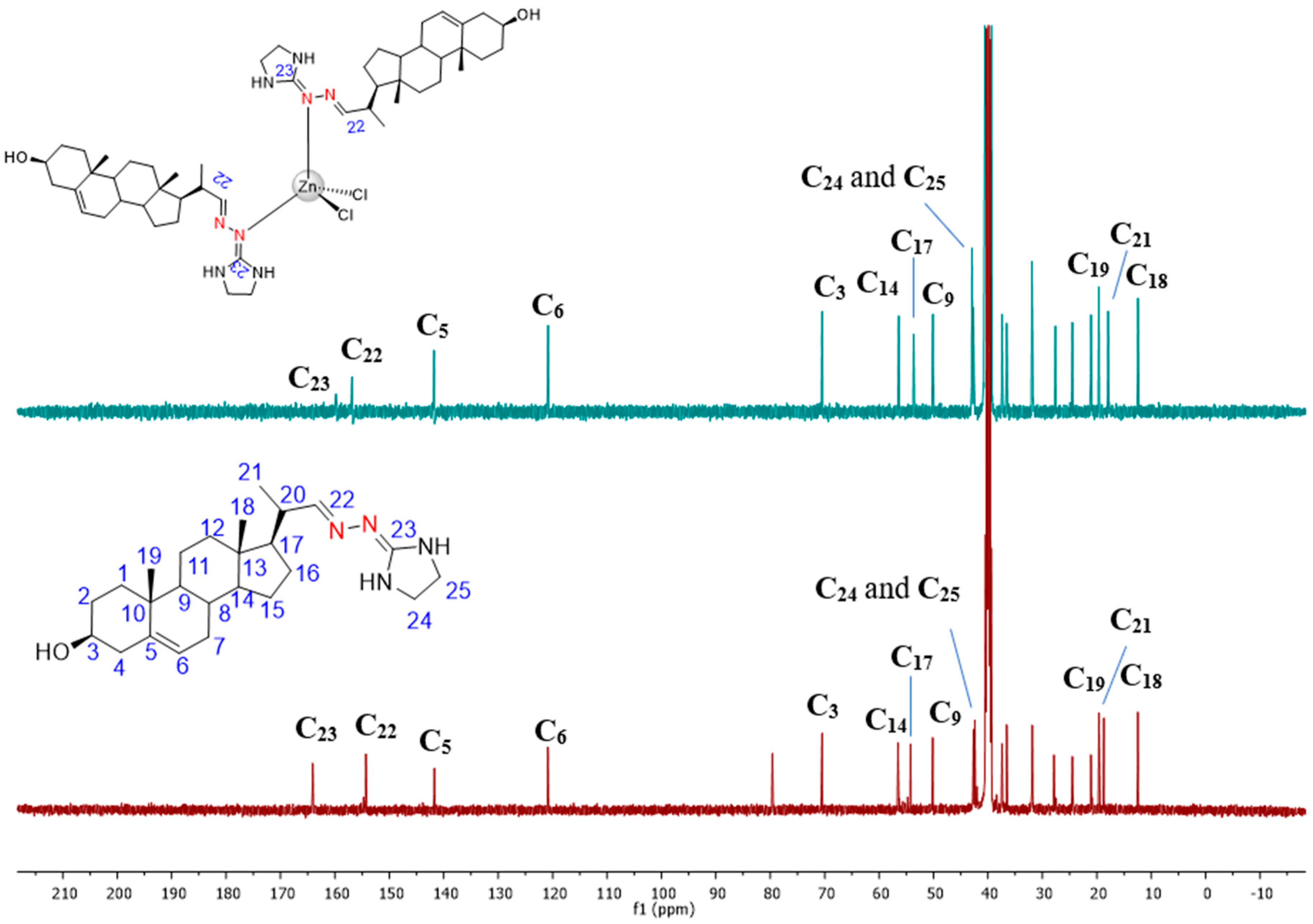
3.1. Chemical Section
3.2. Biological Activity
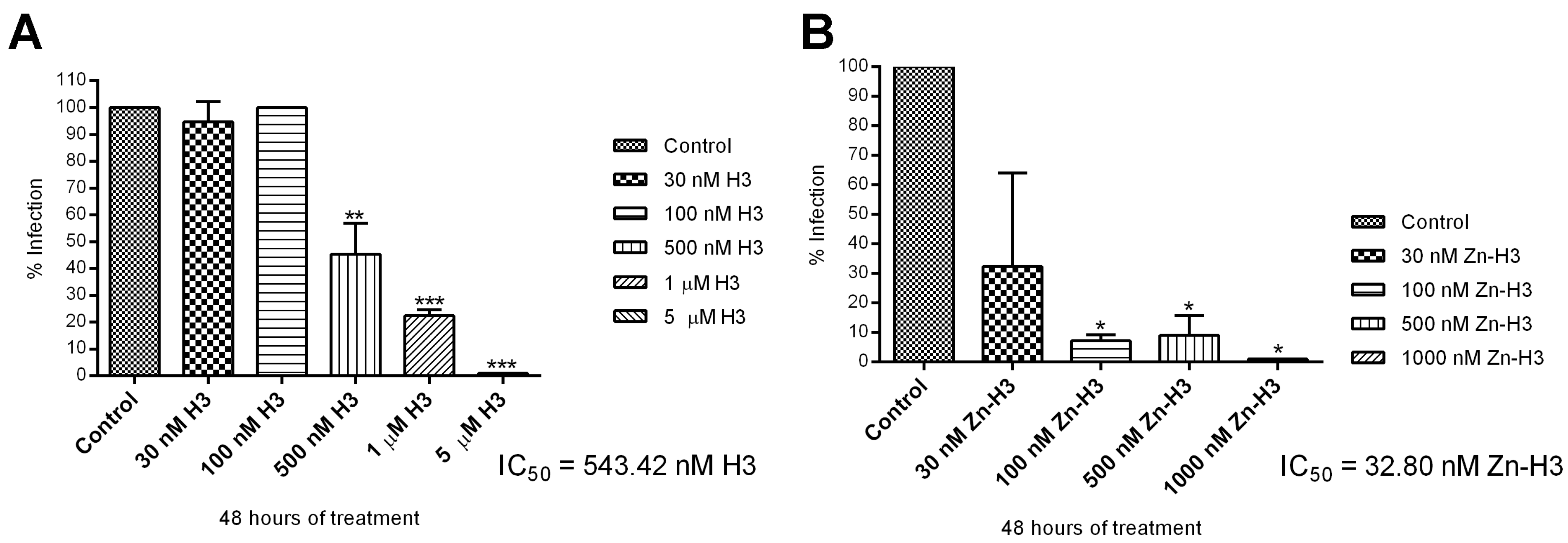
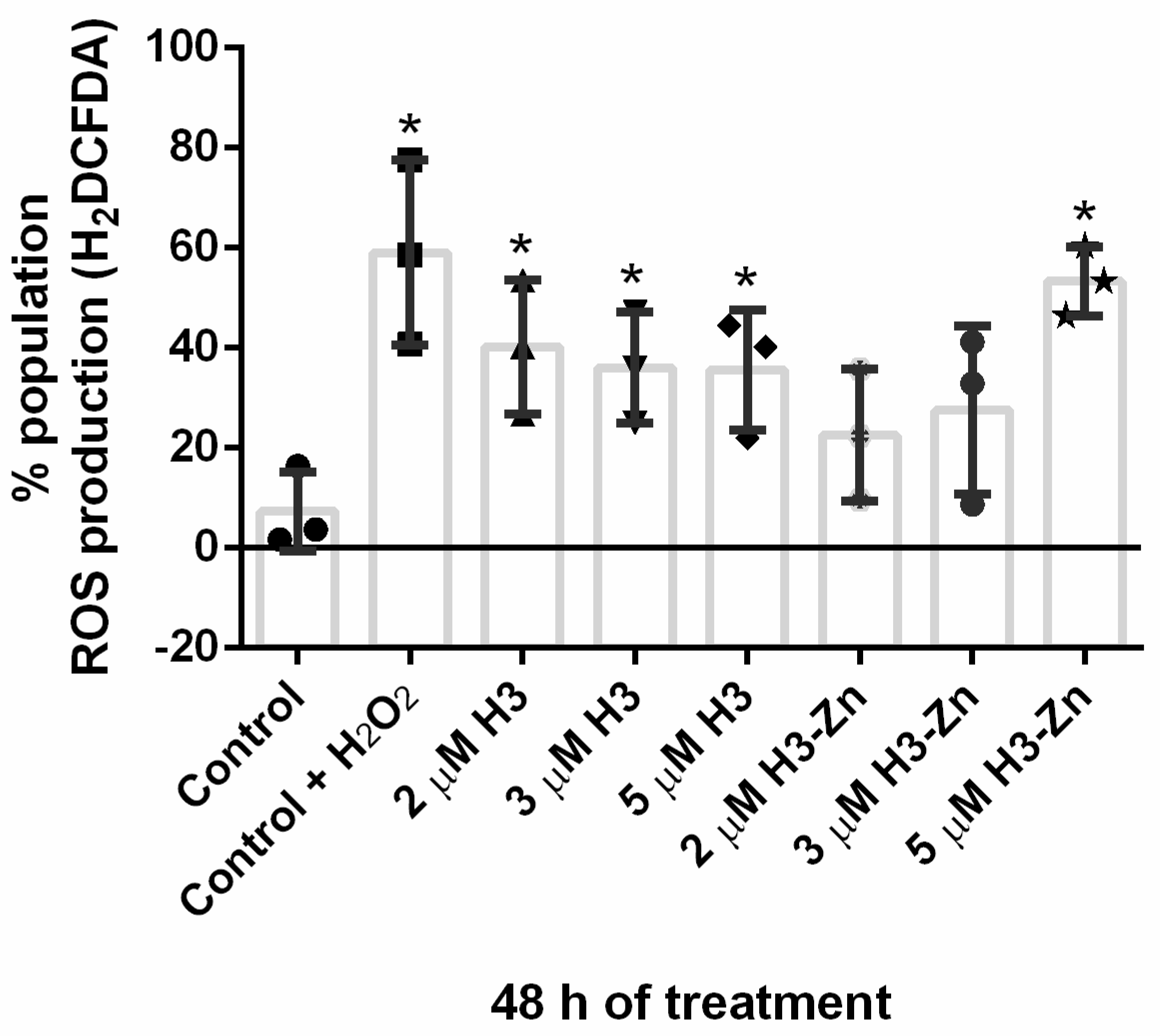
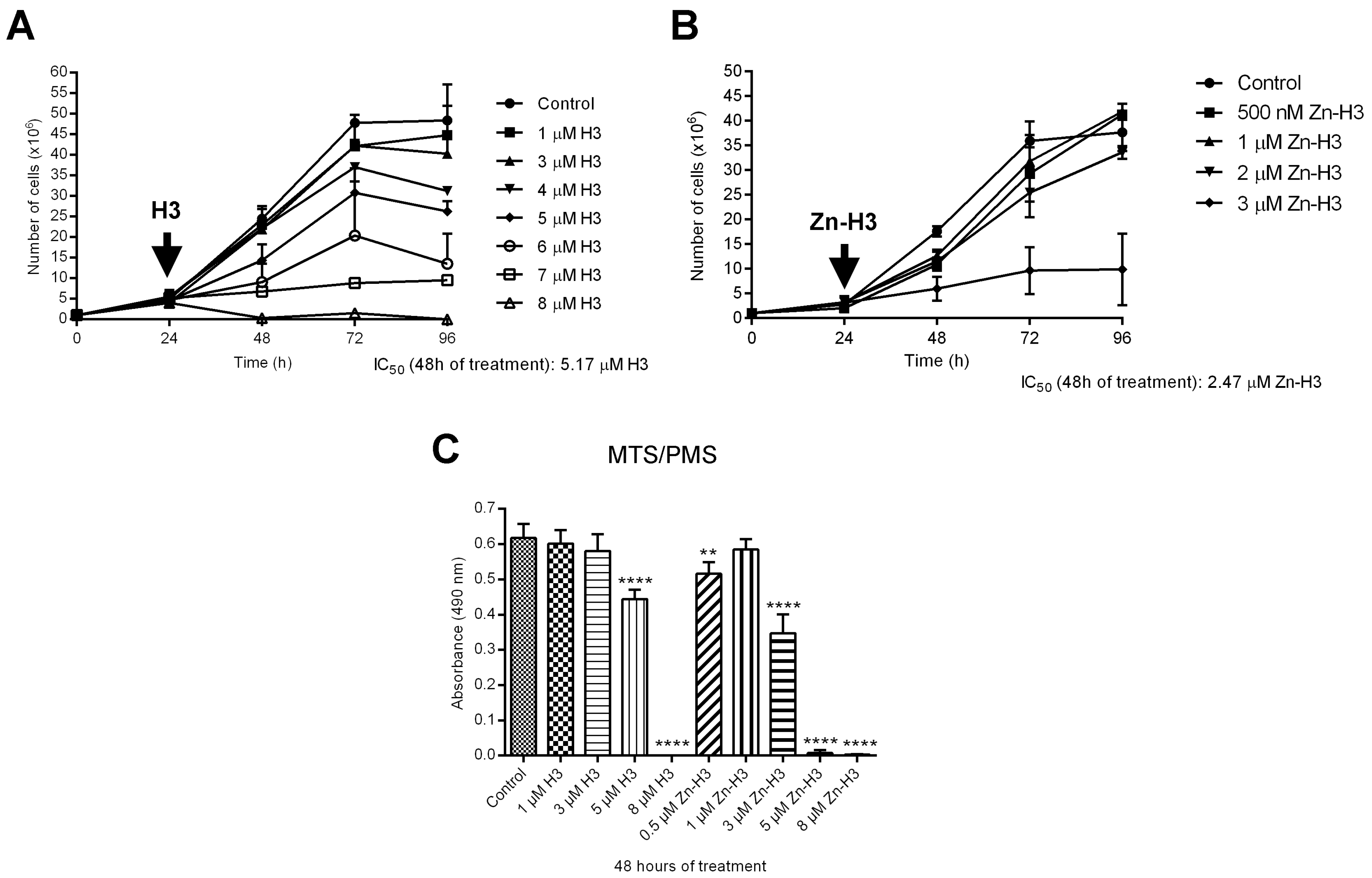
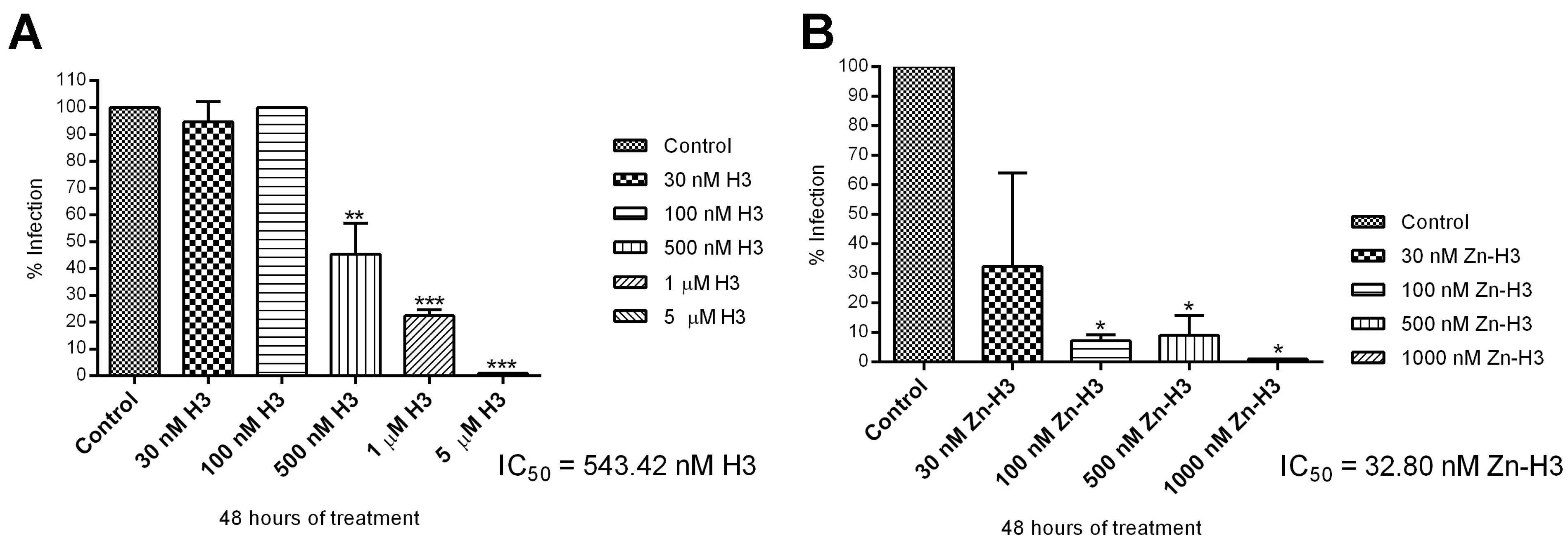
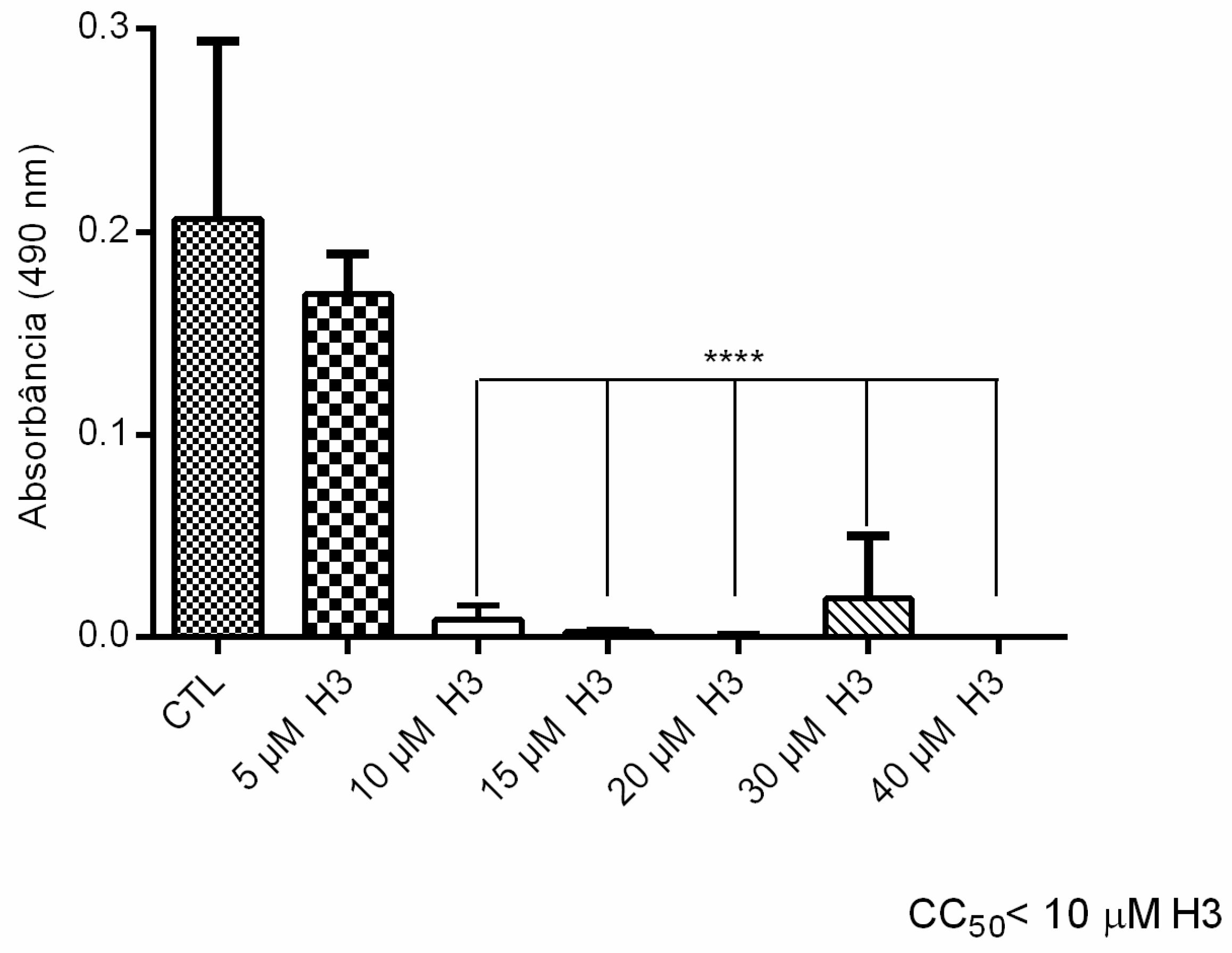
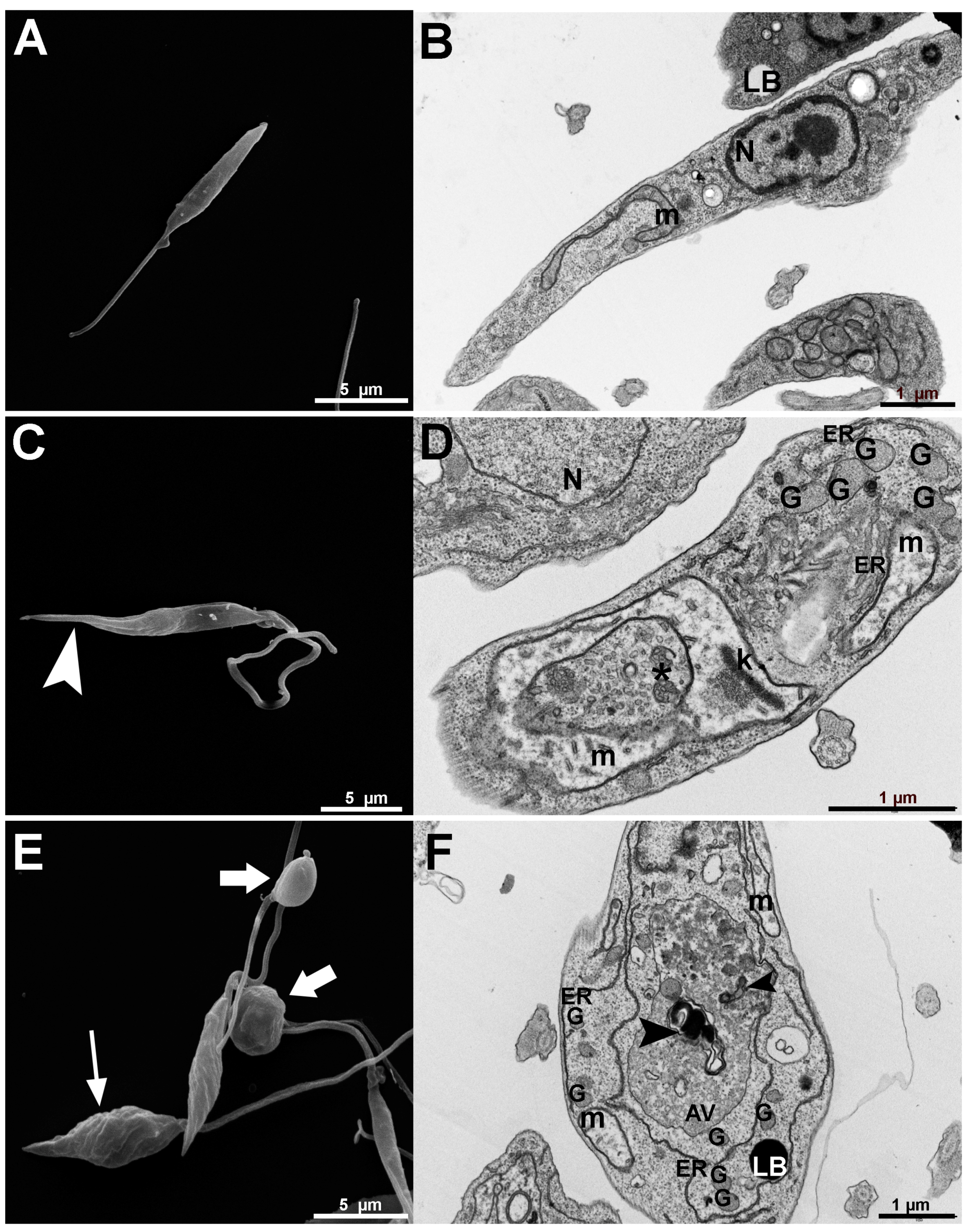
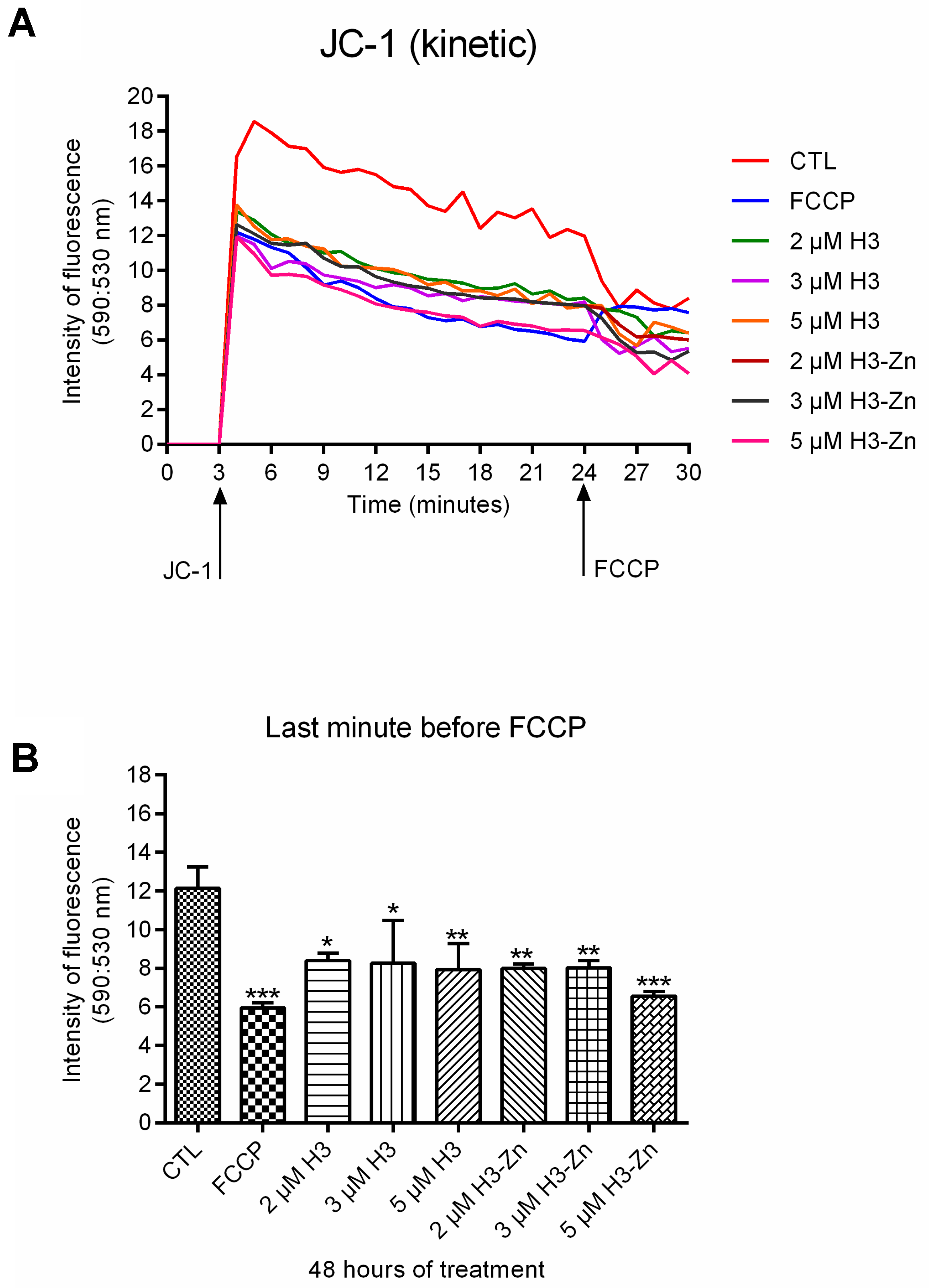
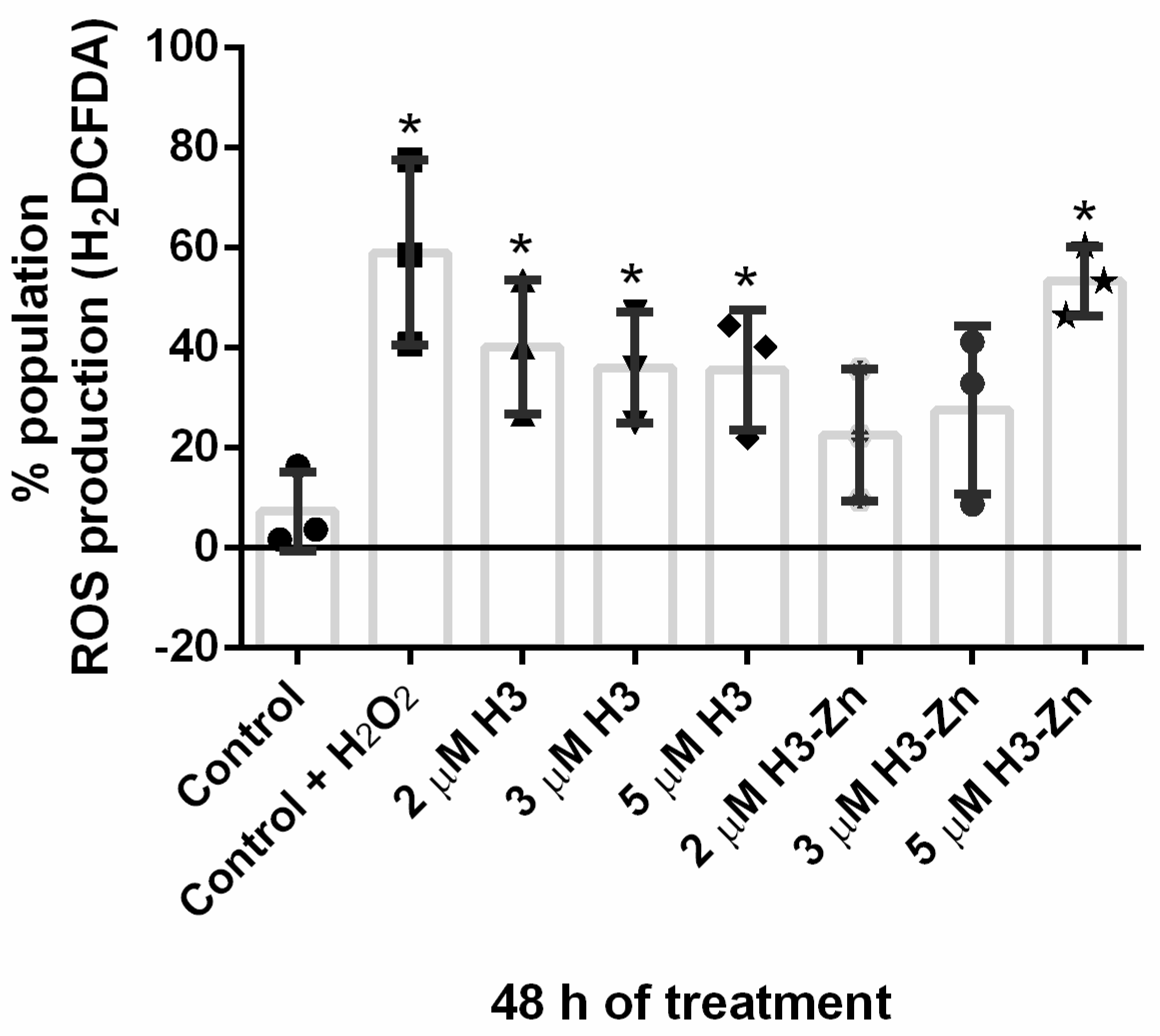
3.2.1. Antiproliferative, Ultrastructural, and Physiological Studies on L. amazonensis Promastigotes and Intracellular Amastigotes
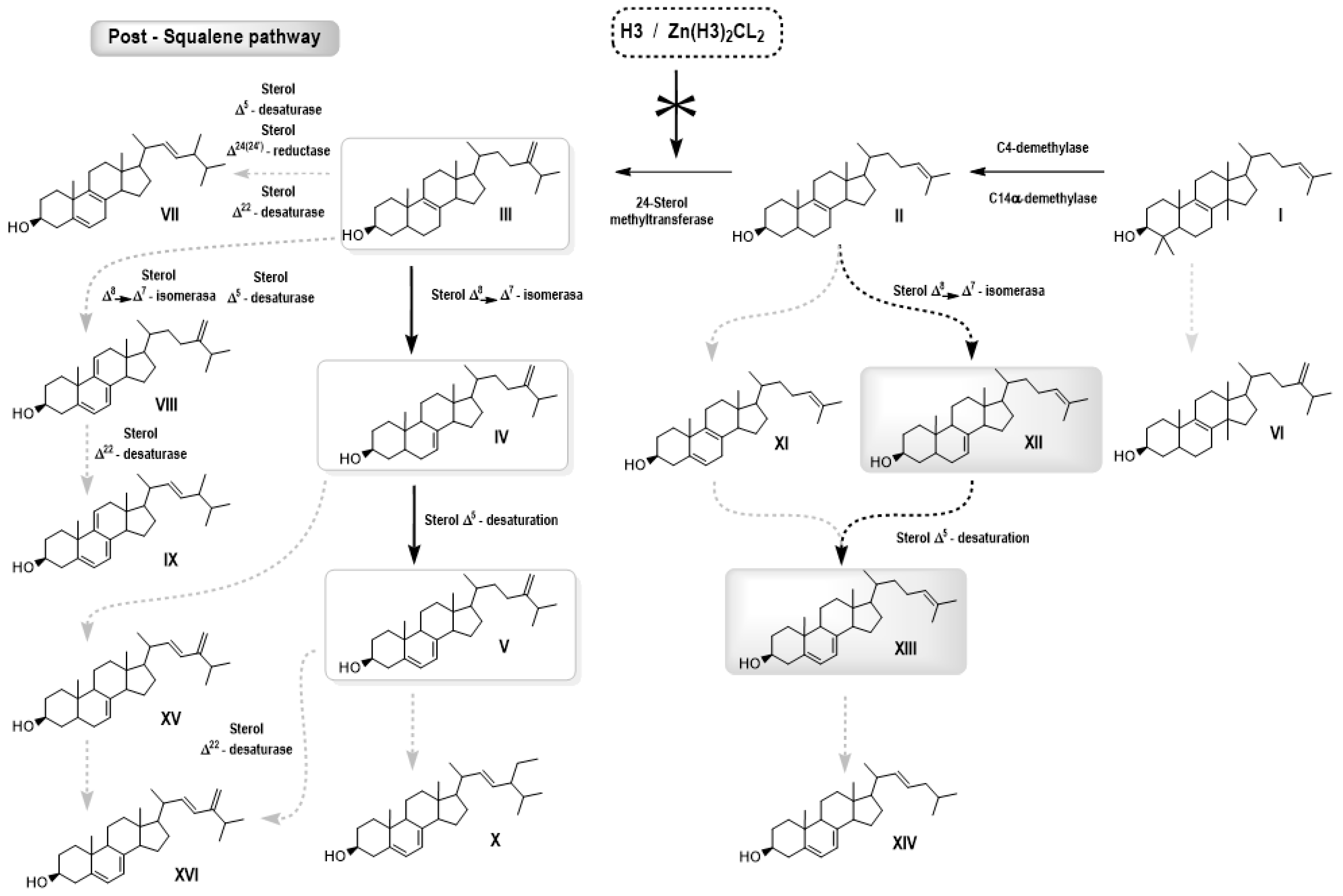
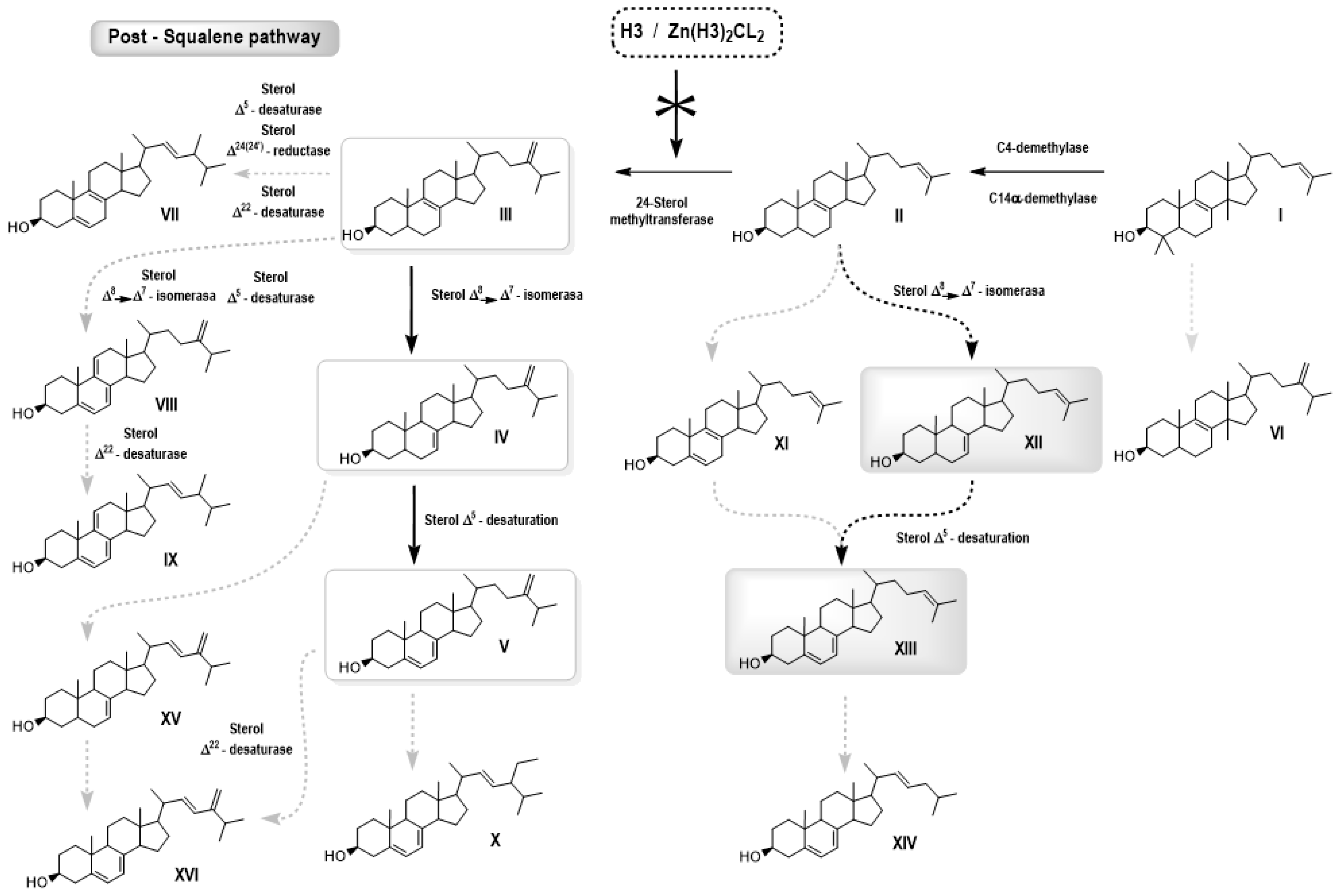
3.2.2. Free Sterol Analyses of L. amazonensis Promastigotes Control and Treated with H3 and ZnCl2(H3)2
4. Conclusions
5. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gossage, S.M.; Rogers, M.E.; Bates, P.A. Two separate growth phases during the development of Leishmania in sand flies: Implications for understanding the life cycle. Int. J. Parasitol. 2003, 33, 1027–1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burza, S.; Simon, L.; Croft, S.L.; Boelaert, M. Leishmaniasis. Lancet 2018, 392, 951–970. