
Salvianolic Acid B: A Review of Pharmacological Effects, Safety, Combination Therapy, New Dosage Forms, and Novel Drug Delivery Routes
Abstract
:1. Introduction
2. Pharmacological Effects
2.1. Effects on the Tumor
2.2. Effects on the Lung
2.3. Effects on the Cerebral Nerve

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pharmacology | Experimental Subject | Experimental Dose | Animal Experimental Drug Delivery Method | Effects | Ref. |
|---|---|---|---|---|---|
| Effects on the cerebral nerve | Oxygenose deprivation/reoxygenation of rat astrocytes | 10 μMol/L | None | Decrease: ROS Increase: p-STAT3, p-AKT, BDNF, IGF1α | [54] |
| Mice were infected with Porphyromonas gingivalis | 20, 40 mg/kg | Intraperitoneal injection | Decrease: IL-1β, IL-6, Aβ, bdnf, ngf, Beta-Site APP Cleaving Enzyme 1, γ-secretase, Receptor for advanced glycation end products, ROS, MDA Increase: Recombinant A Disintegrin, Metalloprotease 10, Low-density lipoprotein receptor-related protein 1, SOD, GSH-Px | [55] | |
| Rats with middle cerebral artery occlusion HUVECs under hypoxia | 10, 20 mg/kg 50 µg/mL | Intraperitoneal injection | Increase: VEGF, VEGFR2, STC1, mTOR, AKT | [56] | |
| Embolization of the left middle cerebral artery occlusion in mice Oxygen–glucose deprivation in mouse cerebral cortex astrocytes | 5, 10, 20, 40, 80, 100 μg/mL 10 mg/kg | Intraperitoneal injection | Decrease: pS2448-m TOR, pS757-ULK1, IL-2, IL-6, IFN-γ, IL-10 Increase: IL-4, Beclin1, pT172-AMPK, LC3B II/LCB I | [57] | |
| Oxygen–glucose-deprivation/reoxygenation-induced primary astrocytes from the cortex of newborn mice Mice with middle cerebral artery occlusion/reperfusion | 800 ng/mL 12 mg/kg | Intraperitoneal injection | Decrease: ROS, LDH Increase: Glucose-6-phosphate, NADPH, GSH, Glycogen phosphorylase activity | [58] | |
| H4 neuroglioma cells from human Bacterial Artificial Chromosome transgenic mice | 50 μM 10 mg/kg | Intraperitoneal injection | Decrease: α-synuclein Increase: LAMP-1, LAMP-2A, LC3-II | [59] | |
| MPTP induced in mice | 6.7 mL/kg | Intraperitoneal injection | Decrease: Calprotectin, TNF-α, TLR4, MyD88, NuclearNF-κB p65, p-NF-κB p65 Increase: IL-1β, Tyrosine hydroxylase, Colonic tight junction protein | [60] | |
| A high-fat diet combined with chronic mild stress-induced mice. Stimulation involved water and food grabbing, 45° tilt of the cage, reversal of circadian rhythm, wetting of the cage horizontal shaking, walking on ice, empty cage, flash screen, tail pinning, and restraint. Palmitic acid binding to LPS stimulates primary microglia. | 10, 20, 40, 60, 80, 100, 200 μM 20 mg/kg | Intraperitoneal injection | Decrease: IL-6, IL-1β, TNF-α, iNOS, CD11b, Caspase3 Increase: NeuN+ | [61] | |
| Rats undergoing without repeating the same stress procedure for two consecutive days, one stress/day, including with cage tilting at 45 °C, wet caging or damp bedding caging, cold water forced swimming, food deprivation, water deprivation, foot shock, tail pinching and suspension, behavior restriction, stroboscopic illumination. | 20, 40 mg/kg | Intraperitoneal injection | Decrease: IL-6, IL-1β, TNF-α, MDA, ASC, cleaved caspase-1 Increase: CAT, SOD, GPx, Sucrose intake | [62] | |
| Rats with food deprivation followed by water deprivation, 45° cage tilting, restraint in an empty water bottle, noise, tail clamping, and damp bedding. | 30 mg/kg | Intraperitoneal injection | Decrease: IL-6, IL-1β, TNF-α, NF-κB, 4-HNE, MDA, CHOP, GRP78, Microglia density of Iba-1-positive cells in the hippocampus Increase: CAT, Nrf2, NQO1, HO-1, pAMPK/AMPK, SIRT1 | [63] | |
| H2O2-induced PC-12 cells | 0, 1, 5, 10 μL | None | Decrease: Bax, cleaved caspase-3, cleaved caspase-9 Increase: Proliferating Cell Nuclear Antigen, Cyclin A, Cyclin E1, Cyclin-Dependent Kinase 2 (CDK2), Cyclin D1, CDK4, Bcl-2, miR-26a, Phosphorylation of PI3K, Phosphorylation of AKT, Phosphorylation of GSK3β | [65] | |
| Rats’ total laminectomy of the T9 vertebral plate. A Kirschner wire (10 g) was inserted into the aorta via a catheter with a weight that fell freely. Following this, a semicircular slice made from thin plastic was used to impact the spinal cord, and the wire was immediately removed, resulting in the incomplete injury of the rat spinal cord. | 2, 10, 20 mg/kg | Intraperitoneal injection | Decrease: Bax, cleaved caspase-3, cleaved caspase-9 Increase: Bcl-2, Lymphatic enhancer factor-1 antibody, Transcription factor-1, Phosphorylation of GSK3β, β-catenin | [66] | |
| Mice’s exposed T9, T10, and T11 vertebrae were struck with a striking rod carrying a 10 g counterweight | 30, 60 mg/kg | Gavage administration | Decrease: TLR4, MyD88, TNF-a, Substance P | [67] |
2.4. Effects on the Heart
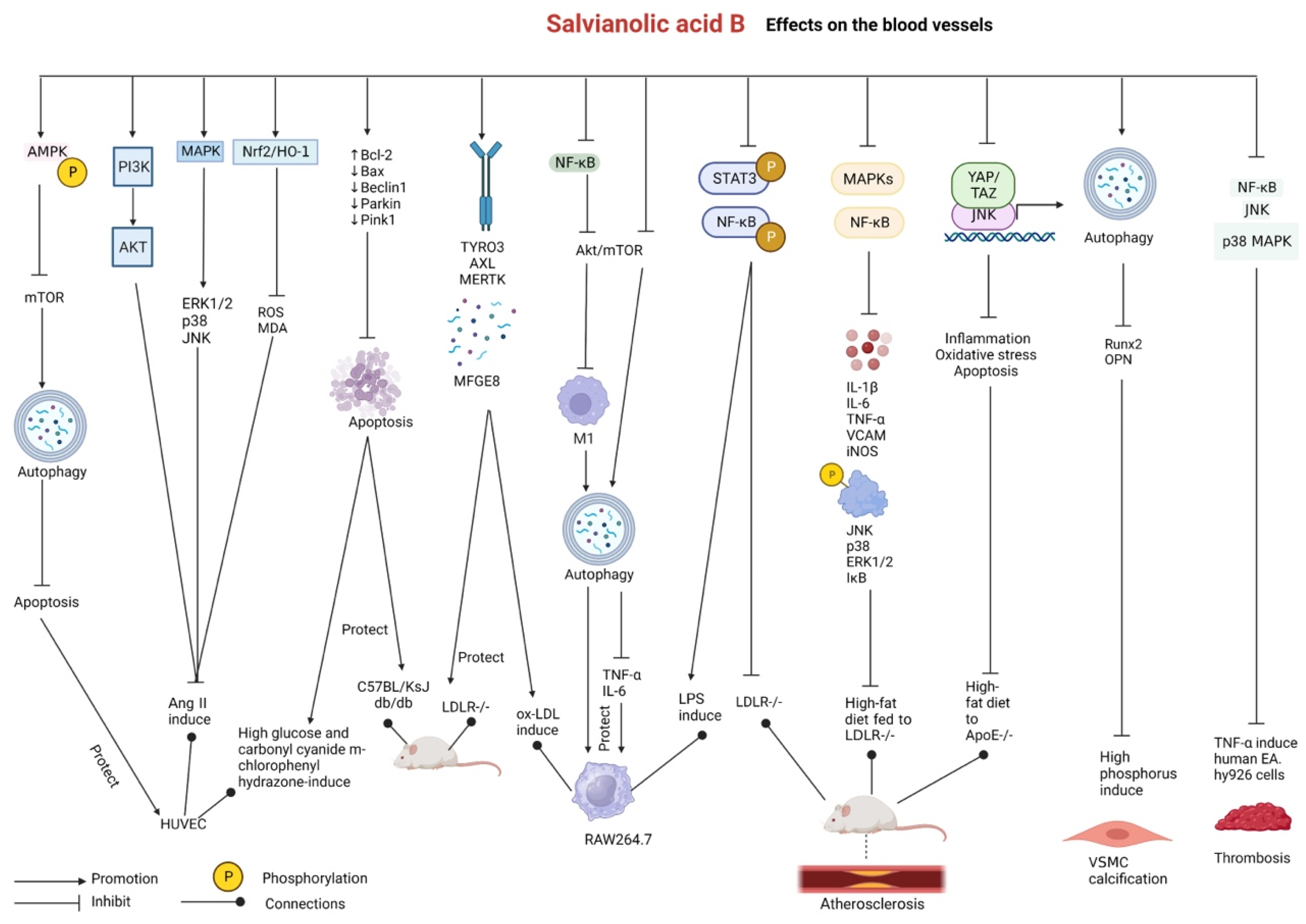
2.5. Effects on Blood Vessels
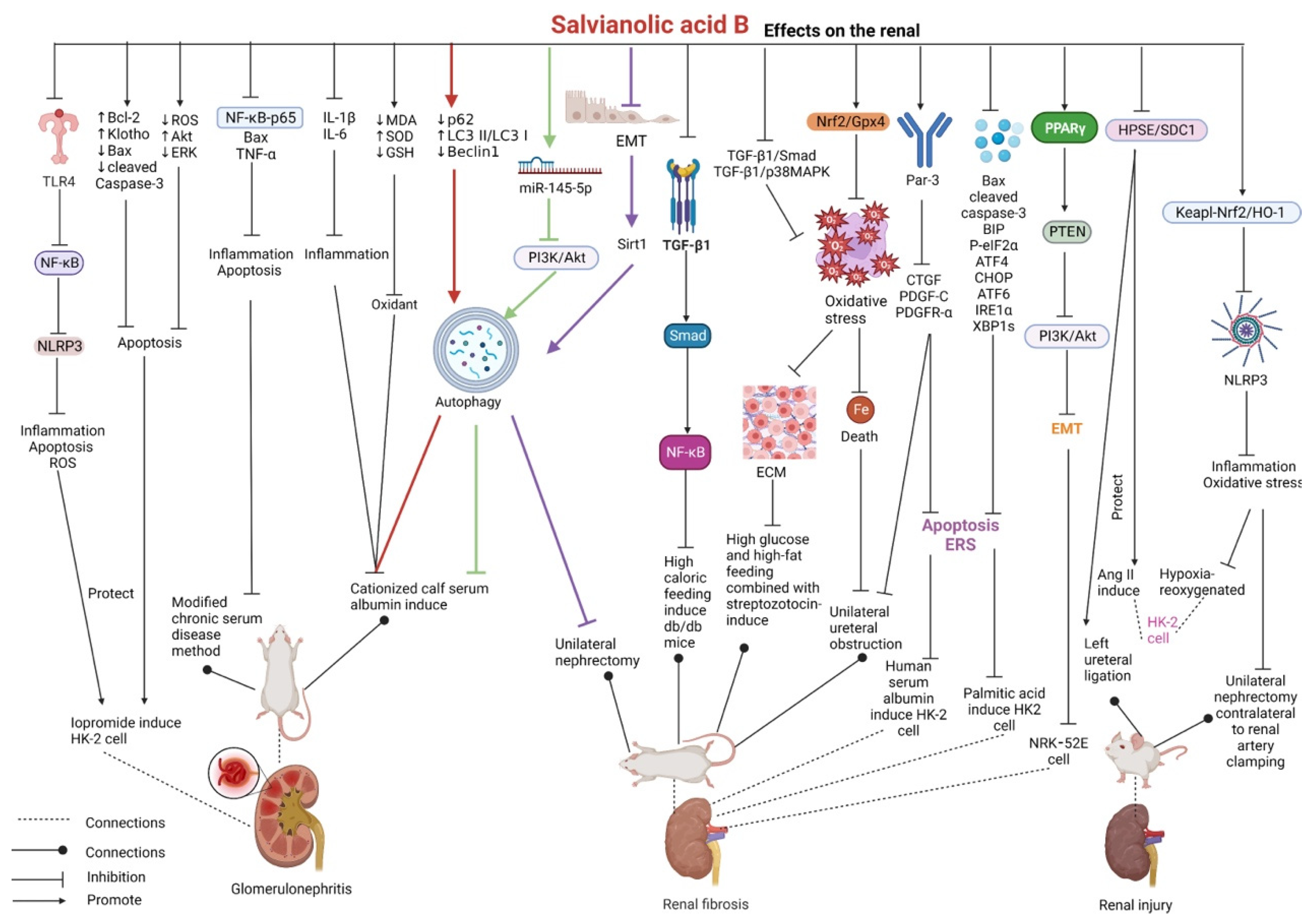
2.6. Effects on the Kidney
2.7. Effects on the Intestine
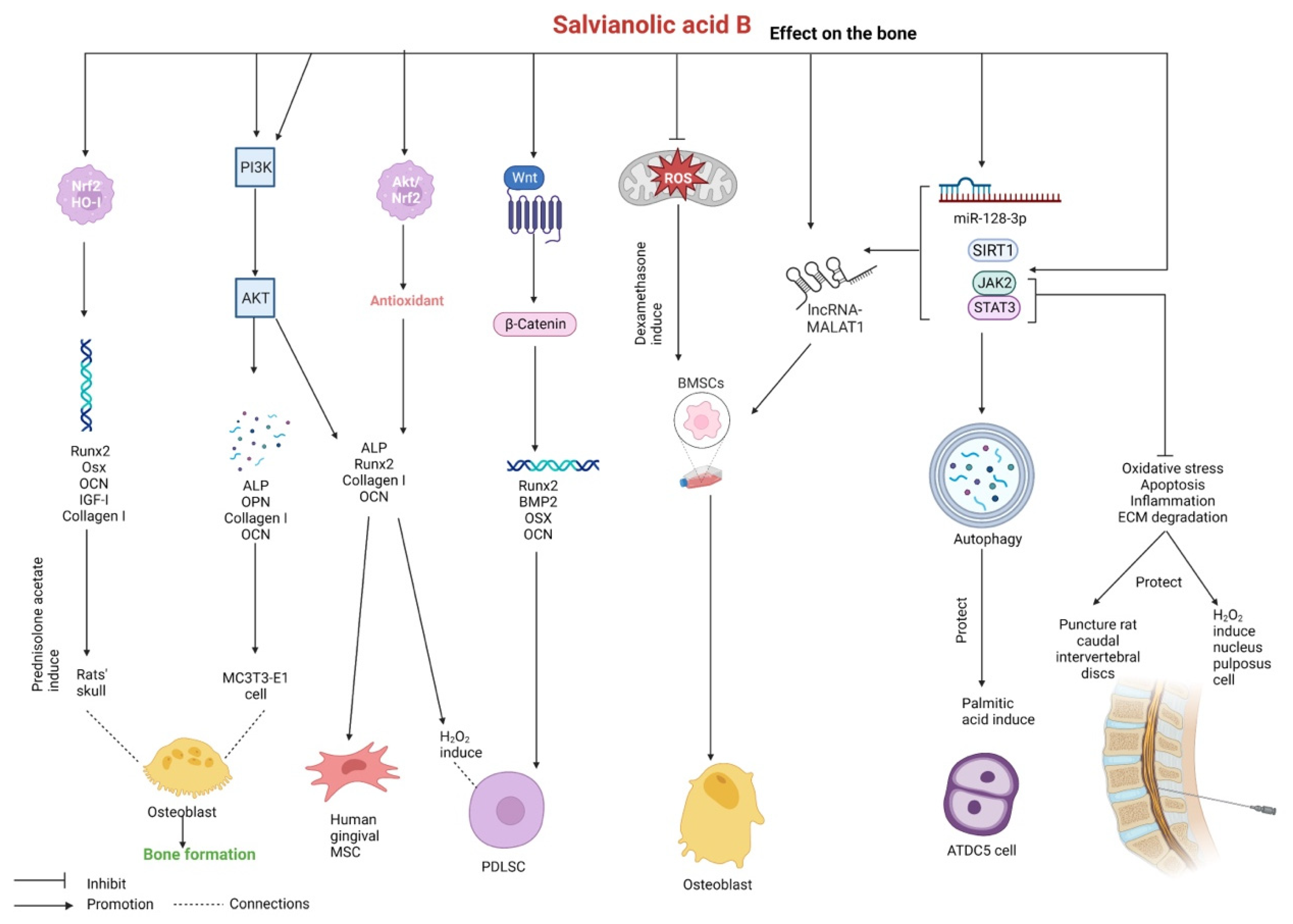
2.8. Effects on the Bone
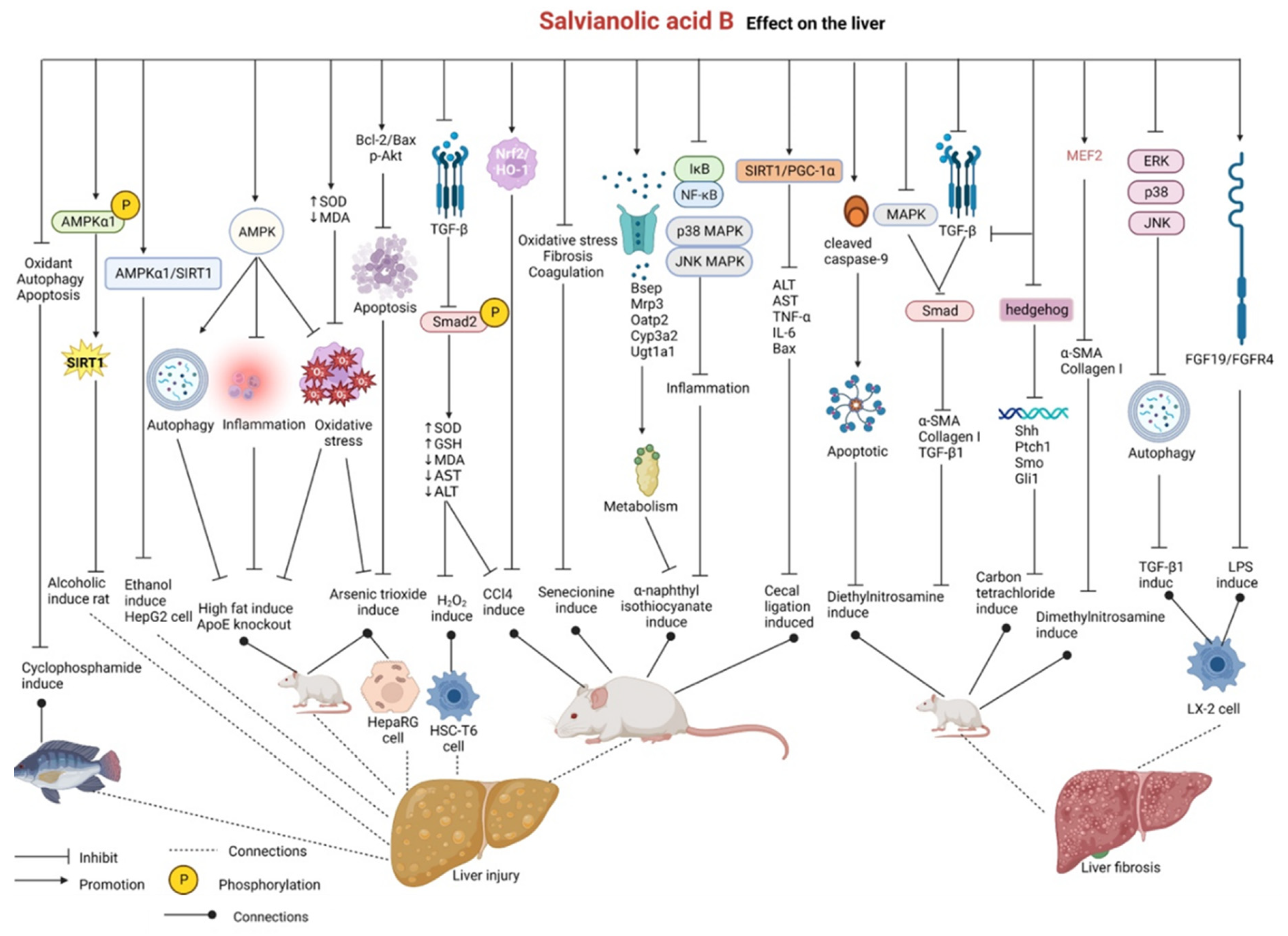
2.9. Effects on the Liver
2.10. Effects on the Skin
2.11. Other Effects
| Pharmacology | Experimental Subject | Experimental Dose | Animal Experimental Drug Delivery Method | Effects | Ref. |
|---|---|---|---|---|---|
| Protect the retina | High-glucose-induced primary Müller cells in the rat retina | 40 μMol/L | None | Decrease: Bax Increase: Beclin1, LC3-II, p-AMPK, Bcl-2 | [171] |
| Improve the fat transplantation survival | Human-adipose-derived stem cells 3T3-L1 Nude mouse fat transplantation | 10, 50, 100 μMol/L 10, 50 μMol/L | None | Increase: Cell proliferation, Adipogenic differentiation | [172] |
| Protect the testicles | Rat testicular ischemia/reperfusion | 10 mg/kg | Intravenous injection | Decrease: MDA Increase: Testicular weight, Seminiferous tubular diameter, Germ cell layer number | [173] |
| Improve crush syndrome | Comprised anesthetized rats with bilateral hindlimb compression by a rubber tourniquet | 10, 20, 50 mg/kg | Intravenous injection | Decrease: IL-6, Cyt c, IL-1β, HMGB1, TNF-α, Nox Increase: SOD | [174] |
| Prevent cystine stones | Cystine-induced HK-2 cell Slc7a9 knockout mice | 500 ug/L 60 mg/kg | Oral administration | Decrease: MDA, Cleavage of the eukaryotic eIF4G1 protein, Enterovirus 71 capsid protein viral polyproteins 1 Increase: SOD, GPx, Heat shock protein 90, Catalase | [175] |
| Treat hand, foot, and mouth disease | Enterovirus 71-induced HeLa cells | 100, 10 μg/mL | None | Increase: Bcl-2, Cyclin-D1 | [176] |
| Protect the airway | PM2.5-induced mice PM2.5-induced human bronchial epithelial 16HBE cells | 2.5, 7.5, 15 mg/mL 2.5, 5, 10 μM | Nebulized inhalation | Decrease: MyD88, IL-1β, TNF-α, KC, TGF-β1, TLR4, TRAF-6, NLRP3, ROS, Phosphorylation-ERK1/2, Phosphorylation-p38 Increase: SOD, CAT, GSH, GSH-Px | [177] |
| Inhibit obesity | High-fat-diet-induced mice | 100 mg/kg | Gavage administration | Decrease: Sterol regulatory element-binding protein 1c, Fatty acid synthase 1, Diacylglycerol O-acyltransferase 2, Insulin-induced gene 1 protein, Insulin-induced gene 2 protein, 3-hydroxy-3 methylglutaryl coenzyme A reductase, TG, TC, LDL, ALT, AST, Gram-negative Desulfovibrionacea, Helicobacteraceae, Deferribacteraceae, Mucispirillum, Odoribacter, TLR4, MYD88 Increase: HDL, Lactobacillaceae, Adlercreutzia, Bifidobacterium | [178] |
| Enhance immune system | CBA/J females mated with DBA/2J males as a spontaneous abortion mouse model | 100 mg/kg | No mention | Decrease: TLR-2, TLR-4, NF-κB, IFN-γ, TNF-α, Nkp46, CD8+T Increase: Placental labyrinth area, SOD, Nrf2 | [181] |
| Mice received γ-irradiation | 5, 12.5, 20 mg/kg | Intraperitoneal injection | Decrease: MDA, Bach1, ROS, Bax Increase: Peripheral white blood cells, Red blood cells, Platelets | [182] | |
| Accelerate wound healing | Human gingival fibroblasts | 25, 50, 75, 100, 150 µg/mL | None | Increase: Cell viability, Wound healing, Collagen III | [183] |
| Anticonvulsant | Pentylenetetrazole-induced rats | 20 mg/kg | Oral administration | Decrease: Bax, cleaved caspase-3 Increase: Bcl-2 | [184] |
3. Safety
4. Combination Therapy
5. New Dosage Forms and Drug Delivery Routes
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Liu, A.H.; Li, L.; Xu, M.; Lin, Y.H.; Guo, H.Z.; Guo, D.A. Simultaneous quantification of six major phenolic acids in the roots of Salvia miltiorrhiza and four related traditional Chinese medicinal preparations by HPLC-DAD method. J. Pharm. Biomed. Anal. 2006, 41, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.; Su, S.; Xiang, X.; Sha, X.; Zhu, Z.; Wang, Y.; Guo, S.; Yan, H.; Qian, D.; Duan, J. Comparative Analysis of the Major Chemical Constituents in Salvia miltiorrhiza Roots, Stems, Leaves and Flowers during Different Growth Periods by UPLC-TQ-MS/MS and HPLC-ELSD Methods. Molecules 2017, 22, 771. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Wei, Y.J.; Qi, L.W.; Li, P.; Qian, Z.M.; Luo, H.W.; Chen, J.; Zhao, J. Determination of fifteen bioactive components in Radix et Rhizoma Salviae Miltiorrhizae by high-performance liquid chromatography with ultraviolet and mass spectrometric detection. Biomed. Chromatogr. 2008, 22, 164–172. [Google Scholar] [CrossRef] [PubMed]
- Li, H.B.; Lai, J.P.; Jiang, Y.; Chen, F. Preparative isolation and purification of salvianolic acid B from the Chinese medicinal plant Salvia miltiorrhiza by high-speed counter-current chromatography. J. Chromatogr. A 2002, 943, 235–239. [Google Scholar] [CrossRef]
- Zhang, M.; Ignatova, S.; Liang, Q.; Wu Jun, F.; Sutherland, I.; Wang, Y.; Luo, G. Rapid and high-throughput purification of salvianolic acid B from Salvia miltiorrhiza Bunge by high-performance counter-current chromatography. J. Chromatogr. A 2009, 1216, 3869–3873. [Google Scholar] [CrossRef]
- Zhou, Y.S.; Xian, Y.K.; Meng, J.; Duan, F.; Liu, L. Study on the Preparative Purification of Salvianolie Acid from Salvia Mihiorrhiza. China Pharm. 2009, 12, 697–698. [Google Scholar]
- Kan, S.; Li, J.; Huang, W.; Shao, L.; Chen, D. Microsphere resin chromatography combined with microbial biotransformation for the separation and purification of salvianolic acid B in aqueous extract of roots of Salvia multiorrihza Bunge. J. Chromatogr. A 2009, 1216, 3881–3886. [Google Scholar] [CrossRef]
- Meng, J.; Zhou, Y.S.; Xian, Y.K.; Duan, F.; Liu, L. Study on the Extraction Procedure for the Salvianolic Acids of Salviamiltiorhiza. Lishizhen Med. Mater. Medica Res. 2009, 20, 2524–2525. [Google Scholar]
- Dong, J.; Liu, Y.; Liang, Z.; Wang, W. Investigation on ultrasound-assisted extraction of salvianolic acid B from Salvia miltiorrhiza root. Ultrason. Sonochem. 2010, 17, 61–65. [Google Scholar] [CrossRef]
- Liu, J.D.; Ye, Z.L.; Li, D.K.; Sun, J.Q.; Peng, F.; Yue, H.S.; Zhou, D.Z. Study on the application of ultrafiltration method in the process of injectable danshen polyphenolic acid preparation. Shaanxi J. Tradit. Chin. Med. 2012, 33, 231–233. [Google Scholar]
- Lai, X.J.; Liu, H.Q.; Li, J.S.; Di, L.Q.; Cai, B.C. Intestinal absorption properties of three components in salvianolic acid extract and the effect of borneol on their absorption in rats. Acta Pharm. Sin. 2010, 45, 1576–1581. [Google Scholar]
- Hou, M.N.; Gu, X.; Xie, Z.; Lou, Y.; Zhao, M.J.; Zheng, J.Q.; Zheng, G.G. Study on Metabolites and Metabolic Pathway of Salvianolic Acid B in Rats. Chin. J. Mod. Appl. Pharm. 2019, 36, 2673–2682. [Google Scholar]
- Ma, B.B.; Lou, T.Y.; Liang, Y.Y.; Wang, T.T.; Li, R.J.; Liu, J.H.; Wang, C.X.; Yu, S.Y.; Guo, Y.D.; Wang, J. Analysis and comparison of metabolic processes of salvianolic acid A and salvianolic acid B in rats based on UHPLC-LTQ-Orbitrap MS technology. China J. Chin. Mater. Medica 2021, 46, 2276–2286. [Google Scholar]
- Wu, Y.T.; Chen, Y.F.; Hsieh, Y.J.; Jaw, I.; Shiao, M.S.; Tsai, T.H. Bioavailability of salvianolic acid B in conscious and freely moving rats. Int. J. Pharm. 2006, 326, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Gao, D.Y.; Han, L.M.; Zhang, L.H.; Fang, X.L.; Wang, J.X. Bioavailability of salvianolic acid B and effect on blood viscosities after oral administration of salvianolic acids in beagle dogs. Arch. Pharm. Res. 2009, 32, 773–779. [Google Scholar] [CrossRef]
- Liu, J. Study on the Apparent Mechanism of Salvianolic Acid B Combined With Stem Cells in the Treatment of Acute Liver Failure; Guangzhou University of Chinese Medicine: Guangzhou, China, 2021. [Google Scholar]
- Zhao, J.; Yang, X.C.; Fujino, M.; Ichimaru, N.; Que, W.; Li, X.K.; Takahara, S. Salvianolic acid B ameliorates liver injury in a murine aGvHD model by decreasing inflammatory responses via upregulation of HO-1. Transpl. Immunol. 2019, 55, 101203. [Google Scholar] [CrossRef]
- Li, S.; Song, F.; Lei, X.; Li, J.; Li, F.; Tan, H. hsa_circ_0004018 suppresses the progression of liver fibrosis through regulating the hsa-miR-660-3p/TEP1 axis. Aging 2020, 12, 11517–11529. [Google Scholar] [CrossRef]
- Pang, Y.; Zhang, P.C.; Lu, R.R.; Li, H.L.; Li, J.C.; Fu, H.X.; Cao, Y.W.; Fang, G.X.; Liu, B.H.; Wu, B.; et al. Andrade-Oliveira Salvianolic Acid B Modulates Caspase-1-Mediated Pyroptosis in Renal Ischemia-Reperfusion Injury via Nrf2 Pathway. Front. Pharmacol. 2020, 11, 541426. [Google Scholar] [CrossRef]
