

Biomimetic Nucleic Acid Drug Delivery Systems for Relieving Tumor Immunosuppressive Microenvironment


Abstract

1. Introduction
2. Biomimetic Nucleic Acid Delivery System for Antitumor Immunotherapy
2.1. Virus-Derived Delivery Systems
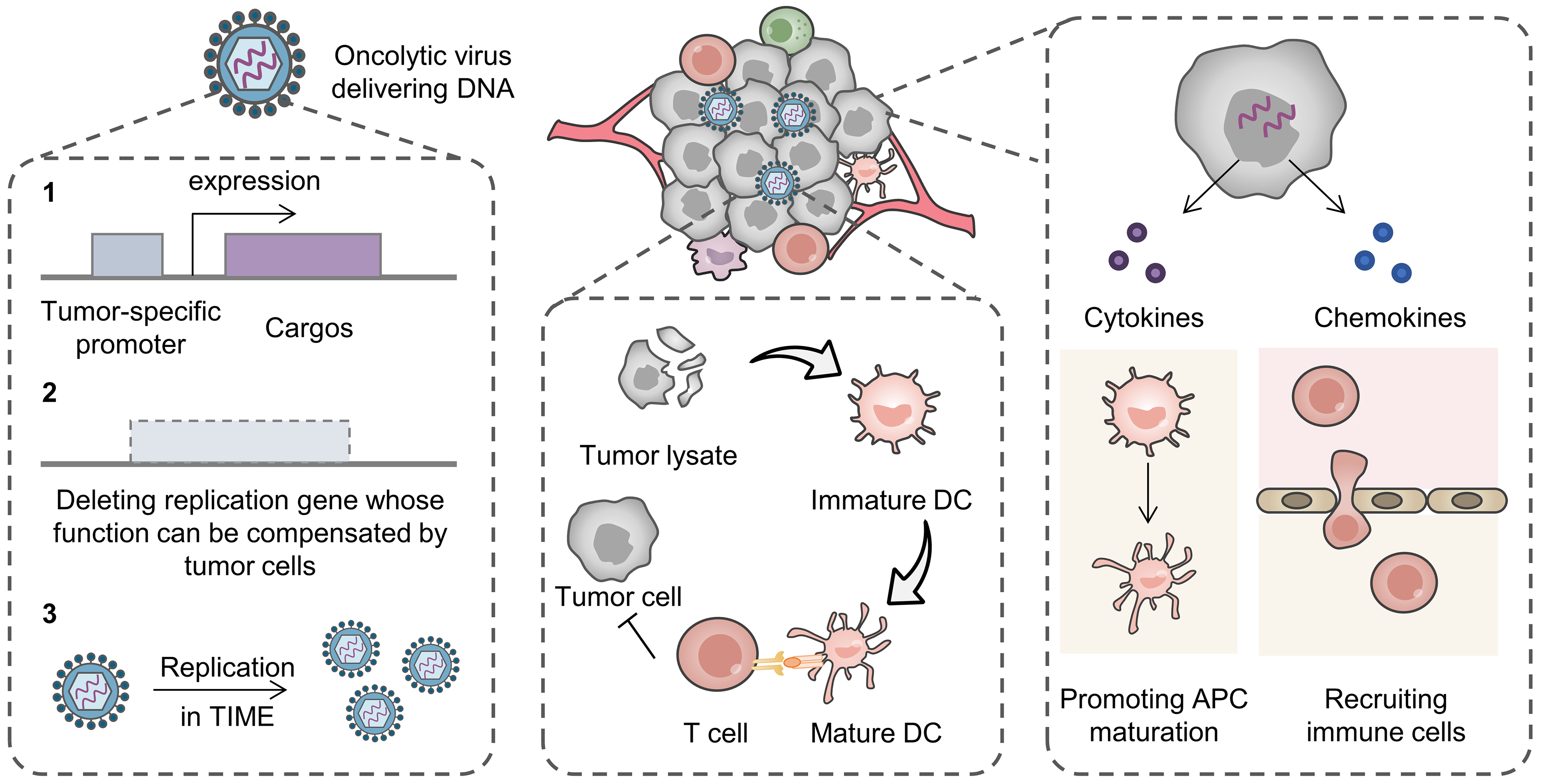
2.1.1. Virus
2.1.2. Virus-like Particles
2.2. Bacteria-Derived Delivery Systems
2.2.1. Bacteria
2.2.2. Bacteria-Derived Nanovesicle
2.3. Cell-Derived Delivery Systems
2.3.1. Cells
2.3.2. Cell Membrane-Coated Nanoparticles
2.3.3. Extracellular Vesicles
3. Conclusions and Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Waldman, A.D.; Fritz, J.M.; Lenardo, M.J. A guide to cancer immunotherapy: From T cell basic science to clinical practice. Nat. Rev. Immunol. 2020, 20, 651–668. [Google Scholar] [CrossRef] [PubMed]
- Hiam-Galvez, K.J.; Allen, B.M.; Spitzer, M.H. Systemic immunity in cancer. Nat. Rev. Cancer 2021, 21, 345–359. [Google Scholar] [CrossRef] [PubMed]
- Dunn, G.P.; Bruce, A.T.; Ikeda, H.; Old, L.J.; Schreiber, R.D. Cancer immunoediting: From immunosurveillance to tumor escape. Nat. Immunol. 2002, 3, 991–998. [Google Scholar] [CrossRef] [PubMed]
- Łuksza, M.; Riaz, N.; Makarov, V.; Balachandran, V.P.; Hellmann, M.D.; Solovyov, A.; Rizvi, N.A.; Merghoub, T.; Levine, A.J.; Chan, T.A.; et al. A neoantigen fitness model predicts tumour response to checkpoint blockade immunotherapy. Nature 2017, 551, 517–520. [Google Scholar] [CrossRef] [PubMed]
- Sanmamed, M.F.; Chen, L. A paradigm shift in cancer immunotherapy: From enhancement to normalization. Cell 2018, 175, 313–326. [Google Scholar] [CrossRef] [PubMed]
- Scott, E.C.; Baines, A.C.; Gong, Y.; Moore, R., Jr.; Pamuk, G.E.; Saber, H.; Subedee, A.; Thompson, M.D.; Xiao, W.; Pazdur, R.; et al. Trends in the approval of cancer therapies by the FDA in the twenty-first century. Nat. Rev. Drug Discov. 2023, 22, 625–640. [Google Scholar] [CrossRef]
- Sharma, P.; Siddiqui, B.A.; Anandhan, S.; Yadav, S.S.; Subudhi, S.K.; Gao, J.; Goswami, S.; Allison, J.P. The next decade of immune checkpoint therapy. Cancer Discov. 2021, 11, 838–857. [Google Scholar] [CrossRef] [PubMed]
- Doroshow, D.B.; Bhalla, S.; Beasley, M.B.; Sholl, L.M.; Kerr, K.M.; Gnjatic, S.; Wistuba, I.I.; Rimm, D.L.; Tsao, M.S.; Hirsch, F.R. PD-L1 as a biomarker of response to immune-checkpoint inhibitors. Nat. Rev. Clin. Oncol. 2021, 18, 345–362. [Google Scholar] [CrossRef] [PubMed]
- de Miguel, M.; Calvo, E. Clinical challenges of immune checkpoint inhibitors. Cancer Cell 2020, 38, 326–333. [Google Scholar] [CrossRef]
- Paunovska, K.; Loughrey, D.; Dahlman, J.E. Drug delivery systems for RNA therapeutics. Nat. Rev. Genet. 2022, 23, 265–280. [Google Scholar] [CrossRef]
- Tang, Q.; Khvorova, A. RNAi-based drug design: Considerations and future directions. Nat. Rev. Drug Discov. 2024, 23, 341–364. [Google Scholar] [CrossRef] [PubMed]
- Huayamares, S.G.; Loughrey, D.; Kim, H.; Dahlman, J.E.; Sorscher, E.J. Nucleic acid-based drugs for patients with solid tumours. Nat. Rev. Clin. Oncol. 2024, 21, 407–427. [Google Scholar] [CrossRef] [PubMed]
- Katti, A.; Diaz, B.J.; Caragine, C.M.; Sanjana, N.E.; Dow, L.E. CRISPR in cancer biology and therapy. Nat. Rev. Cancer 2022, 22, 259–279. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, J.A.; Witzigmann, D.; Thomson, S.B.; Chen, S.; Leavitt, B.R.; Cullis, P.R.; van der Meel, R. The current landscape of nucleic acid therapeutics. Nat. Nanotechnol. 2021, 16, 630–643. [Google Scholar] [CrossRef]
- Lin, Y.X.; Wang, Y.; Blake, S.; Yu, M.; Mei, L.; Wang, H.; Shi, J. RNA nanotechnology-mediated cancer immunotherapy. Theranostics 2020, 10, 281–299. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.; Seok, S.H.; Yoon, H.Y.; Ryu, J.H.; Kwon, I.C. Advancing cancer immunotherapy through siRNA-based gene silencing for immune checkpoint blockade. Adv. Drug Deliv. Rev. 2024, 209, 115306. [Google Scholar] [CrossRef]
- Hattab, D.; Gazzali, A.M.; Bakhtiar, A. Clinical advances of siRNA-based nanotherapeutics for cancer treatment. Pharmaceutics 2021, 13, 1009. [Google Scholar] [CrossRef]
- Liu, C.; Shi, Q.; Huang, X.; Koo, S.; Kong, N.; Tao, W. mRNA-based cancer therapeutics. Nat. Rev. Cancer 2023, 23, 526–543. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, N.; Weissman, D.; Whitehead, K.A. mRNA vaccines for infectious diseases: Principles, delivery and clinical translation. Nat. Rev. Drug Discov. 2021, 20, 817–838. [Google Scholar] [CrossRef]
- Zhang, J.; Hu, Y.; Yang, J.; Li, W.; Zhang, M.; Wang, Q.; Zhang, L.; Wei, G.; Tian, Y.; Zhao, K.; et al. Non-viral, specifically targeted CAR-T cells achieve high safety and efficacy in B-NHL. Nature 2022, 609, 369–374. [Google Scholar] [CrossRef]
- Stadtmauer, E.A.; Fraietta, J.A.; Davis, M.M.; Cohen, A.D.; Weber, K.L.; Lancaster, E.; Mangan, P.A.; Kulikovskaya, I.; Gupta, M.; Chen, F.; et al. CRISPR-engineered T cells in patients with refractory cancer. Science 2020, 367, eaba7365. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Jiang, M.; Yu, W.; Xu, Z.; Liu, X.; Jia, Q.; Guan, X.; Zhang, W. CpG-based nanovaccines for cancer immunotherapy. Int. J. Nanomed. 2021, 16, 5281–5299. [Google Scholar] [CrossRef] [PubMed]
- Dongye, Z.; Li, J.; Wu, Y. Toll-like receptor 9 agonists and combination therapies: Strategies to modulate the tumour immune microenvironment for systemic anti-tumour immunity. Br. J. Cancer 2022, 127, 1584–1594. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.J.; Svensson-Arvelund, J.; Lubitz, G.S.; Marabelle, A.; Melero, I.; Brown, B.D.; Brody, J.D. Cancer vaccines: The next immunotherapy frontier. Nat. Cancer 2022, 3, 911–926. [Google Scholar] [CrossRef] [PubMed]