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Postigo, J.A.; Jain, S.; Mikhailov, A.; Maia-Elkhoury, A.N.; Valadas, S.; Warusavithana, S.; Osman, M.; Lin, Z.; Beshah, A.; Yajima, A.; et al. Global leishmaniasis surveillance: 2019–2020, a baseline for the 2030 roadmap. WHO Ref. Number WER 2021, 96, 401–409. [Google Scholar]
- Silva Santos, S.; de Araujo, R.V.; Giarolla, J.; Seoud, O.E.; Ferreira, E.I. Searching drugs for Chagas disease, leishmaniasis and schistosomiasis: A brief review. Int. J. Antimicrob. Agents 2020, 55, 105906. [Google Scholar] [CrossRef]
- Navarro, M.; Justo, R.M.; Delgado, G.Y.S.; Visbal, G. Metallodrugs for the Treatment of Trypanosomatid Diseases: Recent Advances and New Insights. Curr. Pharm. Des. 2021, 27, 1763–1789. [Google Scholar] [CrossRef]
- Teixeira de Macedo-Silva, S.; de Souza, W.; Fernandes Rodrigues, J.C. Sterol Biosynthesis Pathway as an Alternative for the Anti-Protozoan Parasite Chemotherapy. Curr. Med. Chem. 2015, 22, 2186–2198. [Google Scholar] [CrossRef]
- Nelson, D.L.; Cox, M.M. Lehninger Principles of Biochemistry, 3rd ed.; Worth Publishers: New York, NY, USA, 2000. [Google Scholar]
- Park, J.; Pandya, V.R.; Ezekiel, S.J.; Berghuis, A.M. Phosphonate and Bisphosphonate Inhibitors of Farnesyl Pyrophosphate Synthases: A Structure-Guided Perspective. Front. Chem. 2021, 8, 612728. [Google Scholar] [CrossRef]