- Lee, H.G.; Kwon, S.; Moon, S.K.; Cho, S.Y.; Park, S.U.; Jung, W.S.; Park, J.M.; Ko, C.N.; Cho, K.H. Neuroprotective Effects of Geopung-Chunghyuldan Based on Its Salvianolic Acid B Content Using an In Vivo Stroke Model. Curr. Issues Mol. Biol. 2023, 45, 1613–1626. [Google Scholar] [CrossRef]
- Ma, D.; Mandour, A.S.; Yoshida, T.; Matsuura, K.; Shimada, K.; Kitpipatkun, P.; Uemura, A.; Ifuku, M.; Takahashi, K.; Tanaka, R. Intraventricular pressure gradients change during the development of left ventricular hypertrophy: Effect of salvianolic acid B and beta-blocker. Ultrasound 2021, 29, 229–240. [Google Scholar] [CrossRef]
- Nandi, S.S.; Katsurada, K.; Sharma, N.M.; Anderson, D.R.; Mahata, S.K.; Patel, K.P. MMP9 inhibition increases autophagic flux in chronic heart failure. Am. J. Physiol. Heart Circ. Physiol. 2020, 319, H1414–H1437. [Google Scholar] [CrossRef] [PubMed]
- Griffin, M.F.; Borrelli, M.R.; Garcia, J.T.; Januszyk, M.; King, M.; Lerbs, T.; Cui, L.; Moore, A.L.; Shen, A.H.; Mascharak, S.; et al. JUN promotes hypertrophic skin scarring via CD36 in preclinical in vitro and in vivo models. Sci. Transl. Med. 2021, 13, eabb3312. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Ji, J.; Zhang, D.; Ma, M.; Sun, L. Protective effects and potential mechanism of salvianolic acid B on sodium laurate-induced thromboangiitis obliterans in rats. Phytomedicine 2020, 66, 153110. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Wei, Z.; He, S.; Bi, Y.; Cao, Y.; Wang, W. Mesoporous Bioactive Glass Scaffold Delivers Salvianolic Acid B to Promote Bone Regeneration in a Rat Cranial Defect Model. Curr. Drug Deliv. 2021, 18, 323–333. [Google Scholar] [CrossRef]
- Wang, N.; Li, Y.; Li, Z.; Liu, C.; Xue, P. Sal B targets TAZ to facilitate osteogenesis and reduce adipogenesis through MEK-ERK pathway. J. Cell Mol. Med. 2019, 23, 3683–3695. [Google Scholar] [CrossRef]
- Jiang, L.; Li, Y.; Yu, J.; Wang, J.; Ju, J.; Dai, J. A dry powder inhalable formulation of salvianolic acids for the treatment of pulmonary fibrosis: Safety, lung deposition, and pharmacokinetic study. Drug Deliv. Transl. Res. 2021, 11, 1958–1968. [Google Scholar] [CrossRef]
- Gong, Y.; Li, D.; Li, L.; Yang, J.; Ding, H.; Zhang, C.; Wen, G.; Wu, C.; Fang, Z.; Hou, S.; et al. Smad3 C-terminal phosphorylation site mutation attenuates the hepatoprotective effect of salvianolic acid B against hepatocarcinogenesis. Food Chem. Toxicol. 2021, 147, 111912. [Google Scholar] [CrossRef]
- Ko, Y.S.; Jin, H.; Park, S.W.; Kim, H.J. Salvianolic acid B protects against oxLDL-induced endothelial dysfunction under high-glucose conditions by downregulating ROCK1-mediated mitophagy and apoptosis. Biochem. Pharmacol. 2020, 174, 113815. [Google Scholar] [CrossRef]
- Wang, Q.Q.; Zhai, C.; Wahafu, A.; Zhu, Y.T.; Liu, Y.H.; Sun, L.Q. Salvianolic acid B inhibits the development of diabetic peripheral neuropathy by suppressing autophagy and apoptosis. J. Pharm. Pharmacol. 2019, 71, 417–428. [Google Scholar] [CrossRef]
- Shi, Y.; Pan, D.; Yan, L.; Chen, H.; Zhang, X.; Yuan, J.; Mu, B. Salvianolic acid B improved insulin resistance through suppression of hepatic ER stress in ob/ob mice. Biochem. Biophys. Res. Commun. 2020, 526, 733–737. [Google Scholar] [CrossRef]
- Zhai, J.; Tao, L.; Zhang, Y.; Gao, H.; Qu, X.; Song, Y.; Zhang, S. Salvianolic Acid B Attenuates Apoptosis of HUVEC Cells Treated with High Glucose or High Fat via Sirt1 Activation. Evid. Based Complement. Altern. Med. 2019, 2019, 9846325. [Google Scholar] [CrossRef] [PubMed]
- Qu, H.T.; Zhai, X.J.; Shao, Q.; Cheng, Y.Y. Simultaneous Determination of Seven Active Compounds in Radix Salviae Miltiorrhizae by Temperature-Controlled Ultrasound-Assisted Extraction and HPLC. Chromatographia 2007, 66, 21–27. [Google Scholar] [CrossRef]
- Kuang, Z.A.; Dong, J.W.; Sun, C.; Yin, M.X.; Liu, L.; Deng, H.B.; Liu, X.J.; Feng, Y.C. Salvianolic acid B exerts its anti-tumor immunity by targeting USP2 and reducing the PD-L1 level. Acta Pharm. Sin. 2023, 58, 954–962. [Google Scholar]
- Zhang, L.; Zheng, L.; Yuan, N.P.; Gu, L.Q.; Shen, Q.Y. Inhibition effect and mechanism of the salvianolic acid B on the proliferation of human colon cancer HCT116 cells. Chin. J. Clin. Pharmacol. 2020, 36, 3268–3271. [Google Scholar]
- Tsai, J.H.; Donaher, J.L.; Murphy, D.A.; Chau, S.; Yang, J. Spatiotemporal regulation of epithelial-mesenchymal transition is essential for squamous cell carcinoma metastasis. Cancer Cell 2012, 22, 725–736. [Google Scholar] [CrossRef]
- Han, G.; Wang, Y.; Liu, T.; Gao, J.; Duan, F.; Chen, M.; Yang, Y.; Wu, C. Salvianolic acid B acts against non-small cell lung cancer A549 cells via inactivation of the MAPK and Smad2/3 signaling pathways. Mol. Med. Rep. 2022, 25, 184. [Google Scholar] [CrossRef]
- Zhang, H.; Tang, J.; Cao, Y.; Wang, T. Salvianolic Acid B Suppresses Non-Small-Cell Lung Cancer Metastasis through PKM2-Independent Metabolic Reprogramming. Evid. Based Complement. Altern. Med. 2022, 2022, 9302403. [Google Scholar] [CrossRef]
- Wang, J.; Ma, Y.; Guo, M.; Yang, H.; Guan, X. Salvianolic acid B suppresses EMT and apoptosis to lessen drug resistance through AKT/mTOR in gastric cancer cells. Cytotechnology 2021, 73, 49–61. [Google Scholar] [CrossRef]
- Teng, M.; Hu, C.; Yang, B.; Xiao, W.; Zhou, Q.; Li, Y.; Li, Z. Salvianolic acid B targets mortalin and inhibits the migration and invasion of hepatocellular carcinoma via the RECK/STAT3 pathway. Cancer Cell Int. 2021, 21, 654. [Google Scholar] [CrossRef]
- Chen, L.; Hu, H.J.; Fang, Z.D.; Wang, X.T.; Sun, X.J.; Ge, X.B.; Cheng, C. Salvianolic acid B inhibits rat thoracic aortic aneurysm through JAK2/ STAT3 signaling pathway. J. Chin. Med. Mater. 2021, 44, 1971–1975. [Google Scholar]
- Yuan, P. The Investigation of the Anti-Metastatic Properties of Salvianolic Acid b on Breast Cancer Cells and Its Mechanisms Based on the Interaction between Platelets and Tumor Cells; Jiangxi University of Chinese Medicine: Nanchang, China, 2022. [Google Scholar]
- Ianni, A.; Ruggeri, P.; Bellio, P.; Martino, F.; Celenza, G.; Martino, G.; Franceschini, N. Salvianolic Acid B Strikes Back: New Evidence in the Modulation of Expression and Activity of Matrix Metalloproteinase 9 in MDA-MB-231 Human Breast Cancer Cells. Molecules 2022, 27, 8514. [Google Scholar] [CrossRef] [PubMed]
- Qian, C.A. Study on Exploring the Mechanism of Salvianolic acid B Modulating Tumor Angiogenesis and Metastasis Based on PDGF-B/PDGFR-β Signaling Axis; Nanjing University Of Chinese Medicine: Nanjing, China, 2021. [Google Scholar]
- Katary, M.A.; Abdelsayed, R.; Alhashim, A.; Abdelhasib, M.; Elmarakby, A.A. Salvianolic Acid B Slows the Progression of Breast Cancer Cell Growth via Enhancement of Apoptosis and Reduction of Oxidative Stress, Inflammation and Angiogenesis. Int. J. Mol. Sci. 2019, 20, 5653. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, Q.Q.; Gao, Z.H.; Li, L.; Gao, P.F.; Chen, B.B. Salvianolic acid B inducing autophagic death and senescence of endometrial cancer MFE-280 cells by inhibiting AKT phosphorylation. Chin. J. Immunol. 2022, 38, 671–675. [Google Scholar]
- Yu, J.J. Investigation of the Anti-Tumor Effect of Salvianolic Acid B and Its Mechanism Based on the Effect of Platelets on Tumor Cells; Jiangxi University of Chinese Medicine: Nanchang, China, 2020. [Google Scholar]
- Xiong, L. Study on the Mechanism of Antitumor Effect of Salvianolic Acid B by Inhibiting the Interaction between Platelets and Ovarian Cancer Cells; Jiangxi University of Chinese Medicine: Nanchang, China, 2021. [Google Scholar]
- Begum, G.; Singh, N.D.; Leishangthem, G.D.; Banga, H.S. Amelioration of bleomycin induced pulmonary fibrosis by administration of Salvianolic acid B in mice. Vet. Ital. 2022, 58, 87–101. [Google Scholar]
- Li, J.L.; Zhang, H.J.; Gao, Y.H.; Hou, H.P.; Li, H.; Chen, T.F.; Ma, L.N.; Ye, Z.G.; Zhang, G.P. Effect of Effective Components of Salvia Miltiorrhiza on TGF-β1 Induced Interstitial Transformation of Pulmonary Epithelial Cells. Chin. J. Exp. Tradit. Med. Formulae 2020, 26, 54–59. [Google Scholar]
- Jiang, L.; Wang, J.; Ju, J.; Dai, J. Salvianolic acid B and sodium tanshinone II A sulfonate prevent pulmonary fibrosis through anti-inflammatory and anti-fibrotic process. Eur. J. Pharmacol. 2020, 883, 173352. [Google Scholar] [CrossRef]
- Zhang, T.; Liu, M.; Gao, Y.; Li, H.; Song, L.; Hou, H.; Chen, T.; Ma, L.; Zhang, G.; Ye, Z. Salvianolic acid B inhalation solution enhances antifibrotic and anticoagulant effects in a rat model of pulmonary fibrosis. Biomed. Pharmacother. 2021, 138, 111475. [Google Scholar] [CrossRef]
- Li, J.C.; Feng, B. Effects of Salvianolic acid B on lung tissue repair and cerebral nerve protection in mouse pups with Staphylococcus aureus pneumonia. J. Zhengzhou Univ. (Med. Sci.) 2021, 56, 102–108. [Google Scholar]
- Yang, J.; Cao, Y.S.; Xu, Y.; Du, X.Y.; Zhang, T.; Guo, L.C.; Yuan, Q.; Chai, L.J.; Hu, L.M. The protective effects of salvianolic acid B, ginsenoside Rg1 and notoginsenoside R1 on oxygen-glucose deprivation/reoxygenation and reoxygenation of astrocytes. Chin. Pharmacol. Bull. 2022, 28, 1466–1472. [Google Scholar]
- Liu, J.; Wang, Y.; Guo, J.; Sun, J.; Sun, Q. Salvianolic Acid B improves cognitive impairment by inhibiting neuroinflammation and decreasing Aβ level in Porphyromonas gingivalis-infected mice. Aging 2020, 12, 10117–10128. [Google Scholar] [CrossRef]
- Bi, S.J.; Dong, X.Y.; Wang, Z.Y.; Fu, S.J.; Li, C.L.; Wang, Z.Y.; Xie, F.; Chen, X.Y.; Xu, H.; Cai, X.J.; et al. Salvianolic acid B alleviates neurological injury by upregulating stanniocalcin 1 expression. Ann. Transl. Med. 2022, 10, 739. [Google Scholar] [CrossRef] [PubMed]
- Xin, M.Y. Protection of Autophagy in Ischemic Brain Injury and the Neuroprotective Mechanism of Salvianolic Acid B Base on AMPK/m TOR/ULK1 Pathway; Jilin University: Changchun, China, 2020. [Google Scholar]
- Guo, H.; Zhang, Z.; Gu, T.; Yu, D.; Shi, Y.; Gao, Z.; Wang, Z.; Liu, W.; Fan, Z.; Hou, W.; et al. Astrocytic glycogen mobilization participates in salvianolic acid B-mediated neuroprotection against reperfusion injury after ischemic stroke. Exp. Neurol. 2022, 349, 113966. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.Z.; Ardah, M.; Haikal, C.; Svanbergsson, A.; Diepenbroek, M.; Vaikath, N.N.; Li, W.; Wang, Z.Y.; Outeiro, T.F.; El-Agnaf, O.M.; et al. Dihydromyricetin and Salvianolic acid B inhibit alpha-synuclein aggregation and enhance chaperone-mediated autophagy. Transl. Neurodegener. 2019, 8, 18. [Google Scholar] [CrossRef]
- Nan, Y.Q. Neuroprotective Effect of Salvianolic Acid B on Parkinson’s Disease Model Mice through Gut-Brain Axis Pathway; Lanzhou University: Lanzhou, China, 2022. [Google Scholar]
- Hu, N. Salvianolic Acid B Alleviates Depressive-like Behaviors in High-Fat-Fed and Stress-Treated Mice through the RAGE-DIAPH1 Pathway; University of Electronic Science and Technology of China: Chengdu, China, 2022. [Google Scholar]
- Huang, Q.; Ye, X.; Wang, L.; Pan, J. Salvianolic acid B abolished chronic mild stress-induced depression through suppressing oxidative stress and neuro-inflammation via regulating NLRP3 inflammasome activation. J. Food Biochem. 2019, 43, e12742. [Google Scholar] [CrossRef]
- Liao, D.; Chen, Y.; Guo, Y.; Wang, C.; Liu, N.; Gong, Q.; Fu, Y.; Fu, Y.; Cao, L.; Yao, D.; et al. Salvianolic Acid B Improves Chronic Mild Stress-Induced Depressive Behaviors in Rats: Involvement of AMPK/SIRT1 Signaling Pathway. J. Inflamm. Res. 2020, 13, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Gong, L.; Gu, Y.; Han, X.; Luan, C.; Liu, C.; Wang, X.; Sun, Y.; Zheng, M.; Fang, M.; Yang, S.; et al. Spatiotemporal Dynamics of the Molecular Expression Pattern and Intercellular Interactions in the Glial Scar Response to Spinal Cord Injury. Neurosci. Bull. 2023, 39, 213–244. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Fan, M. Protective functions of salvianolic acid B in PC-12 cells against hydrogen peroxide-triggered damage by mediation of microRNA-26a. Artif. Cells Nanomed. Biotechnol. 2019, 47, 4030–4037. [Google Scholar] [CrossRef]
- Zhou, H.; Liu, Y.; Sun, L.; Fu, M.; Zhao, Y. Salvianolic acid B activates Wnt/β-catenin signaling following spinal cord injury. Exp. Ther. Med. 2020, 19, 825–832. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, X.; Hu, P.; Jia, N.; Ji, S.; Yuan, H. Effect of Toll-Like Receptor 4/Myeloid Differentiation Factor 88 Inhibition by Salvianolic Acid B on Neuropathic Pain After Spinal Cord Injury in Mice. World Neurosurg. 2019, 13, e529–e534. [Google Scholar] [CrossRef]
- Zhao, M.H.; Feng, Y.N.; Yin, Y.W.; Li, C.Q.; Sun, G.J. Effect of Salvianolic Acid B on Antioxidant Capacity and Intestinal Microflora in Mice. Food Sci. 2020, 41, 105–112. [Google Scholar]
- Ji, Q.Q.; Li, Y.J.; Wang, Y.H.; Wang, Z.; Fang, L.; Shen, L.; Lu, Y.Q.; Shen, L.H.; He, B. Salvianolic Acid B Improves Postresuscitation Myocardial and Cerebral Outcomes in a Murine Model of Cardiac Arrest: Involvement of Nrf2 Signaling Pathway. Oxid. Med. Cell Longev. 2020, 2020, 1605456. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.J.; Pan, Y.Z.; Zhang, Q.; Jiang, B.P.; Xu, L. Salvianolic Acid B Attenuates H2O2-induced NLRP3 Inflammasome Activation via Regulating SIRT1 Signaling Pathway. Tradit. Chin. Drug Res. Clin. Pharmacol. 2021, 32, 604–611. [Google Scholar]
