
Optimization of a Modular Nanotransporter Design for Targeted Intracellular Delivery of Photosensitizer
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. Producing MNTs with a Truncated Carrier Module
2.3. MNT Expression and Purification
2.4. Binding Assay
2.5. Liposome Leakage Assay
2.6. Thermophoresis
2.7. Photocytotoxicity
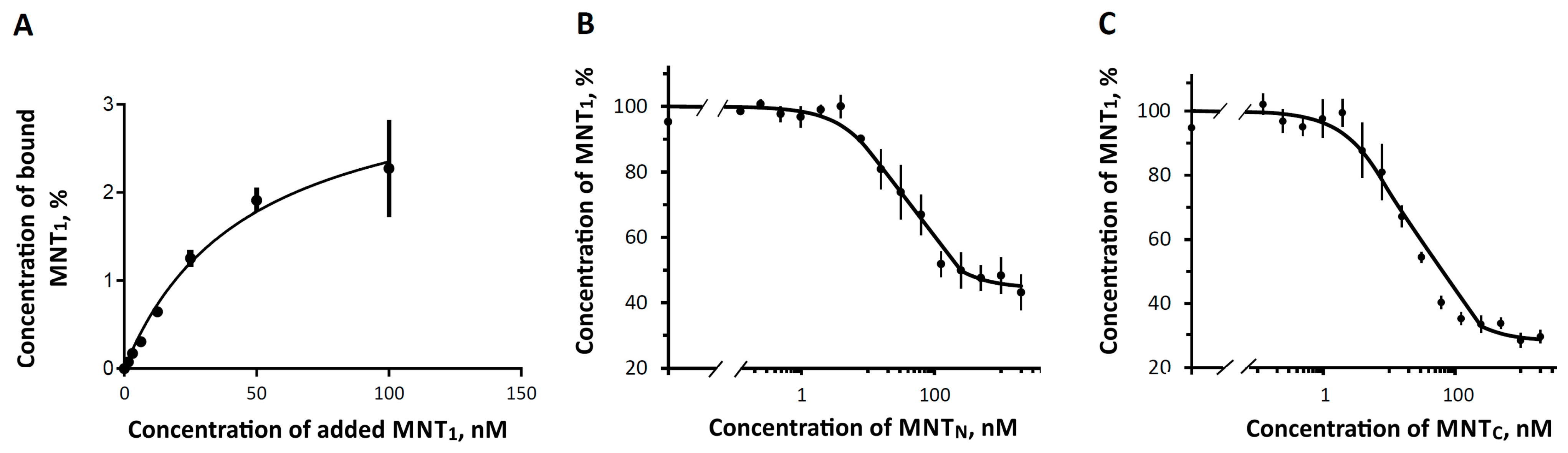
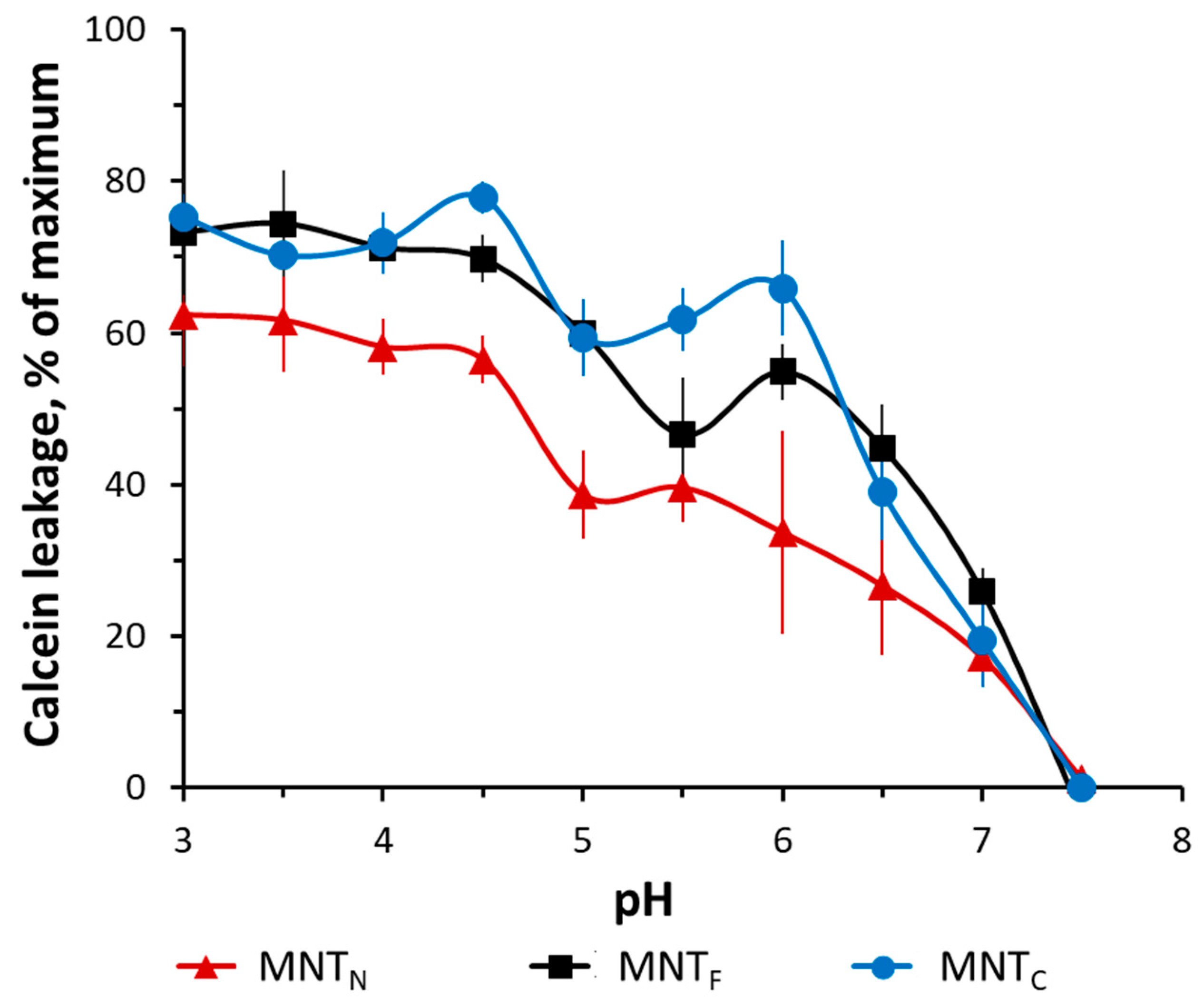
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, Y.T.; Tan, Y.J.; Oon, C.E. Molecular Targeted Therapy: Treating cancer with specificity. Eur. J. Pharmacol. 2018, 834, 188–196. [Google Scholar] [CrossRef]
- Manzari, M.T.; Shamay, Y.; Kiguchi, H.; Rosen, N.; Scaltriti, M.; Heller, D.A. Targeted drug delivery strategies for precision medicines. Nat. Rev. Mater. 2021, 6, 351–370. [Google Scholar] [CrossRef]
- Craik, D.J.; Fairlie, D.P.; Liras, S.; Price, D. The future of peptide-based drugs. Chem. Biol. Drug Des. 2013, 81, 136–147. [Google Scholar] [CrossRef]
- Muttenthaler, M.; King, G.F.; Adams, D.J.; Alewood, P.F. Trends in peptide drug discovery. Nat. Rev. Drug Discov. 2021, 20, 309–325. [Google Scholar] [CrossRef] [PubMed]
- Vargason, A.M.; Anselmo, A.C.; Mitragotri, S. The evolution of commercial drug delivery technologies. Nat. Biomed. Eng. 2021, 5, 951–967. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, N.; Zhang, W.; Cheng, X.; Yan, Z.; Shao, G.; Wang, X.; Wang, R.; Fu, C. Therapeutic peptides: Current applications and future directions. Signal Transduct. Target. Ther. 2022, 7, 48. [Google Scholar] [CrossRef]
- Sobolev, A.S. The Delivery of Biologically Active Agents into the Nuclei of Target Cells for the Purposes of Translational Medicine. Acta Naturae 2020, 12, 47–56. [Google Scholar] [CrossRef]
- Slastnikova, T.A.; Rosenkranz, A.A.; Gulak, P.V.; Schiffelers, R.M.; Lupanova, T.N.; Khramtsov, Y.V.; Zalutsky, M.R.; Sobolev, A.S. Modular nanotransporters: A multipurpose in vivo working platform for targeted drug delivery. Int. J. Nanomed. 2012, 7, 467–482. [Google Scholar]
- Slastnikova, T.A.; Rosenkranz, A.A.; Lupanova, T.N.; Gulak, P.V.; Gnuchev, N.V.; Sobolev, A.S. Study of efficiency of the modular nanotransporter for targeted delivery of photosensitizers to melanoma cell nuclei in vivo. Dokl. Biochem. Biophys. 2012, 446, 235–237. [Google Scholar] [CrossRef]
