
Narrative Review Explaining the Role of HLA-A, -B, and -C Molecules in COVID-19 Disease in and around Africa


Abstract
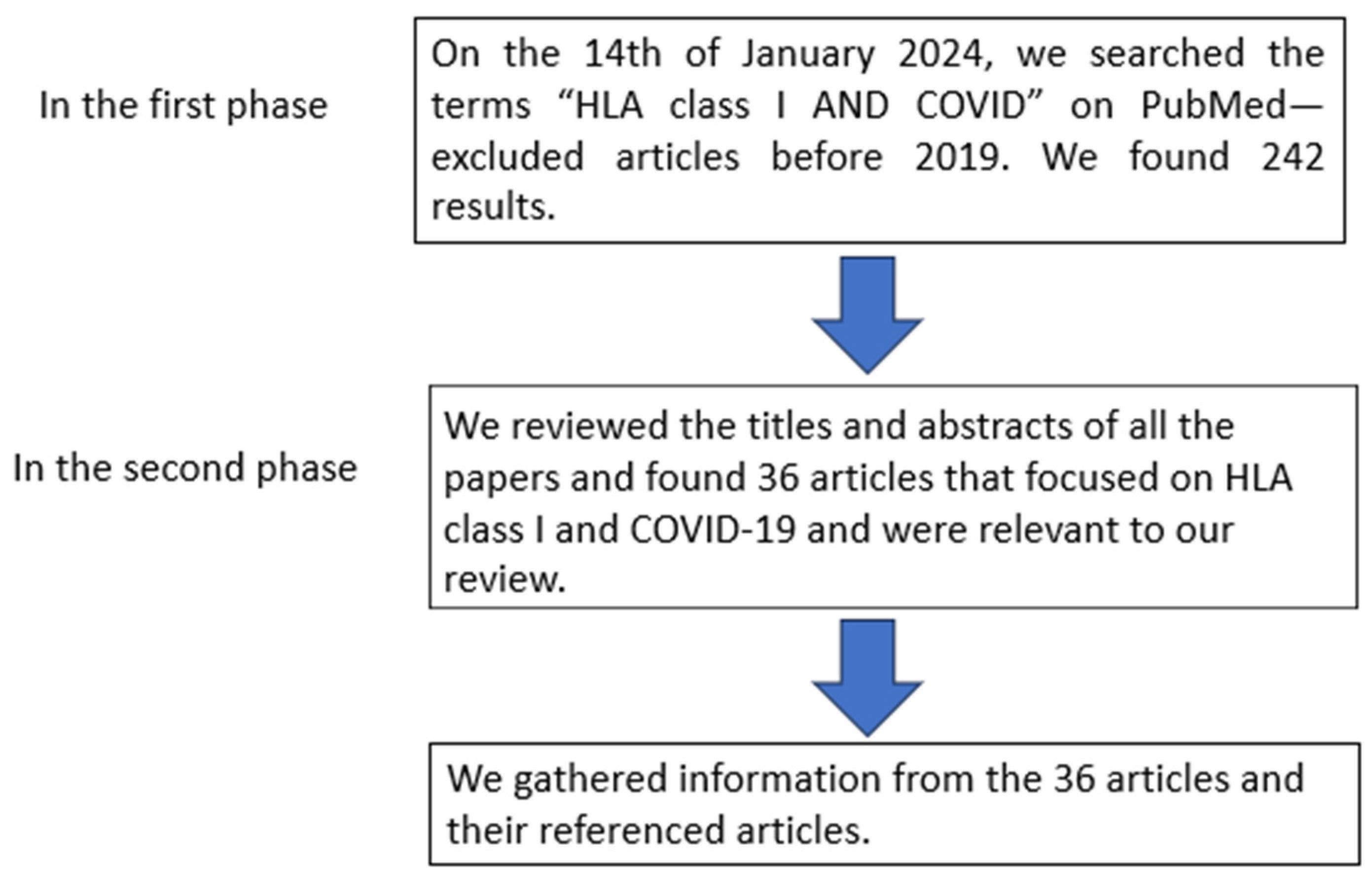
:1. Introduction
2. Selection Criteria
3. The Role of HLA in COVID-19
3.1. HLA-A
3.2. HLA-B
3.3. HLA-C
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lamers, M.M.; Haagmans, B.L. SARS-CoV-2 pathogenesis. Nat. Rev. Microbiol. 2022, 20, 270–284. [Google Scholar] [CrossRef]
- Louis, T.J.; Qasem, A.; Abdelli, L.S.; Naser, S.A. Extra-Pulmonary Complications in SARS-CoV-2 Infection: A Comprehensive Multi Organ-System Review. Microorganisms 2022, 10, 153. [Google Scholar] [CrossRef] [PubMed]
- Zietz, M.; Zucker, J.; Tatonetti, N. Associations between blood type and COVID-19 infection, intubation, and death. Nat. Commun. 2020, 11, 5761. [Google Scholar] [CrossRef] [PubMed]
- Ellinghaus, D.; Degenhardt, F.; Bujanda, L.; Buti, M. Genomewide association study of severe COVID-19 with respiratory failure. N. Engl. J. Med. 2020, 383, 1522–1534. [Google Scholar]
- Pairo-Castineira, E.; Clohisey, S.; Klaric, L.; Bretherick, A.D.; Rawlik, K.; Pasko, D.; Walker, S.; Parkinson, N.; Fourman, M.H.; Russell, C.D. Genetic mechanisms of critical illness in COVID-19. Nature 2021, 591, 92–98. [Google Scholar] [CrossRef]
- Cooke, G.S.; Hill, A.V. Genetics of susceptibility to human infectious disease. Nat. Rev. Genet. 2001, 2, 967–977. [Google Scholar] [CrossRef] [PubMed]
- Tufan, A.; Avanoğlu Güler, A.; Matucci-Cerinic, M. COVID-19, immune system response, hyperinflammation and repurposing antirheumatic drugs. Turk. J. Med. Sci. 2020, 50, 620–632. [Google Scholar] [CrossRef]
- Du, F.; Liu, B.; Zhang, S. COVID-19: The role of excessive cytokine release and potential ACE2 down-regulation in promoting hypercoagulable state associated with severe illness. J. Thromb. Thrombolysis 2021, 51, 313–329. [Google Scholar] [CrossRef]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef]
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R. A novel coronavirus from patients with pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef]
- Holshue, M.L.; DeBolt, C.; Lindquist, S.; Lofy, K.H.; Wiesman, J.; Bruce, H.; Spitters, C.; Ericson, K.; Wilkerson, S.; Tural, A. First case of 2019 novel coronavirus in the United States. New Engl. J. Med. 2020, 382, 929–936. [Google Scholar] [CrossRef]
- Wu, Z.; McGoogan, J.M. Characteristics of and important lessons from the coronavirus disease 2019 (COVID-19) outbreak in China: Summary of a report of 72,314 cases from the Chinese Center for Disease Control and Prevention. J. Am. Med. Assoc. 2020, 323, 1239–1242. [Google Scholar] [CrossRef] [PubMed]
- Osei, S.A.; Biney, R.P.; Anning, A.S.; Nortey, L.N.; Ghartey-Kwansah, G. Low incidence of COVID-19 case severity and mortality in Africa; Could malaria co-infection provide the missing link? BMC Infect. Dis. 2022, 22, 78. [Google Scholar] [CrossRef]
- Wamai, R.G.; Hirsch, J.L.; Van Damme, W.; Alnwick, D.; Bailey, R.C.; Hodgins, S.; Alam, U.; Anyona, M. What Could Explain the Lower COVID-19 Burden in Africa despite Considerable Circulation of the SARS-CoV-2 Virus? Int. J. Environ. Res. Public Health 2021, 18, 8638. [Google Scholar] [CrossRef]
- Soy, A. Coronavirus in Africa: Five Reasons Why COVID-19 Has Been Less Deadly than Elsewhere. Available online: https://dailytrust.com/coronavirus-in-africa-five-reasons-why-covid-19-has-been-less-deadly-than-elsewhere/ (accessed on 3 June 2023).
