
Rigid Macrocycle Metal Complexes as CXCR4 Chemokine Receptor Antagonists: Influence of Ring Size

 , , ,
, , ,  , ,
, ,  , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Synthesis of Ligands L1–L5
2.1.1. Synthesis of p-Linked Homocyclen-Glyoxal (3)
2.1.2. Synthesis of 5
2.1.3. Synthesis of 6
2.1.4. Synthesis of L4
2.1.5. Synthesis of L5
2.1.6. Preparation of HCl Salts of L4 and L5
2.2. Synthesis of Cu(II) and Zn(II) Complexes of L1–L5
2.2.1. Synthesis of Zn2L3
2.2.2. Synthesis of Cu2L4
2.2.3. Synthesis of Zn2L4
2.2.4. Synthesis of Cu2L5
2.2.5. Synthesis of Zn2L5
2.3. Biological Evaluation
2.3.1. General
2.3.2. HIV Infection Assay
2.3.3. Anti-CXCR4 Antibody (Clone 12G5) Binding Inhibition Assay
2.3.4. Chemokine (CXCL12AF647/CCL5AF647) Binding Inhibition Assay
2.3.5. Measurement of Intracellular Chemokine-Induced Calcium Mobilisation
2.3.6. Evaluation of Cellular Cytotoxicity
2.4. X-ray Crystallography
2.5. Computational Methods
3. Results and Discussion
3.1. Synthesis of Ligands and Metal Complexes
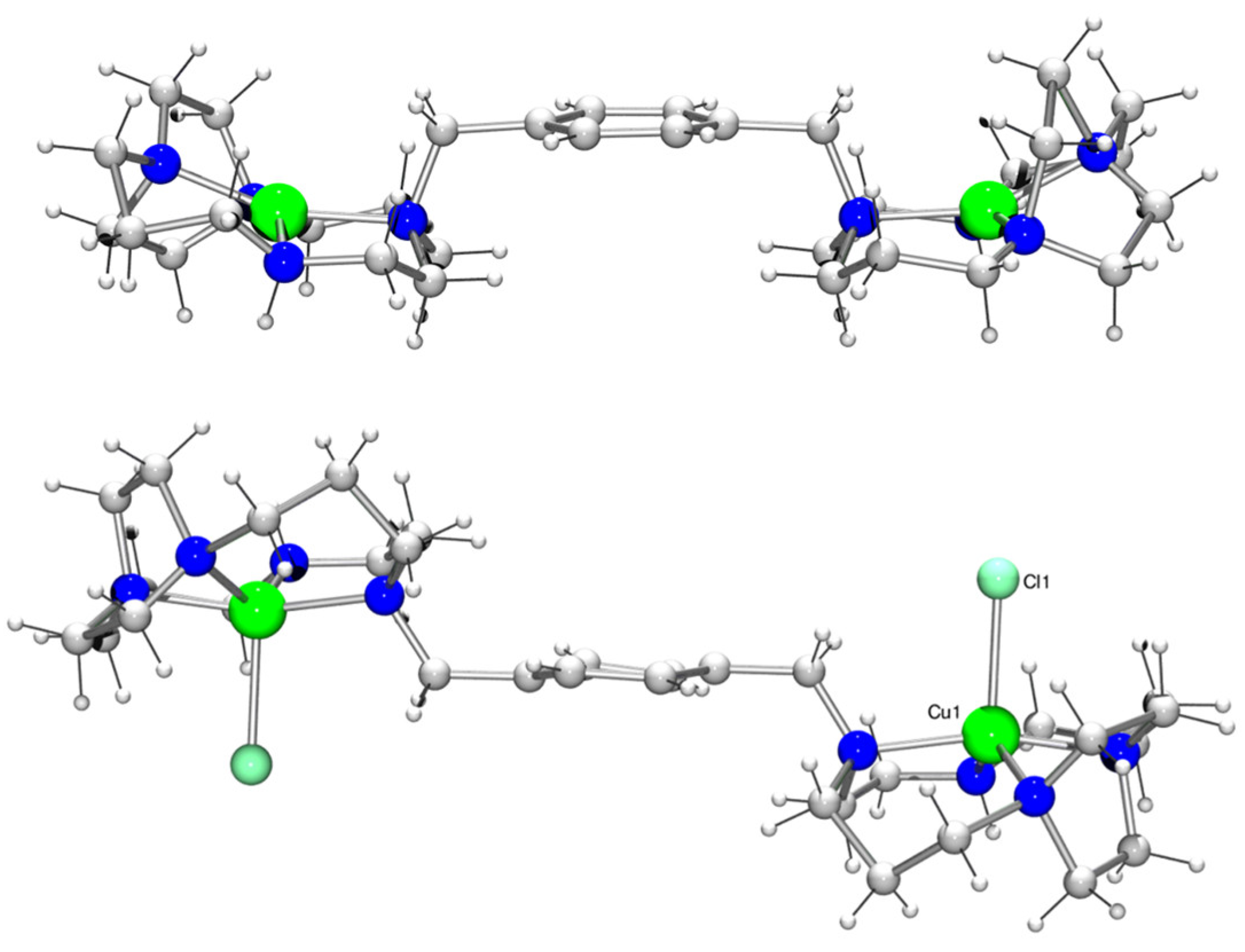
3.2. Crystallographic and Spectroscopic Characterisation of Copper(II) Complexes
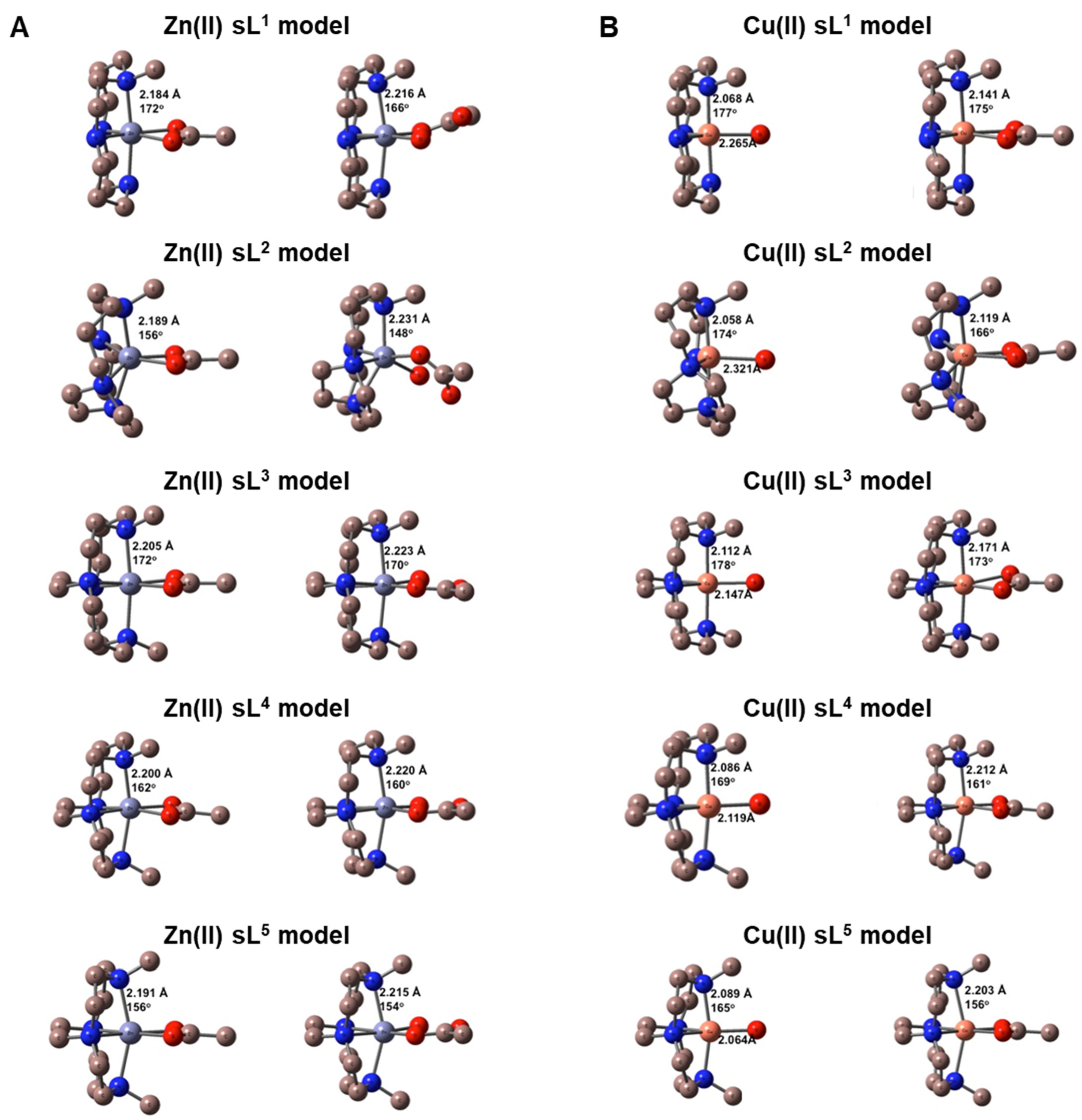
3.3. DFT Calculations
3.4. Biological Evaluation
3.4.1. Assessment of Cellular Cytotoxicity for the Investigated Compounds
3.4.2. Antiviral Activity against HIV-1 NL4-3 (X4) and BaL (R5) Replication
3.4.3. Inhibition of Anti-CXCR4 Monoclonal Antibody (Clone 12G5) Binding
3.4.4. Potent Antagonism of Natural Chemokine Ligand Interaction to CXCR4 and Receptor Selectivity Against CCR5
3.4.5. Inhibition of Chemokine Induced Signalling via CXCR4 or CCR5
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Murdoch, C.; Finn, A. Chemokine receptors and their role in inflammation and infectious diseases. Blood 2000, 95, 3032–3043. [Google Scholar] [CrossRef] [PubMed]
- Nagarsheth, N.; Wicha, M.S.; Zou, W.P. Chemokines in the cancer microenvironment and their relevance in cancer immunotherapy. Nat. Rev. Immunol. 2017, 17, 559–572. [Google Scholar] [CrossRef] [PubMed]
- Adlere, I.; Caspar, B.; Arimont, M.; Dekkers, S.; Visser, K.; Stuijt, J.; de Graaf, C.; Stocks, M.; Kellam, B.; Briddon, S.; et al. Modulators of CXCR4 and CXCR7/ACKR3 Function. Mol. Pharmacol. 2019, 96, 737–752. [Google Scholar] [CrossRef]