- Li, Q.J. Exploring the Mechanism of Salvianolic Acid B against Acute Myocardial Ischemia Injury Based on the SIRT1-NLRP3 Signaling Axis; Nanjing University Of Chinese Medicine: Nanjing, China, 2021. [Google Scholar]
- Li, Q.; Zuo, Z.; Pan, Y.; Zhang, Q.; Xu, L.; Jiang, B. Salvianolic Acid B Alleviates Myocardial Ischemia Injury by Suppressing NLRP3 Inflammasome Activation via SIRT1-AMPK-PGC-1α Signaling Pathway. Cardiovasc. Toxicol. 2022, 22, 842–857. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.P.; Guo, Z. To Analyze the Mechanism of SalB Regulating SIRT1 to Inhibit NLRP3 and Its Ameliorative Effect on Tubulogastric Junction Tumor Lesions Complicated with Myocardial Injury. Biomed. Res. Int. 2022, 2022, 6560693. [Google Scholar] [CrossRef]
- Hu, Y.; Pan, Y.Z.; Li, Q.J.; Zheng, S.Z.; Bian, Y.; Shi, L.; Fan, F.T.; Jiang, B.P.; Xu, L. Sal B attenuates cardiomyocyte injury by regulating priming of NLRP3 inflammasomes. Chin. Pharmacol. Bull. 2020, 36, 210–215. [Google Scholar]
- Hu, Y.; Wang, X.; Li, Q.; Pan, Y.; Xu, L. Salvianolic acid B alleviates myocardial ischemic injury by promoting mitophagy and inhibiting activation of the NLRP3 inflammasome. Mol. Med. Rep. 2020, 22, 5199–5208. [Google Scholar] [CrossRef]
- Hu, Y.; Li, Q.; Pan, Y.; Xu, L. Sal B Alleviates Myocardial Ischemic Injury by Inhibiting TLR4 and the Priming Phase of NLRP3 Inflammasome. Molecules 2019, 24, 4416. [Google Scholar] [CrossRef]
- Xin, G.J.; Fu, J.H.; Han, X.; Li, L.; Guo, H.; Meng, H.X.; Zhao, Y.W.; Jia, F.F.; Liu, J.X. Salvianolic acid B regulates mitochondrial autophagy mediated by NIX to protect H9c2 cardiomyocytes from hypoxia/reoxygenation injury. China J. Chin. Mater. Medica 2020, 45, 2960–2965. [Google Scholar]
- Yang, B.; Zheng, C.; Yu, H.; Zhang, R.; Zhao, C.; Cai, S. Cardio-protective effects of salvianolic acid B on oxygen and glucose deprivation (OGD)-treated H9c2 cells. Artif. Cells Nanomed. Biotechnol. 2019, 47, 2274–2281. [Google Scholar] [CrossRef]
- Liu, W. The Study of SIRT1-Mediated Protect of Salvianolic Acid B against Myocardial Ischemia-Reperfusion Injury; Guangzhou University of Chinese Medicine: Guangzhou, China, 2019. [Google Scholar]
- Liu, H.; Liu, W.; Qiu, H.; Zou, D.; Cai, H.; Chen, Q.; Zheng, C.; Xu, D. Salvianolic acid B protects against myocardial ischaemia-reperfusion injury in rats via inhibiting high mobility group box 1 protein expression through the PI3K/Akt signalling pathway. Naunyn Schmiedebergs Arch. Pharmacol. 2020, 393, 1527–1539. [Google Scholar] [CrossRef]
- Liu, H. Salvianolic Acid B Protects Against Myocardial Ischemia-Reperfusion Injury in Rats via Inhibiting the Expression of High Mobility Group Box 1 Protein through; Guangzhou University of Chinese Medicine: Guangzhou, China, 2019. [Google Scholar]
- Liu, C.; Cheng, X.D.; Sun, J.A.; Zhang, S.H.; Zhang, Q. Salvianolic acid B inhibits ferroptosis through Cx43 in myocardial infarction rat model. Chin. J. Pathophysiol. 2022, 38, 1032–1039. [Google Scholar]
- Zhao, M.; Li, F.; Jian, Y.; Wang, X.; Yang, H.; Wang, J.; Su, J.; Lu, X.; Xi, M.; Wen, A.; et al. Salvianolic acid B regulates macrophage polarization in ischemic/reperfused hearts by inhibiting mTORC1-induced glycolysis. Eur. J. Pharmacol. 2020, 871, 172916. [Google Scholar] [CrossRef]
- Liu, X.L.; Cui, Z.; Li, Y.; Zhang, Y.M. Protective Effect of Salvianolic Acid B on Myocardial Injury in Septic Rats. Chin. J. Exp. Tradit. Med. Formulae 2019, 25, 112–118. [Google Scholar]
- Lu, B.; Li, J.; Gui, M.; Yao, L.; Fan, M.; Zhou, X.; Fu, D. Salvianolic acid B inhibits myocardial I/R-induced ROS generation and cell apoptosis by regulating the TRIM8/GPX1 pathway. Pharm. Biol. 2022, 60, 1458–1468. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Fu, L.J. The study of salvianolic acid B on the improvement of myocardial fibrosis in diabetic rats by RhoA/ROCK1 pathway. Chin. Pharmacol. Bull. 2022, 28, 1487–1491. [Google Scholar]
- Li, C.L.; Liu, B.; Wang, Z.Y.; Xie, F.; Qiao, W.; Cheng, J.; Kuang, J.Y.; Wang, Y.; Zhang, M.X.; Liu, D.S. Salvianolic acid B improves myocardial function in diabetic cardiomyopathy by suppressing IGFBP3. J. Mol. Cell Cardiol. 2020, 139, 98–112. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Sui, H.J.; Fu, L.J. Effect of salvianolic acid B on proliferation and transdifferentiation of high glucose-induced cardiac fibroblasts. Chin. Pharmacol. Bull. 2020, 36, 414–419. [Google Scholar]
- Wang, N. Effect of Salvianolic Acid B on Proliferation and Transdifferentiation of High Glucose-Induced Cardiac Fibroblasts; Jinzhou Medical University: Jinzhou, China, 2020. [Google Scholar]
- Zhao, M.Q.; Ren, Y.N.; Tao, S.J.; Zheng, S.G. Salvianolic acid B inhibits apoptosis in cardiomyocytes from fluctuating high glucose-induced diabetes mellitus rats. Chin. Tradit. Pat. Med. 2021, 43, 2177–2180. [Google Scholar]
- Li, Y.P.; Wang, C.Y.; Shang, H.T.; Hu, R.R.; Fu, H.; Xiao, X.F. A high-throughput and untargeted lipidomics approach reveals new mechanistic insight and the effects of salvianolic acid B on the metabolic profiles in coronary heart disease rats using ultra-performance liquid chromatography with mass spectrometry. RSC Adv. 2020, 10, 17101–17113. [Google Scholar] [CrossRef]
- Pei, L.L.; Sun, G.C.; Wang, D. Salvianolic acid B inhibits oxidative damage of bone marrow mesenchymal stem cells and promotes differentiation into cardiomyocytes. Chin. J. Tissue Eng. Res. 2021, 25, 1032–1036. [Google Scholar]
- Zhao, X.S.; Zheng, B.; Wen, Y.; Sun, Y.; Wen, J.K.; Zhang, X.H. Salvianolic acid B inhibits Ang II-induced VSMC proliferation in vitro and intimal hyperplasia in vivo by downregulating miR-146a expression. Phytomedicine 2019, 58, 152754. [Google Scholar] [CrossRef]
- Tang, Y.; Wa, Q.; Peng, L.; Zheng, Y.; Chen, J.; Chen, X.; Zou, X.; Huang, S.; Shen, H. Salvianolic Acid B Suppresses ER Stress-Induced NLRP3 Inflammasome and Pyroptosis via the AMPK/FoxO4 and Syndecan-4/Rac1 Signaling Pathways in Human Endothelial Progenitor Cells. Oxid. Med. Cell Longev. 2022, 2022, 8332825. [Google Scholar] [CrossRef]
- Song, Y.Q.; Lu, Z.X.; Yu, J.; Dong, M.X.; Guo, S.; Zhuo, F.Q.; Chen, H.N.; Zhang, Y.G.; Han, L.W. Crucial proangiogenic components in Salvia miltiorrhiza based on zebrafish model and molecular docking technique. Chin. Tradit. Herb. Drugs 2022, 53, 3394–3401. [Google Scholar]
- Niu, W.; Wu, F.; Cao, W.; Chen, Y.; Zhang, Y.; Chen, Y.; Ding, R.; Liang, C. Salvianolic Acid B Alleviates Limb Ischemia in Mice via Promoting SIRT1/PI3K/AKT Pathway-Mediated M2 Macrophage Polarization. Evid. Based Complement. Altern. Med. 2022, 2022, 1112394. [Google Scholar] [CrossRef]
- Xiang, J.; Zhang, C.; Di, T.; Chen, L.; Zhao, W.; Wei, L.; Zhou, S.; Wu, X.; Wang, G.; Zhang, Y. Salvianolic acid B alleviates diabetic endothelial and mitochondrial dysfunction by down-regulating apoptosis and mitophagy of endothelial cells. Bioengineered 2022, 13, 3486–3502. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.N. Salvianolic Acid B Regulates Ang II-Induced Vascular Endothelial Cell Oxidative Stress Upon Akt; MAPK Signaling Pathway; Fujian University of Traditional Chinese Medicine: Fuzhou, China, 2020. [Google Scholar]
- Ait-Oufella, H.; Kinugawa, K.; Zoll, J.; Simon, T.; Boddaert, J.; Heeneman, S.; Blanc-Brude, O.; Barateau, V.; Potteaux, S.; Merval, R.; et al. Lactadherin deficiency leads to apoptotic cell accumulation and accelerated atherosclerosis in mice. Circulation 2007, 115, 2168–2177. [Google Scholar] [CrossRef]
- Guo, Z.Z.; Zhang, Y.N.; Liu, J.C.; Guo, Z.G. Mechanism of PSRC1-ANXA2 interaction in attenuating the progression of atherosclerosis. Med. J. Chin. People’s Lib. Army 2021, 46, 440–447. [Google Scholar]
- Guo, Z.Z. Mechanism of PSRC1-ANXA2 Interaction in Attenuating the Progression of Atherosclerosis; Southern Medical University: Guangzhou, China, 2021. [Google Scholar]
- Zhang, Y.F.; Ding, J.; Du, M.; Feng, X.T.; Liu, P. Effects of salvianolic acid B on atherosclerosis and efferocytosis of macrophages of mice and their mechanisms. J. Jilin Univ. (Med. Ed.) 2022, 48, 561–567. [Google Scholar]
- Zhang, Y.F.; Liu, P. Salvianolic acid B suppresses inflammatory responses via STAT3/NF-κB pathway in atherosclerotic mice and RAW264.7 cells. China J. Tradit. Chin. Med. Pharm. 2023, 38, 141–147. [Google Scholar]
- Zhang, Y.F.; Han, X.H.; Liu, P. Salvianolic acid B reduces liver inflammation in mice with atherosclerosis by inhibiting MAPKs / NF-κB signaling pathway. Chin. Pharmacol. Bull. 2022, 38, 525–530. [Google Scholar]
- Yang, Y.; Pei, K.; Zhang, Q.; Wang, D.; Feng, H.; Du, Z.; Zhang, C.; Gao, Z.; Yang, W.; Wu, J.; et al. Salvianolic acid B ameliorates atherosclerosis via inhibiting YAP/TAZ/JNK signaling pathway in endothelial cells and pericytes. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2020, 1865, 158779. [Google Scholar] [CrossRef]
- Zou, T.; Gao, S.; Yu, Z.; Zhang, F.; Yao, L.; Xu, M.; Li, J.; Wu, Z.; Huang, Y.; Wang, S. Salvianolic acid B inhibits RAW264.7 cell polarization towards the M1 phenotype by inhibiting NF-κB and Akt/mTOR pathway activation. Sci. Rep. 2022, 12, 13857. [Google Scholar] [CrossRef]
- Sun, M.; Ye, Y.; Huang, Y.; Yin, W.; Yu, Z.; Wang, S. Salvianolic acid B improves autophagic dysfunction and decreases the apoptosis of cholesterol crystal-induced macrophages via inhibiting the Akt/mTOR signaling pathway. Mol. Med. Rep. 2021, 24, 763. [Google Scholar] [CrossRef]
- Gao, S.; Li, S.; Li, Q.; Zhang, F.; Sun, M.; Wan, Z.; Wang, S. Protective effects of salvianolic acid B against hydrogen peroxide-induced apoptosis of human umbilical vein endothelial cells and underlying mechanisms. Int. J. Mol. Med. 2019, 44, 457–468. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Yao, D.Y.; Shao, T.M.; An, S.J. Effect of tanshinone II A and salvianolic acid B on adventitia fibroblast transformation. Chin. J. Gerontol. 2021, 41, 1889–1892. [Google Scholar]
- Lin, P.L.; Wu, M.; Yang, F.; Xu, L.; Huang, D.; Qiu, F.R.; Ye, C.Y. Mechanism of salvianolic acid B atenuated phosphate induced vascular smooth muscle cels calcification through activating autophagy. Chin. J. Hosp. Pharm. 2022, 42, 1192–1196. [Google Scholar]
- Du, K.Z.; Cui, Y.; Chen, S.; Yang, R.; Shang, Y.; Wang, C.; Yan, Y.; Li, J.; Chang, Y.X. An integration strategy combined progressive multivariate statistics with anticoagulant activity evaluation for screening anticoagulant quality markers in Chinese patent medicine. J. Ethnopharmacol. 2022, 287, 114964. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Qin, N.; Rao, C.; Zhu, J.; Wang, H.; Hu, G. Screening of Potential Anti-Thrombotic Ingredients from Salvia miltiorrhiza in Zebrafish and by Molecular Docking. Molecules 2021, 26, 6807. [Google Scholar] [CrossRef]
- Zheng, X.; Liu, H.; Ma, M.; Ji, J.; Zhu, F.; Sun, L. Anti-thrombotic activity of phenolic acids obtained from Salvia miltiorrhiza f. alba in TNF-α-stimulated endothelial cells via the NF-κB/JNK/p38 MAPK signaling pathway. Arch. Pharm. Res. 2021, 44, 427–438. [Google Scholar] [CrossRef]
- Dong, S.J. Salvianolic acid B attenuates Iopromide-Induced Renal Tubular Epithelial Cell Injury by Inhibiting the TLR4/NF-κB/NLRP3 Signaling Pathway; Zunyi Medical University: Zunyi, China, 2022. [Google Scholar]
- Dong, S.J.; Gao, X.Y.; Pei, M.X.; Luo, T.; Fan, D.; Chen, Y.L.; Jin, J.F.; Zhao, X.D. Effects and Mechanism of Salvianolic Acid B on the Injury of Human Renal Tubular Epithelial Cells Induced by Iopromide. Front. Pharmacol. 2021, 12, 761908. [Google Scholar] [CrossRef]
- Pei, M.X.; Dong, S.J.; Gao, X.Y.; Luo, T.; Fan, D.; Jin, J.F.; Zhao, X.D.; Chen, Y.L. Salvianolic Acid B Attenuates Iopromide-Induced Renal Tubular Epithelial Cell Injury by Inhibiting the TLR4/NF-κB/NLRP3 Signaling Pathway. Evid. Based Complement. Altern. Med. 2022, 2022, 8400496. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.Y.; Chen, Y.; Luo, T.; Zhao, X.D.; Zhuang, X.D.; Wu, X.X.; Fan, D.; Chen, Y.L.; He, L.Y.; Jin, J.F. Effect and mechanism of salvianolic acid B on apoptosis of HK-2 cells induced by iopromide. Chin. Tradit. Herb. Drugs 2019, 50, 2398–2404. [Google Scholar]
- Pang, X.; Zhang, J.W.; Han, J.R.; Wang, X.M.; Wei, L.J. Protective function and mechanism of SAB by regulated the Bax/Bcl-2 on mesangial proliferative glomerulonephritis. Acta Univ. Med. Anhui 2019, 54, 1221–1226. [Google Scholar]
- Li, H.L.; Zhou, J.; Lin, H.; Lu, R.R.; Liu, B.H.; Chen, J.Q.; Zhou, Y.; Zhou, J.Y.; Wu, J.B. Salvianolic Acid B Attenuates Renal Dysfunction in Chronic Glomerulonephritis through Anti-inflammatory.; Anti-oxidation and Promoting Autophagy. Pharmacol. Clin. Chin. Mater. Medica 2019, 35, 34–38. [Google Scholar]
- Chen, J.; Hu, Q.; Luo, Y.; Luo, L.; Lin, H.; Chen, D.; Xu, Y.; Liu, B.; He, Y.; Liang, C.; et al. Salvianolic acid B attenuates membranous nephropathy by activating renal autophagy via microRNA-145-5p/phosphatidylinositol 3-kinase/AKT pathway. Bioengineered 2022, 13, 13956–13969. [Google Scholar] [CrossRef]
