
HSPB4/CRYAA Protect Photoreceptors during Retinal Detachment in Part through FAIM2 Regulation
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Model of Retinal Detachment
2.3. Immunoblot
2.4. Immunofluorescence
2.5. Cell Death Quantification
2.6. Retinal Thickness Measurements
2.7. Cell Culture
2.8. Transfection
2.9. Fas Receptor Activation
2.10. Immunoprecipitation
2.11. Statistical Analysis and ARRIVE Guidelines
3. Results
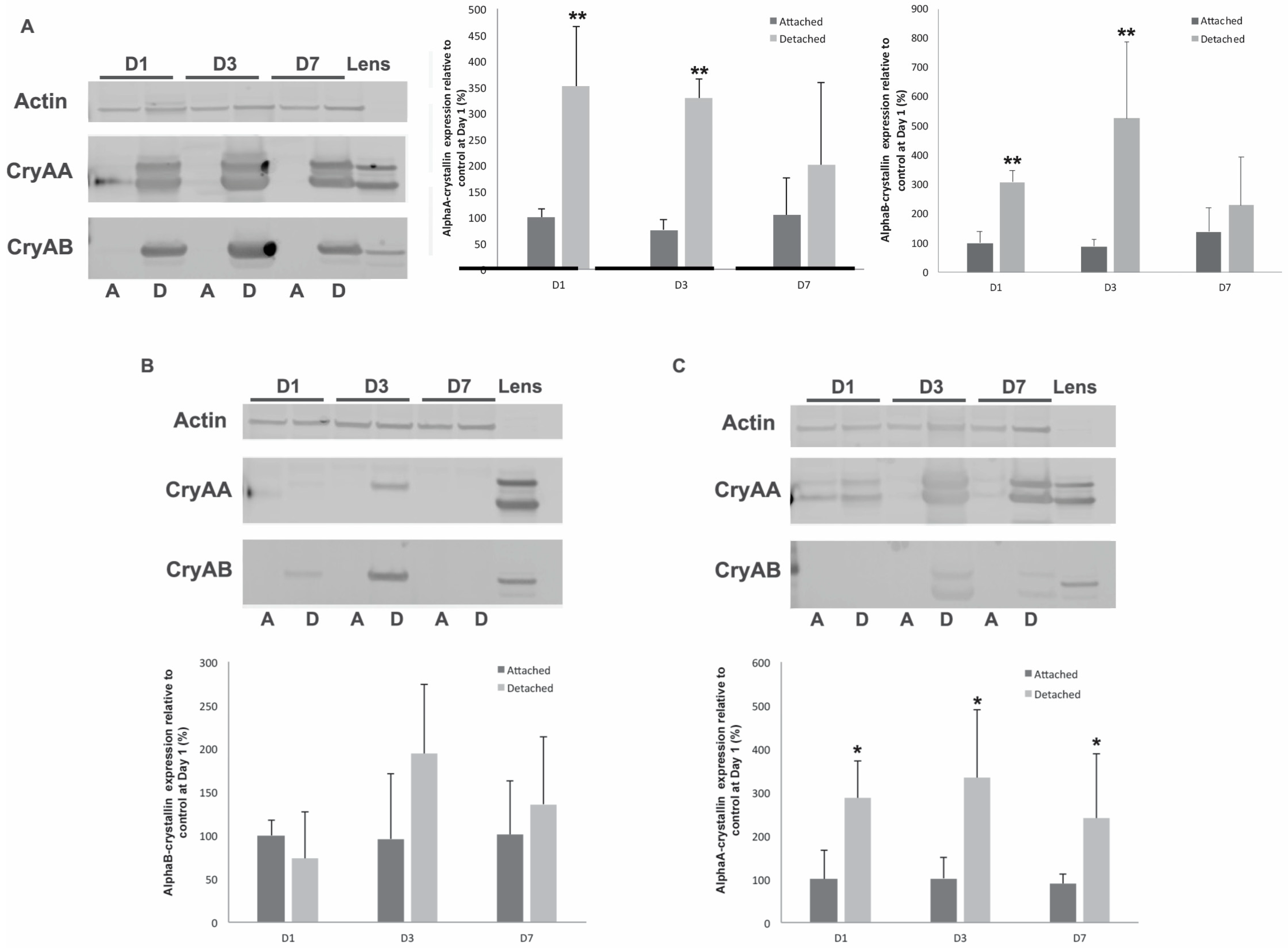
3.1. α-Crystallins Are Induced in the Detached Retina
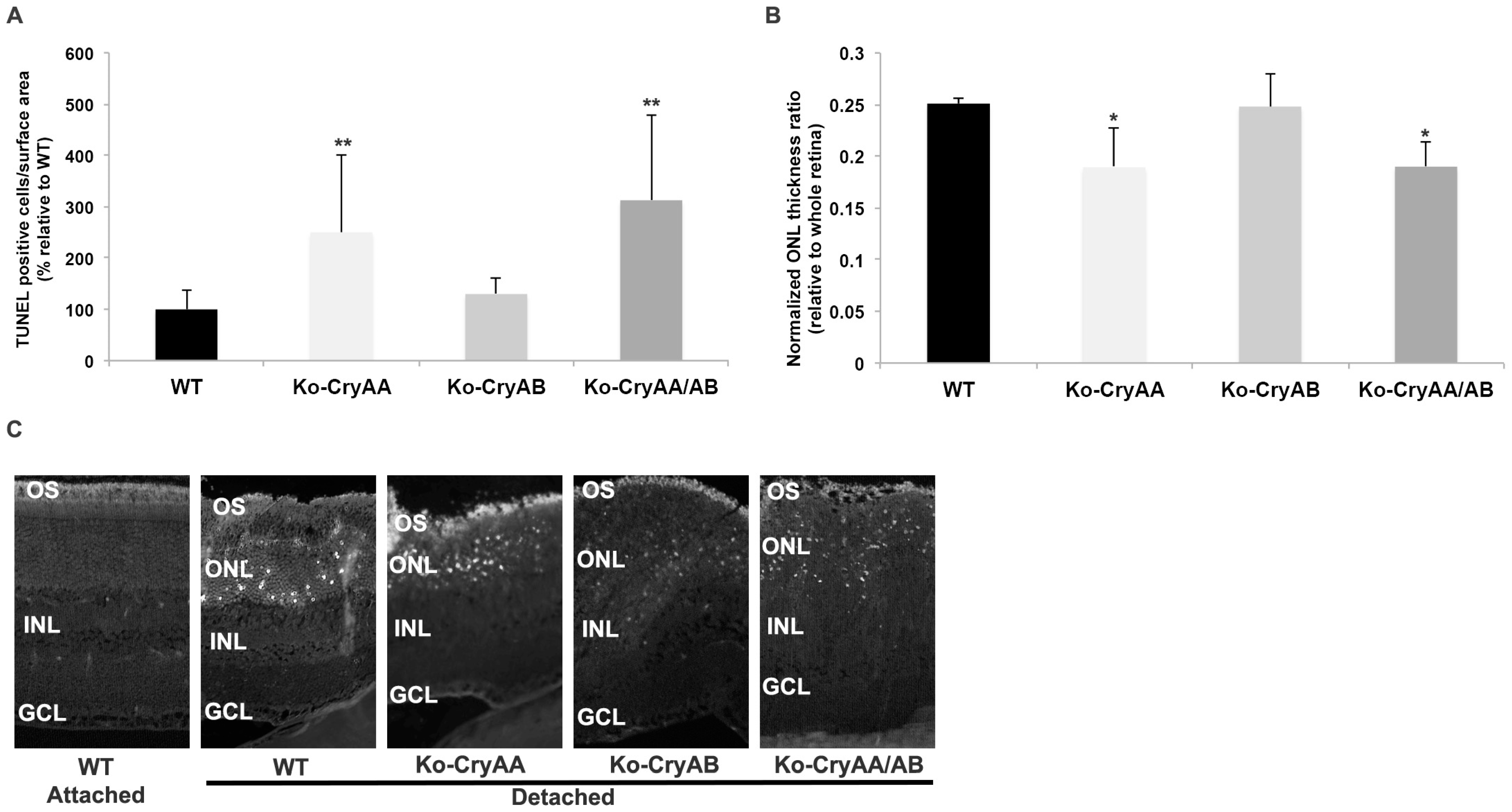
3.2. Loss of αA-Crystallins, but Not αB-Crystallins, Leads to Increased Photoreceptor Death after Retinal Detachment
3.3. α-Crystallins Are Induced and Accumulate in the Photoreceptor Inner and Outer Segments in the Detached Retina
3.4. FAIM2 Expression in Stressed Photoreceptors Requires the Presence of Both α-Crystallins
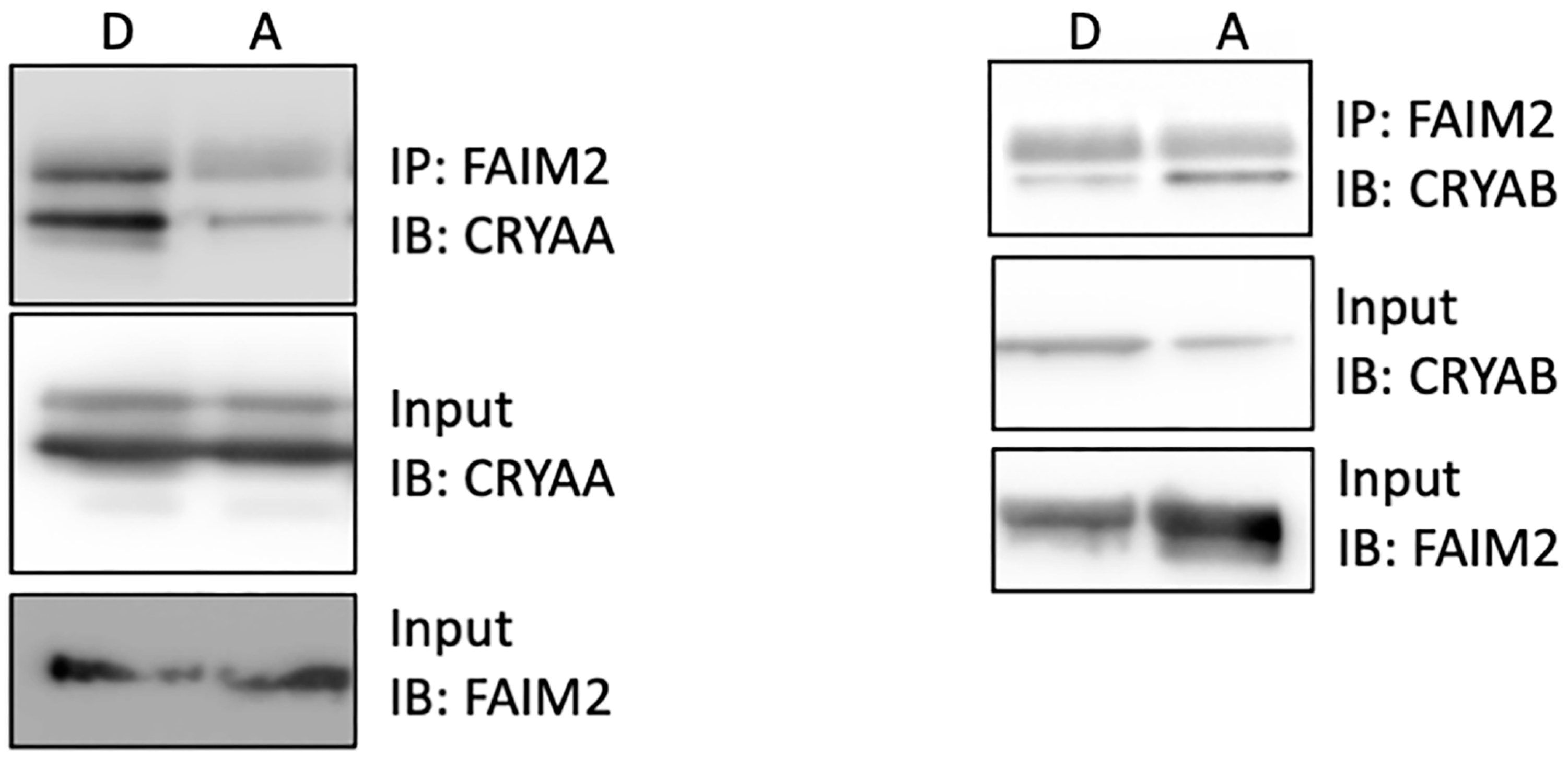
3.5. Increased Interaction of FAIM2 with αA-Crystallin after Retinal Detachment
3.6. FAIM2 Interaction with αA-Crystallin in Photoreceptors during Fas Activation Is T148 Phosphorylation-Dependent
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kwok, J.M.; Yu, C.W.; Christakis, P.G. Retinal detachment. CMAJ 2020, 192, E312. [Google Scholar] [CrossRef]
- Zacks, D.N.; Boehlke, C.; Richards, A.-L.; Zheng, Q.-D. Role of the fas-signaling pathway in photoreceptor neuroprotection. Arch. Ophthalmol. 2007, 125, 1389–1395. [Google Scholar] [CrossRef] [PubMed]
- Pawar, M.; Busov, B.; Chandrasekhar, A.; Yao, J.; Zacks, D.N.; Besirli, C.G. FAS apoptotic inhibitory molecule 2 is a stress-induced intrinsic neuroprotective factor in the retina. Cell Death Differ. 2017, 24, 1799–1810. [Google Scholar] [CrossRef]