- Sealey-Cardona, M.; Cammerer, S.; Jones, S.; Ruiz-Pérez, L.M.; Brun, R.; Gilbert, I.H.; Urbina, J.A.; González-Pacanowska, D. Kinetic Characterization of Squalene Synthase from Trypanosoma cruzi: Selective Inhibition by Quinuclidine Derivatives. Antimicrob. Agents Chemother. 2007, 51, 2123–2129. [Google Scholar] [CrossRef] [Green Version]
- de Macedo-Silva, S.T.; Visbal, G.; Urbina, J.A.; de Souza, W.; Rodrigues, J.C.F. Potent In Vitro Antiproliferative Synergism of Combinations of Ergosterol Biosynthesis Inhibitors against Leishmania amazonensis. Antimicrob. Agents Chemother. 2015, 59, 6402–6418. [Google Scholar] [CrossRef] [Green Version]
- Emami, S.; Tavangar, P.; Keighobadi, M. An overview of azoles targeting sterol 14α-demethylase for antileishmanial therapy. Eur. J. Med. Chem. 2017, 135, 241–259. [Google Scholar] [CrossRef]
- Liu, J.; Nes, W.D. Steroidal Triterpenes: Design of Substrate-Based Inhibitors of Ergosterol and Sitosterol Synthesis. Molecules 2009, 14, 4690–4706. [Google Scholar] [CrossRef] [Green Version]
