

A Bionic “Trojan Horse”-like Nanovesicle Delivery System Hybridized with BCG Cytoplasmic Membrane and Melanoma Cell Membrane for Cancer Immunotherapy
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
3. Results
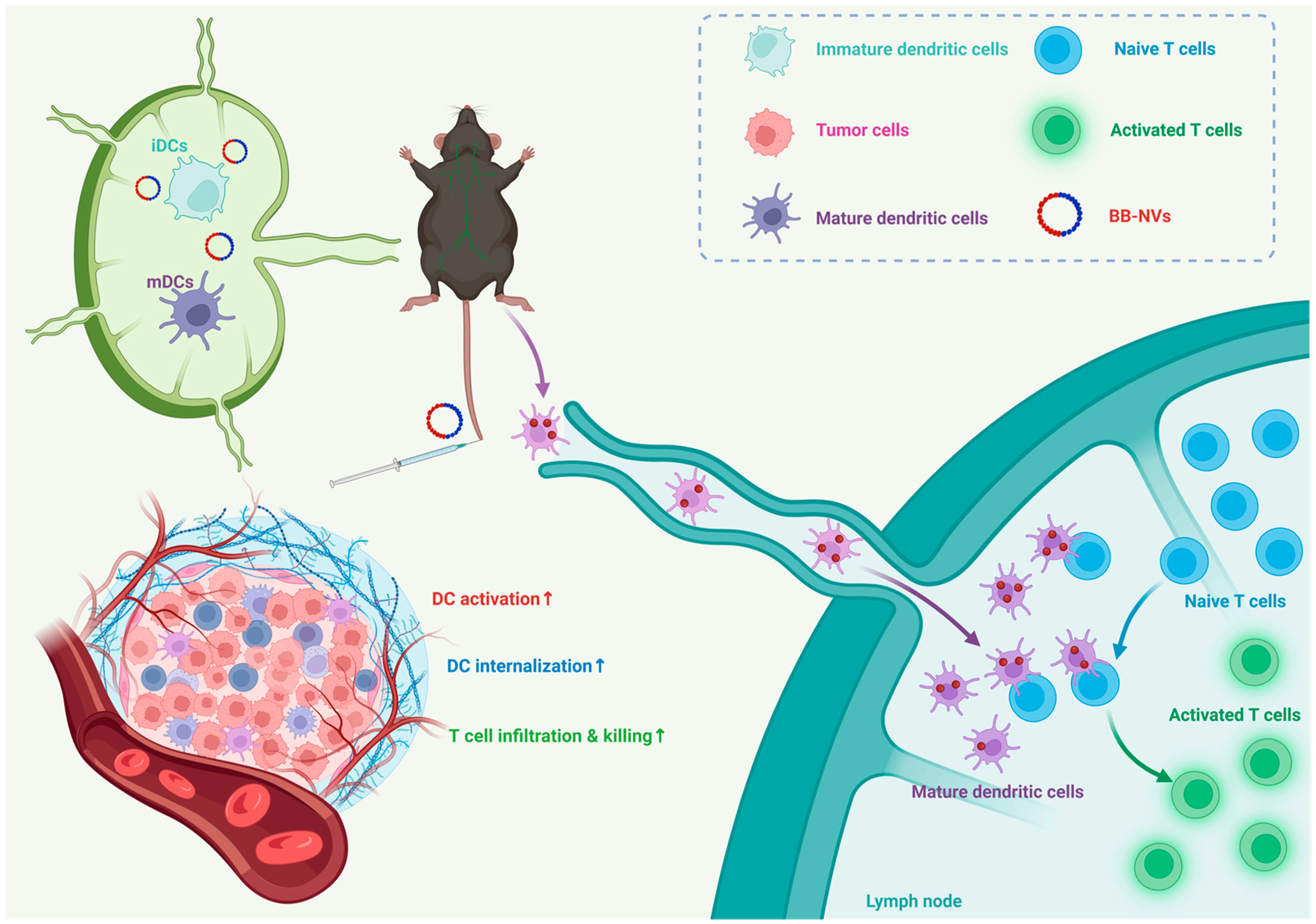
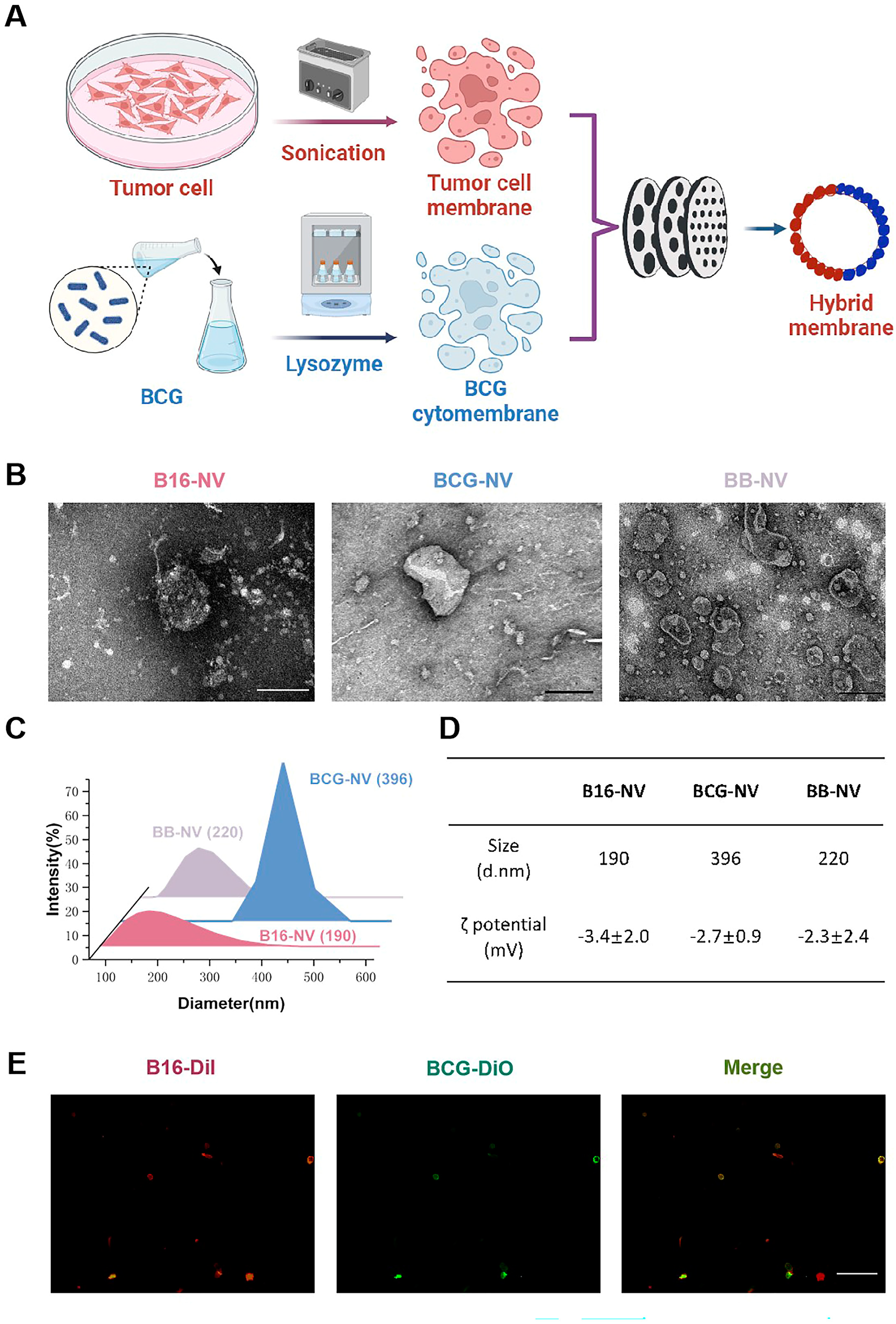
3.1. Preparation and Characterization of B16-NVs, BCG-NVs, and BB-NVs
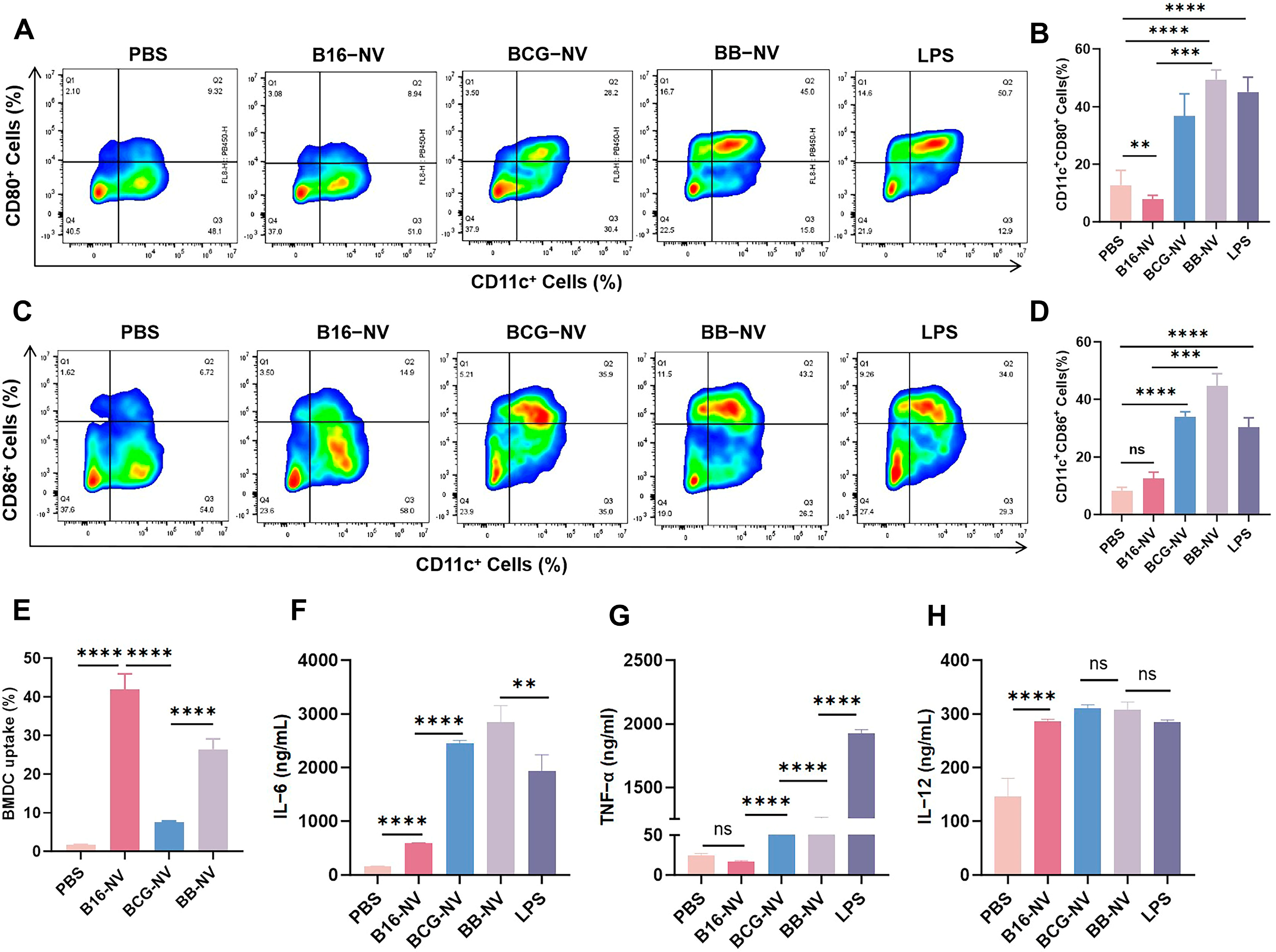
3.2. BB-NVs Increase Tumor Antigen Uptake and Activate Bone Marrow-Derived Dendritic Cells (BMDCs) by Co-Delivery of Antigen and Adjuvant
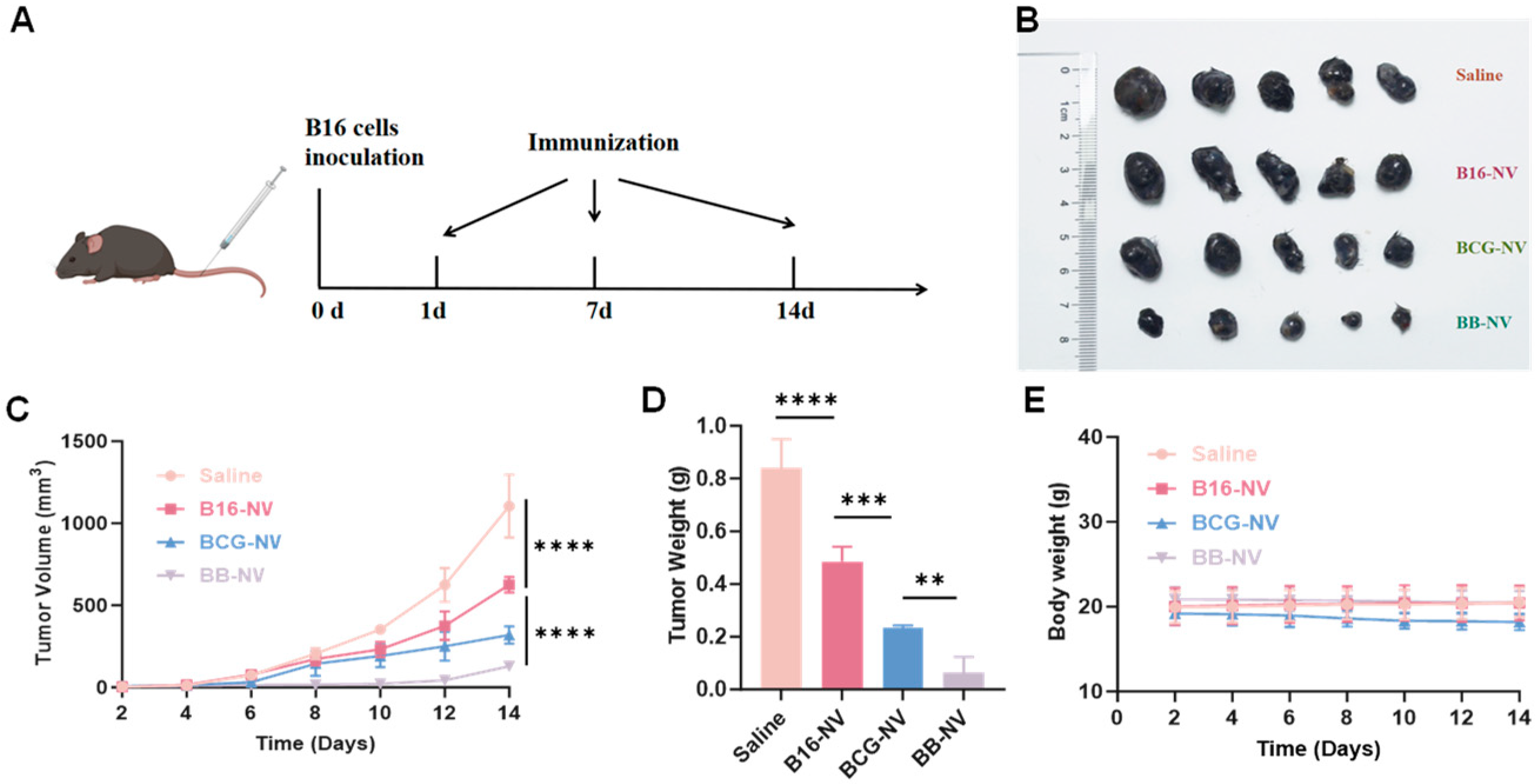
3.3. BB-NVs Vaccination Inhibits Tumor Progression in Murine B16 Tumor Model
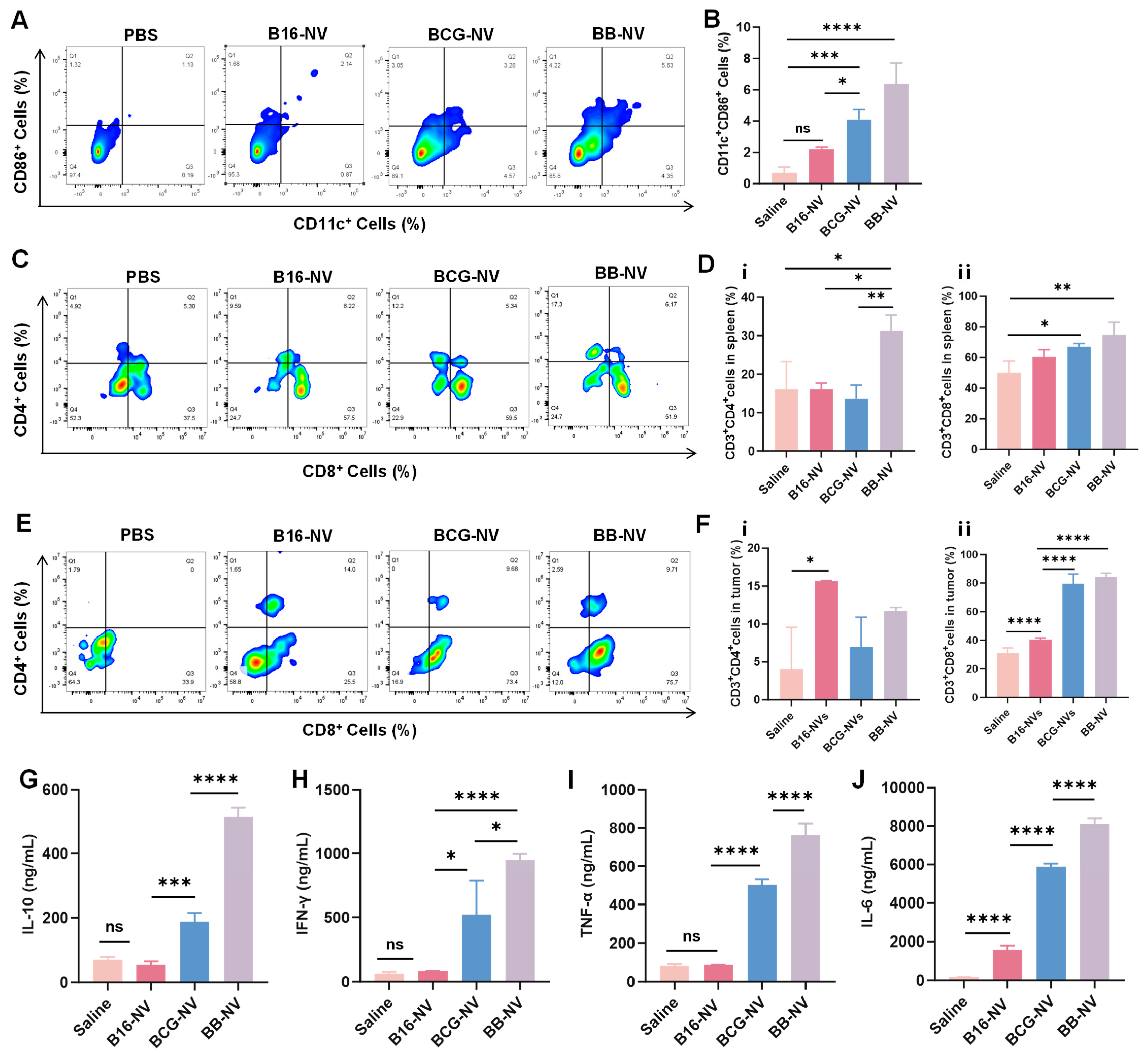
3.4. BB-NVs Activate Immune Cells and Promote Anti-Tumor Cytokine Secretion in Mice
3.5. Biosafety Assessment of B16-NVs, BCG-NVs, and BB-NVs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| APC | Antigen-Presenting Cells |
| BCG | Bacillus Calmette–Guérin |
| DMEM | Dulbecco’s Modified Eagle Medium |
| DCs | Dendritic Cells |
| FBS | Fetal Bovine Serum |
| PBS | Phosphate-Buffered Saline |
| TEM | Transmission Electron Microscopy |
| DLS | Dynamic Light Scattering |
| ELISA | Enzyme-Linked Immunosorbent Assay |
| FITC | Fluoresce-Activated Cell Sorting |
| H&E | Hematoxylin and Eosin Staining |
| SEM | Scanning Electron Microscopy |