- Garcia-Cuesta, E.M.; Santiago, C.A.; Vallejo-Diaz, J.; Juarranz, Y.; Rodriguez-Frade, J.M.; Mellado, M. The Role of the CXCL12/CXCR4/ACKR3 Axis in Autoimmune Diseases. Front. Endocrinol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Miao, M.; De Clercq, E.; Li, G.D. Clinical significance of chemokine receptor antagonists. Expert Opin. Drug Metab. Toxicol. 2020, 16, 11–30. [Google Scholar] [CrossRef] [PubMed]
- Zeelenberg, I.S.; Ruuls-Van Stalle, L.; Roos, E. The chemokine receptor CXCR4 is required for outgrowth of colon carcinoma micrometastases. Cancer Res. 2003, 63, 3833–3839. [Google Scholar] [PubMed]
- Domanska, U.M.; Kruizinga, R.C.; Nagengast, W.B.; Timmer-Bosscha, H.; Huls, G.; de Vries, E.G.; Walenkamp, A.M. A review on CXCR4/CXCL12 axis in oncology: No place to hide. Eur. J. Cancer 2013, 49, 219–230. [Google Scholar] [CrossRef]
- Hattermann, K.; Mentlein, R. An infernal trio: The chemokine CXCL12 and its receptors CXCR4 and CXCR7 in tumor biology. Ann. Anat. Anat. Anz. Off. Organ Anat. Ges. 2013, 195, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Li, J.; Lei, W.; Wang, H.; Ni, Y.; Liu, Y.; Yan, H.; Tian, Y.; Wang, Z.; Yang, Z.; et al. CXCL12-CXCR4/CXCR7 Axis in Cancer: From Mechanisms to Clinical Applications. Int J Biol Sci 2023, 19, 3341–3359. [Google Scholar] [CrossRef] [PubMed]
- Cojoc, M.; Peitzsch, C.; Trautmann, F.; Polishchuk, L.; Telegeev, G.D.; Dubrovska, A. Emerging targets in cancer management: Role of the CXCL12/CXCR4 axis. OncoTargets Ther. 2013, 6, 1347–1361. [Google Scholar] [CrossRef]
- Domanska, U.M.; Timmer-Bosscha, H.; Nagengast, W.B.; Oude Munnink, T.H.; Kruizinga, R.C.; Ananias, H.J.; Kliphuis, N.M.; Huls, G.; De Vries, E.G.; de Jong, I.J.; et al. CXCR4 inhibition with AMD3100 sensitizes prostate cancer to docetaxel chemotherapy. Neoplasia 2012, 14, 709–718. [Google Scholar] [CrossRef] [PubMed]
- Gagliardi, F.; Narayanan, A.; Reni, M.; Franzin, A.; Mazza, E.; Boari, N.; Bailo, M.; Zordan, P.; Mortini, P. The role of CXCR4 in highly malignant human gliomas biology: Current knowledge and future directions. Glia 2014, 62, 1015–1023. [Google Scholar] [CrossRef] [PubMed]
- Marechal, R.; Demetter, P.; Nagy, N.; Berton, A.; Decaestecker, C.; Polus, M.; Closset, J.; Deviere, J.; Salmon, I.; Van Laethem, J.L. High expression of CXCR4 may predict poor survival in resected pancreatic adenocarcinoma. Br J Cancer 2009, 100, 1444–1451. [Google Scholar] [CrossRef] [PubMed]
- Trautmann, F.; Cojoc, M.; Kurth, I.; Melin, N.; Bouchez, L.C.; Dubrovska, A.; Peitzsch, C. CXCR4 as biomarker for radioresistant cancer stem cells. Int. J. Radiat. Biol. 2014, 90, 687–699. [Google Scholar] [CrossRef] [PubMed]