- Mennechet, F.J.D.; Dzomo, G.R.T. Coping with COVID-19 in Sub-Saharan Africa: What Might the Future Hold? Virol. Sin. 2020, 35, 875–884. [Google Scholar] [CrossRef] [PubMed]
- Adams, J.; MacKenzie, M.J.; Amegah, A.K.; Ezeh, A.; Gadanya, M.A.; Omigbodun, A.; Sarki, A.M.; Thistle, P.; Ziraba, A.K.; Stranges, S.; et al. The Conundrum of Low COVID-19 Mortality Burden in sub-Saharan Africa: Myth or Reality? Glob. Health Sci. Pract. 2021, 9, 433–443. [Google Scholar] [CrossRef] [PubMed]
- Blackwell, J.M.; Jamieson, S.E.; Burgner, D. HLA and infectious diseases. Clin. Microbiol. Rev. 2009, 22, 370–385. [Google Scholar] [CrossRef]
- Guan, W.-J.; Ni, Z.-Y.; Hu, Y.; Liang, W.-H.; Ou, C.-Q.; He, J.-X.; Liu, L.; Shan, H.; Lei, C.-L.; Hui, D.S. Clinical characteristics of coronavirus disease 2019 in China. New Engl. J. Med. 2020, 382, 1708–1720. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Tapias, P.; Castiblanco, J.; Anaya, J.-M. Major histocompatibility complex: Antigen processing and presentation. In Autoimmunity: From Bench to Bedside [Internet; El Rosario University Press: Bogota, Colombia, 2013. [Google Scholar]
- Srivastava, A.; Hollenbach, J.A. The immunogenetics of COVID-19. Immunogenetics 2023, 75, 309–320. [Google Scholar] [CrossRef]
- Tumer, G.; Simpson, B.; Roberts, T.K. Genetics, Human Major Histocompatibility Complex (MHC); StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Francis, J.M.; Leistritz-Edwards, D.; Dunn, A.; Tarr, C.; Lehman, J.; Dempsey, C.; Hamel, A.; Rayon, V.; Liu, G.; Wang, Y.; et al. Allelic variation in class I HLA determines CD8(+) T cell repertoire shape and cross-reactive memory responses to SARS-CoV-2. Sci. Immunol. 2022, 7, eabk3070. [Google Scholar]
- Wosen, J.E.; Mukhopadhyay, D.; Macaubas, C.; Mellins, E.D. Epithelial MHC class II expression and its role in antigen presentation in the gastrointestinal and respiratory tracts. Front. Immunol. 2018, 9, 2144. [Google Scholar] [CrossRef]
- Shiina, T.; Hosomichi, K.; Inoko, H.; Kulski, J.K. The HLA genomic loci map: Expression, interaction, diversity and disease. J. Hum. Genet. 2009, 54, 15–39. [Google Scholar] [CrossRef]
- Robinson, J.; Halliwell, J.A.; Hayhurst, J.D.; Flicek, P.; Parham, P.; Marsh, S.G. The IPD and IMGT/HLA database: Allele variant databases. Nucleic Acids Res. 2015, 43, D423–D431. [Google Scholar] [CrossRef]
- Rigden, D.J.; Fernández, X.M. The 2023 Nucleic Acids Research Database Issue and the online molecular biology database collection. Nucleic Acids Res. 2023, 51, D1–D8. [Google Scholar] [CrossRef] [PubMed]
- van Deutekom, H.W.; Keşmir, C. Zooming into the binding groove of HLA molecules: Which positions and which substitutions change peptide binding most? Immunogenetics 2015, 67, 425–436. [Google Scholar] [CrossRef] [PubMed]
- Lenz, T.L.; Spirin, V.; Jordan, D.M.; Sunyaev, S.R. Excess of deleterious mutations around HLA genes reveals evolutionary cost of balancing selection. Mol. Biol. Evol. 2016, 33, 2555–2564. [Google Scholar] [CrossRef]
- Dutta, M.; Dutta, P.; Medhi, S.; Borkakoty, B.; Biswas, D. Polymorphism of HLA class I and class II alleles in influenza A (H1N1) pdm09 virus infected population of Assam, Northeast India. J. Med. Virol. 2018, 90, 854–860. [Google Scholar] [CrossRef]
- Ma, Y.; Yuan, B.; Yi, J.; Zhuang, R.; Wang, J.; Zhang, Y.; Xu, Z.; Zhang, Y.; Liu, B.; Wei, C. The genetic polymorphisms of HLA are strongly correlated with the disease severity after Hantaan virus infection in the Chinese Han population. Clin. Dev. Immunol. 2012, 2012, 308237. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.; Tseng, H.-K.; Trejaut, J.A.; Lee, H.-L.; Loo, J.-H.; Chu, C.-C.; Chen, P.-J.; Su, Y.-W.; Lim, K.H.; Tsai, Z.-U. Association of HLA class I with severe acute respiratory syndrome coronavirus infection. BMC Med. Genet. 2003, 4, 9. [Google Scholar] [CrossRef]
- Saito, H.; Umemura, T.; Joshita, S.; Yamazaki, T.; Fujimori, N.; Kimura, T.; Komatsu, M.; Matsumoto, A.; Tanaka, E.; Ota, M. KIR2DL2 combined with HLA-C1 confers risk of hepatitis C virus-related hepatocellular carcinoma in younger patients. Oncotarget 2018, 9, 19650. [Google Scholar] [CrossRef]
- Knoring, B.; Berkos, A.; IIa, S. Distribution of histocompatibility antigens in patients with pulmonary tuberculosis depending on disease course and immune response pattern. Probl. Tuberk. 1995, 2, 16–19. [Google Scholar]
- Hernández-Doño, S.; Sánchez-González, R.A.; Trujillo-Vizuet, M.G.; Zamudio-Castellanos, F.Y.; García-Silva, R.; Bulos-Rodríguez, P.; Vazquez-Guzmán, C.A.; Cárdenas-Ramos, X.; de León Rodríguez, D.; Elías, F. Protective HLA alleles against severe COVID-19: HLA-A* 68 as an ancestral protection allele in Tapachula-Chiapas, Mexico. Clin. Immunol. 2022, 238, 108990. [Google Scholar] [CrossRef] [PubMed]
- Lorente, L.; Martín, M.; Franco, A.; Barrios, Y.; Cáceres, J.; Solé-Violán, J.; Perez, A.; Marcos, J.; Ramos-Gómez, L.; Ojeda, N. HLA genetic polymorphisms and prognosis of patients with COVID-19. Med. Intensiv. 2021, 45, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Tripathy, A.S.; Wagh, P.; Vishwakarma, S.; Akolkar, K.; Tripathy, S.; Jali, P.; Kakrani, A.L.; Barthwal, M.; Gurav, Y.; Kadgi, N.; et al. Association of human leukocyte antigen class I and class II alleles and haplotypes in COVID-19 infection in a western Indian population. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2023, 113, 105468. [Google Scholar] [CrossRef]
- Suslova, T.A.; Vavilov, M.N.; Belyaeva, S.V.; Evdokimov, A.V.; Stashkevich, D.S.; Galkin, A.; Kofiadi, I.A. Distribution of HLA-A, -B, -C, -DRB1, -DQB1, -DPB1 allele frequencies in patients with COVID-19 bilateral pneumonia in Russians, living in the Chelyabinsk region (Russia). Hum. Immunol. 2022, 83, 547–550. [Google Scholar] [CrossRef]