- Poon, W.; Kingston, B.R.; Ouyang, B.; Ngo, W.; Chan, W.C.W. A framework for designing delivery systems. Nat. Nanotechnol. 2020, 15, 819–829. [Google Scholar] [CrossRef] [PubMed]
- Mendes, B.B.; Conniot, J.; Avital, A.; Yao, D.; Jiang, X.; Zhou, X.; Sharf-Pauker, N.; Xiao, Y.; Adir, O.; Liang, H.; et al. Nanodelivery of nucleic acids. Nat. Rev. Methods Primers 2022, 2, 24. [Google Scholar] [CrossRef] [PubMed]
- Khvorova, A.; Watts, J.K. The chemical evolution of oligonucleotide therapies of clinical utility. Nat. Biotechnol. 2017, 35, 238–248. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Yang, Z.; Tang, X. Chemical modifications of nucleic acid drugs and their delivery systems for gene-based therapy. Med. Res. Rev. 2018, 38, 829–869. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Liu, X.Y.; Lu, A.; Wang, X.Y.; Jiang, L.X.; Wang, J.C. Non-viral vectors for RNA delivery. J. Control. Release 2022, 342, 241–279. [Google Scholar] [CrossRef]
- Yin, H.; Kanasty, R.L.; Eltoukhy, A.A.; Vegas, A.J.; Dorkin, J.R.; Anderson, D.G. Non-viral vectors for gene-based therapy. Nat. Rev. Genet. 2014, 15, 541–555. [Google Scholar] [CrossRef]
- Hou, X.; Zaks, T.; Langer, R.; Dong, Y. Lipid nanoparticles for mRNA delivery. Nat. Rev. Mater. 2021, 6, 1078–1094. [Google Scholar] [CrossRef] [PubMed]
- Binnewies, M.; Roberts, E.W.; Kersten, K.; Chan, V.; Fearon, D.F.; Merad, M.; Coussens, L.M.; Gabrilovich, D.I.; Ostrand-Rosenberg, S.; Hedrick, C.C.; et al. Understanding the tumor immune microenvironment (TIME) for effective therapy. Nat. Med. 2018, 24, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Tang, T.; Huang, X.; Zhang, G.; Hong, Z.; Bai, X.; Liang, T. Advantages of targeting the tumor immune microenvironment over blocking immune checkpoint in cancer immunotherapy. Signal Transduct. Target. Ther. 2021, 6, 72. [Google Scholar] [CrossRef]
- Yan, W.L.; Lang, T.Q.; Yuan, W.H.; Yin, Q.; Li, Y.P. Nanosized drug delivery systems modulate the immunosuppressive microenvironment to improve cancer immunotherapy. Acta Pharmacol. Sin. 2022, 43, 3045–3054. [Google Scholar] [CrossRef]
- Anagnostou, V.; Bardelli, A.; Chan, T.A.; Turajlic, S. The status of tumor mutational burden and immunotherapy. Nat. Cancer 2022, 3, 652–656. [Google Scholar] [CrossRef] [PubMed]
- Blankenstein, T.; Coulie, P.G.; Gilboa, E.; Jaffee, E.M. The determinants of tumour immunogenicity. Nat. Rev. Cancer 2012, 12, 307–313. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Xu, C. Immune checkpoint signaling and cancer immunotherapy. Cell Res. 2020, 30, 660–669. [Google Scholar] [CrossRef]
- Closset, L.; Gultekin, O.; Salehi, S.; Sarhan, D.; Lehti, K.; Gonzalez-Molina, J. The extracellular matrix-immune microenvironment crosstalk in cancer therapy: Challenges and opportunities. Matrix Biol. 2023, 121, 217–228. [Google Scholar] [CrossRef]
- Li, L.; Yu, R.; Cai, T.; Chen, Z.; Lan, M.; Zou, T.; Wang, B.; Wang, Q.; Zhao, Y.; Cai, Y. Effects of immune cells and cytokines on inflammation and immunosuppression in the tumor microenvironment. Int. Immunopharmacol. 2020, 88, 106939. [Google Scholar] [CrossRef]
- Tie, Y.; Tang, F.; Wei, Y.Q.; Wei, X.W. Immunosuppressive cells in cancer: Mechanisms and potential therapeutic targets. J. Hematol. Oncol. 2022, 15, 61. [Google Scholar] [CrossRef]
- Guo, C.; You, Z.; Shi, H.; Sun, Y.; Du, X.; Palacios, G.; Guy, C.; Yuan, S.; Chapman, N.M.; Lim, S.A.; et al. SLC38A2 and glutamine signalling in cDC1s dictate anti-tumour immunity. Nature 2023, 620, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Yu, T.J.; Xu, Y.; Ding, R.; Wang, Y.P.; Jiang, Y.Z.; Shao, Z.M. Emerging therapies in cancer metabolism. Cell Metab. 2023, 35, 1283–1303. [Google Scholar] [CrossRef] [PubMed]
- Kao, K.C.; Vilbois, S.; Tsai, C.H.; Ho, P.C. Metabolic communication in the tumour-immune microenvironment. Nat. Cell Biol. 2022, 24, 1574–1583. [Google Scholar] [CrossRef] [PubMed]
- Sheikhpour, M.; Barani, L.; Kasaeian, A. Biomimetics in drug delivery systems: A critical review. J. Control. Release 2017, 253, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Yoo, J.W.; Irvine, D.J.; Discher, D.E.; Mitragotri, S. Bio-inspired, bioengineered and biomimetic drug delivery carriers. Nat. Rev. Drug Discov. 2011, 10, 521–535. [Google Scholar] [CrossRef] [PubMed]
- Lang, T.; Yin, Q.; Li, Y. Progress of cell-derived biomimetic drug delivery systems for cancer therapy. Adv. Ther. 2018, 1, 1800053. [Google Scholar] [CrossRef]
- Mastrobattista, E.; van der Aa, M.A.; Hennink, W.E.; Crommelin, D.J. Artificial viruses: A nanotechnological approach to gene delivery. Nat. Rev. Drug Discov. 2006, 5, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Eweje, F.; Walsh, M.L.; Ahmad, K.; Ibrahim, V.; Alrefai, A.; Chen, J.; Chaikof, E.L. Protein-based nanoparticles for therapeutic nucleic acid delivery. Biomaterials 2024, 305, 122464. [Google Scholar] [CrossRef] [PubMed]
- Ni, R.; Zhou, J.; Hossain, N.; Chau, Y. Virus-inspired nucleic acid delivery system: Linking virus and viral mimicry. Adv. Drug Deliv. Rev. 2016, 106, 3–26. [Google Scholar] [CrossRef] [PubMed]
- Bulcha, J.T.; Wang, Y.; Ma, H.; Tai, P.W.L.; Gao, G. Viral vector platforms within the gene therapy landscape. Signal Transduct. Target. Ther. 2021, 6, 53. [Google Scholar] [CrossRef]
- Milone, M.C.; O’Doherty, U. Clinical use of lentiviral vectors. Leukemia 2018, 32, 1529–1541. [Google Scholar] [CrossRef] [PubMed]