- Werner, L.; Guzner-Gur, H.; Dotan, I. Involvement of CXCR4/CXCR7/CXCL12 Interactions in Inflammatory bowel disease. Theranostics 2013, 3, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Tahirovic, Y.A.; Pelly, S.; Jecs, E.; Miller, E.J.; Sharma, S.K.; Liotta, D.C.; Wilson, L.J. Small molecule and peptide-based CXCR4 modulators as therapeutic agents. A patent review for the period from 2010 to 2018. Expert Opin. Ther. Pat. 2020, 30, 87–101. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Zhao, H.; Chen, H.T.; Yao, Q.H. CXCR4 in breast cancer: Oncogenic role and therapeutic targeting. Drug Des. Dev. Ther. 2015, 9, 4953–4964. [Google Scholar] [CrossRef]
- Hermann, P.C.; Huber, S.L.; Herrler, T.; Aicher, A.; Ellwart, J.W.; Guba, M.; Bruns, C.J.; Heeschen, C. Distinct populations of cancer stem cells determine tumor growth and metastatic activity in human pancreatic cancer. Cell Stem Cell 2007, 1, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Panneerselvam, J.; Jin, J.K.; Shanker, M.; Lauderdale, J.; Bates, J.; Wang, Q.; Zhao, Y.D.; Archibald, S.J.; Hubin, T.J.; Ramesh, R. IL-24 Inhibits Lung Cancer Cell Migration and Invasion by Disrupting The SDF-1/CXCR4 Signaling Axis. Plos One 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Bao, S.; Darvishi, M.; H Amin, A.; Al-Haideri, M.T.; Patra, I.; Kashikova, K.; Ahmad, I.; Alsaikhan, F.; Al-qaim, Z.H.; Al-Gazally, M.E.; et al. CXC chemokine receptor 4 (CXCR4) blockade in cancer treatment. J. Cancer Res. Clin. Oncol. 2023, 149, 7945–7968. [Google Scholar] [CrossRef] [PubMed]
- Renard, I.; Archibald, S.J. CXCR4-targeted metal complexes for molecular imaging. In Medicinal Chemistry; Sadler, P.J., VanEldik, R., Eds.; 2020; Volume 75, pp. 447–476. [Google Scholar]
- Mayerhoefer, M.E.; Archibald, S.J.; Messiou, C.; Staudenherz, A.; Berzaczy, D.; Schoder, H. MRI and PET/MRI in hematologic malignancies. J. Magn. Reson. Imaging 2020, 51, 1325–1335. [Google Scholar] [CrossRef] [PubMed]
- Burke, B.; Miranda, C.; Lee, R.; Renard, I.; Nigam, S.; Clemente, G.; D’Huys, T.; Ruest, T.; Domarkas, J.; Thompson, J.; et al. Copper-64 PET imaging of the CXCR4 chemokine receptor using a cross-bridged cyclam bis-tetraazamacrocyclic antagonist. J. Nucl. Med. 2019. [Google Scholar] [CrossRef]
- De Clercq, E. The bicyclam AMD3 100 story. Nat. Rev. Drug Discov. 2003, 2, 581–587. [Google Scholar] [CrossRef] [PubMed]