- Poulton, K.; Wright, P.; Hughes, P.; Savic, S.; Welberry Smith, M.; Guiver, M.; Morton, M.; van Dellen, D.; Tholouli, E.; Wynn, R. A role for human leucocyte antigens in the susceptibility to SARS-CoV-2 infection observed in transplant patients. Int. J. Immunogenet. 2020, 47, 324–328. [Google Scholar] [CrossRef]
- Schetelig, J.; Heidenreich, F.; Baldauf, H.; Trost, S.; Falk, B.; Hoßbach, C.; Real, R.; Roers, A.; Lindemann, D.; Dalpke, A.; et al. Individual HLA-A, -B, -C, and -DRB1 Genotypes Are No Major Factors Which Determine COVID-19 Severity. Front. Immunol. 2021, 12, 698193. [Google Scholar] [CrossRef]
- Kiyotani, K.; Toyoshima, Y.; Nemoto, K.; Nakamura, Y. Bioinformatic prediction of potential T cell epitopes for SARS-CoV-2. J. Hum. Genet. 2020, 65, 569–575. [Google Scholar] [CrossRef] [PubMed]
- Tomita, Y.; Ikeda, T.; Sato, R.; Sakagami, T. Association between HLA gene polymorphisms and mortality of COVID-19: An in silico analysis. Immun. Inflamm. Dis. 2020, 8, 684–694. [Google Scholar] [CrossRef]
- Basir, H.R.G.; Majzoobi, M.M.; Ebrahimi, S.; Noroozbeygi, M.; Hashemi, S.H.; Keramat, F.; Mamani, M.; Eini, P.; Alizadeh, S.; Solgi, G.; et al. Susceptibility and Severity of COVID-19 Are Both Associated With Lower Overall Viral-Peptide Binding Repertoire of HLA Class I Molecules, Especially in Younger People. Front. Immunol. 2022, 13, 891816. [Google Scholar] [CrossRef]
- Shkurnikov, M.; Nersisyan, S.; Jankevic, T.; Galatenko, A.; Gordeev, I.; Vechorko, V.; Tonevitsky, A. Association of HLA class I genotypes with severity of coronavirus disease-19. Front. Immunol. 2021, 12, 641900. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Huang, S.; Gao, R.; Zhou, Y.; Lai, C.; Li, Z. Initial whole-genome sequencing and analysis of the host genetic contribution to COVID-19 severity and susceptibility. Cell Discov. 2020, 6, 83. [Google Scholar] [CrossRef] [PubMed]
- Khor, S.S.; Omae, Y.; Nishida, N.; Sugiyama, M.; Kinoshita, N.; Suzuki, T.; Suzuki, M.; Suzuki, S.; Izumi, S.; Hojo, M.; et al. HLA-A*11:01:01:01, HLA-C*12:02:02:01-HLA-B*52:01:02:02, Age and Sex Are Associated With Severity of Japanese COVID-19 With Respiratory Failure. Front. Immunol. 2021, 12, 658570. [Google Scholar] [CrossRef] [PubMed]
- Warren, R.L.; Birol, I. Retrospective in silico HLA predictions from COVID-19 patients reveal alleles associated with disease prognosis. MedRxiv 2020. [Google Scholar] [CrossRef]
- Castro-Santos, P.; Rojas-Martinez, A.; Riancho, J.A.; Lapunzina, P.; Flores, C.; Carracedo, Á.; Díaz-Peña, R. HLA-A*11:01 and HLA-C*04:01 are associated with severe COVID-19. HLA 2023, 102, 731–739. [Google Scholar] [CrossRef] [PubMed]
- de Moura, R.R.; Agrelli, A.; Santos-Silva, C.A.; Silva, N.; Assunção, B.R.; Brandão, L.; Benko-Iseppon, A.M.; Crovella, S. Immunoinformatic approach to assess SARS-CoV-2 protein S epitopes recognised by the most frequent MHC-I alleles in the Brazilian population. J. Clin. Pathol. 2021, 74, 528–532. [Google Scholar] [CrossRef] [PubMed]
- Balas, A.; Moreno-Hidalgo, M.; de la Calle-Prieto, F.; Vicario, J.L.; Arsuaga, M.; Trigo, E.; de Miguel-Buckley, R.; Bellón, T.; Díaz-Menéndez, M. Coronavirus-19 disease risk and protective factors associated with HLA/KIR polymorphisms in Ecuadorian patients residing in Madrid. Hum. Immunol. 2023, 84, 571–577. [Google Scholar] [CrossRef]
- Warren, R.L.; Birol, I. HLA predictions from the bronchoalveolar lavage fluid samples of five patients at the early stage of the wuhan seafood market COVID-19 outbreak. arXiv 2020, arXiv:2004.07108v3. [Google Scholar]
- Schindler, E.; Dribus, M.; Duffy, B.F.; Hock, K.; Farnsworth, C.W.; Gragert, L.; Liu, C. HLA genetic polymorphism in patients with Coronavirus Disease 2019 in Midwestern United States. HLA 2021, 98, 370–379. [Google Scholar] [CrossRef]
- Farahani, R.H.; Esmaeilzadeh, E.; Asl, A.N.; Heidari, M.F.; Hazrati, E. Frequency of HLA Alleles in a Group of Severe COVID-19 Iranian Patients. Iran. J. Public Health 2021, 50, 1882. [Google Scholar]
- Sakuraba, A.; Haider, H.; Sato, T. Population difference in allele frequency of HLA-C* 05 and its correlation with COVID-19 mortality. Viruses 2020, 12, 1333. [Google Scholar] [CrossRef] [PubMed]
- Ng, M.H.; Lau, K.-M.; Li, L.; Cheng, S.-H.; Chan, W.Y.; Hui, P.K.; Zee, B.; Leung, C.-B.; Sung, J.J. Association of human-leukocyte-antigen class I (B* 0703) and class II (DRB1* 0301) genotypes with susceptibility and resistance to the development of severe acute respiratory syndrome. J. Infect. Dis. 2004, 190, 515–518. [Google Scholar] [CrossRef] [PubMed]
- Pisanti, S.; Deelen, J.; Gallina, A.M.; Caputo, M.; Citro, M.; Abate, M.; Sacchi, N.; Vecchione, C.; Martinelli, R. Correlation of the two most frequent HLA haplotypes in the Italian population to the differential regional incidence of COVID-19. J. Transl. Med. 2020, 18, 352. [Google Scholar] [CrossRef] [PubMed]
- Leite, M.M.; Gonzalez-Galarza, F.F.; Silva, B.; Middleton, D.; Santos, E. Predictive immunogenetic markers in COVID-19. Hum. Immunol. 2021, 82, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Correale, P.; Mutti, L.; Pentimalli, F.; Baglio, G.; Saladino, R.E.; Sileri, P.; Giordano, A. HLA-B* 44 and C* 01 prevalence correlates with Covid19 spreading across Italy. Int. J. Mol. Sci. 2020, 21, 5205. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhang, W.; Zhang, J.; He, J.; Zhu, F. Distribution of HLA allele frequencies in 82 Chinese individuals with coronavirus disease-2019 (COVID-19). HLA 2020, 96, 194–196. [Google Scholar] [CrossRef] [PubMed]