- Besirli, C.G.; Chinskey, N.D.; Zheng, Q.-D.; Zacks, D.N. Autophagy activation in the injured photoreceptor inhibits fas-mediated apoptosis. Investig. Opthalmol. Vis. Sci. 2011, 52, 4193. [Google Scholar] [CrossRef] [PubMed]
- Chong, D.Y.; Boehlke, C.S.; Zheng, Q.-D.; Zhang, L.; Han, Y.; Zacks, D.N. Interleukin-6 as a photoreceptor neuroprotectant in an experimental model of retinal detachment. Investig. Opthalmol. Vis. Sci. 2008, 49, 3193–3200. [Google Scholar] [CrossRef] [PubMed]
- Shelby, S.J.; Angadi, P.S.; Zheng, Q.-D.; Yao, J.; Jia, L.; Zacks, D.N. Hypoxia inducible factor 1α contributes to regulation of autophagy in retinal detachment. Exp. Eye Res. 2015, 137, 84–93. [Google Scholar] [CrossRef]
- Besirli, C.G.; Zheng, Q.-D.; Reed, D.M.; Zacks, D.N. ERK-mediated activation of fas apoptotic inhibitory molecule 2 (Faim2) prevents apoptosis of 661W cells in a model of detachment-induced photoreceptor cell death. PLoS ONE 2012, 7, e46664. [Google Scholar] [CrossRef]
- Reich, A.; Spering, C.; Gertz, K.; Harms, C.; Gerhardt, E.; Kronenberg, G.; Nave, K.A.; Schwab, M.; Tauber, S.C.; Drinkut, A.; et al. Fas/CD95 Regulatory protein Faim2 is neuroprotective after transient brain ischemia. J. Neurosci. 2011, 31, 225–233. [Google Scholar] [CrossRef]
- Beier, C.P.; Wischhusen, J.; Gleichmann, M.; Gerhardt, E.; Pekanovic, A.; Krueger, A.; Taylor, V.; Suter, U.; Krammer, P.H.; Endres, M.; et al. FasL (CD95L/APO-1L) resistance of neurons mediated by phosphatidylinositol 3-kinase-Akt/protein kinase B-dependent expression of lifeguard/neuronal membrane protein 35. J. Neurosci. 2005, 25, 6765–6774. [Google Scholar] [CrossRef]