- Amendola, V.; Bergamaschi, G.; Fabbrizzi, L.; Licchelli, M.; Mangano, C. The interaction of Mozobil (TM) with carboxylates. Org. Biomol. Chem. 2016, 14, 905–912. [Google Scholar] [CrossRef] [PubMed]
- Gerlach, L.O.; Skerlj, R.T.; Bridger, G.J.; Schwartz, T.W. Molecular interactions of cyclam and bicyclam non-peptide antagonists with the CXCR4 chemokine receptor. J Biol Chem 2001, 276, 14153–14160. [Google Scholar] [CrossRef] [PubMed]
- Rosenkilde, M.M.; Gerlach, L.-O.; Jakobsen, J.S.; Skerlj, R.T.; Bridger, G.J.; Schwartz, T.W. Molecular Mechanism of AMD3100 Antagonism in the CXCR4 Receptor: TRANSFER OF BINDING SITE TO THE CXCR3 RECEPTOR*. J. Biol. Chem. 2004, 279, 3033–3041. [Google Scholar] [CrossRef]
- Wong, R.S.; Bodart, V.; Metz, M.; Labrecque, J.; Bridger, G.; Fricker, S.P. Comparison of the potential multiple binding modes of bicyclam, monocylam, and noncyclam small-molecule CXC chemokine receptor 4 inhibitors. Mol. Pharmacol. 2008, 74, 1485–1495. [Google Scholar] [CrossRef] [PubMed]
- Gerlach, L.O.; Jakobsen, J.S.; Jensen, K.P.; Rosenkilde, M.R.; Skerlj, R.T.; Ryde, U.; Bridger, G.J.; Schwartz, T.W. Metal ion enhanced binding of AMD3100 to Asp(262) in the CXCR4 receptor. Biochemistry 2003, 42, 710–717. [Google Scholar] [CrossRef] [PubMed]
- Hunter, T.M.; McNae, I.W.; Liang, X.Y.; Bella, J.; Parsons, S.; Walkinshaw, M.D.; Sadler, P.J. Protein recognition of macrocycles: Binding of anti-HIV metallocyclams to lysozyme. Proc. Natl. Acad. Sci. USA 2005, 102, 2288–2292. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.Y.; Parkinson, J.A.; Weishaupl, M.; Gould, R.O.; Paisey, S.J.; Park, H.S.; Hunter, T.M.; Blindauer, C.A.; Parsons, S.; Sadler, P.J. Structure and dynamics of metallomacrocycles: Recognition of zinc xylyl-bicyclam by an HIV coreceptor. J. Am. Chem. Soc. 2002, 124, 9105–9112. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.Y.; Weishaupl, M.; Parkinson, J.A.; Parsons, S.; McGregor, P.A.; Sadler, P.J. Selective recognition of configurational substates of zinc cyclam by carboxylates: Implications for the design and mechanism of action of anti-HIV agents. Chem.-A Eur. J. 2003, 9, 4709–4717. [Google Scholar] [CrossRef] [PubMed]