- Visal, G. Azasterol as possible Antifungal and Antiparasitic Drugs. J. Anal. Pharm. Res. 2018, 7, 00199. [Google Scholar] [CrossRef]
- Urbina, J.A.; Vivas, J.; Lazardi, K.; Molina, J.; Payares, G.; Pirns, M.M.; Piras, R. Antiproliferative Effects of ∆24(25) Sterol Methyl Transferase Inhibitors on Trypanosoma (Schizotrypanum) cruzi: In vitro and in vivo Studies. Chemotherapy 1996, 42, 294–307. [Google Scholar] [CrossRef] [PubMed]
- Visbal, G.; San-Blas, G.; Maldonado, A.; Álvarez-Aular, Á.; Capparelli, M.V.; Murgich, J. Synthesis, in vitro antifungal activity and mechanism of action of four sterol hydrazone analogues against the dimorphic fungus Paracoccidioides brasiliensis. Steroids 2011, 76, 1069–1081. [Google Scholar] [CrossRef] [PubMed]
- Borba-Santos, L.P.; Visbal, G.; Gagini, T.; Rodrigues, A.M.; De Camargo, Z.P.; Lopes-Bezerra, L.M.; Rozental, S. ∆24-sterol methyltransferase plays an important role in the growth and development of Sporothrix schenkii and Sporothrix brasiliensis. Front. Microbiol. 2016, 7, 311. [Google Scholar] [CrossRef] [Green Version]
- Farrell, N.P.; Williamson, J.; Maclaren, D.J. Trypanocidal and antitumor activity of platinum-metal and platinum metal-drug dual-function complexes. Biochem. Pharmacol. 1984, 33, 961–971. [Google Scholar] [CrossRef]