- Rosenkranz, A.A.; Slastnikova, T.A.; Karmakova, T.A.; Vorontsova, M.S.; Morozova, N.B.; Petriev, V.M.; Abrosimov, A.S.; Khramtsov, Y.V.; Lupanova, T.N.; Ulasov, A.V.; et al. Antitumor Activity of Auger Electron Emitter 111In Delivered by Modular Nanotransporter for Treatment of Bladder Cancer with EGFR Overexpression. Front. Pharmacol. 2018, 9, 1331. [Google Scholar] [CrossRef]
- Slastnikova, T.A.; Koumarianou, E.; Rosenkranz, A.A.; Vaidyanathan, G.; Lupanova, T.N.; Sobolev, A.S.; Zalutsky, M.R. Modular nanotransporters: A versatile approach for enhancing nuclear delivery and cytotoxicity of Auger electron-emitting 125I. EJNMMI Res. 2012, 2, 59. [Google Scholar] [CrossRef]
- Correia, J.H.; Rodrigues, J.A.; Pimenta, S.; Dong, T.; Yang, Z. Photodynamic Therapy Review: Principles, Photosensitizers, Applications, and Future Directions. Pharmaceutics 2021, 13, 1332. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Lovell, J.F.; Yoon, J.; Chen, X. Clinical development and potential of photothermal and photodynamic therapies for cancer. Nat. Rev. Clin. Oncol. 2020, 17, 657–674. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.E.; Chang, J.E. Recent Studies in Photodynamic Therapy for Cancer Treatment: From Basic Research to Clinical Trials. Pharmaceutics 2023, 15, 2257. [Google Scholar] [CrossRef]
- Agostinis, P.; Berg, K.; Cengel, K.A.; Foster, T.H.; Girotti, A.W.; Gollnick, S.O.; Hahn, S.M.; Hamblin, M.R.; Juzeniene, A.; Kessel, D.; et al. Photodynamic therapy of cancer: An update. CA Cancer J. Clin. 2011, 61, 250–281. [Google Scholar] [CrossRef] [PubMed]
- Huis In ‘t Veld, R.V.; Heuts, J.; Ma, S.; Cruz, L.J.; Ossendorp, F.A.; Jager, M.J. Current Challenges and Opportunities of Photodynamic Therapy against Cancer. Pharmaceutics 2023, 15, 330. [Google Scholar] [CrossRef]
- Egorov, S.Y.; Kamalov, V.F.; Koroteev, N.I.; Krasnovsky, A.A., Jr.; Toleutaev, B.N.; Zinukov, S.V. Rise and decay kinetics of photosensitized singlet oxygen luminescence in water. Measurements with nanosecond time-correlated single photon counting technique. Chem. Phys. Lett. 1989, 163, 421–424. [Google Scholar] [CrossRef]
- Redmond, R.W.; Kochevar, I.E. Spatially resolved cellular responses to singlet oxygen. Photochem. Photobiol. 2006, 82, 1178–1186. [Google Scholar] [CrossRef]
- Maharjan, P.S.; Bhattarai, H.K. Singlet Oxygen, Photodynamic Therapy, and Mechanisms of Cancer Cell Death. J. Oncol. 2022, 2022, 7211485. [Google Scholar] [CrossRef]
- Gilyazova, D.G.; Rosenkranz, A.A.; Gulak, P.V.; Lunin, V.G.; Sergienko, O.V.; Khramtsov, Y.V.; Timofeyev, K.N.; Grin, M.A.; Mironov, A.F.; Rubin, A.B.; et al. Targeting cancer cells by novel engineered modular transporters. Cancer Res. 2006, 66, 10534–10540. [Google Scholar] [CrossRef]