- Palù, G.; Parolin, C.; Takeuchi, Y.; Pizzato, M. Progress with retroviral gene vectors. Rev. Med. Virol. 2000, 10, 185–202. [Google Scholar] [CrossRef]
- Fischer-Riepe, L.; Kailayangiri, S.; Zimmermann, K.; Pfeifer, R.; Aigner, M.; Altvater, B.; Kretschmann, S.; Völkl, S.; Hartley, J.; Dreger, C.; et al. Preclinical development of CAR T cells with antigen-inducible IL18 enforcement to treat GD2-positive solid cancers. Clin. Cancer Res. 2024, Of1–Of14. [Google Scholar] [CrossRef]
- Scarsella, L.; Ehrke-Schulz, E.; Paulussen, M.; Thal, S.C.; Ehrhardt, A.; Aydin, M. Advances of recombinant adenoviral vectors in preclinical and clinical applications. Viruses 2024, 16, 377. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Tai, P.W.L.; Gao, G. Adeno-associated virus vector as a platform for gene therapy delivery. Nat. Rev. Drug Discov. 2019, 18, 358–378. [Google Scholar] [CrossRef] [PubMed]
- Shalhout, S.Z.; Miller, D.M.; Emerick, K.S.; Kaufman, H.L. Therapy with oncolytic viruses: Progress and challenges. Nat. Rev. Clin. Oncol. 2023, 20, 160–177. [Google Scholar] [CrossRef] [PubMed]
- Greten, F.R.; Grivennikov, S.I. Inflammation and cancer: Triggers, mechanisms, and consequences. Immunity 2019, 51, 27–41. [Google Scholar] [CrossRef]
- Zhao, H.; Wu, L.; Yan, G.; Chen, Y.; Zhou, M.; Wu, Y.; Li, Y. Inflammation and tumor progression: Signaling pathways and targeted intervention. Signal Transduct. Target. Ther. 2021, 6, 263. [Google Scholar] [CrossRef] [PubMed]
- Borden, E.C. Interferons α and β in cancer: Therapeutic opportunities from new insights. Nat. Rev. Drug Discov. 2019, 18, 219–234. [Google Scholar] [CrossRef]
- Sterman, D.H.; Recio, A.; Carroll, R.G.; Gillespie, C.T.; Haas, A.; Vachani, A.; Kapoor, V.; Sun, J.; Hodinka, R.; Brown, J.L.; et al. A phase I clinical trial of single-dose intrapleural IFN-β gene transfer for malignant pleural mesothelioma and metastatic pleural effusions: High rate of antitumor immune responses. Clin. Cancer Res. 2007, 13, 4456–4466. [Google Scholar] [CrossRef]
- Sterman, D.H.; Haas, A.; Moon, E.; Recio, A.; Schwed, D.; Vachani, A.; Katz, S.I.; Gillespie, C.T.; Cheng, G.; Sun, J.; et al. A trial of intrapleural adenoviral-mediated Interferon-α2b gene transfer for malignant pleural mesothelioma. Am. J. Respir. Crit. Care Med. 2011, 184, 1395–1399. [Google Scholar] [CrossRef] [PubMed]
- Ranki, T.; Pesonen, S.; Hemminki, A.; Partanen, K.; Kairemo, K.; Alanko, T.; Lundin, J.; Linder, N.; Turkki, R.; Ristimäki, A.; et al. Phase I study with ONCOS-102 for the treatment of solid tumors—An evaluation of clinical response and exploratory analyses of immune markers. J. Immunother. Cancer 2016, 4, 17. [Google Scholar] [CrossRef] [PubMed]
- Ranki, T.; Joensuu, T.; Jäger, E.; Karbach, J.; Wahle, C.; Kairemo, K.; Alanko, T.; Partanen, K.; Turkki, R.; Linder, N.; et al. Local treatment of a pleural mesothelioma tumor with ONCOS-102 induces a systemic antitumor CD8+ T-cell response, prominent infiltration of CD8(+) lymphocytes and Th1 type polarization. Oncoimmunology 2014, 3, e958937. [Google Scholar] [CrossRef] [PubMed]
- Cerullo, V.; Pesonen, S.; Diaconu, I.; Escutenaire, S.; Arstila, P.T.; Ugolini, M.; Nokisalmi, P.; Raki, M.; Laasonen, L.; Särkioja, M.; et al. Oncolytic adenovirus coding for granulocyte macrophage colony-stimulating factor induces antitumoral immunity in cancer patients. Cancer Res. 2010, 70, 4297–4309. [Google Scholar] [CrossRef] [PubMed]
- Diaconu, I.; Cerullo, V.; Hirvinen, M.L.; Escutenaire, S.; Ugolini, M.; Pesonen, S.K.; Bramante, S.; Parviainen, S.; Kanerva, A.; Loskog, A.S.; et al. Immune response is an important aspect of the antitumor effect produced by a CD40L-encoding oncolytic adenovirus. Cancer Res. 2012, 72, 2327–2338. [Google Scholar] [CrossRef] [PubMed]
- Bortolanza, S.; Bunuales, M.; Otano, I.; Gonzalez-Aseguinolaza, G.; Ortiz-de-Solorzano, C.; Perez, D.; Prieto, J.; Hernandez-Alcoceba, R. Treatment of pancreatic cancer with an oncolytic adenovirus expressing interleukin-12 in Syrian hamsters. Mol. Ther. 2009, 17, 614–622. [Google Scholar] [CrossRef] [PubMed]
- Lapteva, N.; Aldrich, M.; Weksberg, D.; Rollins, L.; Goltsova, T.; Chen, S.Y.; Huang, X.F. Targeting the intratumoral dendritic cells by the oncolytic adenoviral vaccine expressing RANTES elicits potent antitumor immunity. J. Immunother. 2009, 32, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Ramlau, R.; Quoix, E.; Rolski, J.; Pless, M.; Lena, H.; Lévy, E.; Krzakowski, M.; Hess, D.; Tartour, E.; Chenard, M.P.; et al. A phase II study of Tg4010 (Mva-Muc1-Il2) in association with chemotherapy in patients with stage III/IV Non-small cell lung cancer. J. Thorac. Oncol. 2008, 3, 735–744. [Google Scholar] [CrossRef] [PubMed]
- Si, L.; Xu, H.; Zhou, X.; Zhang, Z.; Tian, Z.; Wang, Y.; Wu, Y.; Zhang, B.; Niu, Z.; Zhang, C.; et al. Generation of influenza A viruses as live but replication-incompetent virus vaccines. Science 2016, 354, 1170–1173. [Google Scholar] [CrossRef]
- Ji, D.; Zhang, Y.; Sun, J.; Zhang, B.; Ma, W.; Cheng, B.; Wang, X.; Li, Y.; Mu, Y.; Xu, H.; et al. An engineered influenza virus to deliver antigens for lung cancer vaccination. Nat. Biotechnol. 2024, 42, 518–528. [Google Scholar] [CrossRef]
- High, K.A.; Roncarolo, M.G. Gene therapy. N. Engl. J. Med. 2019, 381, 455–464. [Google Scholar] [CrossRef] [PubMed]
- Lawler, S.E.; Speranza, M.C.; Cho, C.F.; Chiocca, E.A. Oncolytic viruses in cancer treatment: A review. JAMA Oncol. 2017, 3, 841–849. [Google Scholar] [CrossRef] [PubMed]