- de Azevedo-França, J.A.; Borba-Santos, L.P.; Pimentel, G.D.A.; Franco, C.H.J.; Souza, C.; Celestino, J.D.A.; de Menezes, E.F.; dos Santos, N.P.; Vieira, E.G.; Ferreira, A.M.D.C.; et al. Antifungal promising agents of zinc(II) and copper(II) derivatives based on azole drug. J. Inorg. Biochem. 2021, 219, 111401. [Google Scholar] [CrossRef] [PubMed]
- Pereira, L.D.; Vila, T.; Borba-Santos, L.P.; De Souza, W.; Navarro, M.; Rozental, S. Activity of Metal-Azole Complexes Against Biofilms of Candida albicans and Candida glabrata. Curr. Pharm. Des. 2020, 26, 1524–1531. [Google Scholar] [CrossRef] [PubMed]
- Ong, Y.C.; Roy, S.; Andrews, P.C.; Gasser, G. Metal Compounds against Neglected Tropical Diseases. Chem. Rev. 2018, 119, 730–796. [Google Scholar] [CrossRef]
- Visbal, G.; Higuera, A.; Santos, L.B.; de Souza, W.; Rozental, S.; Navarro, M. Metal-azasterol complexes: Synthesis, characterization, interaction studies with DNA and TrxR and Biological Evaluation. J. Mex. Chem. Soc. 2017, 61, 146–157. [Google Scholar] [CrossRef]
- Wilson, J.; Lippard, S.J. Synthetic Methods for the Preparation of Platinum Anticancer Complexes. Chem. Rev. 2014, 114, 4470–4495. [Google Scholar] [CrossRef] [Green Version]