- Yaqoob, M.D.; Xu, L.; Li, C.; Leong, M.M.L.; Xu, D.D. Targeting mitochondria for cancer photodynamic therapy. Photodiagnosis Photodyn. Ther. 2022, 38, 102830. [Google Scholar] [CrossRef] [PubMed]
- Hilf, R. Mitochondria are targets of photodynamic therapy. J. Bioenerg. Biomembr. 2007, 39, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Lv, W.; Zhang, Z.; Zhang, K.Y.; Yang, H.; Liu, S.; Xu, A.; Guo, S.; Zhao, Q.; Huang, W. A Mitochondria-Targeted Photosensitizer Showing Improved Photodynamic Therapy Effects under Hypoxia. Angew. Chem. Int. Ed. Engl. 2016, 55, 9947–9951. [Google Scholar] [CrossRef]
- Biswas, S.; Torchilin, V.P. Nanopreparations for organelle-specific delivery in cancer. Adv. Drug Deliv. Rev. 2014, 66, 26–41. [Google Scholar] [CrossRef]
- Marschall, A.L.J. Targeting the Inside of Cells with Biologicals: Chemicals as a Delivery Strategy. BioDrugs 2021, 35, 643–671. [Google Scholar] [CrossRef]
- Porello, I.; Cellesi, F. Intracellular delivery of therapeutic proteins. New advancements and future directions. Front. Bioeng. Biotechnol. 2023, 11, 1211798. [Google Scholar] [CrossRef]
- Rosenkranz, A.A.; Slastnikova, T.A. Prospects of Using Protein Engineering for Selective Drug Delivery into a Specific Compartment of Target Cells. Pharmaceutics 2023, 15, 987. [Google Scholar] [CrossRef] [PubMed]
- Khramtsov, Y.V.; Ulasov, A.V.; Slastnikova, T.A.; Rosenkranz, A.A.; Lupanova, T.N.; Georgiev, G.P.; Sobolev, A.S. Modular Nanotransporters Delivering Biologically Active Molecules to the Surface of Mitochondria. Pharmaceutics 2023, 15, 2687. [Google Scholar] [CrossRef]
- Ilari, A.; Bonamore, A.; Farina, A.; Johnson, K.A.; Boffi, A. The X-ray structure of ferric Escherichia coli flavohemoglobin reveals an unexpected geometry of the distal heme pocket. J. Biol. Chem. 2002, 277, 23725–23732. [Google Scholar] [CrossRef]
- Dreher, M.R.; Liu, W.; Michelich, C.R.; Dewhirst, M.W.; Yuan, F.; Chilkoti, A. Tumor vascular permeability, accumulation, and penetration of macromolecular drug carriers. J. Natl. Cancer Inst. 2006, 98, 335–344. [Google Scholar] [CrossRef]
- Suzuki, T.; Takahashi, J.; Yamamoto, M. Molecular Basis of the KEAP1-NRF2 Signaling Pathway. Mol. Cells 2023, 46, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.; Kensler, T.W.; Motohashi, H. The KEAP1-NRF2 System: A Thiol-Based Sensor-Effector Apparatus for Maintaining Redox Homeostasis. Physiol. Rev. 2018, 98, 1169–1203. [Google Scholar] [CrossRef] [PubMed]