| °C | Celsius |
References
- Leko, V.; Rosenberg, S.A. Identifying and Targeting Human Tumor Antigens for T Cell-Based Immunotherapy of Solid Tumors. Cancer Cell 2020, 38, 454–472. [Google Scholar] [CrossRef] [PubMed]
- Dagher, O.K.; Schwab, R.D.; Brookens, S.K.; Posey, A.D. Advances in cancer immunotherapies. Cell 2023, 186, 1814–1814.e1. [Google Scholar] [CrossRef]
- Sun, Z.; Zhao, H.; Ma, L.; Shi, Y.; Ji, M.; Sun, X.; Ma, D.; Zhou, W.; Huang, T.; Zhang, D. The quest for nanoparticle-powered vaccines in cancer immunotherapy. J. Nanobiotechnol. 2024, 22, 61. [Google Scholar] [CrossRef] [PubMed]
- Baharom, F.; Ramirez-Valdez, R.A.; Khalilnezhad, A.; Khalilnezhad, S.; Dillon, M.; Hermans, D.; Fussell, S.; Tobin, K.K.; Dutertre, C.-A.; Lynn, G.M.; et al. Systemic vaccination induces CD8+ T cells and remodels the tumor microenvironment. Cell 2022, 185, 4317–4332.e15. [Google Scholar] [CrossRef]
- Lin, M.J.; Svensson-Arvelund, J.; Lubitz, G.S.; Marabelle, A.; Melero, I.; Brown, B.D.; Brody, J.D. Cancer vaccines: The next immunotherapy frontier. Nat. Cancer 2022, 3, 911–926. [Google Scholar] [CrossRef]
- Pan, Y.; Wu, X.; Liu, L.; Zhao, C.; Zhang, J.; Yang, S.; Pan, P.; Huang, Q.; Zhao, X.; Tian, R.; et al. Genetically Engineered Cytomembrane Nanovaccines for Cancer Immunotherapy. Adv. Healthc. Mater. 2024, 13, e2400068. [Google Scholar] [CrossRef]