- Abdelhafiz, A.S.; Ali, A.; Fouda, M.A.; Sayed, D.M.; Kamel, M.M.; Kamal, L.M.; Khalil, M.A.; Bakry, R.M. HLA-B*15 predicts survival in Egyptian patients with COVID-19. Hum. Immunol. 2022, 83, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, A.; David, J.K.; Maden, S.K.; Wood, M.A.; Weeder, B.R.; Nellore, A.; Thompson, R.F. Human leukocyte antigen susceptibility map for severe acute respiratory syndrome coronavirus 2. J. Virol. 2020, 94, e00510–e00520. [Google Scholar] [CrossRef] [PubMed]
- Augusto, D.G.; Yusufali, T.; Peyser, N.D.; Butcher, X.; Marcus, G.M.; Olgin, J.E.; Pletcher, M.J.; Maiers, M.; Hollenbach, J.A. HLA-B* 15: 01 is associated with asymptomatic SARS-CoV-2 infection. MedRxiv 2021. [Google Scholar] [CrossRef]
- Bordon, Y. Asymptomatic SARS-CoV-2 linked to HLA-B*15:01. Nat. Rev. Immunol. 2023, 23, 543. [Google Scholar] [CrossRef]
- Yung, Y.L.; Cheng, C.K.; Chan, H.Y.; Xia, J.T.; Lau, K.M.; Wong, R.S.; Wu, A.K.; Chu, R.W.; Wong, A.C.; Chow, E.Y. Association of HLA-B22 serotype with SARS-CoV-2 susceptibility in Hong Kong Chinese patients. HLA 2021, 97, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Tay, G.K.; Alnaqbi, H.; Chehadeh, S.; Peramo, B.; Mustafa, F.; Rizvi, T.A.; Mahboub, B.H.; Uddin, M.; Alkaabi, N.; Alefishat, E.; et al. HLA class I associations with the severity of COVID-19 disease in the United Arab Emirates. PLoS ONE 2023, 18, e0285712. [Google Scholar] [CrossRef] [PubMed]
- Littera, R.; Campagna, M.; Deidda, S.; Angioni, G.; Cipri, S.; Melis, M.; Firinu, D.; Santus, S.; Lai, A.; Porcella, R.; et al. Human Leukocyte Antigen Complex and Other Immunogenetic and Clinical Factors Influence Susceptibility or Protection to SARS-CoV-2 Infection and Severity of the Disease Course. The Sardinian Experience. Front. Immunol. 2020, 11, 605688. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, A.; David, J.; Maden, S. Human leukocyte antigen susceptibility map for SARS-CoV-2. J. Virol. 2020, 510, 20. [Google Scholar]
- Gutiérrez-Bautista, J.F.; Rodriguez-Nicolas, A.; Rosales-Castillo, A.; López-Ruz, M.; Martín-Casares, A.M.; Fernández-Rubiales, A.; Anderson, P.; Garrido, F.; Ruiz-Cabello, F.; López-Nevot, M. Study of HLA-A, -B, -C, -DRB1 and -DQB1 polymorphisms in COVID-19 patients. J. Microbiol. Immunol. Infect. = Wei Mian Yu Gan Ran Za Zhi 2022, 55, 421–427. [Google Scholar] [CrossRef]
- Naemi, F.M.A.; Al-Adwani, S.; Al-Nazawi, A.; Al-Khatabi, H. Association between HLA genotype and ferritin levels in COVID-19 infection: A study of a Saudi cohort. Infect. Dis. 2021, 53, 891–899. [Google Scholar] [CrossRef] [PubMed]
- Barquera, R.; Collen, E.; Di, D.; Buhler, S.; Teixeira, J.; Llamas, B.; Nunes, J.M.; Sanchez-Mazas, A. Binding affinities of 438 HLA proteins to complete proteomes of seven pandemic viruses and distributions of strongest and weakest HLA peptide binders in populations worldwide. HLA 2020, 96, 277–298. [Google Scholar] [CrossRef] [PubMed]
- Toyoshima, Y.; Nemoto, K.; Matsumoto, S.; Nakamura, Y.; Kiyotani, K. SARS-CoV-2 genomic variations associated with mortality rate of COVID-19. J. Hum. Genet. 2020, 65, 1075–1082. [Google Scholar] [CrossRef] [PubMed]
- Novelli, A.; Andreani, M.; Biancolella, M.; Liberatoscioli, L.; Passarelli, C.; Colona, V.L.; Rogliani, P.; Leonardis, F.; Campana, A.; Carsetti, R. HLA allele frequencies and susceptibility to COVID-19 in a group of 99 Italian patients. HLA 2020, 96, 610–614. [Google Scholar] [CrossRef]
- Gonzalez-Galarza, F.F.; Christmas, S.; Middleton, D.; Jones, A.R. Allele frequency net: A database and online repository for immune gene frequencies in worldwide populations. Nucleic Acids Res. 2010, 39, D913–D919. [Google Scholar] [CrossRef]
- Cacho-Díaz, B.; García-Botello, D.R.; Wegman-Ostrosky, T.; Reyes-Soto, G.; Ortiz-Sánchez, E.; Herrera-Montalvo, L.A. Tumor microenvironment differences between primary tumor and brain metastases. J. Transl. Med. 2020, 18, 1. [Google Scholar] [CrossRef] [PubMed]
- Weiner, J.; Suwalski, P.; Holtgrewe, M.; Rakitko, A.; Thibeault, C.; Müller, M. Increased risk of severe clinical course of COVID-19 in carriers of HLA-C* 04: 01. EClinicalMedicine 2021, 40, 101099. [Google Scholar] [CrossRef]
- Saulle, I.; Vicentini, C.; Clerici, M.; Biasin, M. Antigen presentation in SARS-CoV-2 infection: The role of class I HLA and ERAP polymorphisms. Hum. Immunol. 2021, 82, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Vigón, L.; Galán, M.; Torres, M.; Martín-Galiano, A.J.; Rodríguez-Mora, S.; Mateos, E.; Corona, M.; Malo, R.; Navarro, C.; Murciano-Antón, M.A.; et al. Association between HLA-C alleles and COVID-19 severity in a pilot study with a Spanish Mediterranean Caucasian cohort. PLoS ONE 2022, 17, e0272867. [Google Scholar] [CrossRef] [PubMed]
- Roncati, L.; Palmieri, B. What about the original antigenic sin of the humans versus SARS-CoV-2? Med. Hypotheses 2020, 142, 109824. [Google Scholar] [CrossRef]
- Tavasolian, F.; Rashidi, M.; Hatam, G.R.; Jeddi, M.; Hosseini, A.Z.; Mosawi, S.H.; Abdollahi, E.; Inman, R.D. HLA, immune response, and susceptibility to COVID-19. Front. Immunol. 2021, 11, 601886. [Google Scholar] [CrossRef]
- Shi, Y.; Wang, Y.; Shao, C.; Huang, J.; Gan, J.; Huang, X.; Bucci, E.; Piacentini, M.; Ippolito, G.; Melino, G. COVID-19 Infection: The Perspectives on Immune Responses; Nature Publishing Group: New York, NY, USA, 2020; Volume 27, pp. 1451–1454. [Google Scholar]
- Augusto, D.G.; Hollenbach, J.A. HLA variation and antigen presentation in COVID-19 and SARS-CoV-2 infection. Curr. Opin. Immunol. 2022, 76, 102178. [Google Scholar] [CrossRef]
- Chen, L.-C.; Nersisyan, S.; Wu, C.-J.; Chang, C.-M.; Tonevitsky, A.; Guo, C.-L.; Chang, W.-C. On the peptide binding affinity changes in population-specific HLA repertoires to the SARS-CoV-2 variants Delta and Omicron. J. Autoimmun. 2022, 133, 102952. [Google Scholar] [CrossRef]