- Hayes, J.D.; Dinkova-Kostova, A.T. The Nrf2 regulatory network provides an interface between redox and intermediary metabolism. Trends Biochem. Sci. 2014, 39, 199–218. [Google Scholar] [CrossRef]
- Kopacz, A.; Kloska, D.; Forman, H.J.; Jozkowicz, A.; Grochot-Przeczek, A. Beyond repression of Nrf2: An update on Keap1. Free Radic. Biol. Med. 2020, 157, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Lo, S.C.; Hannink, M. PGAM5 tethers a ternary complex containing Keap1 and Nrf2 to mitochondria. Exp. Cell Res. 2008, 314, 1789–1803. [Google Scholar] [CrossRef]
- Trower, M.K. A rapid PCR-based colony screening protocol for cloned inserts. Methods Mol. Biol. 1996, 58, 329–333. [Google Scholar] [PubMed]
- Ota, M.; Asamura, H.; Oki, T.; Sada, M. Restriction enzyme analysis of PCR products. Methods Mol. Biol. 2009, 578, 405–414. [Google Scholar] [PubMed]
- Sanger, F.; Nicklen, S.; Coulson, A.R. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 1977, 74, 5463–5467. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Slastnikova, T.A.; Rosenkranz, A.A.; Ulasov, A.V.; Khramtsov, Y.V.; Lupanova, T.N.; Georgiev, G.P.; Sobolev, A.S. Mouse Syngeneic Melanoma Model with Human Epidermal Growth Factor Receptor Expression. Pharmaceutics 2022, 14, 2448. [Google Scholar] [CrossRef]
- Szoka, F., Jr.; Papahadjopoulos, D. Procedure for preparation of liposomes with large internal aqueous space and high capture by reverse-phase evaporation. Proc. Natl. Acad. Sci. USA 1978, 75, 4194–4198. [Google Scholar] [CrossRef] [PubMed]
- Karyagina, T.S.; Ulasov, A.V.; Slastnikova, T.A.; Rosenkranz, A.A.; Lupanova, T.N.; Khramtsov, Y.V.; Georgiev, G.P.; Sobolev, A.S. Targeted Delivery of 111In into the Nuclei of EGFR Overexpressing Cells via Modular Nanotransporters with Anti-EGFR Affibody. Front. Pharmacol. 2020, 11, 176. [Google Scholar] [CrossRef] [PubMed]
- Turubanova, V.D.; Balalaeva, I.V.; Mishchenko, T.A.; Catanzaro, E.; Alzeibak, R.; Peskova, N.N.; Efimova, I.; Bachert, C.; Mitroshina, E.V.; Krysko, O.; et al. Immunogenic cell death induced by a new photodynamic therapy based on photosens and photodithazine. J. Immunother. Cancer 2019, 7, 350. [Google Scholar] [CrossRef] [PubMed]
- Barreca, M.; Ingarra, A.M.; Raimondi, M.V.; Spano, V.; De, F.M.; Menilli, L.; Gandin, V.; Miolo, G.; Barraja, P.; Montalbano, A. Insight on pyrimido[5,4-g]indolizine and pyrimido[4,5-c]pyrrolo[1,2-a]azepine systems as promising photosensitizers on malignant cells. Eur. J. Med. Chem. 2022, 237, 114399. [Google Scholar] [CrossRef] [PubMed]