- Heras-Murillo, I.; Adán-Barrientos, I.; Galán, M.; Wculek, S.K.; Sancho, D. Dendritic cells as orchestrators of anticancer immunity and immunotherapy. Nat. Rev. Clin. Oncol. 2024, 21, 257–277. [Google Scholar] [CrossRef] [PubMed]
- Oladejo, M.; Tijani, A.O.; Puri, A.; Chablani, L. Adjuvants in cutaneous vaccination: A comprehensive analysis. J. Control. Release 2024, 369, 475–492. [Google Scholar] [CrossRef]
- Saxena, M.; van der Burg, S.H.; Melief, C.J.; Bhardwaj, N. Therapeutic cancer vaccines. Nat. Rev. Cancer 2021, 21, 360–378. [Google Scholar] [CrossRef]
- Le, Q.-V.; Lee, J.; Lee, H.; Shim, G.; Oh, Y.-K. Cell membrane-derived vesicles for delivery of therapeutic agents. Acta Pharm. Sin. B 2021, 11, 2096–2113. [Google Scholar] [CrossRef]
- Xiao, Y.; Wu, M.; Xue, C.; Wang, Y. Recent Advances in the Development of Membrane-derived Vesicles for Cancer Immunotherapy. Curr. Drug Deliv. 2024, 21, 403–420. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Ju, D.; Zhang, X. Cell Membrane-Derived Vesicle: A Novel Vehicle for Cancer Immunotherapy. Front. Immunol. 2022, 13, 923598. [Google Scholar] [CrossRef]
- Marianecci, C.; Carafa, M. Smart Nanovesicles for Drug Targeting and Delivery. Pharmaceutics 2019, 11, 147. [Google Scholar] [CrossRef]