- Nardon, C.; Boscutti, G.; Fregona, D. Beyond platinums: Gold complexes as anticancer agents. Anticancer. Res. 2014, 34, 487–492. [Google Scholar] [PubMed]
- Cini, M.; Bradshaw, T.D.; Woodward, S. Using titanium complexes to defeat cancer: The view from the shoulders of titans. Chem. Soc. Rev. 2017, 46, 1040–1051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levina, A.; Mitra, A.; Lay, P.A. Recent developments in ruthenium anticancer drugs. Metallomics 2009, 1, 458–470. [Google Scholar] [CrossRef] [PubMed]
- Sun, R.W.-Y.; Ma, D.-L.; Wong, E.L.-M.; Che, C.-M. Some uses of transition metal complexes as anti-cancer and anti-HIV agents. Dalton Trans. 2007, 43, 4884–4892. [Google Scholar] [CrossRef]
- Condé, C.A.S.R.; De Almeida, M.V.; Silva, G.d.S.D.; Sodré, M.B.P.d.A.; Rodrigues, J.C.F.; Navarro, M. Synthesis, characterization and antileishmanial activity of copper(II) and zinc(II) complexes with diamine ligands. Transit. Met. Chem. 2022, 47, 147–156. [Google Scholar] [CrossRef]
- Osredkar, J.; Sustar, N. Copper and zinc, biological role and significance of copper/zinc imbalance. J. Clinic Toxicol. S 2011, 3, 0495. [Google Scholar] [CrossRef] [Green Version]