- Schary, N.; Novak, B.; Kamper, L.; Yousf, A.; Lubbert, H. Identification and pharmacological modification of resistance mechanisms to protoporphyrin-mediated photodynamic therapy in human cutaneous squamous cell carcinoma cell lines. Photodiagnosis Photodyn. Ther. 2022, 39, 103004. [Google Scholar] [CrossRef] [PubMed]
- Jerabek-Willemsen, M.; Andre, T.; Wanner, R.; Roth, H.M.; Duhr, S.; Baaske, P.; Breitsprecher, D. MicroScale Thermophoresis: Interaction analysis and beyond. J. Mol. Struct. 2014, 1077, 101–113. [Google Scholar] [CrossRef]
- Ladokhin, A.S. pH-triggered conformational switching along the membrane insertion pathway of the diphtheria toxin T-domain. Toxins 2013, 5, 1362–1380. [Google Scholar] [CrossRef] [PubMed]
- Leka, O.; Vallese, F.; Pirazzini, M.; Berto, P.; Montecucco, C.; Zanotti, G. Diphtheria toxin conformational switching at acidic pH. FEBS J. 2014, 281, 2115–2122. [Google Scholar] [CrossRef] [PubMed]
- Rodnin, M.V.; Kyrychenko, A.; Kienker, P.; Sharma, O.; Posokhov, Y.O.; Collier, R.J.; Finkelstein, A.; Ladokhin, A.S. Conformational switching of the diphtheria toxin T domain. J. Mol. Biol. 2010, 402, 1–7. [Google Scholar] [CrossRef]
- Di, G.A.; Bonamore, A. Globin interactions with lipids and membranes. Methods Enzymol. 2008, 436, 239–253. [Google Scholar]
- Sui, M.; Liu, W.; Shen, Y. Nuclear drug delivery for cancer chemotherapy. J. Control. Release 2011, 155, 227–236. [Google Scholar] [CrossRef]
- Pan, L.; Liu, J.; Shi, J. Cancer cell nucleus-targeting nanocomposites for advanced tumor therapeutics. Chem. Soc. Rev. 2018, 47, 6930–6946. [Google Scholar] [CrossRef]
- Tiwari, R.; Jain, P.; Asati, S.; Haider, T.; Soni, V.; Pandey, V. State-of-art based approaches for anticancer drug-targeting to nucleus. J. Drug Deliv. Sci. Technol. 2018, 48, 383–392. [Google Scholar] [CrossRef]
- Goyal, P.; Malviya, R. Advances in nuclei targeted delivery of nanoparticles for the management of cancer. Biochim. Biophys. Acta Rev. Cancer 2023, 1878, 188881. [Google Scholar] [CrossRef] [PubMed]
- Bellezza, I.; Giambanco, I.; Minelli, A.; Donato, R. Nrf2-Keap1 signaling in oxidative and reductive stress. Biochim. Biophys. Acta Mol. Cell Res. 2018, 1865, 721–733. [Google Scholar] [CrossRef]
- Nguyen, T.; Nioi, P.; Pickett, C.B. The Nrf2-antioxidant response element signaling pathway and its activation by oxidative stress. J. Biol. Chem. 2009, 284, 13291–13295. [Google Scholar] [CrossRef]