- Cheng, Q.; Kang, Y.; Yao, B.; Dong, J.; Zhu, Y.; He, Y.; Ji, X. Genetically Engineered-Cell-Membrane Nanovesicles for Cancer Immunotherapy. Adv. Sci. 2023, 10, e2302131. [Google Scholar] [CrossRef]
- Zhu, L.; Zhong, Y.; Wu, S.; Yan, M.; Cao, Y.; Mou, N.; Wang, G.; Sun, D.; Wu, W. Cell membrane camouflaged biomimetic nanoparticles: Focusing on tumor theranostics. Mater. Today Bio 2022, 14, 100228. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.; Huang, J. Application of Biomimetic Nanoparticles based on the Cell Membrane in Tumor Therapy. Curr. Top. Med. Chem. 2023, 23, 907–920. [Google Scholar] [CrossRef] [PubMed]
- Jakubechova, J.; Altanerova, U.; Altaner, C. Tumor-targeted suicide gene-directed enzyme prodrug therapy mediated by extracellular vesicles. Neoplasma 2023, 70, 333–339. [Google Scholar] [CrossRef]
- Sautès-Fridman, C.; Lawand, M.; Giraldo, N.A.; Kaplon, H.; Germain, C.; Fridman, W.H.; Dieu-Nosjean, M.-C. Tertiary Lymphoid Structures in Cancers: Prognostic Value, Regulation, and Manipulation for Therapeutic Intervention. Front. Immunol. 2016, 7, 407. [Google Scholar] [CrossRef] [PubMed]
- Pettenati, C.; Ingersoll, M.A. Mechanisms of BCG immunotherapy and its outlook for bladder cancer. Nat. Rev. Urol. 2018, 15, 615–625. [Google Scholar] [CrossRef]