- Faller, P.; Hureau, C. Bioinorganic chemistry of copper and zinc ions coordinated to amyloid-β peptide. Dalton Trans. 2009, 1080–1094. [Google Scholar] [CrossRef]
- Yoshikawa, Y.; Ueda, E.; Kawabe, K.; Miyake, H.; Takino, T.; Sakurai, H.; Kojima, Y. Development of new insulinomimetic zinc(II) picolinate complexes with a Zn(N2O2) coordination mode: Structure characterization, in vitro, and in vivo studies. JBIC J. Biol. Inorg. Chem. 2002, 7, 68–73. [Google Scholar] [CrossRef]
- Justo, R.M.S.; Borba-Santos, L.P.; Sade, Y.B.; Ferreira, F.H.C.; Rosa, N.M.P.; Costa, L.A.S.; Visbal, G.; Rozental, S.; Navarro, M. Synthesis, characterization and biological evaluation of zinc and copper azasterol complexes against Sporothrix brasiliensis. New J. Chem. 2021, 45, 20840–20849. [Google Scholar] [CrossRef]
- Visbal, G.; Navarro, M.; Justo, R.M.S.; Rodrigues, J.C.F.; Silva, S.T.M.; De Souza, W.; Da Silva, G.S. Delta 24-Sterol Methyltransferase Enzyme Inhibitor Metalloazasterol Compound, Pharmaceutical Composition and Use of the Compound. WO2022/061431 A1; Patent Application. Granted 2022. Available online: https://www.lens.org/lens/search/patent/list?q=WO2022%2F061431%20A1 (accessed on 9 March 2023).