- Baird, L.; Dinkova-Kostova, A.T. The cytoprotective role of the Keap1-Nrf2 pathway. Arch. Toxicol. 2011, 85, 241–272. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Motohashi, H.; Yamamoto, M. Toward clinical application of the Keap1-Nrf2 pathway. Trends Pharmacol. Sci. 2013, 34, 340–346. [Google Scholar] [CrossRef] [PubMed]
- Guntas, G.; Lewis, S.M.; Mulvaney, K.M.; Cloer, E.W.; Tripathy, A.; Lane, T.R.; Major, M.B.; Kuhlman, B. Engineering a genetically encoded competitive inhibitor of the KEAP1-NRF2 interaction via structure-based design and phage display. Protein Eng. Des. Sel. 2016, 29, 1–9. [Google Scholar] [CrossRef]
- Mou, Y.; Wen, S.; Li, Y.X.; Gao, X.X.; Zhang, X.; Jiang, Z.Y. Recent progress in Keap1-Nrf2 protein-protein interaction inhibitors. Eur. J. Med. Chem. 2020, 202, 112532. [Google Scholar] [CrossRef]
- Ulasov, A.V.; Rosenkranz, A.A.; Georgiev, G.P.; Sobolev, A.S. Nrf2/Keap1/ARE signaling: Towards specific regulation. Life Sci. 2022, 291, 120111. [Google Scholar] [CrossRef]
- Zeb, A.; Choubey, V.; Gupta, R.; Kuum, M.; Safiulina, D.; Vaarmann, A.; Gogichaishvili, N.; Liiv, M.; Ilves, I.; Tamm, K.; et al. A novel role of KEAP1/PGAM5 complex: ROS sensor for inducing mitophagy. Redox. Biol. 2021, 48, 102186. [Google Scholar] [CrossRef] [PubMed]
- McCutcheon, D.C.; Lee, G.; Carlos, A.; Montgomery, J.E.; Moellering, R.E. Photoproximity Profiling of Protein-Protein Interactions in Cells. J. Am. Chem. Soc. 2020, 142, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Li, X.; Yoon, J. Organelle-Targeted Photosensitizers for Precision Photodynamic Therapy. ACS Appl. Mater. Interfaces 2021, 13, 19543–19571. [Google Scholar] [CrossRef] [PubMed]
- Pham, T.C.; Nguyen, V.N.; Choi, Y.; Lee, S.; Yoon, J. Recent Strategies to Develop Innovative Photosensitizers for Enhanced Photodynamic Therapy. Chem. Rev. 2021, 121, 13454–13619. [Google Scholar] [CrossRef]
- Desai, V.M.; Choudhary, M.; Chowdhury, R.; Singhvi, G. Photodynamic Therapy Induced Mitochondrial Targeting Strategies for Cancer Treatment: Emerging Trends and Insights. Mol. Pharm. 2024, 21, 1591–1608. [Google Scholar] [CrossRef]
- Bock, F.J.; Tait, S.W.G. Mitochondria as multifaceted regulators of cell death. Nat. Rev. Mol. Cell Biol. 2020, 21, 85–100. [Google Scholar] [CrossRef]
- Nath, R.; Sengupta, S.; Bhattacharjee, A. Enzymatic and Non-Enzymatic Response during Nitrosative Stress in. POSTĘPY MIKROBIOLOGII-Adv. Microbiol. 2022, 61, 81–93. [Google Scholar]