- Benitez, M.L.R.; Bender, C.B.; Oliveira, T.L.; Schachtschneider, K.M.; Collares, T.; Seixas, F.K. Mycobacterium bovis BCG in metastatic melanoma therapy. Appl. Microbiol. Biotechnol. 2019, 103, 7903–7916. [Google Scholar] [CrossRef]
- Ramalingam, S.; Gunasekaran, K.; Arora, H.; Muruganandam, M.; Nagaraju, S.; Padmanabhan, P. Disseminated BCG Infection after intravesical BCG Immunotherapy of Bladder Cancer. QJM Int. J. Med. 2021, 114, 410–411. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Jiang, Y.-L.; Men, Y.; Jiao, Y.-Z.; Chen, S. Kinetic cation effect in alkaline hydrogen electrocatalysis and double layer proton transfer. Nat. Commun. 2025, 16, 1844. [Google Scholar] [CrossRef]
- Storgard, R.; Markova, A. Cutaneous hypersensitivity reactions to immune checkpoint inhibitors. J. Allergy Clin. Immunol. Pract. 2024, 12, 1132–1136. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Mu, J.; Chen, Y.; Liu, Y.; Li, X.; Li, H.; Cao, P. Hybrid Ginseng-derived Extracellular Vesicles-Like Particles with Autologous Tumor Cell Membrane for Personalized Vaccination to Inhibit Tumor Recurrence and Metastasis. Adv. Sci. 2024, 11, e2308235. [Google Scholar] [CrossRef]
- Kuai, R.; Ochyl, L.J.; Bahjat, K.S.; Schwendeman, A.; Moon, J.J. Designer vaccine nanodiscs for personalized cancer immunotherapy. Nat. Mater. 2017, 16, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Sang, W.; Zhang, Z.; Dai, Y.; Chen, X. Recent advances in nanomaterial-based synergistic combination cancer immunotherapy. Chem. Soc. Rev. 2019, 48, 3771–3810. [Google Scholar] [CrossRef]