- Henriques, C.; Moreira, T.L.B.; Maia-Brigagão, C.; Henriques-Pons, A.; Carvalho, T.M.U.; de Souza, W. Tetrazolium salt based methods for high-throughput evaluation of anti-parasite chemotherapy. Anal. Methods. 2011, 3, 2148–2155. [Google Scholar] [CrossRef]
- Silva, S.T.D.M.; Visbal, G.; Godinho, J.L.P.; Urbina, J.A.; De Souza, W.; Rodrigues, J. In vitro antileishmanial activity of ravuconazole, a triazole antifungal drug, as a potential treatment for leishmaniasis. J. Antimicrob. Chemother. 2018, 73, 2360–2373. [Google Scholar] [CrossRef] [PubMed]
- Geary, W.J. The use of conductivity measurements in organic solvents for the characterisation of coordination compounds. Coord. Chem. Rev. 1971, 7, 81–122. [Google Scholar] [CrossRef]
- Hay, P.J.; Wadt, W.R. Ab initio effective core potentials for molecular calculations. Potentials for K to Au including the outermost core orbitals. J. Chem. Phys. 1985, 82, 299–310. [Google Scholar] [CrossRef]
- De Souza, W.; Rodrigues, J.C.F. Ultrastructural Alterations in Organelles of Parasitic Protozoa Induced by Different Classes of Metabolic Inhibitors. Curr. Pharm. Des. 2008, 14, 925–938. [Google Scholar]
- Rodrigues, C.O.; Catisti, R.; Uyemura, S.A.; Vercesi, A.E.; Lira, R.; Rodriguez, C.; Urbina, J.A.; Docampo, R. The Sterol Composition of Trypanosoma cruzi Changes After Growth in Different Culture Media and Results in Different Sensitivity to Digitonin-Permeabilization. J. Eukaryot. Microbiol. 2001, 48, 588–594. [Google Scholar] [CrossRef]
- Palmie-Peixoto, I.V.; Rocha, M.R.; Urbina, J.A.; de Souza, W.; Einecker-Lamas, M.; Motta, M.C. Effects of sterol biosynthesis inhibitors on endosymbiont-bearing trypanoso-matids. FEMS Microbiol. Lett. 2006, 255, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Mathur, R.; Das, R.P.; Ranjan, A.; Shaha, C. Elevated ergosterol protects Leishmania parasites against antimony-generated stress. FASEB J. 2015, 29, 4201–4213. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, S.; Xu, W.; Hsu, F.-F.; Patel, J.; Huang, J.; Zhang, K. Sterol methyltransferase is required for optimal mitochondrial function and virulence in Leishmania major. Mol. Microbiol. 2019, 111, 65–81. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sterol | RT (min) | Control | H3 3 μM | H3 5 μM | H3 7 μM |
|---|---|---|---|---|---|
| Cholesterol | 22.37 | 6.8 | 8.1 | 10.7 | 13.5 |
| Zymosterol (II) | 23.19 | 0.7 | 2.1 | 2.1 | 1.4 |
| Cholesta-5,8,24-trien-3β-ol (XI) | 23.40 | nd | 1.6 | 1.5 | 0.5 |
| Cholesta-5,7,24-trien-3β-ol (XIII) | 24.1 | 2.5 | 69.0 | 64.9 | 57.2 |
| Cholesta-7,24-dien-3β-ol (XII) | 24.3 | 1.2 | 12.0 | 11.9 | 12.0 |
| Ergosta-5,7,9(11)-22-tetraen-3β-ol (IX) | 24.6 | 0.7 | 1.1 | nd | nd |
| Ergosta-5,8,22-trien-3β-ol (VII) | 24.8 | 3.0 | nd | nd | nd |
| 14α-methyl-ergosta-8,24(24′)-dien-3β-ol (VI) | 25.0 | nd | 0.2 | nd | nd |
| Ergosta-5,7,24(24′)-trien-3β-ol (V) | 25.7 | 69.6 | 5.1 | 8.8 | 15.0 |
| Ergosta-7,24(24′)-dien-3β-ol (IV) | 26.0 | 11.2 | nd | nd | nd |
| Ergosta-5,7,9(11)-24(24′)-tetraen-3β-ol (VIII) | 26.2 | 1.4 | nd | nd | nd |
| Lanosterol (I) | 27.3 | 0.8 | 0.6 | nd | nd |
| Stigmasta-5,7,22-trien-3β-ol (X) | 28.3 | 2.0 | 0.2 | nd | nd |
| Sterol | RT (min) | Control | ZnH3 2 μM | ZnH3 5 μM | ZnH3 7 μM |
|---|---|---|---|---|---|
| Cholesterol | 22.37 | 7.1 | 8.9 | 17.6 | 16.7 |
| Zymosterol (II) | 23.18 | 0.80 | 2.3 | 2.7 | 2.5 |
| Cholesta-5,7,22-trien-3β-ol (XIV) | 23.34 | nd | 1.0 | 0.9 | 1.2 |
| Cholesta-5,8,24-trien-3β-ol (XI) | 23.40 | nd | 1.4 | 2.9 | 3.3 |
| Cholesta-5,7,24-dien-3β-ol (XIII) | 24.12 | nd | 54.4 | 40.0 | 37.6 |
| Ergosta-7,22,24(24′)-trien-3β-ol (XV) | 24.20 | 0.70 | nd | nd | nd |
| Cholesta-7,24-dien-3β-ol ((XII) | 24.36 | nd | 20.8 | 18.2 | 17.1 |
| Ergosta-5,7,22,24(24′)-tetraen-3β-ol (XVI) | 24.37 | 2.0 | nd | nd | nd |
| Ergosta-5,7,9(11)-22- tetraen-3β-ol (IX) | 24.56 | nd | 1.2 | 2.5 | nd |
| Ergosta-5,8,22-trien-3β-ol (VII) | 24.77 | 0.80 | nd | nd | nd |
| 14α-methyl-ergosta-8,24(24′)-dien-3ß-ol (VI) | 24.87 | 1.1 | 1.0 | 2.5 | 3.9 |
| Ergosta-5,7,24(24′)-trien-3β-ol (V) | 25.7 | 65.7 | 6.6 | 12.4 | 15.7 |
| Ergosta-7,24(24′)-dien-3β-ol (IV) | 26.0 | 14.1 | nd | nd | nd |
| Ergosta-5,7,9(11)-24(24′)-tetraen-3β-ol (VIII) | 26.2 | 2.1 | nd | nd | nd |
| Lanosterol (I) | 27.3 | 2.1 | 1.1 | 1.2 | nd |
| Stigmasta-5,7,22-trien-3β-ol (X) | 28.3 | 2.6 | nd | nd | nd |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Visbal, G.; Justo, R.M.S.; dos Santos da Silva e Miranda, G.; Teixeira de Macedo Silva, S.; de Souza, W.; Rodrigues, J.C.F.; Navarro, M. Zinc(II)-Sterol Hydrazone Complex as a Potent Anti-Leishmania Agent: Synthesis, Characterization, and Insight into Its Mechanism of Antiparasitic Action. Pharmaceutics 2023, 15, 1113. https://doi.org/10.3390/pharmaceutics15041113
Visbal G, Justo RMS, dos Santos da Silva e Miranda G, Teixeira de Macedo Silva S, de Souza W, Rodrigues JCF, Navarro M. Zinc(II)-Sterol Hydrazone Complex as a Potent Anti-Leishmania Agent: Synthesis, Characterization, and Insight into Its Mechanism of Antiparasitic Action. Pharmaceutics. 2023; 15(4):1113. https://doi.org/10.3390/pharmaceutics15041113
Chicago/Turabian StyleVisbal, Gonzalo, Rodrigo M. S. Justo, Gabrielle dos Santos da Silva e Miranda, Sara Teixeira de Macedo Silva, Wanderley de Souza, Juliany Cola Fernandes Rodrigues, and Maribel Navarro. 2023. "Zinc(II)-Sterol Hydrazone Complex as a Potent Anti-Leishmania Agent: Synthesis, Characterization, and Insight into Its Mechanism of Antiparasitic Action" Pharmaceutics 15, no. 4: 1113. https://doi.org/10.3390/pharmaceutics15041113
APA StyleVisbal, G., Justo, R. M. S., dos Santos da Silva e Miranda, G., Teixeira de Macedo Silva, S., de Souza, W., Rodrigues, J. C. F., & Navarro, M. (2023). Zinc(II)-Sterol Hydrazone Complex as a Potent Anti-Leishmania Agent: Synthesis, Characterization, and Insight into Its Mechanism of Antiparasitic Action. Pharmaceutics, 15(4), 1113. https://doi.org/10.3390/pharmaceutics15041113