- Ling, A.L.; Solomon, I.H.; Landivar, A.M.; Nakashima, H.; Woods, J.K.; Santos, A.; Masud, N.; Fell, G.; Mo, X.; Yilmaz, A.S.; et al. Clinical trial links oncolytic immunoactivation to survival in glioblastoma. Nature 2023, 623, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Ding, X.; Liao, Q.; Gao, N.; Chen, Y.; Zhao, C.; Zhang, X.; Xu, J. IL-21 arming potentiates the anti-tumor activity of an oncolytic vaccinia virus in monotherapy and combination therapy. J. Immunother. Cancer 2021, 9, e001647. [Google Scholar] [CrossRef] [PubMed]
- de Queiroz, N.; Xia, T.; Konno, H.; Barber, G.N. Ovarian cancer cells commonly exhibit defective STING signaling which affects sensitivity to viral oncolysis. Mol. Cancer Res. 2019, 17, 974–986. [Google Scholar] [CrossRef]
- Tatarūnas, V.; Čiapienė, I.; Giedraitienė, A. Precise therapy using the selective endogenous encapsidation for cellular delivery vector system. Pharmaceutics 2024, 16, 292. [Google Scholar] [CrossRef] [PubMed]
- Berger, S.; Lächelt, U.; Wagner, E. Dynamic carriers for therapeutic RNA delivery. Proc. Natl. Acad. Sci. USA 2024, 121, e2307799120. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Chen, X.; Tang, G.; Zhang, R.; Li, J.; Sun, G.; Yan, X.; Fan, K. Constructing a nanocage-based universal carrier for delivering TLR-activating nucleic acids to enhance antitumor immunotherapy. Nano Today 2022, 46, 101564. [Google Scholar] [CrossRef]
- Li, J.; Sun, Y.; Jia, T.; Zhang, R.; Zhang, K.; Wang, L. Messenger RNA vaccine based on recombinant MS2 virus-like particles against prostate cancer. Int. J. Cancer 2014, 134, 1683–1694. [Google Scholar] [CrossRef]
- Meng, C.; Chen, Z.; Mai, J.; Shi, Q.; Tian, S.; Hinkle, L.; Li, J.; Zhang, Z.; Ramirez, M.; Zhang, L.; et al. Virus-mimic mRNA vaccine for cancer treatment. Adv. Ther. 2021, 4, 2100144. [Google Scholar] [CrossRef]
- Mai, Y.; Guo, J.; Zhao, Y.; Ma, S.; Hou, Y.; Yang, J. Intranasal delivery of cationic liposome-protamine complex mRNA vaccine elicits effective anti-tumor immunity. Cell. Immunol. 2020, 354, 104143. [Google Scholar] [CrossRef] [PubMed]
- Segel, M.; Lash, B.; Song, J.; Ladha, A.; Liu, C.C.; Jin, X.; Mekhedov, S.L.; Macrae, R.K.; Koonin, E.V.; Zhang, F. Mammalian retrovirus-like protein PEG10 packages its own mRNA and can be pseudotyped for mRNA delivery. Science 2021, 373, 882–889. [Google Scholar] [CrossRef]
- Madigan, V.; Zhang, Y.; Raghavan, R.; Wilkinson, M.E.; Faure, G.; Puccio, E.; Segel, M.; Lash, B.; Macrae, R.K.; Zhang, F. Human paraneoplastic antigen Ma2 (PNMA2) forms icosahedral capsids that can be engineered for mRNA delivery. Proc. Natl. Acad. Sci. USA 2024, 121, e2307812120. [Google Scholar] [CrossRef] [PubMed]
- Gurbatri, C.R.; Arpaia, N.; Danino, T. Engineering bacteria as interactive cancer therapies. Science 2022, 378, 858–864. [Google Scholar] [CrossRef] [PubMed]
- Zhu, R.; Lang, T.; Yan, W.; Zhu, X.; Huang, X.; Yin, Q.; Li, Y. Gut microbiota: Influence on carcinogenesis and modulation strategies by drug delivery systems to improve cancer therapy. Adv. Sci. 2021, 8, 2003542. [Google Scholar] [CrossRef] [PubMed]
- Toyofuku, M.; Schild, S.; Kaparakis-Liaskos, M.; Eberl, L. Composition and functions of bacterial membrane vesicles. Nat. Rev. Microbiol. 2023, 21, 415–430. [Google Scholar] [CrossRef] [PubMed]
- Raman, V.; Van Dessel, N.; Hall, C.L.; Wetherby, V.E.; Whitney, S.A.; Kolewe, E.L.; Bloom, S.M.K.; Sharma, A.; Hardy, J.A.; Bollen, M.; et al. Intracellular delivery of protein drugs with an autonomously lysing bacterial system reduces tumor growth and metastases. Nat. Commun. 2021, 12, 6116. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Wang, Y.; Liu, J.; Rawding, P.; Bu, J.; Hong, S.; Hu, Q. Chemically and biologically engineered bacteria-based delivery systems for emerging diagnosis and advanced therapy. Adv. Mater. 2021, 33, 2102580. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Yu, J.; Liu, Y.; Xiong, B.; Fang, Y.; Zhu, Y.; Li, S.; Sun, L.; Zhou, B.; Sun, Y.; et al. Photosynthetic bacteria-hitchhiking 2D iMXene-mRNA vaccine to enable photo-immunogene cancer therapy. Adv. Sci. 2024, 11, e2307225. [Google Scholar] [CrossRef]
- Lynch, J.P.; Goers, L.; Lesser, C.F. Emerging strategies for engineering Escherichia coli Nissle 1917-based therapeutics. Trends Pharmacol. Sci. 2022, 43, 772–786. [Google Scholar] [CrossRef]
- Leventhal, D.S.; Sokolovska, A.; Li, N.; Plescia, C.; Kolodziej, S.A.; Gallant, C.W.; Christmas, R.; Gao, J.R.; James, M.J.; Abin-Fuentes, A.; et al. Immunotherapy with engineered bacteria by targeting the STING pathway for anti-tumor immunity. Nat. Commun. 2020, 11, 2739. [Google Scholar] [CrossRef] [PubMed]
- Luke, J.J.; Piha-Paul, S.A.; Medina, T.; Verschraegen, C.F.; Varterasian, M.; Brennan, A.M.; Riese, R.J.; Sokolovska, A.; Strauss, J.; Hava, D.L.; et al. Phase I Study of SYNB1891, an engineered E. coli Nissle strain expressing STING agonist, with and without atezolizumab in advanced malignancies. Clin. Cancer Res. 2023, 29, 2435–2444. [Google Scholar] [CrossRef] [PubMed]
- Chien, T.; Harimoto, T.; Kepecs, B.; Gray, K.; Coker, C.; Hou, N.; Pu, K.; Azad, T.; Nolasco, A.; Pavlicova, M.; et al. Enhancing the tropism of bacteria via genetically programmed biosensors. Nat. Biomed. Eng. 2022, 6, 94–104. [Google Scholar] [CrossRef] [PubMed]
- Din, M.O.; Danino, T.; Prindle, A.; Skalak, M.; Selimkhanov, J.; Allen, K.; Julio, E.; Atolia, E.; Tsimring, L.S.; Bhatia, S.N.; et al. Synchronized cycles of bacterial lysis for in vivo delivery. Nature 2016, 536, 81–85. [Google Scholar] [CrossRef] [PubMed]