- Valks, G.C.; McRobbie, G.; Lewis, E.A.; Hubin, T.J.; Hunter, T.M.; Sadler, P.J.; Pannecouque, C.; De Clercq, E.; Archibald, S.J. Configurationally Restricted Bismacrocyclic CXCR4 Receptor Antagonists. J. Med. Chem. 2006, 49, 6162–6165. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Nicholson, G.; Greenman, J.; Madden, L.; McRobbie, G.; Pannecouque, C.; De Clercq, E.; Ullom, R.; Maples, D.L.; Maples, R.D.; et al. Binding Optimization through Coordination Chemistry: CXCR4 Chemokine Receptor Antagonists from Ultrarigid Metal Complexes. J. Am. Chem. Soc. 2009, 131, 3416–3417. [Google Scholar] [CrossRef] [PubMed]
- Cain, A.N.; Freeman, T.N.C.; Roewe, K.D.; Cockriel, D.L.; Hasley, T.R.; Maples, R.D.; Allbritton, E.M.A.; D’Huys, T.; van Loy, T.; Burke, B.P.; et al. Acetate as a model for aspartate-based CXCR4 chemokine receptor binding of cobalt and nickel complexes of cross-bridged tetraazamacrocycles. Dalton Trans. 2019, 48, 2785–2801. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.; Huskens, D.; Daelemans, D.; Mewis, R.E.; Garcia, C.D.; Cain, A.N.; Freeman, T.N.C.; Pannecouque, C.; De Clercq, E.; Schols, D.; et al. CXCR4 chemokine receptor antagonists: Nickel(II) complexes of configurationally restricted macrocycles. Dalton Trans. 2012, 41, 11369–11377. [Google Scholar] [CrossRef] [PubMed]
- Alzahrani, S.O.; McRobbie, G.; Khan, A.; D’Huys, T.; Van Loy, T.; Walker, A.N.; Renard, I.; Hubin, T.J.; Schols, D.; Burke, B.P.; et al. Trans-IV restriction: A new configuration for metal bis-cyclam complexes as potent CXCR4 inhibitors. Dalton Trans 2024, 53, 5616–5623. [Google Scholar] [CrossRef] [PubMed]
- Baghdadi, N.E.; Burke, B.P.; Alresheedi, T.; Nigam, S.; Saeed, A.; Almutairi, F.; Domarkas, J.; Khan, A.; Archibald, S.J. Multivalency in CXCR4 chemokine receptor targeted iron oxide nanoparticles. Dalton Trans. 2021, 50, 1599–1603. [Google Scholar] [CrossRef] [PubMed]
- McRobbie, G. Configurationally Restricted Bis-Tetraazamacrocyclic Complexes: Chemokine Receptor Antagonists. Ph.D. Thesis, University of Hull, UK, 2009. [Google Scholar]
- D’Huys, T. Biological Characterization of Novel CXCR4 and CXCR7/ACKR3 Receptor Inhibitors. Ph.D. Thesis, KU Leuven (Katholieke Universiteit Leuven), Leuven, Belgium, 2018. [Google Scholar]
- Brandes, S.; Gros, C.; Denat, F.; Pullumbi, P.; Guilard, R. New facile and convenient synthesis of bispolyazamacrocycles using Boc protection. Determination of geometric parameters of dinuclear copper(II) complexes using ESR spectroscopy and molecular mechanics calculations. Bull. Soc. Chim. Fr. 1996, 133, 65–73. [Google Scholar]