- Zhao, J.; Xu, Y.; Ma, S.; Wang, Y.; Huang, Z.; Qu, H.; Yao, H.; Zhang, Y.; Wu, G.; Huang, L.; et al. A Minimalist Binary Vaccine Carrier for Personalized Postoperative Cancer Vaccine Therapy. Adv. Mater. 2022, 34, 2109254. [Google Scholar] [CrossRef]
- Raposo, G.; Stahl, P.D. Extracellular vesicles, genetic programmers. Nat. Cell Biol. 2024, 26, 22–23. [Google Scholar] [CrossRef]
- Zhao, X.; Zhao, R.; Nie, G. Nanocarriers based on bacterial membrane materials for cancer vaccine delivery. Nat. Protoc. 2022, 17, 2240–2274. [Google Scholar] [CrossRef]
- Kremenovic, M.; Chan, A.A.; Feng, B.; Bäriswyl, L.; Robatel, S.; Gruber, T.; Tang, L.; Lee, D.J.; Schenk, M. BCG hydrogel promotes CTSS-mediated antigen processing and presentation, thereby suppressing metastasis and prolonging survival in melanoma. J. Immunother. Cancer 2022, 10, e004133. [Google Scholar] [CrossRef]
- van Puffelen, J.H.; Keating, S.T.; Oosterwijk, E.; van der Heijden, A.G.; Netea, M.G.; Joosten, L.A.; Vermeulen, S.H. Trained immunity as a molecular mechanism for BCG immunotherapy in bladder cancer. Nat. Rev. Urol. 2020, 17, 513–525. [Google Scholar] [CrossRef] [PubMed]