- Albayrak, A.; Ertek, M.; Tasyaran, M.A.; Pirim, I. Role of HLA Allele Polymorphism in Chronic Hepatitis B Virus Infection and HBV Vaccine Sensitivity in Patients from Eastern Turkey. Biochemical Genetics. 2011, 49, 258–269. [Google Scholar] [CrossRef]
- Ben Shachar, S.; Barda, N.; Manor, S.; Israeli, S.; Dagan, N.; Carmi, S.; Balicer, R.; Zisser, B.; Louzoun, Y. MHC haplotyping of SARS-CoV-2 patients: HLA subtypes are not associated with the presence and severity of COVID-19 in the Israeli population. J. Clin. Immunol. 2021, 41, 1154–1161. [Google Scholar] [CrossRef]
- Fakhkhari, M.; Caidi, H.; Sadki, K. HLA alleles associated with COVID-19 susceptibility and severity in different populations: A systematic review. Egypt. J. Med. Hum. Genet. 2023, 24, 10. [Google Scholar] [CrossRef] [PubMed]
- Gill, C.J.; Mwananyanda, L.; MacLeod, W.; Kwenda, G.; Pieciak, R.; Etter, L.; Bridges, D.; Chikoti, C.; Chirwa, S.; Chimoga, C. Sustained high prevalence of COVID-19 deaths from a systematic post-mortem study in Lusaka, Zambia: One year later. medRxiv 2022. [Google Scholar] [CrossRef]
- Ishii, T. Human Leukocyte Antigen (HLA) Class I Susceptible Alleles Against COVID-19 Increase Both Infection and Severity Rate. Cureus 2020, 12, e12239. [Google Scholar] [CrossRef] [PubMed]
- González-Galarza, F.F.; Takeshita, L.Y.; Santos, E.J.; Kempson, F.; Maia, M.H.T.; Silva, A.L.S.d.; Silva, A.L.T.e.; Ghattaoraya, G.S.; Alfirevic, A.; Jones, A.R. Allele frequency net 2015 update: New features for HLA epitopes, KIR and disease and HLA adverse drug reaction associations. Nucleic Acids Res. 2015, 43, D784–D788. [Google Scholar] [CrossRef] [PubMed]
- Cheranev, V.; Bulusheva, I.; Vechorko, V.; Korostin, D.; Rebrikov, D. The Search of Association of HLA Class I and Class II Alleles with COVID-19 Mortality in the Russian Cohort. Int. J. Mol. Sci. 2023, 24, 3068. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Galarza, F.F.; McCabe, A.; Melo dos Santos, E.J.; Takeshita, L.; Ghattaoraya, G.; Jones, A.R.; Middleton, D. Allele frequency net database. In HLA Typing; Springer: Berlin/Heidelberg, Germany, 2018; pp. 49–62. [Google Scholar]
- Deb, P.; Molla, M.M.A.; Saif-Ur-Rahman, K.M. An update to monoclonal antibody as therapeutic option against COVID-19. Biosaf. Health 2021, 3, 87–91. [Google Scholar] [CrossRef]
- Yung, Y.-L.; Cheng, C.-K.; Chan, H.-Y.; Xia, J.T.; Lau, K.-M.; Wong, R.S.M.; Wu, A.K.L.; Chu, R.W.; Wong, A.C.C.; Chow, E.Y.D.; et al. Association of HLA-B22 serotype with SARS-CoV-2 susceptibility in Hong Kong Chinese patients. HLA 2021, 97, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Migliorini, F.; Torsiello, E.; Spiezia, F.; Oliva, F.; Tingart, M.; Maffulli, N. Association between HLA genotypes and COVID-19 susceptibility, severity and progression: A comprehensive review of the literature. Eur. J. Med. Res. 2021, 26, 84. [Google Scholar] [CrossRef] [PubMed]
- Ueta, M.; Kannabiran, C.; Wakamatsu, T.H.; Kim, M.K.; Yoon, K.-C.; Seo, K.Y.; Joo, C.-K.; Sangwan, V.; Rathi, V.; Basu, S. Trans-ethnic study confirmed independent associations of HLA-A* 02: 06 and HLA-B* 44: 03 with cold medicine-related Stevens-Johnson syndrome with severe ocular surface complications. Sci. Rep. 2014, 4, 5981. [Google Scholar] [CrossRef]
- Grams, S.; Moonsamy, P.; Mano, C.; Oksenberg, J.; Begovich, A. Two new HLA-B alleles, B* 4422 and B* 4704, identified in a study of families with autoimmunity. Tissue Antigens 2002, 59, 338–340. [Google Scholar] [CrossRef]
- Johnston, D.T.; Mehaffey, G.; Thomas, J.; Young Jr, K.R.; Wiener, H.; Li, J.; Go, R.C.; Schroeder Jr, H.W. Increased frequency of HLA-B44 in recurrent sinopulmonary infections (RESPI). Clin. Immunol. 2006, 119, 346–350. [Google Scholar] [CrossRef] [PubMed]
- Deb, P.; Zannat, K.e.; Talukder, S.; Bhuiyan, A.H.; Jilani, M.S.A.; Saif-Ur-Rahman, K. Association of HLA gene polymorphism with susceptibility, severity, and mortality of COVID-19: A systematic review. HLA 2022, 99, 281–312. [Google Scholar] [CrossRef] [PubMed]
- Tuttolomondo, A.; Di Raimondo, D.; Pecoraro, R.; Casuccio, A.; Di Bona, D.; Aiello, A.; Accardi, G.; Arnao, V.; Clemente, G.; Corte, V.D. HLA and killer cell immunoglobulin-like receptor (KIRs) genotyping in patients with acute ischemic stroke. J. Neuroinflammation 2019, 16, 88. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Shen, J.; Cox, C.; Wakefield, J.C.; Ehm, M.G.; Nelson, M.R.; Weir, B.S. HIBAG—HLA genotype imputation with attribute bagging. Pharmacogenomics J. 2014, 14, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Roberto, L.; Campagna, M.; Silvia, D.; Goffredo, A.; Selene, C.; Melis, M.; Firinu, D.; Simonetta, S.; Alberto, L.; Rita, P. Human Leukocyte Antigen Complex and Other Immunogenetic and Clinical Factors Influence Susceptibility or Protection to SARS-CoV-2 Infection and Severity of the Disease Course. The Sardinian Experience. Front. Immunol. 2020, 11, 605688. [Google Scholar]
- Amoroso, A.; Magistroni, P.; Vespasiano, F.; Bella, A.; Bellino, S.; Puoti, F.; Alizzi, S.; Vaisitti, T.; Boros, S.; Grossi, P.A. HLA and AB0 polymorphisms may influence SARS-CoV-2 infection and COVID-19 severity. Transplantation 2021, 105, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Iturrieta-Zuazo, I.; Rita, C.G.; García-Soidán, A.; de Malet Pintos-Fonseca, A.; Alonso-Alarcón, N.; Pariente-Rodríguez, R.; Tejeda-Velarde, A.; Serrano-Villar, S.; Castañer-Alabau, J.L.; Nieto-Gañán, I. Possible role of HLA class-I genotype in SARS-CoV-2 infection and progression: A pilot study in a cohort of COVID-19 Spanish patients. Clin. Immunol. 2020, 219, 108572. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.-B.; Xu, C.; Zhang, R.-B.; Wu, M.; Pan, C.-K.; Li, X.-J.; Wang, Q.; Zeng, F.-F.; Zhu, S. Associations of procalcitonin, C-reaction protein and neutrophil-to-lymphocyte ratio with mortality in hospitalized COVID-19 patients in China. Sci. Rep. 2020, 10, 15058. [Google Scholar] [CrossRef] [PubMed]