- Logtenberg, M.E.W.; Scheeren, F.A.; Schumacher, T.N. The CD47-SIRPα Immune Checkpoint. Immunity 2020, 52, 742–752. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, S.; Castro, S.; Coker, C.; Hinchliffe, T.E.; Arpaia, N.; Danino, T. Programmable bacteria induce durable tumor regression and systemic antitumor immunity. Nat. Med. 2019, 25, 1057–1063. [Google Scholar] [CrossRef] [PubMed]
- Gurbatri, C.R.; Lia, I.; Vincent, R.; Coker, C.; Castro, S.; Treuting, P.M.; Hinchliffe, T.E.; Arpaia, N.; Danino, T. Engineered probiotics for local tumor delivery of checkpoint blockade nanobodies. Sci. Transl. Med. 2020, 12, eaax0876. [Google Scholar] [CrossRef] [PubMed]
- Schwechheimer, C.; Kuehn, M.J. Outer-membrane vesicles from Gram-negative bacteria: Biogenesis and functions. Nat. Rev. Microbiol. 2015, 13, 605–619. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Ma, X.; Yue, Y.; Zhang, K.; Cheng, K.; Feng, Q.; Ma, N.; Liang, J.; Zhang, T.; Zhang, L.; et al. Rapid surface display of mRNA antigens by bacteria-derived outer membrane vesicles for a personalized tumor vaccine. Adv. Mater. 2022, 34, e2109984. [Google Scholar] [CrossRef]
- Qing, S.; Lyu, C.; Zhu, L.; Pan, C.; Wang, S.; Li, F.; Wang, J.; Yue, H.; Gao, X.; Jia, R.; et al. Biomineralized bacterial outer membrane vesicles potentiate safe and efficient tumor microenvironment reprogramming for anticancer therapy. Adv. Mater. 2020, 32, e2002085. [Google Scholar] [CrossRef]
- Li, S.; Li, X.; Meng, J.; Bao, W.; Wang, S.; Ye, P.; Gao, X.-D.; Wei, W. Dynamic shielding of bacterial outer membrane vesicles for safe and efficient chemo-immunotherapy against tumors. Nano Res. 2023, 17, 836–847. [Google Scholar] [CrossRef]
- Cassetta, L.; Pollard, J.W. A timeline of tumour-associated macrophage biology. Nat. Rev. Cancer 2023, 23, 238–257. [Google Scholar] [CrossRef]
- Zhao, M.; Cheng, X.; Shao, P.; Dong, Y.; Wu, Y.; Xiao, L.; Cui, Z.; Sun, X.; Gao, C.; Chen, J.; et al. Bacterial protoplast-derived nanovesicles carrying CRISPR-Cas9 tools re-educate tumor-associated macrophages for enhanced cancer immunotherapy. Nat. Commun. 2024, 15, 950. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Shi, X.; Kuang, Y.; Wei, R.; Feng, L.; Chen, J.; Wu, X. Cell-nanocarrier drug delivery system: A promising strategy for cancer therapy. Drug Deliv. Transl. Res. 2024, 14, 581–596. [Google Scholar] [CrossRef]
- Yu, H.; Yang, Z.; Li, F.; Xu, L.; Sun, Y. Cell-mediated targeting drugs delivery systems. Drug Deliv. 2020, 27, 1425–1437. [Google Scholar] [CrossRef]
- Yang, L.; Yang, Y.; Chen, Y.; Xu, Y.; Peng, J. Cell-based drug delivery systems and their in vivo fate. Adv. Drug Deliv. Rev. 2022, 187, 114394. [Google Scholar] [CrossRef]
- Meng, J.; Lv, Y.; Bao, W.; Meng, Z.; Wang, S.; Wu, Y.; Li, S.; Jiao, Z.; Tian, Z.; Ma, G.; et al. Generation of whole tumor cell vaccine for on-demand manipulation of immune responses against cancer under near-infrared laser irradiation. Nat. Commun. 2023, 14, 4505. [Google Scholar] [CrossRef] [PubMed]
- Tanyi, J.L.; Bobisse, S.; Ophir, E.; Tuyaerts, S.; Roberti, A.; Genolet, R.; Baumgartner, P.; Stevenson, B.J.; Iseli, C.; Dangaj, D.; et al. Personalized cancer vaccine effectively mobilizes antitumor T cell immunity in ovarian cancer. Sci. Transl. Med. 2018, 10, eaao5931. [Google Scholar] [CrossRef] [PubMed]
- Padmanaban, V.; Krol, I.; Suhail, Y.; Szczerba, B.M.; Aceto, N.; Bader, J.S.; Ewald, A.J. E-cadherin is required for metastasis in multiple models of breast cancer. Nature 2019, 573, 439–444. [Google Scholar] [CrossRef]
- Ci, T.; Li, H.; Chen, G.; Wang, Z.; Wang, J.; Abdou, P.; Tu, Y.; Dotti, G.; Gu, Z. Cryo-shocked cancer cells for targeted drug delivery and vaccination. Sci. Adv. 2020, 6, eabc3013. [Google Scholar] [CrossRef]
- Zhao, Z.; Fang, L.; Xiao, P.; Sun, X.; Zhou, L.; Liu, X.; Wang, J.; Wang, G.; Cao, H.; Zhang, P.; et al. Walking dead tumor cells for targeted drug delivery against lung metastasis of triple-negative breast cancer. Adv. Mater. 2022, 34, 2205462. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Xin, M.; Feng, H.; Zhang, W.; Liao, Z.; Sheng, T.; Wen, P.; Wu, Q.; Liang, T.; Shi, J.; et al. Cryo-shocked tumor cells deliver CRISPR-Cas9 for lung cancer regression by synthetic lethality. Sci. Adv. 2024, 10, eadk8264. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; De May, H.; Franco, S.; Noureddine, A.; Tang, L.; Brinker, C.J.; Kusewitt, D.F.; Adams, S.F.; Serda, R.E. Cancer vaccines from cryogenically silicified tumour cells functionalized with pathogen-associated molecular patterns. Nat. Biomed. Eng. 2022, 6, 19–31. [Google Scholar] [CrossRef]
- Jiang, Q.; Wei, H.; Tian, Z. Poly I:C enhances cycloheximide-induced apoptosis of tumor cells through TLR3 pathway. BMC Cancer 2008, 8, 12. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; He, H.; Liang, R.; Pan, H.; Chen, Z.; Deng, G.; Zhang, S.; Ma, Y.; Liu, L.; Cai, L. In situ poly I:C released from living cell drug nanocarriers for macrophage-mediated antitumor immunotherapy. Biomaterials 2021, 269, 120670. [Google Scholar] [CrossRef] [PubMed]
- Fang, R.H.; Gao, W.; Zhang, L. Targeting drugs to tumours using cell membrane-coated nanoparticles. Nat. Rev. Clin. Oncol. 2023, 20, 33–48. [Google Scholar] [CrossRef] [PubMed]
- Xia, Q.; Zhang, Y.; Li, Z.; Hou, X.; Feng, N. Red blood cell membrane-camouflaged nanoparticles: A novel drug delivery system for antitumor application. Acta Pharm. Sin. B 2019, 9, 675–689. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Qin, D.; Zou, J.; Li, X.; Guo, X.D.; Tang, Y.; Liu, C.; Chen, W.; Kong, N.; Zhang, C.Y.; et al. Living leukocyte-based drug delivery systems. Adv. Mater. 2023, 35, 2207787. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Xu, L.; Kang, Y.; Cheng, Q.; He, Y.; Ji, X. Platelet-derived drug delivery systems: Pioneering treatment for cancer, cardiovascular diseases, infectious diseases, and beyond. Biomaterials 2024, 306, 122478. [Google Scholar] [CrossRef]