- Komnig, D.; Gertz, K.; Habib, P.; Nolte, K.W.; Meyer, T.; Brockmann, M.A.; Endres, M.; Rathkolb, B.; de Angelis, M.H.; German Mouse Clinic Consortium; et al. Faim2 contributes to neuroprotection by erythropoietin in transient brain ischemia. J. Neurochem. 2018, 145, 258–270. [Google Scholar] [CrossRef]
- Ruebsam, A.; Dulle, J.E.; Myers, A.M.; Sakrikar, D.; Green, K.M.; Khan, N.W.; Schey, K.; Fort, P.E. A specific phosphorylation regulates the protective role of αA-crystallin in diabetes. J. Clin. Investig. 2018, 3, e97919. [Google Scholar] [CrossRef]
- Rao, N.A.; Saraswathy, S.; Wu, G.S.; Katselis, G.S.; Wawrousek, E.F.; Bhat, S. Elevated retina-specific expression of the small heat shock protein, αA-crystallin, is associated with photoreceptor protection in experimental uveitis. Investig. Opthalmol. Vis. Sci. 2008, 49, 1161–1171. [Google Scholar] [CrossRef] [PubMed]
- Baba, T.; Oshitari, T.; Yamamoto, S. Level of vitreous alpha-B crystallin in eyes with rhegmatogenous retinal detachment. Graefe’s Arch. Clin. Exp. Ophthalmol. 2015, 253, 1251–1254. [Google Scholar] [CrossRef] [PubMed]
- Va’zquez-Chona, F.; Song, B.K.; Geisert, E.E. Temporal changes in gene expression after injury in the rat retina. Investig. Opthalmol. Vis. Sci. 2004, 45, 2737–2746. [Google Scholar] [CrossRef] [PubMed]
- Hamadmad, S.; Shah, M.H.; Kusibati, R.; Kim, B.; Erickson, B.; Heisler-Taylor, T.; Bhattacharya, S.K.; Abdel-Rahman, M.H.; Cebulla, C.M.; Van Law, H.; et al. Significant upregulation of small heat shock protein αA-crystallin in retinal detachment. Exp. Eye Res. 2019, 189, 107811. [Google Scholar] [CrossRef]