- Mueller, A.A.; Tamura, T.; Crowley, C.P.; DeGrado, J.R.; Haider, H.; Jezmir, J.L.; Keras, G.; Penn, E.H.; Massaro, A.F.; Kim, E.Y. Inflammatory biomarker trends predict respiratory decline in COVID-19 patients. Cell Rep. Med. 2020, 1, 100144. [Google Scholar] [CrossRef]
- Balnis, J.; Madrid, A.; Hogan, K.J.; Drake, L.A.; Chieng, H.C.; Tiwari, A.; Vincent, C.E.; Chopra, A.; Vincent, P.A.; Robek, M.D. Blood DNA methylation and COVID-19 outcomes. Clin. Epigenetics 2021, 13, 118. [Google Scholar] [CrossRef]
- Olvera, A.; Pérez-Álvarez, S.; Ibarrondo, J.; Ganoza, C.; Lama, J.R.; Lucchetti, A.; Cate, S.; Hildebrand, W.; Bernard, N.; Gomez, L. The HLA-C* 04: 01: KIR2DS4: Gene combination and human leukocyte antigen alleles with high population frequency drive rate of HIV disease progression. Aids 2015, 29, 507–517. [Google Scholar] [CrossRef] [PubMed]
- Cowling, B.J.; Chan, K.H.; Fang, V.J.; Lau, L.L.; So, H.C.; Fung, R.O.; Ma, E.S.; Kwong, A.S.; Chan, C.-W.; Tsui, W.W. Comparative epidemiology of pandemic and seasonal influenza A in households. N. Engl. J. Med. 2010, 362, 2175–2184. [Google Scholar] [CrossRef] [PubMed]
- Peiris, J.S.M.; Chu, C.-M.; Cheng, V.C.-C.; Chan, K.; Hung, I.; Poon, L.L.; Law, K.-I.; Tang, B.; Hon, T.; Chan, C. Clinical progression and viral load in a community outbreak of coronavirus-associated SARS pneumonia: A prospective study. Lancet 2003, 361, 1767–1772. [Google Scholar] [CrossRef] [PubMed]
- Jones, T.C.; Biele, G.; Mühlemann, B.; Veith, T.; Schneider, J.; Beheim-Schwarzbach, J.; Bleicker, T.; Tesch, J.; Schmidt, M.L.; Sander, L.E. Estimating infectiousness throughout SARS-CoV-2 infection course. Science 2021, 373, eabi5273. [Google Scholar] [CrossRef] [PubMed]
- Zou, L.; Ruan, F.; Huang, M.; Liang, L.; Huang, H.; Hong, Z.; Yu, J.; Kang, M.; Song, Y.; Xia, J. SARS-CoV-2 viral load in upper respiratory specimens of infected patients. N. Engl. J. Med. 2020, 382, 1177–1179. [Google Scholar] [CrossRef] [PubMed]
- Tindale, L.C.; Stockdale, J.E.; Coombe, M.; Garlock, E.S.; Lau, W.Y.V.; Saraswat, M.; Zhang, L.; Chen, D.; Wallinga, J.; Colijn, C. Evidence for transmission of COVID-19 prior to symptom onset. Elife 2020, 9, e57149. [Google Scholar] [CrossRef] [PubMed]
- The COVID-19 Host Genetics Initiative; Ganna, A. Mapping the human genetic architecture of COVID-19 by worldwide meta-analysis. medRxiv 2021. [Google Scholar] [CrossRef]
- Hovhannisyan, A.; Madelian, V.; Avagyan, S.; Nazaretyan, M.; Hyussyan, A.; Sirunyan, A.; Arakelyan, R.; Manukyan, Z.; Yepiskoposyan, L.; Mayilyan, K.R.; et al. HLA-C*04:01 Affects HLA Class I Heterozygosity and Predicted Affinity to SARS-CoV-2 Peptides, and in Combination With Age and Sex of Armenian Patients Contributes to COVID-19 Severity. Front Immunol 2022, 13, 769900. [Google Scholar] [CrossRef] [PubMed]
- Johansson, T.; Partanen, J.; Saavalainen, P. HLA allele-specific expression: Methods, disease associations, and relevance in hematopoietic stem cell transplantation. Front. Immunol. 2022, 13, 1007425. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Luo, W.; Szatmary, P.; Zhang, X.; Lin, J.W.; Chen, L.; Liu, D.; Sutton, R.; Xia, Q.; Jin, T.; et al. Monocytic HLA-DR Expression in Immune Responses of Acute Pancreatitis and COVID-19. Int. J. Mol. Sci. 2023, 24, 3246. [Google Scholar] [CrossRef]
- Bouayad, A. Features of HLA class I expression and its clinical relevance in SARS-CoV-2: What do we know so far? Rev. Med. Virol. 2021, 31, e2236. [Google Scholar] [CrossRef]
- Asmaa, N.; Alaa, R.; Nafady-Hego, H.; Nasif, K.A.; Ebtessam, E.; Elrahman, N.A.; Osman, H.A. Lymphocyte Phenotyping and HLA-DR Expression over the Course of COVID-19 Infection in Patients with Different Disease Severity. Clin. Lab. 2022, 68, 2383. [Google Scholar] [CrossRef]
- Arab, F.; Mollazadeh, S.; Ghayourbabaei, F.; Moghbeli, M.; Saburi, E. The role of HLA genotypes in understanding the pathogenesis of severe COVID-19. Egypt. J. Med. Hum. Genet. 2023, 24, 14. [Google Scholar] [CrossRef]
- Nersisyan, S.; Zhiyanov, A.; Zakharova, M.; Ishina, I.; Kurbatskaia, I.; Mamedov, A.; Galatenko, A.; Shkurnikov, M.; Gabibov, A.; Tonevitsky, A. Alterations in SARS-CoV-2 Omicron and Delta peptides presentation by HLA molecules. PeerJ 2022, 10, e13354. [Google Scholar] [CrossRef] [PubMed]
- Reynisson, B.; Alvarez, B.; Paul, S.; Peters, B.; Nielsen, M. NetMHCpan-4.1 and NetMHCIIpan-4.0: Improved predictions of MHC antigen presentation by concurrent motif deconvolution and integration of MS MHC eluted ligand data. Nucleic Acids Res. 2020, 48, W449–W454. [Google Scholar] [CrossRef] [PubMed]
- Duong, B.V.; Larpruenrudee, P.; Fang, T.; Hossain, S.I.; Saha, S.C.; Gu, Y.; Islam, M.S. Is the SARS-CoV-2 Omicron Variant Deadlier and More Transmissible Than Delta Variant? Int. J. Environ. Res. Public Health 2022, 19, 4586. [Google Scholar] [CrossRef]
- Araf, Y.; Akter, F.; Tang, Y.D.; Fatemi, R.; Parvez, M.S.A.; Zheng, C.; Hossain, M.G. Omicron variant of SARS-CoV-2: Genomics, transmissibility, and responses to current COVID-19 vaccines. J. Med. Virol. 2022, 94, 1825–1832. [Google Scholar] [CrossRef] [PubMed]
- Prugnolle, F.; Manica, A.; Charpentier, M.; Guégan, J.F.; Guernier, V.; Balloux, F. Pathogen-driven selection and worldwide HLA class I diversity. Curr. Biol. CB 2005, 15, 1022–1027. [Google Scholar] [CrossRef]
- Tshabalala, M.; Mellet, J.; Pepper, M.S. Human Leukocyte Antigen Diversity: A Southern African Perspective. J. Immunol. Res. 2015, 2015, 746151. [Google Scholar] [CrossRef] [PubMed]