- Ma, Y.; Yi, J.; Ruan, J.; Ma, J.; Yang, Q.; Zhang, K.; Zhang, M.; Zeng, G.; Jin, L.; Huang, X.; et al. Engineered cell membrane-coated nanoparticles: New strategies in glioma targeted therapy and immune modulation. Adv. Healthc. Mater. 2024, 2400514. [Google Scholar] [CrossRef]
- Peng, X.; Yang, L.; Yuan, P.; Ding, X. Hybrid cell membrane-based nanoplatforms for enhanced immunotherapy against cancer and infectious diseases. Adv. Healthc. Mater. 2024, 2304477. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, N.; Jiang, Y.; Zhou, J.; Mohapatra, A.; Peng, F.X.; Duan, Y.; Holay, M.; Chekuri, S.; Guo, Z.; Gao, W.; et al. A modular approach to enhancing cell membrane-coated nanoparticle functionality using genetic engineering. Nat. Nanotechnol. 2024, 19, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Chen, M.; He, J. Cancer cell membrane cloaking nanoparticles for targeted co-delivery of doxorubicin and PD-L1 siRNA. Artif. Cells Nanomed. Biotechnol. 2019, 47, 1635–1641. [Google Scholar] [CrossRef] [PubMed]
- Mu, X.; Zhang, M.; Wei, A.; Yin, F.; Wang, Y.; Hu, K.; Jiang, J. Doxorubicin and PD-L1 siRNA co-delivery with stem cell membrane-coated polydopamine nanoparticles for the targeted chemoimmunotherapy of PCa bone metastases. Nanoscale 2021, 13, 8998–9008. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Chen, S.; Zhang, M.; Lin, X.; Jin, X.; Zhang, M.; Liu, Y.; Wang, Y.; Shi, K. A trojan horse delivery vehicle carrying siRNA nanotherapeutics with multiple tumor microenvironment responsiveness elicits robust antitumor immune responses in situ via a "self-synergistic" approach. Adv. Healthc. Mater. 2023, 12, 2301401. [Google Scholar] [CrossRef] [PubMed]
- Gong, C.; Yu, X.; Zhang, W.; Han, L.; Wang, R.; Wang, Y.; Gao, S.; Yuan, Y. Regulating the immunosuppressive tumor microenvironment to enhance breast cancer immunotherapy using pH-responsive hybrid membrane-coated nanoparticles. J. Nanobiotechnol. 2021, 19, 58. [Google Scholar] [CrossRef] [PubMed]
- Lv, H.; Wang, T.; Ma, F.; Zhang, K.; Gao, T.; Pei, R.; Zhang, Y. Aptamer-functionalized targeted siRNA delivery system for tumor immunotherapy. Biomed. Mater. 2022, 17, 024108. [Google Scholar] [CrossRef] [PubMed]
- Kroll, A.V.; Fang, R.H.; Jiang, Y.; Zhou, J.; Wei, X.; Yu, C.L.; Gao, J.; Luk, B.T.; Dehaini, D.; Gao, W.; et al. Nanoparticulate delivery of cancer cell membrane elicits multiantigenic antitumor immunity. Adv. Mater. 2017, 29, 1703969. [Google Scholar] [CrossRef]
- Park, J.H.; Mohapatra, A.; Zhou, J.; Holay, M.; Krishnan, N.; Gao, W.; Fang, R.H.; Zhang, L. Virus-mimicking cell membrane-coated nanoparticles for cytosolic delivery of mRNA. Angew. Chem. 2022, 61, e202113671. [Google Scholar] [CrossRef]
- Li, W.; Zhang, X.; Zhang, C.; Yan, J.; Hou, X.; Du, S.; Zeng, C.; Zhao, W.; Deng, B.; McComb, D.W.; et al. Biomimetic nanoparticles deliver mRNAs encoding costimulatory receptors and enhance T cell mediated cancer immunotherapy. Nat. Commun. 2021, 12, 7264. [Google Scholar] [CrossRef]
- El Andaloussi, S.; Mäger, I.; Breakefield, X.O.; Wood, M.J. Extracellular vesicles: Biology and emerging therapeutic opportunities. Nat. Rev. Drug Discov. 2013, 12, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Vader, P.; Mol, E.A.; Pasterkamp, G.; Schiffelers, R.M. Extracellular vesicles for drug delivery. Adv. Drug Deliv. Rev. 2016, 106, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Liu, Z.; Wu, J.; Li, B. Cell-derived vesicles for mRNA delivery. Pharmaceutics 2022, 14, 2699. [Google Scholar] [CrossRef] [PubMed]
- Roerig, J.; Mitrach, F.; Schmid, M.; Hause, G.; Hacker, M.C.; Wölk, C.; Schulz-Siegmund, M. Synergistic siRNA loading of extracellular vesicles enables functional delivery into cells. Small Methods 2022, 6, 2201001. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, K.; Breyne, K.; Ughetto, S.; Laurent, L.C.; Breakefield, X.O. RNA delivery by extracellular vesicles in mammalian cells and its applications. Nat. Rev. Mol. Cell Biol. 2020, 21, 585–606. [Google Scholar] [CrossRef]
- Tian, T.; Liang, R.; Erel-Akbaba, G.; Saad, L.; Obeid, P.J.; Gao, J.; Chiocca, E.A.; Weissleder, R.; Tannous, B.A. Immune checkpoint inhibition in GBM primed with radiation by engineered extracellular vesicles. ACS Nano 2022, 16, 1940–1953. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wu, Q.; Gong, Y.; Qin, Q.; Han, Q.; Cheng, Z.; Yan, Z. Biomimetic exosomal vesicles loaded with siRNA improves antitumor immune responses by inhibiting the secretion of tumor-derived exosome PD-L1. Int. Immunopharmacol. 2024, 129, 111659. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Huang, L.; Mao, M.; Ding, J.; Wu, G.; Fan, W.; Yang, T.; Zhang, M.; Huang, Y.; Xie, H.-Y. Viral protein-pseudotyped and siRNA-electroporated extracellular vesicles for cancer immunotherapy. Adv. Funct. Mater. 2020, 30, 2006515. [Google Scholar] [CrossRef]
- Morishita, M.; Takahashi, Y.; Matsumoto, A.; Nishikawa, M.; Takakura, Y. Exosome-based tumor antigens-adjuvant co-delivery utilizing genetically engineered tumor cell-derived exosomes with immunostimulatory CpG DNA. Biomaterials 2016, 111, 55–65. [Google Scholar] [CrossRef]