- Zacks, D.N. Gene transcription profile of the detached retina (An AOS Thesis). Trans. Am. Ophthalmol. Soc. 2009, 107, 343–382. [Google Scholar]
- Brady, J.P.; Garland, D.; Duglas-Tabor, Y.; Robison, W.G.; Groome, A.; Wawrousek, E.F. Targeted disruption of the mouse αA-crystallin gene induces cataract and cytoplasmic inclusion bodies containing the small heat shock protein αB-crystallin. Proc. Natl. Acad. Sci. USA 1997, 94, 884–889. [Google Scholar] [CrossRef] [PubMed]
- Brady, J.P.; Garland, D.L.; Green, D.E.; Tamm, E.R.; Giblin, F.J.; Wawrousek, E.F. AlphaB-crystallin in lens development and muscle integrity: A gene knockout approach. Invest. Ophthalmol. Vis. Sci. 2001, 42, 2924–2934. [Google Scholar]
- Reiter, C.E.; Wu, X.; Sandirasegarane, L.; Nakamura, M.; Gilbert, K.A.; Singh, R.S.; Fort, P.E.; Antonetti, D.A.; Gardner, T.W. Diabetes reduces basal retinal insulin receptor signaling: Reversal with systemic and local insulin. Diabetes 2006, 55, 1148–1156. [Google Scholar] [CrossRef]
- Losiewicz, M.K.; Fort, P.E. Diabetes impairs the neuroprotective properties of retinal alpha-crystallins. Investig. Opthalmol. Vis. Sci. 2011, 52, 5034–5042. [Google Scholar] [CrossRef]
- Barber, A.J.; Lieth, E.; Khin, S.A.; Antonetti, D.A.; Buchanan, A.G.; Gardner, T.W. Neural apoptosis in the retina during experimental and human diabetes. Early onset and effect of insulin. J. Clin. Investig. 1998, 102, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Yaung, J.; Kannan, R.; Wawrousek, E.F.; Spee, C.; Sreekumar, P.G.; Hinton, D.R. Exacerbation of retinal degeneration in the absence of alpha crystallins in an in vivo model of chemically induced hypoxia. Exp. Eye Res. 2007, 86, 355–365. [Google Scholar] [CrossRef]