- Sun, Y.B.; Song, Y.R.; Wang, S.Q.; Sun, L.X.; Li, G.Q. Salvianolic acid B attenuates renal interstitial fibrosis in rats with unilateral ureteral obstruction via Nrf2-Gpx4 pathway-mediated ferroptosis. J. Army Med. Univ. 2022, 44, 1018–1024. [Google Scholar]
- Sun, L.; Tian, P.P.; Zhang, F.; Xiao, Y.; Guo, B. Effect of salvianolic acid B on high-glucose induced renal tubular epithelial-mesenchymal transition in rats and its mechanism. China J. Chin. Mater. Medica 2020, 45, 3922–3930. [Google Scholar]
- He, Y.; Lu, R.; Wu, J.; Pang, Y.; Li, J.; Chen, J.; Liu, B.; Zhou, Y.; Zhou, J. Salvianolic acid B attenuates epithelial-mesenchymal transition in renal fibrosis rats through activating Sirt1-mediated autophagy. Biomed. Pharmacother. 2020, 128, 110241. [Google Scholar] [CrossRef]
- Zhang, P.C. Salvianolic Acid-B Regulates Pyroptosis by the Nrf2 in Acute Kidney Injury; Guangzhou University of Chinese Medicine: Guangzhou, China, 2019. [Google Scholar]
- Hu, Y.; Wang, M.; Pan, Y.; Li, Q.; Xu, L. Salvianolic acid B attenuates renal interstitial fibrosis by regulating the HPSE/SDC1 axis. Mol. Med. Rep. 2020, 22, 1325–1334. [Google Scholar] [CrossRef]
- Yao, L.; Zhao, R.; He, S.; Feng, Q.; Qiao, Y.; Wang, P.; Li, J. Effects of salvianolic acid A and salvianolic acid B in renal interstitial fibrosis via PDGF-C/PDGFR-α signaling pathway. Phytomedicine 2022, 106, 154414. [Google Scholar] [CrossRef]
- Mai, X.; Yin, X.; Chen, P.; Zhang, M. Salvianolic Acid B Protects Against Fatty Acid-Induced Renal Tubular Injury via Inhibition of Endoplasmic Reticulum Stress. Front. Pharmacol. 2020, 11, 574229. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Ye, Q.L.; Zhuang, H.S.; Jiang, J.P. Effect of salvianolic acid B on renal fibrosis and inflammation in diabetic nephropathy db/db mice. Chin. Tradit. Herb. Drugs 2022, 53, 1084–1092. [Google Scholar]
- Zhu, Y.M. Protective Effects of Salvianolic Acid B on Renal Fibrosis in Diabetic Rats and the Potential Mechanisms Implicated; Wannan Medical College: Wuhu, China, 2019. [Google Scholar]
- Zhu, Y.M.; Yin, B.J.; Zhang, X.; Tang, B.L.; Cheng, Y.P.; Yang, J.R.; Zheng, S.G. Effect of salvianolic acid B on high glucose-induced phenotypic transition and extracellular matrix secretion in human glomerular mesangial cells. Chin. J. Pathophysiol. 2019, 35, 248–252. [Google Scholar]
- Liu, B.R. Regulatory Mechanism of Salvianolic Acid B on Metabolic Disorders in Mice with Diabetic Nephropathy; Southern Medical University: Guangzhou, China, 2019. [Google Scholar]
- Wang, D. Molecular Mechanism of Salvianolic Acid B Activating Akt/GSK3 β Signal Pathway Preventing Oxidative Stress Injury of Intestinal Mucosa; China Medical University: Shenyang, China, 2022. [Google Scholar]
- Wang, D.; Lu, X.; Wang, E.; Shi, L.; Ma, C.; Tan, X. Salvianolic acid B attenuates oxidative stress-induced injuries in enterocytes by activating Akt/GSK3β signaling and preserving mitochondrial function. Eur. J. Pharmacol. 2021, 909, 174408. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.X. Protective Effect of Salvianolic Acid B Against Intestinal Ischemia-Reperfusion Injury in a Rat Model; Nanchang University: Nanchang, China, 2020. [Google Scholar]
- Chen, G.L.; Xu, S.; Wu, Z.J.; Wu, Y. Protective Effect of Salvianolic Acid B on Intestinal Ischemia-reperfusion Injury in Rats. Acta Acad. Med. Sin. 2020, 42, 30–36. [Google Scholar]
- Feng, P.P.; Fang, X.S.; Zhao, S.H.; Fu, J.Y.; Zhang, H.T.; Yi, Y.L.; Li, C.Y.; Jiang, C.L.; Chen, D.P. Salvianolic acid B decreases interleukin-1β-induced colitis recurrence in mice. Chin. Med. J. 2020, 133, 1436–1444. [Google Scholar] [CrossRef]
- Dai, S.Q. Salvianolic Acid B Alleviates Intervertebral Disc Degeneration by Regulation of Oxidative Stress In Vitro and In Vivo; Soochow University: Suzhou, China, 2021. [Google Scholar]
- Dai, S.; Liang, T.; Shi, X.; Luo, Z.; Yang, H. Salvianolic Acid B Protects Intervertebral Discs from Oxidative Stress-Induced Degeneration via Activation of the JAK2/STAT3 Signaling Pathway. Oxid Med Cell Longev 2021, 2021, 6672978. [Google Scholar] [CrossRef]
- Yan, H.S.; Hang, C.; Chen, S.W.; Wang, K.K.; Bo, P. Salvianolic acid B combined with mesenchymal stem cells contributes to nucleus pulposus regeneration. Connect. Tissue Res. 2020, 61, 435–444. [Google Scholar] [CrossRef]
- Kayalar, E.; Deynek, G.T.; Tok, O.E.; Kucuk, S. Effect of salvianolic acid B on new bone formation in the orthopedically expanded suture. Angle Orthod. 2021, 91, 248–254. [Google Scholar] [CrossRef]
- Li, Y. Salvianolic Acid B Accelerates the Healing of Osteoporotic Fractures by Promoting the Expression of lnc RNAMALAT1; Guangzhou University of Chinese Medicine: Guangzhou, China, 2021. [Google Scholar]
- Ma, L. Salvianolic Acid B Promotes Osteogenesis of Mesenchymal Stem Cells by Protecting Dexamethasone-Induced Mitochondrial Dynamics Disorder; Huazhong University of Science and Technology: Wuhan, China, 2021. [Google Scholar]
- Lin, Y.; Xu, J.; Lu, F.Q.; Xiao, Y.H. Study of the regulation of osteoblast bone formation by salvianolic acid B based on PI3K/AKT signaling pathway. Chin. J. Gerontol. 2019, 39, 2760–2763. [Google Scholar]
- Qiao, J.; Liu, A.; Liu, J.; Guan, D.; Chen, T. Salvianolic acid B (Sal B) alleviates the decreased activity induced by prednisolone acetate on osteoblasts by up-regulation of bone formation and differentiation genes. Food Funct. 2019, 10, 6184–6192. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Huang, Y.; Li, Y.S.; Zhao, W. Salvianolic acid B promotes the proliferatio; migration and osteogenic differentiation of human gingival mesenchymal stem cells by activating the PI3K / AKT pathway. Chin. J. Cell. Mol. Immunol. 2021, 37, 815–820. [Google Scholar]
- Zhang, H.L.; Chen, X. Effects of Salvianolic Acid B on Osteogenic Differentiation and Oxidative Stress of Periodontal Ligament Stem Cells. Genom. Appl. Biol. 2020, 39, 3232–3240. [Google Scholar]
- Bian, Y.; Xiang, J. Salvianolic acid B promotes the osteogenic differentiation of human periodontal ligament cells through Wnt/β-catenin signaling pathway. Arch. Oral. Biol. 2020, 113, 104693. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Wang, F.; Hu, G.; Li, Z. Salvianolic acid B activates chondrocytes autophagy and reduces chondrocyte apoptosis in obese mice via the KCNQ1OT1/miR-128-3p/SIRT1 signaling pathways. Nutr. Metab. 2022, 19, 53. [Google Scholar] [CrossRef]
- Qiu, L.L.; Cai, Z.M.; Zhang, N. Protective effects of Salvianolic acid B on alcoholic liver injury in rats. Chin. J. Clin. Pharmacol. 2020, 36, 2419–2421. [Google Scholar]
- Qiu, L.L.; Chu, J.Y.; Zhang, N. Protective effects of Salvianolic acid B on HepG2 cells with alcoholic injury. Chin. J. Clin. Pharmacol. 2020, 36, 3643–3646. [Google Scholar]
- Lu, W.P.; Wen, Z.F.; Liu, J.Y.; Liao, P.Y.; Chen, Y.L.; Yang, Y.; Chen, M. Improvement and mechanism of salvianolic acid B on non-alcoholic fatty liver in ApoE knockout mice. Chin. Pharmacol. Bull. 2020, 36, 31–37. [Google Scholar]
- Sun, J.F.; Sun, X.; Luo, Y.; Gao, Y.; Wang, S.; Nan, F.W.; Sun, X.B.; Sim, G.B. Protective Effect of Salvianolic Acid B on Arsenic Trioxide-induced HepaRG Human Hepatocyte Injury. Chin. J. Exp. Tradit. Med. Formulae 2020, 26, 68–74. [Google Scholar]
- Sun, J.F. Toxicity Mechanism of Arsenic Trioxide and Protective Effect of Salvianolic Acid B; Harbin University of Commerce: Harbin, China, 2020. [Google Scholar]
- Tao, X.M.; Li, D.; Zhang, C.; Wen, G.H.; Wu, C.; Xu, Y.Y.; Kan, Y.; Lu, W.P.; Ding, H.Y.; Yang, Y. Salvianolic acid B protects against acute and chronic liver injury by inhibiting Smad2C/L phosphorylation. Exp. Ther. Med. 2021, 21, 341. [Google Scholar] [CrossRef]
- Jin, Y.M.; Tao, X.M.; Shi, Y.N.; Lu, Y.; Mei, J.Y. Salvianolic acid B exerts a protective effect in acute liver injury by regulating the Nrf2/HO-1 signaling pathway. Can. J. Physiol. Pharmacol. 2020, 98, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.L.; Jiang, K.Y.; Yang, L.; Xiong, A.Z.; Wang, Z.T. The protective effect of salvianolic acid B against senecionine- induced hepatotoxicity in mice. Acta Pharm. Sin. 2021, 56, 1079–1085. [Google Scholar]
- Li, S.N. The Protective Effect and Mechanism of Salvianolic Acid B Against ANIT-Induced Liver Injury in Rats; Shanghai Jiao Tong University: Shanghai, China, 2019. [Google Scholar]
- Li, S.; Wang, R.; Wu, B.; Wang, Y.; Song, F.; Gu, Y.; Yuan, Y. Salvianolic acid B protects against ANIT-induced cholestatic liver injury through regulating bile acid transporters and enzymes.; and NF-κB/IκB and MAPK pathways. Naunyn Schmiedebergs Arch. Pharmacol. 2019, 392, 1169–1180. [Google Scholar] [CrossRef] [PubMed]
- Su, H.; Ma, Z.; Guo, A.; Wu, H.; Yang, X. Salvianolic acid B protects against sepsis-induced liver injury via activation of SIRT1/PGC-1α signaling. Exp. Ther. Med. 2020, 20, 2675–2683. [Google Scholar] [CrossRef]
- Cao, L.; Yin, G.; Du, J.; Jia, R.; Gao, J.; Shao, N.; Li, Q.; Zhu, H.; Zheng, Y.; Nie, Z.; et al. Salvianolic Acid B Regulates Oxidative Stress.; Autophagy and Apoptosis against Cyclophosphamide-Induced Hepatic Injury in Nile Tilapia (Oreochromis niloticus). Animals 2023, 13, 341. [Google Scholar] [CrossRef]
- Kan, Y.; Yang, Y. Effect of salvianolic acid B on apoptosis of liver fibrotic cells based on cleaved caspase-9 protein. Chin. Pharmacol. Bull. 2019, 35, 827–832. [Google Scholar]
- Kan, Y. Effect of Salvianolic Acid B on Apoptosis of Fibrotic-HCC Cells Based on Cleaved Caspase-9; Anhui Medical University: Hefei, China, 2019. [Google Scholar]
- Wu, C.; Chen, W.; Ding, H.; Li, D.; Wen, G.; Zhang, C.; Lu, W.; Chen, M.; Yang, Y. Salvianolic acid B exerts anti-liver fibrosis effects via inhibition of MAPK-mediated phospho-Smad2/3 at linker regions in vivo and in vitro. Life Sci. 2019, 239, 116881. [Google Scholar] [CrossRef]
- Tao, S.; Duan, R.; Xu, T.; Hong, J.; Gu, W.; Lin, A.; Lian, L.; Huang, H.; Lu, J.; Li, T. Salvianolic acid B inhibits the progression of liver fibrosis in rats via modulation of the Hedgehog signaling pathway. Exp. Ther. Med. 2022, 23, 116. [Google Scholar] [CrossRef]
- Tian, S.; Chen, M.; Wang, B.; Han, Y.; Shang, H.; Chen, J. Salvianolic acid B blocks hepatic stellate cell activation via FGF19/FGFR4 signaling. Ann. Hepatol. 2021, 20, 100259. [Google Scholar] [CrossRef]
- Jiang, N.; Zhang, J.; Ping, J.; Xu, L. Salvianolic acid B inhibits autophagy and activation of hepatic stellate cells induced by TGF-β1 by downregulating the MAPK pathway. Front. Pharmacol. 2022, 13, 938856. [Google Scholar] [CrossRef]
- Zhang, W.; Ping, J.; Zhou, Y.; Chen, G.; Xu, L. Salvianolic Acid B Inhibits Activation of Human Primary Hepatic Stellate Cells Through Downregulation of the Myocyte Enhancer Factor 2 Signaling Pathway. Front. Pharmacol. 2019, 10, 322. [Google Scholar] [CrossRef] [PubMed]
- Ye, S.Q.; Jin, L.Y.; Wang, Y.J.; Li, C.Y.; Zheng, Y.F.; Peng, G.P. Effect of Salvianolic Acid B on Proliferation, Apoptosis and Cell Cycle of HaCaT Cells. Chin. Arch. Tradit. Chin. Med. 2020, 38, 143–146+270. [Google Scholar]
- Liu, Q.; Lu, J.; Lin, J.; Tang, Y.; Pu, W.; Shi, X.; Jiang, S.; Liu, J.; Ma, Y.; Li, Y.; et al. Salvianolic acid B attenuates experimental skin fibrosis of systemic sclerosis. Biomed. Pharmacother. 2019, 110, 546–553. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhu, L.; Xu, Y.; Qin, Z.; Xu, A. Salvianolic acid B ameliorates psoriatic changes in imiquimod-induced psoriasis on BALB/c mice by inhibiting inflammatory and keratin markers via altering phosphatidylinositol-3-kinase/protein kinase B signaling pathway. Korean J. Physiol. Pharmacol. 2020, 24, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Dai, Y.; Yang, X.Z. Effects of salvianolic acid B on high glucose-induced apoptosis and autophagy of retinal Müller cells and AMPK signaling pathway. Recent. Adv. Ophthalmol. 2022, 42, 122–127. [Google Scholar]
- Sun, J.M.; Ho, C.K.; Gao, Y.; Chong, C.H.; Zheng, D.N.; Zhang, Y.F.; Yu, L. Salvianolic acid-B improves fat graft survival by promoting proliferation and adipogenesis. Stem Cell Res. Ther. 2021, 12, 507. [Google Scholar] [CrossRef]
- Wei, S.M.; Huang, Y.M. Attenuation Effect of Salvianolic Acid B on Testicular Ischemia-Reperfusion Injury in Rats. Oxid. Med. Cell Longev. 2022, 2022, 7680182. [Google Scholar] [CrossRef]
- Murata, I.; Sugai, T.; Murakawa, Y.; Miyamoto, Y.; Kobayashi, J.; Inoue, Y.; Kanamoto, I. Salvianolic acid B improves the survival rate; acute kidney dysfunction.; inflammation and NETosis-mediated antibacterial action in a crush syndrome rat model. Exp. Ther. Med. 2022, 23, 320. [Google Scholar] [CrossRef]
- Yifan, Z.; Luwei, X.; Kai, L.; Liuhua, Z.; Yuzheng, G.; Ruipeng, J. Protective effect of salvianolic acid B against oxidative injury associated with cystine stone formation. Urolithiasis 2019, 47, 503–510. [Google Scholar] [CrossRef]
- Kim, S.H.; Lee, J.; Jung, Y.L.; Hong, A.; Nam, S.J.; Lim, B.K. Salvianolic Acid B Inhibits Hand-Foot-Mouth Disease Enterovirus 71 Replication through Enhancement of AKT Signaling Pathway. J. Microbiol. Biotechnol. 2020, 30, 38–43. [Google Scholar]