- Kleinnijenhuis, J.; Quintin, J.; Preijers, F.; Joosten, L.A.; Jacobs, C.; Xavier, R.J.; van der Meer, J.W.; van Crevel, R.; Netea, M.G. BCG-induced trained immunity in NK cells: Role for non-specific protection to infection. Clin. Immunol. 2014, 155, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Jones, B.W.; Means, T.K.; Heldwein, K.A.; Keen, M.A.; Hill, P.J.; Belisle, J.T.; Fenton, M.J. Different Toll-like receptor agonists induce distinct macrophage responses. J. Leukoc. Biol. 2001, 69, 1036–1044. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-W.; Jung, J.; Lee, Y.; Kim, T.-Y.; Choi, S.-Y.; Park, J.; Kim, D.-S.; Kwon, H.-J. Immunostimulatory oligodeoxynucleotide isolated from genome wide screening of Mycobacterium bovis chromosomal DNA. Mol. Immunol. 2006, 43, 2107–2118. [Google Scholar] [CrossRef]
- Tsuji, S.; Matsumoto, M.; Takeuchi, O.; Akira, S.; Azuma, I.; Hayashi, A.; Toyoshima, K.; Seya, T. Maturation of human dendritic cells by cell wall skeleton of Mycobacterium bovis bacillus Calmette-Guérin: Involvement of toll-like receptors. Infect. Immun. 2000, 68, 6883–6890. [Google Scholar] [CrossRef]
- Sfakianos, J.P.; Salome, B.; Daza, J.; Farkas, A.; Bhardwaj, N.; Horowitz, A. Bacillus Calmette-Guerin (BCG): Its fight against pathogens and cancer. Urol. Oncol. Semin. Orig. Investig. 2020, 39, 121–129. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, Y.; Chen, K.; Hu, T.; Wang, Y.; Wang, J.; Lv, C.; Xu, J.; Zhang, X.; Li, A.; Chen, B.; et al. A Bionic “Trojan Horse”-like Nanovesicle Delivery System Hybridized with BCG Cytoplasmic Membrane and Melanoma Cell Membrane for Cancer Immunotherapy. Pharmaceutics 2025, 17, 507. https://doi.org/10.3390/pharmaceutics17040507
Xiao Y, Chen K, Hu T, Wang Y, Wang J, Lv C, Xu J, Zhang X, Li A, Chen B, et al. A Bionic “Trojan Horse”-like Nanovesicle Delivery System Hybridized with BCG Cytoplasmic Membrane and Melanoma Cell Membrane for Cancer Immunotherapy. Pharmaceutics. 2025; 17(4):507. https://doi.org/10.3390/pharmaceutics17040507
Chicago/Turabian StyleXiao, Yuai, Kexin Chen, Tianchi Hu, Yuchong Wang, Jing Wang, Chuan Lv, Jianguo Xu, Xinyi Zhang, Ang Li, Bingdi Chen, and et al. 2025. "A Bionic “Trojan Horse”-like Nanovesicle Delivery System Hybridized with BCG Cytoplasmic Membrane and Melanoma Cell Membrane for Cancer Immunotherapy" Pharmaceutics 17, no. 4: 507. https://doi.org/10.3390/pharmaceutics17040507
APA StyleXiao, Y., Chen, K., Hu, T., Wang, Y., Wang, J., Lv, C., Xu, J., Zhang, X., Li, A., Chen, B., Zhu, J., Wu, M., & Xue, C. (2025). A Bionic “Trojan Horse”-like Nanovesicle Delivery System Hybridized with BCG Cytoplasmic Membrane and Melanoma Cell Membrane for Cancer Immunotherapy. Pharmaceutics, 17(4), 507. https://doi.org/10.3390/pharmaceutics17040507