- Johnson, P.T.; Brown, M.N.; Pulliam, B.C.; Anderson, D.H.; Johnson, L.V. Synaptic pathology, altered gene expression, and degeneration in photoreceptors impacted by drusen. Investig. Opthalmol. Vis. Sci. 2005, 46, 4788–4795. [Google Scholar] [CrossRef]
- Nakata, K.; Crabb, W.J.; Hollyfield, J.G. Crystallin distribution in Bruch’s membrane-choroid complex from AMD and age-matched donor eyes. Exp. Eye Res. 2005, 80, 821–826. [Google Scholar] [CrossRef]
- Kase, S.; Meghpara, B.B.; Ishida, S.; Rao, N.A. Expression of α-crystallin in the retina of human sympathetic ophthalmia. Mol. Med. Rep. 2011, 5, 395–399. [Google Scholar] [CrossRef] [PubMed]
- Saraswathy, S.; Nguyen, A.M.; Rao, N.A. The role of TLR4 in photoreceptor {alpha}a crystallin upregulation during early experimental autoimmune uveitis. Invest. Ophthalmol. Vis. Sci. 2010, 51, 3680–3686. [Google Scholar] [CrossRef]
- Hamann, S.; Métrailler, S.; Schorderet, D.F.; Cottet, S. Analysis of the cytoprotective role of alpha-Crystallins in cell survival and implication of the alpha A-crystallin c-terminal extension domain in preventing Bax-induced apoptosis. PLoS ONE 2013, 8, e55372. [Google Scholar] [CrossRef]
- Hoover, H.E.; Thuerauf, D.J.; Martindale, J.J.; Glembotski, C.C. alpha B-crystallin gene induction and phosphorylation by MKK6-activated p38. A potential role for alpha B-crystallin as a target of the p38 branch of the cardiac stress response. J. Biol. Chem. 2000, 275, 23825–23833. [Google Scholar] [CrossRef] [PubMed]