- Poole, R.K.; Hughes, M.N. New functions for the ancient globin family: Bacterial responses to nitric oxide and nitrosative stress. Mol. Microbiol. 2000, 36, 775–783. [Google Scholar] [CrossRef]
- Forrester, M.T.; Eyler, C.E.; Rich, J.N. Bacterial flavohemoglobin: A molecular tool to probe mammalian nitric oxide biology. Biotechniques 2011, 50, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Mukai, M.; Mills, C.E.; Poole, R.K.; Yeh, S.R. Flavohemoglobin, a globin with a peroxidase-like catalytic site. J. Biol. Chem. 2001, 276, 7272–7277. [Google Scholar] [CrossRef] [PubMed]
- Membrillo-Hernandez, J.; Coopamah, M.D.; Anjum, M.F.; Stevanin, T.M.; Kelly, A.; Hughes, M.N.; Poole, R.K. The flavohemoglobin of Escherichia coli confers resistance to a nitrosating agent, a “Nitric oxide Releaser,” and paraquat and is essential for transcriptional responses to oxidative stress. J. Biol. Chem. 1999, 274, 748–754. [Google Scholar] [CrossRef] [PubMed]
- Frey, A.D.; Farres, J.; Bollinger, C.J.; Kallio, P.T. Bacterial hemoglobins and flavohemoglobins for alleviation of nitrosative stress in Escherichia coli. Appl. Environ. Microbiol. 2002, 68, 4835–4840. [Google Scholar] [CrossRef] [PubMed]
- Buytaert, E.; Dewaele, M.; Agostinis, P. Molecular effectors of multiple cell death pathways initiated by photodynamic therapy. Biochim. Biophys. Acta 2007, 1776, 86–107. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alieva, R.T.; Ulasov, A.V.; Khramtsov, Y.V.; Slastnikova, T.A.; Lupanova, T.N.; Gribova, M.A.; Georgiev, G.P.; Rosenkranz, A.A. Optimization of a Modular Nanotransporter Design for Targeted Intracellular Delivery of Photosensitizer. Pharmaceutics 2024, 16, 1083. https://doi.org/10.3390/pharmaceutics16081083
Alieva RT, Ulasov AV, Khramtsov YV, Slastnikova TA, Lupanova TN, Gribova MA, Georgiev GP, Rosenkranz AA. Optimization of a Modular Nanotransporter Design for Targeted Intracellular Delivery of Photosensitizer. Pharmaceutics. 2024; 16(8):1083. https://doi.org/10.3390/pharmaceutics16081083
Chicago/Turabian StyleAlieva, Rena T., Alexey V. Ulasov, Yuri V. Khramtsov, Tatiana A. Slastnikova, Tatiana N. Lupanova, Maria A. Gribova, Georgii P. Georgiev, and Andrey A. Rosenkranz. 2024. "Optimization of a Modular Nanotransporter Design for Targeted Intracellular Delivery of Photosensitizer" Pharmaceutics 16, no. 8: 1083. https://doi.org/10.3390/pharmaceutics16081083