- Bridger, G.J.; Skerlj, R.T.; Thornton, D.; Padmanabhan, S.; Martellucci, S.A.; Henson, G.W.; Abrams, M.J.; Yamamoto, N.; Devreese, K.; Pauwels, R.; et al. Synthesis and Structure-Activity-Relationships of Phenylenebis(Methylene)-Linked Bis-Tetraazamacrocycles That Inhibit HIV Replication - Effects of Macrocyclic Ring Size and Substituents on the Aromatic Linker. J. Med. Chem. 1995, 38, 366–378. [Google Scholar] [CrossRef] [PubMed]
- McRobbie, G.; Valks, G.C.; Empson, C.J.; Khan, A.; Silversides, J.D.; Pannecouque, C.; De Clercq, E.; Fiddy, S.G.; Bridgeman, A.J.; Young, N.A.; et al. Probing key coordination interactions: Configurationally restricted metal activated CXCR4 antagonists. Dalton Trans. 1039. [Google Scholar] [CrossRef] [PubMed]
- Weisman, G.R.; Ho, S.C.H.; Johnson, V. Tetracyclic tetraamines by glyoxal-macrocyclic tetraamine condensation. Tetrahedron Lett. 1980, 21, 335–338. [Google Scholar] [CrossRef]
- Le Baccon, M.; Chuburu, F.; Toupet, L.; Handel, H.; Soibinet, M.; Dechamps-Olivier, I.; Barbier, J.-P.; Aplincourt, M. Bis-aminals: Efficient tools for bis-macrocycle synthesis. New J. Chem. 2001, 25, 1168–1174. [Google Scholar] [CrossRef]
- Björndal, A.; Deng, H.; Jansson, M.; Fiore, J.R.; Colognesi, C.; Karlsson, A.; Albert, J.; Scarlatti, G.; Littman, D.R.; Fenyö, E.M. Coreceptor usage of primary human immunodeficiency virus type 1 isolates varies according to biological phenotype. J Virol 1997, 71, 7478–7487. [Google Scholar] [CrossRef] [PubMed]
- Schols, D.; Esté José, A.; Cabrera, C.; De Clercq, E. T-Cell-Line-Tropic Human Immunodeficiency Virus Type 1 That Is Made Resistant to Stromal Cell-Derived Factor 1α Contains Mutations in the Envelope gp120 but Does Not Show a Switch in Coreceptor Use. J. Virol. 1998, 72, 4032–4037. [Google Scholar] [CrossRef] [PubMed]
- Princen, K.; Hatse, S.; Vermeire, K.; De Clercq, E.; Schols, D. Establishment of a novel CCR5 and CXCR4 expressing CD4+ cell line which is highly sensitive to HIV and suitable for high-throughput evaluation of CCR5 and CXCR4 antagonists. Retrovirology 2004, 1, 2. [Google Scholar] [CrossRef] [PubMed]
- Montefiori, D.C. Measuring HIV Neutralization in a Luciferase Reporter Gene Assay. In Methods in Molecular Biology; Prasad, V.R., Kalpana, G.V., Eds.; 2009; Volume 485, pp. 395–405. [Google Scholar]
- X-AREA v 1.64. STOE & Cie GmbH: Darmstadt, Germany, 2012.
- Sheldrick, G.M. A short history of SHELX. Acta Crystallogr. Sect. A: Found. Crystallogr. 2008, 64, 112–122. [Google Scholar] [CrossRef] [PubMed]
- Gaussian 09; Gaussian, Inc.: Wallingford CT, USA, 2009.