- Miller, L.H. Impact of malaria on genetic polymorphism and genetic diseases in Africans and African Americans. Proc. Natl. Acad. Sci. USA 1994, 91, 2415–2419. [Google Scholar] [CrossRef]
- Apps, R.; Qi, Y.; Carlson, J.M.; Chen, H.; Gao, X.; Thomas, R.; Yuki, Y.; Del Prete, G.Q.; Goulder, P.; Brumme, Z.L. Influence of HLA-C expression level on HIV control. Science 2013, 340, 87–91. [Google Scholar] [CrossRef]
- Payne, R.P.; Branch, S.; Kløverpris, H.; Matthews, P.C.; Koofhethile, C.K.; Strong, T.; Adland, E.; Leitman, E.; Frater, J.; Ndung’u, T.; et al. Differential escape patterns within the dominant HLA-B*57:03-restricted HIV Gag epitope reflect distinct clade-specific functional constraints. J. Virol. 2014, 88, 4668–4678. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Xu, S.; Yu, M.; Wang, K.; Tao, Y.; Zhou, Y.; Shi, J.; Zhou, M.; Wu, B.; Yang, Z. Risk factors for severity and mortality in adult COVID-19 inpatients in Wuhan. J. Allergy Clin. Immunol. 2020, 146, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Guan, W.-J.; Liang, W.-H.; Zhao, Y.; Liang, H.-R.; Chen, Z.-S.; Li, Y.-M.; Liu, X.-Q.; Chen, R.-C.; Tang, C.-L.; Wang, T. Comorbidity and its impact on 1590 patients with COVID-19 in China: A nationwide analysis. Eur. Respir. J. 2020, 55, 2000547. [Google Scholar] [CrossRef] [PubMed]
- Gomez, F.; Hirbo, J.; Tishkoff, S.A. Genetic variation and adaptation in Africa: Implications for human evolution and disease. Cold Spring Harb. Perspect. Biol. 2014, 6, a008524. [Google Scholar] [CrossRef] [PubMed]
- Ellinghaus, D.; Degenhardt, F.; Bujanda, L.; Buti, M.; Albillos, A.; Invernizzi, P.; Fernández, J.; Prati, D.; Baselli, G.; Asselta, R.; et al. The ABO blood group locus and a chromosome 3 gene cluster associate with SARS-CoV-2 respiratory failure in an Italian-Spanish genome-wide association analysis. MedRxiv 2020. [Google Scholar] [CrossRef]
- The Severe COVID-19 GWAS Group. Genomewide association study of severe COVID-19 with respiratory failure. N. Engl. J. Med. 2020, 383, 1522–1534. [Google Scholar] [CrossRef] [PubMed]
- COVID-19 Host Genetics Initiative. Mapping the human genetic architecture of COVID-19. Nature 2021, 600, 472–477. [Google Scholar] [CrossRef]
- Poland, G.; Ovsyannikova, I.; Jacobson, R.; Smith, D. Heterogeneity in vaccine immune response: The role of immunogenetics and the emerging field of vaccinomics. Clin. Pharmacol. Ther. 2007, 82, 653–664. [Google Scholar] [CrossRef]


{kind=link}
| Gene | Ethnicity | Effect/Association | No. of Samples | p Value | Reference |
|---|---|---|---|---|---|
| HLA-A | |||||
| HLA-A*01 | Mexican | Risk of fatal COVID-19 | 146 111 COVID-19-infected individuals and 35 controls | Pc = 0.03 | [35] |
| Spain | Associated with higher mortality | 3886 | Apache p = 0.04 or sofa p = 0.02 | [36] | |
| West Indian | Prevalent in controls than COVID-19 patients | 228 controls, 235 COVID-19 patients | p = 0.011 | [37] | |
| HLA-A*01:01 | Russian | Protective against severe COVID-19 | 100 pneumonia caused by COVID-19 patients and 100 controls | p = 0.009 | [38] |
| HLA-A*02 | UK (Manchester and Leeds) | Might cause a protective effect or effective immune response against COVID-19 Protective against susceptibility and mortality | 80 COVID-19-infected individuals 308 wait-listed renal transplants 10,000 deceased | p = 0.0179 Insignificant after correction | [39] |
| West Indian | Prevalent among COVID-19-infected individuals of varying severity | 228 controls, 235 COVID-19 patients | p < 0.001 | [37] | |
| HLA-A*02:01 | German | Associated with symptomatic COVID-19 | 6919 COVID-19-infected individuals | p = 0.03 | [40] |
| Japanese | Low COVID-19 risk. | 1336 | [41] | ||
| 19 countries | Increased risk of COVID susceptibility and mortality. | p = 0.20 | [42] | ||
| HLA-A*02:05 | German | Associated with the risk of severe respiratory infection | 6919 COVID-19-infected individuals | p = 0.04 | [40] |
| HLA-A*26 | Manchester and Leeds | Increased in patients than in controls | 80 COVID-19-infected individuals, 308 wait-listed renal transplants, 10,000 deceased donors | p = 0.0049 | [39] |
| HLA-A*02:06 | Asia, North America, Europe, Oceania | 6421 sequences | [41] | ||
| HLA-A*03 | Prevalent in COVID-19 patients. | p = 0.047 | |||
| Iranian | Associated with risk | 142 COVID-19-infected individuals and 143 controls | p = 0.0025 | [43] | |
| HLA-A*03:01 | Low COVID-19 risk | [44] | |||
| HLA-A*11 | Spain | Higher mortality Increased frequency in deceased than in survivors | 3886 | SOFA (p = 0.04) APACHE (p = 0.02) p = 0.051 | [36] |
| Chinese | Possibly confers susceptibility in SARS-CoV-2 infection. | 332 patients | p = 8.5 × 10−3 | [45] | |
| HLA-A*11:01 | Japanese | Severe disease, hospitalization, and mortality COVID-19 severity Protective against COVID-19 susceptibility and mortality | 190 | p = 3.34 × 10−3 p = 0.013 p = 0.0078 | [46] |
| Albany, NY, USA | Increased risk of hospitalization | 100 hospitalized COVID-19 patients and 26 controls | p = 0.0078 | [47] | |
| Spanish | COVID-19 severity | 5943 controls 9373 COVID-19-infected individuals | p = 0.033 | [48] | |
| HLA-A*23:01 | Brazilian | Protection against COVID-19 | [49] | ||
| HLA-A*24 | Iranian | 48 severe COVID-19 cases | p = 0.003 | ||
| HLA-A*24:02 | Brazilian | Protection against COVID-19 | [49] | ||
| Ecuadorians | Protection against severe COVID-19 | [50] | |||
| HLA-A*24:02:01 | Chinese | Susceptibility | 5 | [51] | |
| HLA-A*24:02 | Japanese | Worse COVID-19 outcomes | 1336 | [41] | |
| HLA-A*26:01 | Russian | Worse COVID-19 outcome | 111 COVID-19-infected individuals and 428 controls | p = 0.0459 | [44] |
| South Han Chinese | Associated with diabetes a risk factor for COVID-19 | 5 | [51] | ||
| Russian | 111 | 0.0400 | |||