- Kamei, A.; Iwase, H.; Masuda, K. Cleavage of amino acid residue(s) from the N-terminal region of alpha A- and alpha B-Crystallins in human crystalline lens during aging. Biochem. Biophys. Res. Commun. 1997, 231, 373–378. [Google Scholar] [CrossRef]
- Kannan, R.; Sreekumar, P.G.; Hinton, D.R. Novel roles for α-crystallins in retinal function and disease. Prog. Retin. Eye Res. 2012, 31, 576–604. [Google Scholar] [CrossRef]
- Li, R.; Zhu, Z.; Reiser, G. Specific phosphorylation of αA-crystallin is required for the αA-crystallin-induced protection of astrocytes against staurosporine and C2-ceramide toxicity. Neurochem. Int. 2012, 60, 652–658. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Besirli, C.G.; Nath, M.; Yao, J.; Pawar, M.; Myers, A.M.; Zacks, D.; Fort, P.E. HSPB4/CRYAA Protect Photoreceptors during Retinal Detachment in Part through FAIM2 Regulation. Neurol. Int. 2024, 16, 905-917. https://doi.org/10.3390/neurolint16050068
Besirli CG, Nath M, Yao J, Pawar M, Myers AM, Zacks D, Fort PE. HSPB4/CRYAA Protect Photoreceptors during Retinal Detachment in Part through FAIM2 Regulation. Neurology International. 2024; 16(5):905-917. https://doi.org/10.3390/neurolint16050068
Chicago/Turabian StyleBesirli, Cagri G., Madhu Nath, Jingyu Yao, Mercy Pawar, Angela M. Myers, David Zacks, and Patrice E. Fort. 2024. "HSPB4/CRYAA Protect Photoreceptors during Retinal Detachment in Part through FAIM2 Regulation" Neurology International 16, no. 5: 905-917. https://doi.org/10.3390/neurolint16050068