- Zhao, Y.; Truhlar, D.G. The M06 suite of density functionals for main group thermochemistry, thermochemical kinetics, noncovalent interactions, excited states, and transition elements: Two new functionals and systematic testing of four M06-class functionals and 12 other functionals. Theor. Chem. Acc. 2008, 120, 215–241. [Google Scholar] [CrossRef]
- Kowallick, R.; Neuburger, M.; Zehnder, M.; Kaden, T.A. Metal complexes with macrocyclic ligands.45. Axial coordination tendency in reinforced tetraazamacrocyclic complexes. Helv. Chim. Acta 1997, 80, 948–959. [Google Scholar] [CrossRef]
- Silversides, J.D.; Allan, C.C.; Archibald, S.J. Copper (II) cyclam-based complexes for radiopharmaceutical applications: Synthesis and structural analysis. Dalton Trans. 2007, 971–978. [Google Scholar] [CrossRef] [PubMed]
- Maples, R.D.; Cain, A.N.; Burke, B.P.; Silversides, J.D.; Mewis, R.E.; D’Huys, T.; Schols, D.; Linder, D.P.; Archibald, S.J.; Hubin, T.J. Aspartate-Based CXCR4 Chemokine Receptor Binding of Cross-Bridged Tetraazamacrocyclic Copper(II) and Zinc(II) Complexes. Chem. -A Eur. J. 2016, 22, 12916–12930. [Google Scholar] [CrossRef] [PubMed]
- Schols, D.; Este, J.A.; Henson, G.; DeClercq, E. Bicyclams, a class of potent anti-HIV agents, are targeted at the HIV coreceptor Fusin/CXCR-4. Antivir. Res. 1997, 35, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Schols, D.; Struyf, S.; VanDamme, J.; Este, J.A.; Henson, G.; DeClercq, E. Inhibition of T-tropic HIV strains by selective antagonization of the chemokine receptor CXCR4. J. Exp. Med. 1997, 186, 1383–1388. [Google Scholar] [CrossRef] [PubMed]
- McKnight, A.; Wilkinson, D.; Simmons, G.; Talbot, S.; Picard, L.; Ahuja, M.; Marsh, M.; Hoxie, J.A.; Clapham, P.R. Inhibition of human immunodeficiency virus fusion by a monoclonal antibody to a coreceptor (CXCR4) is both cell type and virus strain dependent. J. Virol. 1997, 71, 1692–1696. [Google Scholar] [CrossRef] [PubMed]
- Carnec, X.; Quan, L.; Olson, W.C.; Hazan, U.; Dragic, T. Anti-CXCR4 monoclonal antibodies recognizing overlapping epitopes differ significantly in their ability to inhibit entry of human immunodeficiency virus type 1. J. Virol. 2005, 79, 1930–1933. [Google Scholar] [CrossRef] [PubMed]
- Hatse, S.; Princen, K.; Liekens, S.; Vermeire, K.; De Clercq, E.; Schols, D. Fluorescent CXCL12(AF647) as a novel probe for nonradioactive CXCL12/CXCR4 cellular interaction studies. Cytom. Part A 2004, 61A, 178–188. [Google Scholar] [CrossRef] [PubMed]
- Schoofs, G.; Van Hout, A.; D’Huys, T.; Schols, D.; Van Loy, T. A Flow Cytometry-based Assay to Identify Compounds That Disrupt Binding of Fluorescently-labeled CXC Chemokine Ligand 12 to CXC Chemokine Receptor 4. J. Vis. Exp. 2018. [Google Scholar] [CrossRef]
- Princen, K.; Hatse, S.; Vermeire, K.; De Clercq, E.; Schols, D. Evaluation of SDF-1/CXCR4-induced Ca2+ signaling by fluorometric imaging plate reader (FLIPR) and flow cytometry. Cytom. Part A 2003, 51A, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Wiesmann, N.; Tremel, W.; Brieger, J. Zinc oxide nanoparticles for therapeutic purposes in cancer medicine. J. Mater. Chem. B 2020, 8, 4973–4989. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Anti-HIV Activity a IC50 (nM) | Toxicity b CC50 (µM) | |
|---|---|---|---|
| NL4-3 (X4) | BaL (R5) | PMBCs | |