- Guan, Y.; Li, L.; Kan, L.; Xie, Q. Inhalation of Salvianolic Acid B Prevents Fine Particulate Matter-Induced Acute Airway Inflammation and Oxidative Stress by Downregulating the LTR4/MyD88/NLRP3 Pathway. Oxid. Med. Cell Longev. 2022, 2022, 5044356. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Li, R.; Zhu, R.; Chen, B.; Tian, Y.; Zhang, H.; Xia, B.; Jia, Q.; Wang, L.; Zhao, D.; et al. Salvianolic acid B prevents body weight gain and regulates gut microbiota and LPS/TLR4 signaling pathway in high-fat diet-induced obese mice. Food Funct. 2020, 11, 8743–8756. [Google Scholar] [CrossRef] [PubMed]
- An, T.; Zhang, J.; Lv, B.; Liu, Y.; Huang, J.; Lian, J.; Wu, Y.; Gao, S.; Jiang, G. Salvianolic acid B plays an anti-obesity role in high fat diet-induced obese mice by regulating the expression of mRNA, circRNA, and Lncrna. PeerJ 2019, 7, e6506. [Google Scholar] [CrossRef] [PubMed]
- Lv, B.; Wu, Y.; Lian, J.; Yu, N.; An, T.; Wang, T.; Bao, X.; Mo, F.; Zhao, D.; Yang, X.; et al. Effects of Salvianolic acid B on RNA expression and co-expression network of lncRNAs in brown adipose tissue of obese mice. J. Ethnopharmacol. 2021, 278, 114289. [Google Scholar] [CrossRef]
- Wang, J.; Liu, C.; Que, W.; Fujino, M.; Tong, G.; Yan, H.; Li, X.K. Immunomodulatory effects of Salvianolic acid B in a spontaneous abortion mouse model. J. Reprod. Immunol. 2020, 137, 103075. [Google Scholar] [CrossRef]
- Zhou, R.; Long, H.; Zhang, B.; Lao, Z.; Zheng, Q.; Wang, T.; Zhang, Y.; Wu, Q.; Lai, X.; Li, G.; et al. Salvianolic acid B.; an antioxidant derived from Salvia militarize.; protects mice against γ-radiation-induced damage through Nrf2/Bach1. Mol. Med. Rep. 2019, 19, 1309–1317. [Google Scholar] [CrossRef]
- Szwedowicz, U.; Szewczyk, A.; Gołąb, K.; Choromańska, A. Evaluation of Wound Healing Activity of Salvianolic Acid B on In Vitro Experimental Model. Int. J. Mol. Sci. 2021, 22, 7728. [Google Scholar] [CrossRef]
- Yu, X.; Guan, Q.; Wang, Y.; Shen, H.; Zhai, L.; Lu, X.; Jin, Y. Anticonvulsant and anti-apoptosis effects of salvianolic acid B on pentylenetetrazole-kindled rats via AKT/CREB/BDNF signaling. Epilepsy Res. 2019, 154, 90–96. [Google Scholar] [CrossRef]
- Cai, M.; Wang, M.; Qiao, H.Q.; Liu, J. Embryo-fetal developmental toxicity and genotoxicity of salvianolic acid B. Lishizhen Med. Mater. Medica. Res. 2018, 29, 2351–2353. [Google Scholar]
- Ding, S.S.; Yao, J.J.; Ling, Z.; Li, Y.Y. Study on the safety of salvianolic acid B. West. China J. Pharm. Sci. 2017, 32, 49–51. [Google Scholar]
- Cheng, J.; Long, J.; Zhang, J.; Han, L.; Hu, Y.; Liu, J.; Qiu, R.; Zhu, Z.; Fan, H. Safety, tolerance and pharmacokinetics of salvianolic acid B in healthy Chinese volunteers: A randomized double-blind placebo-controlled phase 1 clinical trial. Front. Pharmacol. 2023, 14, 1146309. [Google Scholar] [CrossRef]
- Han, S.; Zhao, X.Z.; Shi, L.W.; Lin, L.K.; Li, D.K.; Chen, K.X.; Xuan, L.J.; Ma, C.F.J.; Ma, H.Y.; Fang, W.Y.; et al. Clinical Application Evaluation of Salvia Miltiorrhiza Depside Salts for Injection. Chin. Pharm. J. 2021, 56, 422–428. [Google Scholar]
- Chen, Z.W.; Xie, Y.M.; Liao, X.; Wang, G.Q. Systematic review on safety of Salvianolate injection. China J. Chin. Mater. Medica 2016, 41, 3686–3695. [Google Scholar]
- Li, Y.; Wang, L.X.; Xi, Y.M. Efficacy and safety of smiltiorrhizadepsidesal combined with conventional western medicine treatment for stable angina pectoris. China J. Chin. Mater. Medica 2016, 41, 4488–4493. [Google Scholar]
- Zhang, D.L.; Wang, H.Z. A case study of conjunctival hemorrhage due to combination of warfarin and Salvianolate for injection. Cap. Food Medica. 2018, 25, 66. [Google Scholar]
- Li, Y.; Wang, L.; Dong, Z.; Wang, S.; Qi, L.; Cho, K.; Zhang, Z.; Li, N.; Hu, Y.; Jiang, B. Cardioprotection of salvianolic acid B and ginsenoside Rg1 combination on subacute myocardial infarction and the underlying mechanism. Phytomedicine 2019, 57, 255–261. [Google Scholar] [CrossRef]
- Bai, Y.; Bao, X.; Mu, Q.; Fang, X.; Zhu, R.; Liu, C.; Mo, F.; Zhang, D.; Jiang, G.; Li, P.; et al. Ginsenoside Rb1.; salvianolic acid B and their combination modulate gut microbiota and improve glucolipid metabolism in high-fat diet induced obese mice. PeerJ 2021, 9, e10598. [Google Scholar] [CrossRef]
- Wang, J.H. Study on the Anti-Pulmonary Fibrosis Effect of Salvianolic Acid B and Tanshinone IIA Based on THP-1 and MRC-5 Cell Models; Beijing University of Chinese Medicine: Beijing, China, 2019. [Google Scholar]
- Qin, C. Study on the Effect and Mechanism of Salvia miltiorrhiza Extract on Myocardial Tissue of Mice with Kawasaki Disease Model; Liaoning University Of Traditional Chinese Medicine: Shenyang, China, 2020. [Google Scholar]
- Hu, H.Y. Effects of Salvianolic Acid B Combined with Hydroxysafflor Yellow A on Myocardial Ischemia Reperfusion Injury; Nanchang University: Nanchang, China, 2020. [Google Scholar]
- Yang, C.; Cheng, C.; Wang, J.S.; Chen, K.; Zhan, J.; Pan, X.Y.; Xu, X.F.; Xu, W.; Liu, S.W. Salvianolic acid B and its magnesium salt inhibit SARS-CoV-2 infection of Vero-E6 cells by blocking spike protein-mediated membrane fusion. J. South. Med. Univ. 2021, 41, 475–482. [Google Scholar]
- Liang, Y.S.; Ding, C.C.; Luo, P.; Zhang, Z.F.; Wen, L.; Chen, G. A two-dimensional g-C3N4 nanosheet for high loading and sustained release of water-soluble drug salvianolic acid B. Acta Pharm. Sin. 2020, 55, 1296–1305. [Google Scholar]
- Zhang, Y.L. Construction and Preliminary Study of Carboxymethyl Panax Notoginseng Polysaccharide-Chitosan Nanoparticles Loaded with Salvianolic acid B/Doxorubicin; Tianjin University of Traditional Chinese Medicine: Tianjin, China, 2021. [Google Scholar]
- Zhou, L.Y. Study on Salvianolic Acid B Polymer Nanoparticle Brain Targeted Delivery System; Harbin University of Commerce: Harbin, China, 2020. [Google Scholar]
- Ye, R.; Sun, L.; Peng, J.; Wu, A.; Chen, X.; Wen, L.; Bai, C.; Chen, G. Design, Synthesis and Biological Evaluation of Dexamethasone-Salvianolic Acid B Conjugates and Nanodrug Delivery against Cisplatin-Induced Hearing Loss. J. Med. Chem. 2021, 64, 3115–3130. [Google Scholar]
- Liu, W.; Zhou, Z.; Zhu, L.; Li, H.; Wu, L. Chemopreventive efficacy of salvianolic acid B phospholipid complex loaded nanoparticles against experimental oral carcinogenesis: Implication of sustained drug release. Ann. Transl. Med. 2022, 10, 244. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Cui, L.; Chen, G.; Huang, J.; Yang, Y.; Zou, K.; Lai, Y.; Wang, X.; Zou, L.; Wu, T.; et al. PLGA/β-TCP composite scaffold incorporating salvianolic acid B promotes bone fusion by angiogenesis and osteogenesis in a rat spinal fusion model. Biomaterials 2019, 196, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Xu, Z.; Hu, J.; Wu, Z.; Zheng, Y. Preparation and sustained-release properties of poly(lactic acid)/graphene oxide porous biomimetic composite scaffolds loaded with salvianolic acid B. RSC Adv. 2022, 12, 28867–28877. [Google Scholar] [CrossRef]
- Wang, W.; Liu, Y.; Yang, C.; Jia, W.; Qi, X.; Liu, C.; Li, X. Delivery of Salvianolic Acid B for Efficient Osteogenesis and Angiogenesis from Silk Fibroin Combined with Graphene Oxide. ACS Biomater. Sci. Eng. 2020, 6, 3539–3549. [Google Scholar] [CrossRef] [PubMed]
- Ji, C.; Bi, L.; Li, J.; Fan, J. Salvianolic Acid B-Loaded Chitosan/hydroxyapatite Scaffolds Promotes The Repair Of Segmental Bone Defect By Angiogenesis And Osteogenesis. Int. J. Nanomed. 2019, 14, 8271–8284. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Li, W.; Xu, Z.; Hu, J.; Wu, F.; Zheng, Y. Preparation and Synergistic Effect of Biomimetic Poly(lactic acid)/Graphene Oxide Composite Scaffolds Loaded with Dual Drugs. Polymers 2022, 14, 5348. [Google Scholar] [CrossRef]
- Chen, R.; Zhu, C.; Xu, L.; Gu, Y.; Ren, S.; Bai, H.; Zhou, Q.; Liu, X.; Lu, S.; Bi, X.; et al. An injectable peptide hydrogel with excellent self-healing ability to continuously release salvianolic acid B for myocardial infarction. Biomaterials 2021, 274, 120855. [Google Scholar] [CrossRef]
- Hu, J.; Li, C.; Jin, S.; Ye, Y.; Fang, Y.; Xu, P.; Zhang, C. Salvianolic acid B combined with bone marrow mesenchymal stem cells piggybacked on HAMA hydrogel re-transplantation improves intervertebral disc degeneration. Front. Bioeng. Biotechnol. 2022, 10, 950625. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Kong, L.W.; Xu, K.; Sun, Z.J. Effect of gelatin methacryloyl hydrogel loaded with salvianolic acid B on intervertebral disc degeneration. Chin. J. Tissue Eng. Res. 2024, 28, 380–386. [Google Scholar]
- Luo, X.; Xu, X.Q.; Liu, D.R.; Wang, J.; Han, B.; Shi, J. Preparation and properties of tanshinone IIA-salvianolic acid B co-loaded liposomes hydrogel. Chin. Tradit. Herb. Drugs 2022, 53, 2977–2984. [Google Scholar]
- Xu, X.Q. Preparation and Evaluation of Tanshinone IIA-Salvianolic Acid B Co-Loaded Liposome Hydrogel; Guangdong Pharmaceutical University: Guangzhou, China, 2021. [Google Scholar]
- Wu, Y.T.; Guo, S.Y.; Shi, J.; Chen, G.T.; Xu, X.Q.; Zhang, T.Q.; Wu, Y. Preparation of cell penetrating peptide TAT-modified liposomes loaded with salvianolic acid B and its effect on proliferation and migration of human skin fibroblasts. Chin. Tradit. Herb. Drugs 2019, 50, 59–68. [Google Scholar]
- Wu, Y.T. Preparation of Cell Penetrating Peptide TAT-Modified Liposome Loaded with Salvianolic Acid B and Its Effect on Proliferation of HSF; Guangdong Pharmaceutical University: Guangzhou, China, 2019. [Google Scholar]
- Shi, J.; Guo, S.; Wu, Y.; Chen, G.; Lai, J.; Xu, X. Behaviour of cell penetrating peptide TAT-modified liposomes loaded with salvianolic acid B on the migration, proliferation, and survival of human skin fibroblasts. J. Liposome Res. 2020, 30, 93–106. [Google Scholar] [PubMed]
- Liu, R.P.; Zhan, Y.S.; Xu, X.Q.; Luo, X.; Wang, J.; Han, B.; Shi, J. Preparation and characterization of tanshinone IIA-salvianolic acid B co-loaded liposomes and their effects on the biological properties of human skin fibroblasts. Chin. Tradit. Herb. Drugs 2022, 53, 2633–2641. [Google Scholar]
- Chen, Y.N. Study on the Interaction Mechanism of Pegylated Salvianolic Acid B Liposomes and Pegylated Docetaxel Liposomes Based on Tumor Microenvironment; Anhui University of Chinese Medicine: Heifei, China, 2021. [Google Scholar]
- Zhang, X.R.; Lin, T.; Wang, X.L.; Wang, X.J.; Gu, H. Preparation of salvianolic acid B.; tanshinone IIA.; and glycyrrhetinic acid lipid emulsion and its protective effect against acute liver injury induced by acetaminophen. China J. Chin. Mater. Medica 2022, 47, 4634–4642. [Google Scholar]
- Lu, P.; Li, J.; Liu, C.; Yang, J.; Peng, H.; Xue, Z.; Liu, Z. Salvianolic acid B dry powder inhaler for the treatment of idiopathic pulmonary fibrosis. Asian J. Pharm. Sci. 2022, 17, 447–461. [Google Scholar] [CrossRef]
- Guo, J.W.; Cheng, Y.P.; Liu, C.Y.; Thong, H.Y.; Huang, C.J.; Lo, Y.; Wu, C.Y.; Jee, S.H. Salvianolic Acid B in Microemulsion Formulation Provided Sufficient Hydration for Dry Skin and Ameliorated the Severity of Imiquimod-Induced Psoriasis-Like Dermatitis in Mice. Pharmaceutics 2020, 12, 457. [Google Scholar] [CrossRef]
- Cao, F.M.; Hu, D.H. Evaluation of microneedle transdermal administration of salvianolic acid B- tanshinone IIA-glycyrrhetinic acid microemulsion gel. Drug Eval. Res. 2022, 45, 2039–2048. [Google Scholar]
- Zhang, S.; Shi, Y.; Tang, L.; Wang, J.; Guo, D.; Wang, M.; Zhang, X. Evaluation of brain targeting in rats of Salvianolic acid B nasal delivery by the microdialysis technique. Xenobiotica 2018, 48, 851–859. [Google Scholar] [CrossRef]
- Ma, L.Y.; Wang, Z.W.; Fan, J.; Hu, S.S. Interpretation of Report on Cardiovascular Health and Diseases in China 2022. Chin. Gen. Pract. 2023, 26, 3975–3994. [Google Scholar]
- Elmaaty, A.A.; Darwish, K.M.; Khattab, M.; Elhady, S.S.; Salah, M.; Hamed, M.I.A.; Al-Karmalawy, A.A.; Saleh, M.M. In a search for potential drug candidates for combating COVID-19: Computational study revealed salvianolic acid B as a potential therapeutic targeting 3CLpro and spike proteins. J. Biomol. Struct. Dyn. 2022, 40, 8866–8893. [Google Scholar] [CrossRef]
- Hu, S.; Wang, J.; Zhang, Y.; Bai, H.; Wang, C.; Wang, N.; He, L. Three salvianolic acids inhibit 2019-nCoV spike pseudovirus viropexis by binding to both its RBD and receptor ACE2. J. Med. Virol. 2021, 93, 3143–3151. [Google Scholar] [CrossRef] [PubMed]
- Lewinska, A.; Sodagam, L.; Bloniarz, D.; Siems, K.; Wnuk, M.; Rattan, S.I.S. Plant-Derived Molecules α-Boswellic Acid Acetate.; Praeruptorin-A.; and Salvianolic Acid-B Have Age-Related Differential Effects in Young and Senescent Human Fibroblasts In Vitro. Molecules 2019, 25, 141. [Google Scholar] [CrossRef] [PubMed]
- Li, E.; Wang, Y.; Li, Q.; Li, L.; Wei, L. Protective Effects of Sal B on Oxidative Stress-Induced Aging by Regulating the Keap1/Nrf2 Signaling Pathway in Zebrafish. Molecules 2021, 26, 5239. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, Y.; Xu, X.; Cao, W.; Shen, Z.; Wang, N.; Leng, J.; Zou, N.; Shang, E.; Zhu, Z.; et al. Improved dialysis removal of protein-bound uremic toxins by salvianolic acids. Phytomedicine 2019, 57, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.X.; Chen, L.N.; Wang, H.X.; Cheng, X.M.; Ji, L.L.; Li, X.F.; Liu, S.; Kang, X.D.; Wei, X.H.; Wang, C.H. Simultaneous determination of five phenolic acids in Shenkangning Capsules by QAMS. Chin. Tradit. Pat. Med. 2023, 45, 2153–2157. [Google Scholar]