- Yu, T.; Wang, X.; Zhi, T.; Zhang, J.; Wang, Y.; Nie, E.; Zhou, F.; You, Y.; Liu, N. Delivery of MGMT mRNA to glioma cells by reactive astrocyte-derived exosomes confers a temozolomide resistance phenotype. Cancer Lett. 2018, 433, 210–220. [Google Scholar] [CrossRef]
- Wang, J.H.; Forterre, A.V.; Zhao, J.; Frimannsson, D.O.; Delcayre, A.; Antes, T.J.; Efron, B.; Jeffrey, S.S.; Pegram, M.D.; Matin, A.C. Anti-Her2 scFv-directed extracellular vesicle-mediated mRNA-based gene delivery inhibits growth of Her2-positive human breast tumor xenografts by prodrug activation. Mol. Cancer Ther. 2018, 17, 1133–1142. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Liu, X.; Bi, Y.; Wang, Y.; Antony, A.; Lee, D.; Huntoon, K.; Jeong, S.; Ma, Y.; Li, X.; et al. Adaptive design of mRNA-loaded extracellular vesicles for targeted immunotherapy of cancer. Nat. Commun. 2023, 14, 6610. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Shi, J.; Xie, J.; Wang, Y.; Sun, J.; Liu, T.; Zhao, Y.; Zhao, X.; Wang, X.; Ma, Y.; et al. Large-scale generation of functional mRNA-encapsulating exosomes via cellular nanoporation. Nat. Biomed. Eng. 2020, 4, 69–83. [Google Scholar] [CrossRef] [PubMed]
- Gu, W.; Luozhong, S.; Cai, S.; Londhe, K.; Elkasri, N.; Hawkins, R.; Yuan, Z.; Su-Greene, K.; Yin, Y.; Cruz, M.; et al. Extracellular vesicles incorporating retrovirus-like capsids for the enhanced packaging and systemic delivery of mRNA into neurons. Nat. Biomed. Eng. 2024, 8, 415–426. [Google Scholar] [CrossRef]
- Yin, D.; Zhong, Y.; Ling, S.; Lu, S.; Wang, X.; Jiang, Z.; Wang, J.; Dai, Y.; Tian, X.; Huang, Q.; et al. Dendritic-cell-targeting virus-like particles as potent mRNA vaccine carriers. Nat. Biomed. Eng. 2024. [CrossRef] [PubMed]
- Stranford, D.M.; Simons, L.M.; Berman, K.E.; Cheng, L.; DiBiase, B.N.; Hung, M.E.; Lucks, J.B.; Hultquist, J.F.; Leonard, J.N. Genetically encoding multiple functionalities into extracellular vesicles for the targeted delivery of biologics to T cells. Nat. Biomed. Eng. 2024, 8, 397–414. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, J.R.; Chen, E.; Perez, B.S.; Sandoval Espinoza, C.R.; Kang, M.H.; Trinidad, M.; Ngo, W.; Doudna, J.A. In vivo human T cell engineering with enveloped delivery vehicles. Nat. Biotechnol. 2024. [CrossRef] [PubMed]
- Liu, Z.; Shi, M.; Ren, Y.; Xu, H.; Weng, S.; Ning, W.; Ge, X.; Liu, L.; Guo, C.; Duo, M.; et al. Recent advances and applications of CRISPR-Cas9 in cancer immunotherapy. Mol. Cancer 2023, 22, 35. [Google Scholar] [CrossRef]
- Harrington, K.; Freeman, D.J.; Kelly, B.; Harper, J.; Soria, J.C. Optimizing oncolytic virotherapy in cancer treatment. Nat. Rev. Drug Discov. 2019, 18, 689–706. [Google Scholar] [CrossRef]
- Barati, M.; Mirzavi, F.; Atabaki, M.; Bibak, B.; Mohammadi, M.; Jaafari, M.R. A review of PD-1/PD-L1 siRNA delivery systems in immune T cells and cancer cells. Int. Immunopharmacol. 2022, 111, 109022. [Google Scholar] [CrossRef]
- Kuai, R.; Ochyl, L.J.; Bahjat, K.S.; Schwendeman, A.; Moon, J.J. Designer vaccine nanodiscs for personalized cancer immunotherapy. Nat. Mater. 2017, 16, 489–496. [Google Scholar] [CrossRef]
- Manzari, M.T.; Shamay, Y.; Kiguchi, H.; Rosen, N.; Scaltriti, M.; Heller, D.A. Targeted drug delivery strategies for precision medicines. Nat. Rev. Mater. 2021, 6, 351–370. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Li, S.; Lin, X.; Chen, Y. A perspective of lipid nanoparticles for RNA delivery. Exploration 2024, 20230147. [Google Scholar] [CrossRef]
- Akinc, A.; Maier, M.A.; Manoharan, M.; Fitzgerald, K.; Jayaraman, M.; Barros, S.; Ansell, S.; Du, X.; Hope, M.J.; Madden, T.D.; et al. The Onpattro story and the clinical translation of nanomedicines containing nucleic acid-based drugs. Nat. Nanotechnol. 2019, 14, 1084–1087. [Google Scholar] [CrossRef] [PubMed]
- Tenchov, R.; Bird, R.; Curtze, A.E.; Zhou, Q. Lipid Nanoparticles—From Liposomes to mRNA Vaccine Delivery, a Landscape of Research Diversity and Advancement. ACS Nano 2021, 15, 16982–17015. [Google Scholar] [CrossRef]
- Kon, E.; Ad-El, N.; Hazan-Halevy, I.; Stotsky-Oterin, L.; Peer, D. Targeting cancer with mRNA-lipid nanoparticles: Key considerations and future prospects. Nat. Rev. Clin. Oncol. 2023, 20, 739–754. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Zhao, Y.; Hu, X.; Jiao, J.; Wang, W.; Yao, H. Advances in the clinical development of oncolytic viruses. Am. J. Transl. Res. 2022, 14, 4192–4206. [Google Scholar] [PubMed]
- Huang, H.; Liu, Y.; Liao, W.; Cao, Y.; Liu, Q.; Guo, Y.; Lu, Y.; Xie, Z. Oncolytic adenovirus programmed by synthetic gene circuit for cancer immunotherapy. Nat. Commun. 2019, 10, 4801. [Google Scholar] [CrossRef] [PubMed]
- Peng, S.; Huang, H.; Wei, P.; Xie, Z. Synthetic gene circuits moving into the clinic. Quant. Biol. 2021, 9, 100–103. [Google Scholar] [CrossRef]
- Kwon, S.-Y.; Thi-Thu Ngo, H.; Son, J.; Hong, Y.; Min, J.-J. Exploiting bacteria for cancer immunotherapy. Nat. Rev. Clin. Oncol. 2024, 21, 569–589. [Google Scholar] [CrossRef] [PubMed]
- Soiffer, R.J.; Kooshesh, K.A.; Ho, V. Whole tumor cell vaccines engineered to secrete GM-CSF (GVAX). ImmunoMedicine 2021, 1, e1025. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nucleic Acid Drug | Delivery Platform | Strategy of Constructing the Biomimetic Nucleic Acid Drug Delivery System | Mechanism of Activating Antitumor Immune Response | Antitumor Effects | Ref. |
|---|---|---|---|---|---|
| CpG | Cryo-silicified tumor cells | Polyethyleneimine (PEI)-adsorbed cryo-silicified tumor cells were coated with CpG. | CpG activated Toll like receptor (TLR) 9 and silicified tumor cells served as the vaccine. | The vaccine eradicated the tumors in C57BL/6 mice bearing ovarian cancer. | [113] |