| Cu2L1 | 10.1 ± 1.6 | >20,000 | >100 |
| Zn2L1 | 0.73 ± 0.34 | >15,000 | >90 |
| Cu2L2 | 12.9 ± 2.8 | >20,000 | >100 |
| Zn2L2 | 2.9 ± 1.2 | >15,000 | >100 |
| Cu2L3 | 0.74 ± 0.60 | >5000 | >40 |
| Zn2L3 | 6.2 ± 2.5 | >1000 | >100 |
| Cu2L4 | 0.87 ± 0.32 | >2000 | >10 |
| Zn2L4 | 4.8 ± 1.5 | >2000 | >90 |
| Cu2L5 | 31.6 ± 2.4 | >20,000 | >30 |
| Zn2L5 | 16.9 ± 0.9 | >20,000 | >100 |
| AMD3100 | 0.42 ± 0.14 | >20,000 | >100 |
| Maraviroc | >20,000 | 2.9 ± 1.2 | >100 |
| Compound | Competition Binding Assay a IC50 (nM) | Ca2+ Signalling Assay b IC50 (nM) | |||
|---|---|---|---|---|---|
| 12G5 | CXCL12 | CCL5 | CXCL12 | CCL3L1 | |
| Cu2L1 | 50.1 ± 7.2 | 5.1 ± 0.9 | >1000 | 113 ± 12.7 | >20,000 |
| Zn2L1 | 5.9 ± 2.4 | 0.41 ± 0.03 | >1000 | 4.9 ± 2.9 | >2000 |
| Cu2L2 | 465 ± 155 | 8.9 ± 1.1 | >1000 | 185 ± 75 | >20,000 |
| Zn2L2 | 1.4 ± 0.4 | 0.43 ± 0.00 | 255 ± 58 | 5.6 ± 1.9 | 120 ± 28 |
| Cu2L3 | 1.12 ± 0.15 | 0.54 ± 0.03 | 741 ± 145 | 3.1 ± 0.3 [23] | >1000 |
| Zn2L3 | 1.9 ± 0.3 | 0.90 ± 0.23 | >1000 | 17.5 ± 11.7 | >5000 |
| Cu2L4 | 1.3 ± 0.4 | 0.53 ± 0.03 | 702 ± 219 | 11.9 ± 8.3 | >10,000 |
| Zn2L4 | 3.4 ± 0.2 | 2.1 ± 0.1 | >1000 | 66.1 ± 0.9 | >20,000 |
| Cu2L5 | 1.9 ± 0.6 | 0.83 ± 0.02 | >1000 | 11.3 ± 4.7 | >15,000 |
| Zn2L5 | 8.6 ± 4.7 | 0.49 ± 0.01 | >1000 | 10.3 ± 6.6 | >20,000 |
| AMD3100 | 83.7 ± 8.3 | 18.0 ± 4.2 [37] | >1000 | 203.5 ± 19.4 [23] | >20,000 |
| Maraviroc | - | >1000 | 10.3 ± 3.5 | >20,000 | 3.1 ± 0.6 |
| Single Ring Chelator Model | ΔG298 (kJ/mol) [CusL(H2O)]2+ + OAc– → [CusL(OAc)]+ + H2O | ΔG298 (kJ/mol) [ZnsL(OAc)]+ + H2O → [ZnsL(OAc)(H2O)]+ | ΔG298 (kJ/mol) [ZnsL(H2O)2]2+ + OAc– → [ZnsL(OAc)(H2O)]+ + H2O |
|---|---|---|---|
| sL1 | −822.2 | −14.9 | −825.8 |
| sL2 | −800.4 | 4.0 | − |
| sL3 | −833.5 | −25.1 | −830.6 |
| sL4 | −845.9 | −23.5 | −840.4 |
| sL5 | −866.7 | −22.5 | −849.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Renard, I.; D’huys, T.; Burke, B.P.; Ajoleza, T.; Cain, A.N.; Funwie, N.L.; Khan, A.; Maples, D.L.; Maples, R.D.; Matz, D.L.; et al. Rigid Macrocycle Metal Complexes as CXCR4 Chemokine Receptor Antagonists: Influence of Ring Size. Pharmaceutics 2024, 16, 1000. https://doi.org/10.3390/pharmaceutics16081000
Renard I, D’huys T, Burke BP, Ajoleza T, Cain AN, Funwie NL, Khan A, Maples DL, Maples RD, Matz DL, et al. Rigid Macrocycle Metal Complexes as CXCR4 Chemokine Receptor Antagonists: Influence of Ring Size. Pharmaceutics. 2024; 16(8):1000. https://doi.org/10.3390/pharmaceutics16081000
Chicago/Turabian StyleRenard, Isaline, Thomas D’huys, Benjamin P. Burke, Trisha Ajoleza, Amy N. Cain, Neil L. Funwie, Abid Khan, Danny L. Maples, Randall D. Maples, Dallas L. Matz, and et al. 2024. "Rigid Macrocycle Metal Complexes as CXCR4 Chemokine Receptor Antagonists: Influence of Ring Size" Pharmaceutics 16, no. 8: 1000. https://doi.org/10.3390/pharmaceutics16081000
APA StyleRenard, I., D’huys, T., Burke, B. P., Ajoleza, T., Cain, A. N., Funwie, N. L., Khan, A., Maples, D. L., Maples, R. D., Matz, D. L., McRobbie, G., Ullom, R., Prior, T. J., Linder, D. P., Van Loy, T., Hubin, T. J., Schols, D., & Archibald, S. J. (2024). Rigid Macrocycle Metal Complexes as CXCR4 Chemokine Receptor Antagonists: Influence of Ring Size. Pharmaceutics, 16(8), 1000. https://doi.org/10.3390/pharmaceutics16081000