- Yuan, H.; Wang, Y.L.; Zhong, J.Q.; Chen, Z.W.; Wu, A.Y.; Lu, J.G. Simultaneous determination of salvianolic acid B, cryptotanshinone and tanshinone IIA in Yigan Fuzheng capsules by HPLC. Drug Stand. China 2023, 24, 77–80. [Google Scholar]
- Chen, L.Y. Simultaneous Determination of Puerarin Paeoniflorin and Salvianolic Acid B in ChangMaiLe I Capsules by HPLC. Strait Pharm. J. 2021, 33, 77–80. [Google Scholar]
- Yang, X.L.; Wang, H.Y.; Jiang, Z. Determination of content of salvianolic acid B in Xinkeshu Capsules by HPLC. Drug Stand. China 2020, 21, 567–570. [Google Scholar]
- Gao, R.L.; Yang, P.S.; Xu, G.; Wu, X.W.; Yang, C.; Shi, X.Y. Study on Establishment of Near-Infrared Quantitative Model for Salvianolic Acid B in Naoxintong Capsule Based on the System Modeling Idea. Spectrosc. Spectr. Anal. 2020, 40, 3573–3578. [Google Scholar]
- Zhu, Y.L.; Liu, L.; Zhang, J.J.; Lv, M.Y.; Zhang, W.Y.; Hou, X.J.; Wang, W.Q.; Wang, J.F.; Gu, H.S.; Wang, L.Y. Determination of Puerarin, Paeoniflorin, and Salvianolic Acid B in Lutai Danshen Baishao Granules by HPLC. World Chin. Med. 2022, 17, 3139–3142. [Google Scholar]
- Hao, Z.M.; Zhang, Y.O.; Xie, J.; Kang, Y.; Li, T.K. Study on the Contents of Tanshinone IIA, Salvianolic Acid B, Danshensu and Protocatechuic Aldehyde in Danshen Formula Granules. Lishizhen Med. Mater. Medica Res. 2022, 33, 605–607. [Google Scholar]
- Jiang, C.; Huang, Y.F.; He, L.F.; Zhuo, S. Determination of Paeoniflorin and Salvianolic Acid B in DanGua HuMai Granules by HPLC. Strait Pharm. J. 2021, 33, 20–22. [Google Scholar]
- Tang, J.Y.; Zhang, W.M.; Fei, Q.Q.; Bian, Z.H.; Yuan, X.H.; Hu, M.M. Simultaneous Determination of Ferulic Acid and Salvianolic Acid B in Heying Mixture by HPLC. China Pharm. 2022, 31, 68–70. [Google Scholar]
- Zhou, W.Y.; Wang, Y.M.; Huang, Y.; Chen, S.Y. Determination of salvianolic acid B in compound realgar and natural indigo tablets by HPLC. Anhui Med. Pharm. J. 2022, 26, 463–465. [Google Scholar]
- Bian, Y.; Yang, M.S.; Fu, L.J.; Qu, J.L.; Gao, L.; Jiang, Q.R.; Tong, Z.J. Simultaneous Determination of Salvianolic acid B, Puerarin and Icariin in SheXiang XinNaoTong Tablets by HPLC. Strait Pharm. J. 2021, 33, 71–74. [Google Scholar]
- Liu, D.H.; Gao, C.Y.; Zheng, Y.B.; Xu, Y.; Chen, W.K. Determination of Salvianolic Acid B in Fufang Xiatianwu Tablets by Ultra Performance Liquid Chromatography. Drug Eval. 2021, 18, 1300–1302. [Google Scholar]
- Zhao, X.L.; Shang, X.H.; Zhan, C.S. Establishment of a quality control method for the determination of salvianolic acid B in Jianyao Migu tablets. Shanghai Med. Pharm. J. 2021, 42, 73–76. [Google Scholar]
- Shi, F.; Wu, Y.; Wang, Y. Simultaneous Determination of Seven Active Ingredients in Compound Danshen Dropping Pills by HPLC. China Pharm. 2013, 16, 339–341. [Google Scholar]
- Bai, J.; Cai, M.R.; Zhang, Z.Q.; Wu, H.M.; Yao, Y.; Yin, D.G.; Liu, J.; Yin, X.B.; Dong, X.X.; Qu, C.H.; et al. Content determination of salidroside and salvianolic acid B in Jingshen Pills investigation of drying process. Acad. J. Shanghai Univ. Tradit. Chin. Med. 2022, 36, 20–26. [Google Scholar]
- Ren, J.; Jiang, T.; Li, C.; Gu, L.H.; Li, J.M. Content Determination of Tanshinol and Salvianolic Acid B in Zhongfenghuichun Pilule by HPLC. Pharm. Today 2021, 31, 32–34+42. [Google Scholar]
- Qiu, Y.J.; Wang, L.; Zhou, W.G.; Song, H.L.; Li, J.T.; Liu, J.S.; Li, X.J.; Gao, H.; Zhu, G.Q. Content Determination of Salvianolic Acid B in Qingbutongluo Pills by HPLC. China Pharm. 2020, 29, 40–42. [Google Scholar]
- Ma, W.G.; Ma, L.Z. Determination of verbascoside, jionoside B1, rosmarinic acid, lithospermic acid, salvianolic acid B, 2,3,5,4′- tetrahydroxystilbene-2-O-β-D-glucoside, and polydatin in Ruanmailing Oral Liquid by HPLC. Drugs Clin. 2020, 35, 2321–2325. [Google Scholar]
- Duan, G.X.; Niu, X.P.; Cheng, H.J. Determination of rosmarinic acid, lithospermic acid, salvianolic acid B, icariin, baohuoside I, 2,3,5,4′-tetrahydroxystilbene-2-O-β-D-glucoside, and polydatin in Huolisu Oral Liquid by HPLC. Drugs Clin. 2020, 35, 1950–1953. [Google Scholar]
- Zhao, S.S.; Wu, F.; Luo, D.Q.; Kang, X.F.; Liu, H.J. Study on the determination method for salvianolic acid B and the chromatographic fingerprint of Shenkang Suppositories. Northwest Pharm. J. 2019, 34, 466–469. [Google Scholar]
- Li, W.L.; Liu, M.J.; Cao, X.P.; Cui, X.M.; Gao, X.L.; Shi, H.L. Simultaneous Determination of Paeoniflorin, Salvianolic Acid B and Icarin in Kun angao by HPLC. Asia-Pac. Tradit. Med. 2019, 15, 48–51. [Google Scholar]






| Pharmacology | Experimental Subject | Experimental Dose | Animal Experimental Drug Delivery Method | Effects | Ref. |
|---|---|---|---|---|---|
| Antitumor effects | Colon cancer cells RKO Prostate cancer cells PC3 Mice inoculated with MC38 colon cancer cells | 5, 10, 20 μMol/L 10, 20 mg/kg | Intraperitoneal injection | Decrease: PD-L1, USP2 Increase: CD8+ T | [34] |
| Human-derived HCT116 colon cancer cell line | 50, 100, 200 μMol/L | None | Decrease: LC3-I, p62, AKT, mTOR Increase: LC3-II, Atg5 | [35] | |
| TGF-β1-induced NSCLC A549 cells | 25, 50, 100 µM | None | Decrease: N-cadherin, vimentin, Snail, cyclin B1, LC3α, p62, Bcl-2, ERK1/2, JNK1/2, p38, p-Smad2, p-Smad3 Increase: p21, LC3β, Beclin1, Bax, caspase-3, cleaved caspase-3 | [37] | |
| NSCLC NCI-H2030 and NCI-H1650 | 100, 200, 300, 400, 500, 600 μM | None | Decrease: Cell migration, Cell invasion, Glucose uptake, Lactate production, Enolase activity, ATP, Oxygen consumption rate, Extracellular acidification rate, Pyruvate kinase M2, LDHA, GLUT1, β-catenin, E-cadherin | [38] | |
| Normal gastric mucosa epithelial cell line GSE1 Gastric cancer cells AGS and AGS/cisplatin | 25, 50, 100, 200, 250 µM | None | Decrease: Cell viability, Cell clone, Cell migration, Cell invasion, Vimentin, N-cadherin, MMP2, MMP9, EMT formation Increase: ROS, E-cadherin | [39] | |
| Human hepatocellular carcinoma cell lines HepG2 and HCCLM3 | 100, 200, 300, 400, 500, 600 μM | None | Decrease: N-cadherin, vimentin, Mortalin, MMP2, MMP9, p-STAT3Y705, Ac-STAT3K685 Increase: RECK, E-cadherin | [40] | |
| 3-Aminopropionitrile induced in rats | 20 mg/kg | Intraperitoneal injection | Decrease: MMP -2, MMP -9, CD8+, ROS, MDA, IL-1β, IL-6, IL-8, TNF-α, JAK2, STAT3, p-JAK2, p-STAT3 Increase: CD4+, CD4+/CD8+, IgA, IgG, IgM, SOD | [41] | |
| Triple-negative breast cancer cells MDA-MB-231 and MDA-MB-468 | 3.12, 6.25, 12.5, 25, 50 μM | None | Decrease: Cell migration, Cell invasion, E-cadherin, Cytokeratin 19 Increase: N-cadherin, Vimentin, p-Snail/Snail, p-Slug/Slug | [42] | |
| Human MDA-MB-231 breast cancer cells | 0–100 μM | None | Decrease: MMP-9 | [43] | |
| Mice inoculated with 4T1 murine-derived breast cancer cells and MDA-MB-231 human-derived breast cancer cells | 100, 40 mg/kg | Intraperitoneal injection | Increase: NG2+, αSMA+, Collagen IV+, VE-Cadherin+, Claudin 5+ | [44] | |
| Human breast cancer adenocarcinoma MCF-7 Mice injected with Ehrlich solid carcinoma cell line | 0 to 1.0 mg/mL 25 mg/kg | Intraperitoneal injection | Decrease: TNF-α, MMP-8, Cyclin D1, VEGF, COX-2, MDA Increase: caspase-3, P53, GSH | [45] | |
| Endometrial cancer MFE-280 cells | 0.5, 1, 2.5 μMol/L | None | Decrease: Cell clone formation rate, p-PI3K/PI3K, p-AKT/AKT Increase: Apoptosis rate, Beclin1, LC3-II/LC3-I, ATG7, Mitochondrial membrane potential | [46] | |
| Ovarian cancer cells ID8 | 0.87, 1.74, 3.48, 6.96, 13.92 μM | None | Decrease: Cell proliferation, Cell cloning ability, PI3K110β, PDPK1, p-PDPK1 (Ser241), pan-Akt, p-Akt1 (Ser473), p-GSK3β (Y216) | [47] | |
| Ovarian cancer cells ID8 Mice inoculated with ovarian cancer cells ID8 | 3.75, 0.9375, 1.875, 3.75, 7.5, 15 μM | None | Decrease: Cell proliferation, Cell cloning ability, p-IκBα, IκBα, p-p65, p65 | [48] |
| Pharmacology | Experimental Subject | Experimental Dose | Animal Experimental Drug Delivery Method | Effects | Ref. |
|---|---|---|---|---|---|
| Effects on the lung | TGF-β1-induced human lung epithelial A549 cells | 10, 20, 40, 80, 160 μMol/L | None | Decrease: Cell proliferation, FN, Collagen type I Increase: E-calcium viscosity | [50] |
| LPS-induced human acute monocytic leukemia cell line THP-1 TGF-β1-induced human lung fibroblast cell line MRC-5 | 20, 50, 100, 200, 300, 400, 500, 600, 700, 800, 900 μg/mL | None | Decrease: TNF-α, IL-1β, IL-6, α-SMA, Collagen 1α1 | [51] | |
| Bleomycin-induced rats | 1, 10, 25, 40, 50 mg/mL | Nebulized inhalation | Decrease: Hydroxyproline, Collagen-1, Tissue factor/Coagulation factor VII, Activated coagulation factor X, Thrombin–antithrombin complex, Fibrinogen degradation product, Plasminogen activator inhibitor-1, PAR-1, p-PKC Increase: Coagulation factor Ⅱ, Coagulation factor X, Tissue-type plasminogen activator, Urokinase-type plasminogen activator | [52] | |
| Nasal drops of Staphylococcus aureus in young rats | 5, 10, 20 mg/kg | Intraperitoneal injection | Decrease: IL-1β, IL-6, TNF-α, NF-κBp65, MDA Increase: Arterial partial pressure of oxygen, Oxygenation index, SOD | [53] |
| Pharmacology | Experimental Subject | Experimental Dose | Animal Experimental Drug Delivery Method | Effects | Ref. |
|---|---|---|---|---|---|
| Effects on the heart | Mice | 30, 60, 120 mg/kg | Gavage administration | Decrease: MDA, Bacterbidetes, Proteobacteria, Verrucomicrobia, Actinobacteria Increase: GSH, SOD, Firmicutes A, Gordonibacter | [68] |
| Mice were subjected to cardiac arrest induced by an intravenous injection of potassium chloride, followed by cardiopulmonary resuscitation H9c2 cardiomyocytes with hypoxia/reoxygenation | 20 mg/kg 10 μM | Intravenously | Decrease: Bax, caspase-3, SOD, NADPH oxidase activity, Keap1 Increase: Bcl-2, Bcl-2/Bax, HO-1, NQO1 | [69] | |
| H2O2-induced cardiomyocyte H9c2 cells Ligation of the left anterior descending coronary artery in rats | 5, 10, 20 μMol/L 8, 32 mg/kg | Intravenously | Decrease: LDH, CK-MB, cTnI, ROS, IL-1β, NLRP3, Caspase-1, ASC, p-AMPKα/AMPKα, PGC-1α Increase: Mitochondrial membrane potential | [70,71,72,73] | |
| Hypoxia-induced rat cardiomyocytes H9C2 | 1, 5, 10, 15, 20, 25, 30, 35 μMol/L | None | Decrease: LDH, cTnI, IL-1β, NLRP3, TLR4, Myd88, IRAK1, NF-κB | [74,75,76] | |
| H9c2 hypoxia/reoxygenation injury in cardiac myocytes | 50 μMol/L | None | Decrease: LDH, ROS, LC3-II/LC3-I, cleaved caspase-3, NIX Increase: ATP, Mitochondrial membrane potential | [77] | |
| H9c2 cells with oxygen and glucose deprivation | 1, 5, 10 μM | None | Decrease: Bax, caspase-3, caspase-9, MEG3, P53 Increase: cyclin D1, AMPK | [78] | |
| Ischemia/reperfusion of the left anterior descending branch of the ligated coronary artery in rats Cardiomyocyte H9c2 oxygen–glucose deprivation/reperfusion | 200, 100, 50 μM 30, 60 mg/kg | Intraperitoneal injection | Decrease: LDH, CK-MB, C-HMGB1, ac-HMGB1, TLR4, NF-κB, Bax Increase: SIRT1, Bcl-2, N-HMGB1 | [79] | |
| Ischemia/reperfusion of coronary artery ligation in rats | 15, 60 mg/kg | Intraperitoneal injection | Decrease: L-LDH, CK-MB, TNF-α, IL-18, IL-β, TLR4 Increase: PI3K, AKT | [80,81] | |
| Ligation of the left coronary artery in rats | 40 mg/kg | Intraperitoneal injection | Decrease: CK-MB, cTnI, LDH, MDA, ROS, Iron ion content, Area of Perl blue positivity Increase: GSH, Cx43 | [82] | |
| LPS-induced bone-marrow-derived macrophages Myocardial ischemia/reperfusion in mice induced by ligation of the left anterior descending artery | 80 mg/kg 5, 10, 20 μM | Gavage administration | (M1 biomarkers) Decrease: IL-6, iNOS, CCL2, TNF-α (M2 biomarkers) Decrease: RagD, mTORC1 Increase: Arg1, Clec10a, Mrc, CX3CR1, CD36, MerTK, IL-10 | [83] | |
| Rats undergoing cecum ligation and perforation | 6, 12, 24 mg/kg | Intravenously | Decrease: RagD, mTORC1, Myocardial necrosis markers troponin T, CK-MB, cTnI, IL-1β, IL-6, TNF-α, MDA Increase: SOD, LC3II/LC3I, Beclin-1 | [84] | |
| AC16 cardiomyocytes were treated with ischemia/reperfusion The rat coronary artery was clamped with a plastic tube, and then the coronary artery was restored by releasing the clamp. | 10, 25, 50 μM 20, 40, 60 mg/kg | Intravenously | Decrease: Apoptotic, ROS, MDA, TRIM8 Increase: SOD | [85] | |
| Streptozotocin injection in rats | 12.5, 25, 50 μMol/L | Gavage administration | Decrease: α-SMA, Collagen I, Collagen III, Bcl-2 Increase: RhoA, ROCK1, Bax, Beclin1, LC3-I, LC3-II | [86] | |
| Streptozotocin-induced mice HUVECs under hypoxia | 15, 30 mg/kg 50–100 μg/mL | Intraperitoneal injection | Decrease: ECM, Collagen I, Collagen III, IGFBP3, AKT phosphorylation, ERK phosphorylation Increase: VEGF, VEGFR2 | [87] | |
| High-glucose-induced cardiac fibroblasts in rats | 12.5, 25, 50 μMol/L | None | Decrease: Collagen fiber deposition, α-SMA, β-catenin, p-GSK 3β | [88,89] | |