| Poly I:C | Macrophages | Poly(lactic-co-glycolic acid) (PLGA) nanoparticles encapsulating poly I:C were coupled onto the macrophage surface. | Poly I:C triggered apoptosis of tumor cells, promoted dendritic cells (DCs) maturation, and polarized macrophage vectors to the M1 type. | The biomimetic system induced 84.9% tumor cell apoptosis in vivo and inhibited lung metastases by 74.9% in 4T1 tumor-bearing mice. | [115] |
| Programmed death ligand 1 (PD-L1) small interfering RNA (siRNA) | Tumor cell membrane | Tumor cell membrane was utilized to coat PLGA nanoparticles containing PD-L1 siRNA. | Cell membrane-coated nanoparticles were more easily taken up by tumor cells, resulting in a more effective PD-L1 knockdown. | The nanoparticle induced cytotoxicity in the source cells of the cell membrane coating. | [123] |
| PD-L1 siRNA | Mesenchymal stem cell membrane | Mesenchymal stem cell membrane was utilized for coating PD-L1 siRNA and doxorubicin co-loaded polydopamine nanoparticle. | Doxorubicin exerted antitumor cytotoxicity and upregulated the expression of PD-L1, which could be inhibited by PD-L1 siRNA. | The biomimetic nanoparticle inhibited PC-3 cell growth both in vitro and in vivo. | [124] |
| PD-L1 siRNA | Macrophage cell membrane | Spermine-based nanoparticle compressing PD-L1 siRNA and a photosensitizer ICG was encapsulated with a macrophage cell membrane. | PD-L1 siRNA degraded PD-L1 mRNA, while IGG mediated photodynamic therapy for generating the in situ tumor vaccine. | The combination therapy achieved the apoptotic rate of 46.1% in vitro and the tumor growth inhibition rate of ~80% in vivo. | [125] |
| siRNA targeting fibrinogen-like protein 1 | Tumor cell–macrophage hybrid membrane | PLGA nanoparticles encapsulating siRNA and metformin were coated with the hybrid membrane. | Metformin and siRNA synergistically promoted T-cell-mediated immune responses. | The nanoparticle achieved an apoptosis rate of 75.71% in 4T1 cells in vitro and a tumor inhibitory rate of 97.3% in vivo. | [126] |
| CpG | Melanoma cell membrane | Melanoma cell membrane was loaded with a CpG-containing PLGA nanoparticle. | CpG induced maturation of DCs and the tumor cell membrane provided tumor antigens. | The vaccine prevented tumor occurrence in 86% of the mice and inhibited tumor growth. | [128] |
| PD-L1 siRNA | Neuronal cell-derived extracellular vesicles | Cerebrovascular endothelial cell-targeting peptides were coupled onto neuronal cell-derived extracellular vesicles to form delivery vehicles for PD-L1 siRNA. | Radiotherapy upregulated the intratumoral PD-L1 level, providing targets for siRNA. | The combination therapy inhibited glioblastoma in vivo and extended the median survival time from 22.5 to 47 d. | [136] |
| siRNA targeting neutral sphingomyelinase type 2 | ApoA1-modified tumor-derived exosome | ApoA1-modified tumor-derived exosome was coupled with cholesterol-decorated PD-L1 siRNA. | The downregulation of neutral sphingomyelinase type 2 reduced the level of PD-L1. | The exosome achieved the PD-L1 silencing efficiency of 94.07% at the protein level and significantly delayed HepG2 tumor growth. | [137] |
| PD-L1 siRNA | M1-type macrophage extracellular vesicles | PD-L1 siRNA was loaded by electroporation into vesicles expressing a virus-derived fusogenic protein with pH sensitivity. | PD-L1 siRNA downregulated the expression of PD-L1 in tumor cells, and the proinflammatory cytokines in the M1 macrophage-derived vesicles reprogramed M2-type tumor associated macrophages (TAMs). | The combination immunotherapy achieved the tumor inhibition rate of over 80% in vivo. | [138] |
| Interferon (IFN)-γ mRNA | Extracellular vesicles expressing CD64 | Antibodies-decorated vesicles loaded with IFN-γ mRNA were generated with nanosecond pulse electroporation. | The secreted IFN-γ upregulated tumor major histocompatibility complex (MHC) class I expression, enabling the immune system to recognize tumor cells. | The vesicle prolonged the median survival time from 29 to 53 d in GL261-bearing mice. | [142] |
| sgRNA and Cas protein | Extracellular vesicles expressing cell-targeting antibodies | Extracellular vesicles containing CRISPR/Cas9 systems were decorated with antibodies for T cell targeting and a mutant form of the vesicular stomatitis virus glycoprotein for cell fusion. | The engineered vesicles generated gene-edited chimeric antigen receptor (CAR)-T cells for killing tumor cells. | The in vivo-generated CAR-T cells depleted CD19 B cells. | [147] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, W.; Cao, Y.; Yin, Q.; Li, Y. Biomimetic Nucleic Acid Drug Delivery Systems for Relieving Tumor Immunosuppressive Microenvironment. Pharmaceutics 2024, 16, 1028. https://doi.org/10.3390/pharmaceutics16081028
Yan W, Cao Y, Yin Q, Li Y. Biomimetic Nucleic Acid Drug Delivery Systems for Relieving Tumor Immunosuppressive Microenvironment. Pharmaceutics. 2024; 16(8):1028. https://doi.org/10.3390/pharmaceutics16081028
Chicago/Turabian StyleYan, Wenlu, Ying Cao, Qi Yin, and Yaping Li. 2024. "Biomimetic Nucleic Acid Drug Delivery Systems for Relieving Tumor Immunosuppressive Microenvironment" Pharmaceutics 16, no. 8: 1028. https://doi.org/10.3390/pharmaceutics16081028
APA StyleYan, W., Cao, Y., Yin, Q., & Li, Y. (2024). Biomimetic Nucleic Acid Drug Delivery Systems for Relieving Tumor Immunosuppressive Microenvironment. Pharmaceutics, 16(8), 1028. https://doi.org/10.3390/pharmaceutics16081028