| High-sugar, high-fat diet combined with streptozotocin injections induced in rats | 160, 80 mg/kg | Gavage administration | Decrease: Fasting Glucose, CK-MB, Bax, caspase-3 Increase: Bcl-2 | [90] | |
| Hydrogen peroxide intervention in rat bone marrow mesenchymal stem cells | 50, 100, 150, 200 μg/L | None | Decrease: CK, LDH, MDA Increase: SOD Decrease: Keap1 Increase: Nrf2, Bcl-2/Bax, GATA4 genes, cTnT | [92] | |
| The left common carotid artery was dissected and ligated near the carotid bifurcation using suture to induce intima formation in mice Ang II-induced mouse aortic vascular smooth muscle cells | 7 mg/kg 2.5, 5, 10 μMol/L | Intraperitoneal injection | Decrease: KLF5, cyclin D1, miR-146a | [93] | |
| Tunicamycin-induced bone-marrow-derived endothelial progenitor cells | 10, 20 μM | None | Decrease: ROS, TXNIP, NLRP3, Cleavage caspase-1, IL-1β, IL-18, Caspase, Syndencan-4 Increase: HO-1, SOD2, p-AMPKα, KLF2 | [94] |
| Pharmacology | Experimental Subject | Experimental Dose | Animal Experimental Drug Delivery Method | Effects | Ref. |
|---|---|---|---|---|---|
| Effects on blood vessels | PTK787 causes vascular damage in zebrafish | 10, 20, 40, 80 μg/mL | None | Increase: Generation of blood vessels | [95] |
| Mice with hind limb ischemia model Mouse bone-marrow-derived macrophages isolated from the femur and the tibia bone marrow cells treated with salvianolic acid B, then collected the culture supernatant to culture HUVECs | 15 mg/kg 1, 5, 25 μM | Intraperitoneal injection | Increase: M2 Polarization, Cell migration, Tube formation | [96] | |
| High-glucose-induced HUVECs exposed to carbonyl cyanide m-chlorophenyl hydrazone C57BL/KsJ db/db mice | 5, 10, 30, 50, 80, 100 μM 50 mg/kg | Oral administration | Decrease: Bax, Beclin1, Parkin, PTEN Induced Kinase 1 (Pink1), TC, TG, LDL-C, HDL-C, Fasting blood glucose, Fasting insulin, Homeostasis model assessment insulin resistance Increase: Bcl-2, Mitochondrial activity, Ca2+ | [97] | |
| Ang II-induced HUVEC | 25, 50, 100, 200, 400, 800 μM | None | Decrease: ROS, MDA, NOX2 Increase: NO, Intracellular calcium ions, ERK1/2, p38, JNK, PI3K, AKT, NRF2, HO-1 | [98] | |
| High-fat diet fed to LDLR knockout mice ox-LDL-induced macrophage RAW264.7 in mice | 1.25, 2.50, 5.00 mg/L 25 mg/kg | Intraperitoneal injection | Decrease: TC, TG, LDL-c Increase: AXL, MERTK, TYRO3, MFGE8 | [102] | |
| High-fat diet fed to LDLR knockout mice LPS-induced macrophage RAW264.7 in mice | 200 μL 1.25, 2.5, 5 μg/mL | Intraperitoneal injection | Decrease: IL-6, IL-1β, TNF-α, TC, TG, LDL-c, STAT3, IκB, NF-κB | [103] | |
| High-fat diet fed to LDLR knockout mice ox-LDL-induced or LPS-induced RAW264.7 cells | 25 mg/kg 1.25, 2.5, 5 μg/mL | Intraperitoneal injection | Decrease: TC, TG, LDL-C, AST, ALT, IL-6, IL-1β, TNF-α, VCAM, iNOS, p-JNK, p-P38, p-ERK 1/2, p-IκB, p-NF-κB p65 | [104] | |
| ApoE-/- mice with high-fat-diet ox-LDL-induced Ecs and pericytes | 30, 25 μg/mL 30 mg/kg | Intraperitoneal injection | Decrease: YAP, TAZ, JNK, Nuclear NF-κB P65, Total P65, TNF-α, ROS, MDA, TG, TC, LDL-C, IL-6, IL-1 β, TNF-α Increase: p-YAP, p-TAZ, SOD, Gpx | [105] | |
| RAW264.7 macrophages | 50, 100, 150, 200, 250, 300, 350 μM | None | Decrease: iNOS, TNF-α, IL-6, p62, NF-κB p65 Increase: Arg-1, IL-10, LC3-II, Beclin-1, p-AKT, p-mTOR | [106] | |
| Cholesterol crystals stimulate RAW264.7 macrophages | 40, 80, 120, 160, 200, 240, 280, 320, 360 µM | None | Decrease: TNF-α, IL-6, cleaved-caspase 3/pro-caspase 3, Bax/Bcl-2, p62 Increase: LC3 II, Beclin-1 | [107] | |
| H2O2-induced HUVECs | 5, 10, 20 µg/ml | None | Decrease: p62, Phosphorylation of mTOR, caspase-3, cytochrome c Increase: LC3-II, Beclin-1, Phosphorylation of AMPK | [108] | |
| Ang II-induced rat thoracic aortic outer membrane | 10−5 mol/L | None | Decrease: ROS, Cell migration capacity, α-SMA | [109] | |
| High-phosphorus-induced vascular smooth muscle cells in rats | 3, 10, 30 nmol/g | None | Decrease: Runx2, OPN Increase: Beclin-1, LC3-I/II, Calponin, SM22 | [110] | |
| TNF-α-induced human EA.hy 926 cells Abdominal aorta of the rats | 1, 5, 25, 50 μM | None | Decrease: PAI-1, Factor IIa, Factor Xa, Tissue factor, p-NF-κB p65, p-p38 MAPK, pJNK Increase: Tissue plasminogen activator | [113] |
| Pharmacology | Experimental Subject | Experimental Dose | Animal Experimental Drug Delivery Method | Effects | Ref. |
|---|---|---|---|---|---|
| Effects on the kidney | Iopromide-induced damage to HK-2 human renal cortical proximal tubular epithelial cells | 10, 50, 100 μMol/L | None | Decrease: Apoptosis, Bax/Bcl-2, cleaved caspase-3, ROS, Caspase-1, NLRP3, TLR4, p-NF-κB, IL-18, IL-1β, TNF-α, ASC, GRP78, p-eIF2α, p-JNK, CHOP Increase: Cellular mitochondrial membrane potential, Cellular viability | [114,115,116] |
| Iopromide-induced HK-2 human renal proximal tubular epithelial cells | 1, 10, 50, 100, 200 μMol/L | None | Decrease: ROS, p-ERK1/2, Bax, cleaved Caspase-3 Increase: p-Akt, Klotho, Bcl-2 | [117] | |
| Modified chronic serum disease method for induction of rats | 100 mg/kg | Gavage administration | Decrease: Serum creatinine, Urea nitrogen, Glomerular cell apoptosis, Caspase-3, Bax, Bax/Bcl-2, IL-1β, IL-6, IL-18, TNF-α, NF-κB p65 Increase: Bcl-2 | [118] | |
| Rats injected with cationized calf serum albumin and Freund’s incomplete adjuvant | 50, 100 mg/kg | Gavage administration | Decrease: Serum creatinine, Urea nitrogen, Urine protein, MDA, C3 deposition, IgG deposition, IL-1β, IL-6, p62 Increase: GSH, SOD, LC3 II/LC3 I, Beclin1 | [119] | |
| Unilateral ureteral obstruction in rats | 100 mg/kg | Gavage administration | Decrease: Fe2+, MDA, Collagen-I, Collagen-III Increase: SOD, Nrf2, Gpx4 | [121] | |
| High-glucose-induced rat proximal renal tubular epithelial cell line NRK-52E | 1, 5, 10, 20, 50, 100 μMol/L | None | Decrease: α-SMA, p-Akt Increase: PPARγ, PTEN, E-cadherin | [122] | |
| Rats subjected to unilateral nephrectomy TGF-β1-induced HK-2 cells | 200, 100, 50 mg/kg 100 μM | Oral administration | Decrease: Fibronectin, α-SMA, TGF-β, Beclin1, LC3B Increase: p62, Sirt1 | [123] | |
| Unilateral nephrectomy in mice with contralateral renal artery clamping of the renal tip Hypoxia-reoxygenated renal tubular epithelial HK-2 cells | 200, 100, 50 mg/kg 80, 40, 20 μM | Gavage administration | Decrease: IL-1β, TNF-α, Caspase-1, MDA, NLRP3, TXNIP, Keap1, GSDMD Increase: HO-1, SOD, GSH | [124] | |
| Cationic bovine serum albumin-induced rats LPS-induced human mesangial cells | 100 mg/kg | Oral administration | Decrease: TNF-α, IL-6, IL-1β, IL-2, CD68 Increase: miR-145-5p, LC3B, Beclin-1 | [120] | |
| Mice with left ureteral ligation Ang II-induced HK-2 cells | 6.25, 12.5, 25 mg/kg 0.1, 1, 10 µM | Intraperitoneal injection | Decrease: Serum creatinine, Blood urea nitrogen, TGF-β1, FGF-2, SDC1, α-SMA Increase: HPSE, E-cadherin | [125] | |
| Rat with the left ureter was exposed via a left flank incision and ligated Human serum albumin induced the HK-2 human renal proximal tubular cell line | 12.5 mg/kg 20 μM/L | Oral administration | Decrease: β2-microglobulin, N-Acetamidoglucosidase, Blood urea nitrogen, Serum creatinine, CTGF, PDGF-C, PDGFR-α, caspase-3, Early growth response protein 1, CHOP, GRP78 Increase: Par-3 | [126] | |
| High-fat diet fed in mice Palmitic acid-, tunicamycin-, and thapsigargin-induced human proximal tubule epithelial HK-2 cells | 3, 6.25, 12.5 mg/kg 1, 10, 100 μM | Intraperitoneal injection | Decrease: ICAM-1, VCAM-1, IL-1β, IL-6, TNF-α, Bax, cleaved caspase-3, BIP, P-eIF2α, ATF4, CHOP, ATF6, IRE1α, XBP1s Increase: Bcl-2 | [127] | |
| db/db mice fed a high-calorie diet | 50, 100, 150 mg/kg | Gavage administration | Decrease: p-Smad3/Smad3, Collagen type IV, TGF-β1, IL-6, TNF-α, Urine microalbumin, Urine creatinine, Blood urea nitrogen, TC, TG | [128] | |
| High-glucose and high-fat feeding combined with streptozotocin-induced rats High-glucose-induced glomerular-tethered HGMC cells | 80, 160 mg/kg 10−7, 10−6, 10−5 Mol/L | Gavage administration | Decrease: Fasting blood glucose, TG, TC, Urea nitrogen, Urine microalbumin, Serum creatinine, MDA, NO, Collagen content, TGF-β1, Phosphorylated Smad2, Phosphorylation of p38MAPK, Collagen I, Collagen III, Fibronectin, Laminin, α-SMA, Smad2, p38MAPK Increase: Smad7, Albumin, Total protein | [129,130] |
| Pharmacology | Experimental Subject | Experimental Dose | Animal Experimental Drug Delivery Method | Effects | Ref. |
|---|---|---|---|---|---|
| Effects on the intestine | H2O2-induced rat small intestine IEC-6 cells Ischemia/reperfusion injury of rat small intestinal mucosa | 10, 50, 100, 500 μM 40 mg/kg | Intraperitoneal injection | Decrease: Apoptosis, Mitochondrial membrane potential depolarization, ROS, Cyt-c, caspase-3, Bax Increase: Bcl-2 | [132,133] |
| Clamping of the superior mesenteric artery in rats | 40 mg/kg | Intraperitoneal injection | Decrease: MDA, IL-1β, IL-6, TNF-α, MPO, NF-κB p65 Increase: PI3Kp85α, p-AKT (ser473), SOD, CAT, GSH | [134] | |
| Intestinal ischemia/reperfusion injury in rats | 40 mg/kg | Intraperitoneal injection | Decrease: MDA, TNF-α, IL-1β, IL-6, NF-κB Increase: SOD | [135] | |
| Mice with free drinking water containing dextran sulfate sodium and injected with IL-1β | 80, 100 mg/kg | Gavage administration | Decrease: MLCK | [136] |
| Pharmacology | Experimental Subject | Experimental Dose | Animal Experimental Drug Delivery Method | Effects | Ref. |
|---|---|---|---|---|---|
| Effects on the bone | H2O2-induced extraction of nucleus pulposus cells from rat caudal intervertebral discs Induction of degeneration of the caudal intervertebral disc in rats by percutaneous needling | 0.001, 0.01, 0.1, 1, 10, 100 nM 200 mg/kg | Gavage administration | Decrease: ROS, MDA, Bax, Cleaved caspase 3, IL-1β, IL-6, TNF-α, MMP3, MMP13, ADAMTS4, ADAMTS5 Increase: SOD, CAT, GSH-Px, Bcl-2, Collagen II, Aggrecan | [137,138] |
| Aspiration of the nucleus pulposus in New Zealand white rabbits | 0.01, 0.1, 1, 10 mg/L | Injection passed through the annulus fibrosus | Increase: Runx2, Sox9, Collagen II, Proteoglycan content, TGF-β1 | [139] | |
| Expand the premaxillary suture of rats | 40 mg/kg | Intraperitoneal injection | Increase: Vascular, Osteoblasts, Osteoclasts | [140] | |
| Human bone marrow mesenchymal stem cells Bilateral ovarian removal combined with sawing of the femur in rats | 0.5, 1, 5 μM 2.5 mg/kg | Local injection | Increase: Endomucin, Hypoxia-inducible factor 1-α, OPN | [141] | |
| Extraction of mesenchymal stem cells from human bone marrow | 5 μM | None | Decrease: ROS | [142] | |
| Mouse osteoblasts MC3T3-E1 | 0.01, 0.03, 0.10, 0.30, 1.00 nmol/L | None | Increase: OPN, OCN, Collagen I, ALP, PI3K, p-AKT | [143] | |
| Prednisolone acetate-induced osteoblasts isolated from the skull of rats | None | None | Decrease: Nrf2 Increase: Runx2, Osx, OCN, IGF-I, Collagen I, HO-I | [144] | |
| Human gingival mesenchymal stem cells | 0, 5, 10 μMol/L | None | Decrease: PPARγ, C/EBPα Increase: Runx2, OCN, Collagen I, ALP | [145] | |
| H2O2-induced periodontal ligament stem cells in orthodontic patients | 0.5, 1, 5, 10 μMol/L | None | Decrease: PPARγ, LPL, C/EBPα, ROS Increase: ALP, Collagen 1, RUNX2, OCN, Akt, SOD, GSH, Nrf2 | [136] | |
| Human periodontal ligament cell tissues were isolated from periodontal ligament of the middle third of healthy molars | 0.1, 0.5, 1, 5 μM | None | Increase: RUNX2, BMP2, OSX, OCN, β-catenin, LEF1, Cyclin D1, ALP, Phosphorylated GSK-3β | [147] | |
| High-fat diet feeding and anterior cruciate ligament transection/medial meniscus removal in mice Palmitic-acid-induced mouse chondrocyte cell line ATDC5 | 25 mg/kg 25, 50, 100 μM | Intraperitoneal injection | Decrease: TNF-α, IL-6, PGE2, JAK2, STAT3 Increase: KCNQ1OT1, Bcl-2, LC3II/LC3I, Beclin-1 | [148] |
| Pharmacology | Experimental Subject | Experimental Dose | Animal Experimental Drug Delivery Method | Effects | Ref. |
|---|---|---|---|---|---|
| Effects on the liver | Alcoholic liquid feed for rats | 30, 15, 30 mg/kg | Gavage administration | Decrease: ALT, AST, TG, TC Increase: Ethanol dehydrogenase, SIRT1, p-AMPKα1/AMPKα1 | [149] |
| Ethanol-induced HepG2 cells | 8 μMol/L | None | Increase: Cell survival rate, p-AMPKα1/AMPKα1 | [150] | |
| High-fat-diet-induced ApoE knockout mice | 15 mg/kg | Intraperitoneal injection | Decrease: AST, ALT, TNF-α, NF-κB, IL-6, Nrf2, HO- 1 Increase: Beclin1, p62, LC3, AMPK | [151] | |
| Arsenic-trioxide-induced HepaRG human hepatocytes | 10, 5, 2.5 μMol/L | None | Decrease: Apoptosis Increase: Cell survival, Mitochondrial membrane potential, Bcl-2/Bax, p-Akt/Akt | [152] | |
| Arsenic-trioxide-induced rat | 5, 10, 20, 40 mg/kg | Intraperitoneal injection | Decrease: MDA Increase: SOD | [153] | |
| Mice injected with CCl4 H2O2-induced rat hepatic stellate cells HSC-T6 | 7.5, 15, 30 mg/kg 25, 50, 100 μMol/L | Gavage administration | Decrease: ALT, AST, MDA, pSmad2C, pSmad2L Increase: SOD, GSH | [154,155] | |
| Senecionine-induced mice | 10 mg/kg | Gavage administration | Decrease: GSH, PAI-1, MMP-9, TGF-β1 Increase: SOD | [156] | |
| α-Naphthyl isothiocyanate-induced rat | 15, 30 mg/kg | Intraperitoneal injection | Decrease: ALT, ALP, AST, γ-glutamyl transferase, Total bilirubin, Direct bilirubin, Total bile acids, IL-1β, IL-6, TNF-α, TGF-β, COX-2 Increase: Cyp3a2, Ugt1a1 | [157,158] | |
| A cecal ligation and puncture method to induce mice sepsis | 30 mg/kg | Intraperitoneal injection | Decrease: TNF-α, IL-6, Bax, AST, ALT, MPO, caspase-3 Increase: Bcl-2, SIRT1, PGC-1α | [159] | |
| Cyclophosphamide-induced Nile tilapia | 0.25, 0.50, 0.75 g/kg | Mixed with feed for oral administration | Decrease: ROS, Atg-3, -5, -7, -13, Beclin1, MP-activated protein kinase, unc-51-like autophagy-activating kinase 1, mucolipin-1, lysosomal associated membrane protein 1, UV-radiation-resistance-associated gene, transcription factor EB, caspase-3, -8, and -9, Cytc, Bax, p53, JNK1, 2, apoptosis signal-regulating kinase 1, p38, extracellular signal-regulated protein kinase Increase: SOD, GSH, CAT, NO, Gpx3, glutamate–cysteine ligase catalytic subunit, glutathione reductase, glutathione s-transfer α, catalase, nrf2, uncoupling protein 2, heme oxygenase-1, p62, mTOR, Bcl-2 | [160] | |
| Diethylnitrosamine-induced mouse TGF-β1-induced HSC-T6 cells | 25, 50, 100 μMol/L 15, 30 mg/kg | Gavage administration | Increase: Cleaved caspase-9, Apoptosis | [161,162] | |
| Diethylnitrosamine-induced mouse TGF-β1-induced HSC-T6 cells and LX-2 cells | 25, 50, 100 μMol/L 15, 30 mg/kg | Gavage administration | Decrease: α-SM, TGF-β1, Collagen I, P-Smad2C, P-Smad2L, P-ERK1/2, P-JNK1/2, P-p38, P-Smad3C, P-Smad3L, Plasminogen activator inhibitor 1 | [163] | |
| Carbon-tetrachloride-induced rats | 60 mg/kg | Oral administration | Decrease: TGF-β1, Shh, Ptch1, Smo, Gli1, α-SMA, ALT, AST, Total bilirubin Increase: Albumin | [164] | |
| LPS-induced human hepatic stellate cell line LX-2 | 1.0–5.0 μM | None | Decrease: Cell activation, Cell proliferation Increase: FGF19, FGFR4 | [165] | |
| TGF-β1-induced mouse immortalized stellate cell lines JS1 TGF-β1-induced human hepatic stellate cell lines LX2 | 10−5 M | None | Decrease: LC3B II, Collagen Ⅰ, α-SMA, Atg5 Increase: C-Caspase 3 | [166] | |
| Rats injected with dimethylnitrosamine TGF-β1-induced hepatic stellate cells from liver tissue and rats | 12.5 mg/kg 1 μM | Gavage administration | Decrease: MEF2, α-SMA, Collagen I | [167] |
| Pharmacology | Experimental Subject | Experimental Dose | Animal Experimental Drug Delivery Method | Effects | Ref. |
|---|---|---|---|---|---|
| Effects on the skin | Human immortalized cells HaCaT | 3.125, 6.25, 12.5 μMol/L | None | Decrease: Cell proliferation, Bcl-2 Increase: Apoptosis, Caspase3, Caspase9, Bax | [168] |
| Bleomycin-induced mice TGF-β-induced human skin fibroblasts | 10 mg/kg 12, 25, 50 μg/mL | Intraperitoneal injection | Decrease: Collagen content, Connective tissue growth factor, Profibrotic gene: FN1, Plasminogen activator inhibitor-1, α-SMA, phosphorylation of SMAD3, phosphorylation of ERK1/2 | [169] | |
| Imiquimod-induced mice psoriasis | 40 mg/kg | Intraperitoneal injection | Decrease: Values of psoriasis area, IL-22, IL-23, IL-17A, IL-1β, IL-6, MDA, Keratin markers (K16 and K17), pAkt/Akt, pPI3K/PI3K Increase: SOD, CAT | [170] |
| Combination Therapy with | Test Subject | Mechanism/Effect | Ref. |
|---|---|---|---|
| Ginsenoside Rg1 | Rat with ligature below the left descending coronary artery | Decrease: Infarct area, Inflammatory infiltration, Collagen volume fraction, Fibrosis, Collagen I/III, α-SMA, MMP-9 | [192] |
| High-fat-diet-induced mice | Decrease: TC, TG, LDL-c, Non-esterified fatty acid | [193] | |
| Sodium tanshinone IIA sulfonate | Foponate combined with LPS-induced THP-1 macrophages TGF-β1-induced MRC-5 cells | Decrease: IL-1β, TNF-α, α-SMA, Collagen 1α1, Fibronectin | [194] |
| Tanshinone IIA | Bovine serum albumin-induced myocardial necrosis in mice | Decrease: Creatine kinase, CK-MB, LDH, α-Hydroxybutyrate dehydrogenase, TNF-α, IL-6, IL-1β, AST | [195] |
| Hydroxysafflor yellow A | Oxygen–glucose deprivation/reperfusion-induced H9c2 cardiomyocyte | Decrease: TNF-α, IL-6, IL-1, MDA, LDH, Creatine kinase Increase: SOD | [196] |
| Magnesium salt Salvia Miltiorrhiza Polyphenolate Injection | HEK293T cells, human hepatocellular carcinoma Huh-7 cells, 293T cells stably overexpressing human-derived SAR-CoV-2 receptor protein ACE2 | Anti-SARS-CoV-2 | [197] |
| Dosage Form | Preparation | Test Subject | Drug Delivery Routes | Effects | Ref. |
|---|---|---|---|---|---|
| Nanosheet | Graphitic carbon nitride nanosheets were prepared using an alkali–chemical–ultrasonic-assisted exfoliation method, added with PEI aqueous solution to obtain PEI–graphitic carbon nitride nanosheets | Mouse fibroblast L929 Wild AB lineage zebrafish | None | Increase: Slow release performance, Drug delivery rate | [198] |
| Nanoparticles | Chitosan powder was added to deionized water, glacial acetic acid was added and dissolved by sonication, an aqueous solution of carboxymethyl panax notoginseng polysaccharides was added, and the precipitate was collected and lyophilized | Mouse breast cancer cells 4T1 Mouse cardiomyocytes H9C2 | None | Sustained release | [199] |
| Nanoparticles | Salvianolic acid B and poloxamer 188 were dissolved by sonication in distilled water; ice chips and 1.2% n-butyl cyanoacrylate were dissolved with an appropriate amount of acetone and added dropwise to the above solution | Mice | Intravenous injection | Increase: Brain targeting | [200] |
| Nanoparticles | Dexamethasone−salvianolic acid B conjugate 1 was dissolved in ethanol and added to phosphate-buffered saline (PBS). Subsequently, the ethanol was removed under vacuum to give conjugate 1 nanoparticles | Auditory hair cell line HEI-OC1 Zebrafish Cisplatin-induced guinea pig | Transtympanically administered | Decrease: Ototoxicity | [201] |
| Nanoparticles | Free salvianolic acid B embedding phospholipid complex nanoparticles, free salvianolic acid B, and salvianolic-acid-B–phospholipid complex-loaded nanoparticles | 4-nitroquinoline-N-oxide-induced mice | Orally administered | Decrease: Cell proliferation (Ki67 and PCNA) markers, Cell cycle (cyclin D1 and p16) markers | [202] |
| Composite porous scaffold | PLGA and β-TCP were prepared by dissolving in 1,4-dioxane, and salvianolic acid B was dissolved in 1,4-dioxane. Then, different volumes of salvianolic acid B solution were added and entirely dried in a freeze-dryer | EA-hy9.26 endothelial cells Rat subperiosteal dissection with elevation of all paraspinal muscles and periosteum from the posterior elements was performed to expose the L5 and L6 transverse processes of the rat, and implanted with the same size of PLGA/β-TCP | Intracorporeal implantation | Increase: Bony fusion, Osteogenesis, Migration, Angiogenesis | [203] |
| Composite scaffold | GO was ultrasonically dispersed in dioxane/water, the solution was added with PLA, and different amounts of salvianolic acid B were then added. The homogeneous solution was poured into a mold until gel formed and finally frozen | Osteoblast cells MC3T3-E1 | None | Increase: Bone regeneration, Blood vessel formation | [204] |
| Scaffold | GO/salvianolic acid B conjugates and GO were dissolved in ddH2O with silk fibroin, added to a mold, and placed in a freeze-dryer | EA-hy9.26 endothelial cell Rat BMSCs Rat cranial defect model implantation of scaffolds | Intracorporeal implantation | Increase: Cell migration, Osteogenesis, Angiogenesis, Bone regeneration | [205] |
| Scaffold | Chitosan was added to 2% acetic acid, and salvianolic acid B was dissolved in ethanol and added to the chitosan solution. Then, Ca (NO3)2 and KH2PO4 were added to prepare the salvianolic acid B–chitosan/hydroxyapatite precursor solution. The solution was poured into a plastic mold and freeze-dried | Rabbit radius defect MC3T3-E1 cells HUVECs | Intracorporeal implantation | Increase: Vessel area, Vessel size | [206] |
| Biomimetic composite scaffolds | GO and ASA were ultrasonically dispersed in a dioxane/water mixture, and PLA was successively added to the mixed solution. Then, salvianolic acid B was added. The solution was poured into the mold until the solution formed a gel and was then frozen | Pre-osteoblast cells MC3T3-E1 | None | Decrease: Platelet aggregation Increase: Cell proliferation | [207] |
| Hydrogel | Peptide was dissolved in PBS, and transglutaminase stock solution with calcium chloride and dithiothreitol was added to form a hydrogel. A polydopamine nanoparticle solution was added to salvianolic acid B solution | HUVEC Rat coronary artery ligation | Intramyocardial injection | Decrease: Ventricular remodeling | [208] |
| Hydrogel | 1% HAMA hydrogel as a carrier and co-encapsulated BMSCs and salvianolic acid B into the hydrogel | BMSCs Needling segments 7–10 of the rat caudal spine using a 21G puncture needle | Inject into the intervertebral disc | Decrease: Apoptosis, Nucleus pulposus loss, Annulus fibrosus rupture disorder | [209] |
| Hydrogel | Photoinitiator LAP was added to PBS and dissolved. Salvianolic acid B was dissolved in PBS and added to the methacrylate-based gelatin hydrogel precursor solution to form a gel | Needling segments 7–10 of the rat caudal spine using a 20G puncture needle | Inject into the intervertebral disc | Decrease: TLR4, NF-kB, MMP3, MMP13, ADAMTS4, ADAMTS5, Collagen II, Proteoglycan | [210] |
| Hydrogel | Phospholipids, cholesterol, and tanshinone IIA were dissolved in dichloromethane. Salvianolic acid B was dissolved in 1% glycine–hydrochloric acid buffer solution and added to the above solution. The lyophilized powder of tanshinone IIA-salvianolic acid B was redissolved in oxidized hyaluronic acid oxidized hyaluronic acid solution, added to N-succinyl chitosan, and stirred | Rat abdominal skin New Zealand rabbit | Ex vivo dermal administration | Increase: Transdermal absorption, Drug dermal retention | [211,212] |
| Liposomes | Glycol chemically coupled to the membrane penetrating peptide TAT dissolved in an organic solvent, salvianolic acid B was dissolved in a 1% glycine–hydrochloric acid buffer solution. The two-phase system was mixed to form a water-in-oil emulsion, and then extruded with liposome extruder | Human skin fibroblast (HSF) cells Rat | Ex vivo dermal administration | Decrease: Cell proliferation, Cell migration, Cell invasion | [213,214,215] |
| Liposomes | Soy lecithin, cholesterol, and tanshinone IIA were dissolved in chloroform. Salvianolic acid B was dissolved in 1% glycine–HCl buffer solution and added dropwise to tanshinone IIA liposomes, obtain the liposome suspension, and extruded with liposome extruder | Human skin fibroblast HSF cells Rat | Ex vivo dermal administration | Decrease: Cell proliferation, Cell migration, Cell invasion, Collagen I Increase: MMP2 | [216] |
| Liposomes | Cholesterol, distearoyl phosphatidylethanolamine–polyethylene glycol 2000, and egg yolk lecithin were dissolved in anhydrous ethanol, and salvianolic acid B was dissolved in ultrapure water. The anhydrous ethanol mixture was injected into the drug-containing ultrapure water | Mouse mammary carcinoma 4T1 cells and mouse embryonic fibroblast NIH3T3 cells injected into mice | Intravenous injection | Decrease: α-SMA, TGF-β1, Smad2, Smad3, pSmad2, pSmad3, Smad4 Increase: IFN-γ, TNF-α, IL-12α, IL-2, IL-4, IL-6, IL-10 | [217] |
| Lipid emulsion | The medium-chain triglycerides and natural lecithin were heated and sonicated in a water bath; the aqueous phase was added to the oil phase and sheared at high speed for 2 min, and placed in an ultrasonic cell disruptor with fixed power | Acetaminophen-induced mouse | Gavage administration | Decrease: MDA, ALT, AST | [218] |
| Dry powder | Upon dissolving salvianolic acid B and L-leucine in water, the spray would begin | Mouse embryonic fibroblast NIH-3T3 Bleomycin saline solution-induced rat | Noninvasive tracheal administration | Decrease: Collagen 1A1, Collagen 3A1, ICAM-1, FN, iNOS, Arg-1, Cell migration, MDA, MPO, TGF-β1, IL-1β, IL-4, IL-6, IL-18 Increase: IFN-γ, SOD | [219] |
| Microemulsion | The oil phase was mixed with a surfactant–cosurfactant mixture. And the mixture was titrated with PBS buffer until it turned turbid | Mice skin Imiquimod-induced mice | Applied to the dorsal skin | Decrease: IL-17A, IL-17F, IL-22, IL-23, PCNA | [220] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, G.; Chen, G.; Liu, W.; Ye, D.; Liu, X.; Liang, X.; Song, J. Salvianolic Acid B: A Review of Pharmacological Effects, Safety, Combination Therapy, New Dosage Forms, and Novel Drug Delivery Routes. Pharmaceutics 2023, 15, 2235. https://doi.org/10.3390/pharmaceutics15092235
He G, Chen G, Liu W, Ye D, Liu X, Liang X, Song J. Salvianolic Acid B: A Review of Pharmacological Effects, Safety, Combination Therapy, New Dosage Forms, and Novel Drug Delivery Routes. Pharmaceutics. 2023; 15(9):2235. https://doi.org/10.3390/pharmaceutics15092235
Chicago/Turabian StyleHe, Guannan, Guangfeng Chen, Weidong Liu, Dongxue Ye, Xuehuan Liu, Xiaodong Liang, and Jing Song. 2023. "Salvianolic Acid B: A Review of Pharmacological Effects, Safety, Combination Therapy, New Dosage Forms, and Novel Drug Delivery Routes" Pharmaceutics 15, no. 9: 2235. https://doi.org/10.3390/pharmaceutics15092235