| HLA-A*30:02 | African American | Increased COVID-19 susceptibility | 234 COVID-19 cases and 22,000 controls | p = 0.01 | [52] |
| Albany, NY, USA | Enriched in COVID-19-positive individuals | 100 hospitalized COVID-19-infected individuals and 26 controls | (Exact test) p = 0.0417 | [47] | |
| Brazilian | Protection against COVID-19 | [49] | |||
| HLA-A*31:01 | Brazilian | Protection against COVID-19 | [49] | ||
| HLA-A*32 | Spain | Higher in healthy controls than COVID-19 patients | 3886 | (p = 0.004) | [36] |
| Iranian | Protection against COVID-19 | 143 controls and 142 COVID-19-infected individuals | p = 0.0388 | [43] | |
| HLA-A*68 | Iranian | Prevalent in COVID-19 | 48 severe COVID-19 and 500 controls | p = 0.001 | [53] |
| Mexican | Protective against severe COVID-19 | 146 111 COVID-19-infected individuals and 35 controls | PC = 0.03 | [35] | |
| HLA-A*68:01 | Brazilian | Protection against COVID-19 | [49] | ||
| HLA-B | |||||
| HLA-B*07 | 74 countries | Risk of mortality | p = 0.00081 Insignificant after multivariable regression | [54] | |
| HLA-B*07:03 | Hong Kong | Disease | 90 | p = 0.00072 | [55] |
| HLA-B*08 | Italians | Increased COVID-19 and death rate | HLA−A*01:01g−B*08:01g−C*07:01g−DRB1*03:01gG (p = 0.00042, p = 0.013) | [56] | |
| HLA-B*08:01 | 74 countries | Increased COVID-19 and death rate | 104,135 | p = 0.047 (insignificant after multivariate regression with backward elimination) | [54] |
| HLA-B*08:01 | 209 populations | 420 HLA-B alleles | <0.0001 | [57] | |
| HLA-B*14 | Italians | Inversely associated with COVID-19 | 370,000 | p < 0.0001 | [58] |
| HLA-B*14:02 | Chinese | Patients entering the severe stage. | 332 | p = 3 × 10−3 | [59] |
| HLA-B*15 | West Indian | Protection against COVID-19 | 228 controls, 235 COVID-19 patients | p = 0.008 | [37] |
| Egyptian | Protection | 69 | p < 0.001 | [60] | |
| HLA-B*15:03 | Protective against COVID-19 | [61] | |||
| HLA-B*15:01 | 805 district populations from 101 countries | Positively associated with COVID-19 | [62] | ||
| Asymptomatic SARS-CoV-2 | [63] | ||||
| HLA-B1527 | Chinese | More frequent in COVID19-infected individuals than in healthy controls | 82 | p = 0.001 | [59] |
| HLA-B*18 | Italians | Inversely associated with COVID-19 | 370,000 and additional 120,926 individuals | p < 0.0001 | [58] |
| HLA-B*18:01 | Italian | Protects against COVID-19 incidence and mortality | HLA-A*02.01g-B*18.01g-C*07.01g-DRB1*11.04g (p = 0.0053, p = 0.034) | [56] | |
| Brazilian | Protection against COVID-19 | [49] | |||
| HLA-B*22 | Chinese | SARS-CoV-2 susceptibility | 190 COVID-19-infected individuals and 294 controls | p = 0.032 | [64] |
| HLA-B*27 | Chinese | More prevalent among controls than COVID-19 patients Susceptibility and resistance to all SARS-CoV-2 strains | 190 COVID-19-infected individuals and 294 controls | p = 0.068 | [64] |
| HLA-B*35 | United Arab Emirates (15 nationalities) | Severe COVID-19 | 92 patients | p = 0.0051 | [65] |
| South Asian | Severe COVID-19 | ||||
| HLA-B*37:01 | Russia | Associated with deceased COVID-19 individuals | 111 COVID-19-infected individuals and 428 controls | p = 0.0331 | [44] |
| HLA-B*38 | Iranian | Disease susceptibility | 48 severe cases of COVID-19 | p < 0.001 | [53] |
| HLA-B*39 | Spain | Higher COVID-19 rates | 72 infected out of 3886 | p = 0.02 | [62] |
| Ecuadorian | Associated with COVID-19 risk | 52 COVID-19-infected individuals and 87 controls | [50] | ||
| HLA-B*41 | Egyptian | Associated with severe COVID-19 | 69 | [60] | |
| HLA-B*44 | Italian | High risk for COVID-19 susceptibility, severity, and mortality in population-based studies. | 182 patients, 619 controls | p = 0.175 | [66] |
| 74 countries | Risk of mortality | p = 0.0022 (insignificant after multivariable regression) | [54] | ||
| UK (Manchester and Leeds) | Protective effect | 10,000 deceased donors, 308 wait-listed renal patients, and 80 COVID-19-infected individuals | p = 0.0052 (did not remain significant after correction). | [39] | |
| HLA-B*44:02 | 60 countries 209 populations | High risk for COVID-19 susceptibility, severity, and mortality in population-based studies. | 420 Hla-b alleles | 0.0003 | [57] |
| Brazilian | Protection against COVID-19 | [49] | |||
| German | Associated with the risk of hospitalization | 6919 COVID-19-infected individuals | p = 0.008 | ||
| HLA-B*46:01 | Chinese, Vietnamese, Taiwan, Singaporean | Increased COVID-19 susceptibility | [67] | ||
| Prevalent in mild COVID-19 | [68] | ||||
| HLA-B*49 | Italians | Inversely associated with COVID-19 | 370,000 and additional 120,926 individuals | p < 0.0001 | [58] |
| HLA-B*51 | South Asian | Fatal COVID-19 | [69] | ||
| HLA-B*52:01 | African, European, Asian, Australian, Oceanian, American. | 158 and 374 typed samples | [70] | ||
| HLA-B*54:01 | Six areas (Asia, North America, South America, Europe, Oceania, and Africa) | SARS-CoV-2 susceptibility | 158 and 374 typed samples | [70] | |
| Protection against COVID-19 infection | 12,343 SARS-CoV-2 | p = 0.017 (insignificant after adjusted p = 0.45) | [71] | ||
| HLA-B*55 | Iranian | Protection | 142 COVID-19 patients and 143 controls | p = 0.0033 | [43] |
| HLA-B*55:01 | African, European, Asian, Australian, Oceanian, American. | SARS-CoV-2 susceptibility | 158 and 374 typed samples | [70] | |
| HLA-B*55:07 | African, European, Asian, Australian, Oceanian, American. | SARS-CoV-2 susceptibility | 158 and 374 typed samples | [70] | |
| HLA-B*55:12 | African, European, Asian, Australian, Oceanian, American. | SARS-CoV-2 susceptibility | 158 and 374 typed samples | [70] | |
| HLA-B*51:01 | Chinese | Severe COVID-19 | 332 patients | p = 0.007 | [45] |
| Brazilian | Protection against COVID-19 | [49] | |||
| HLA-B*56:01 | African, European, Asian, Australian, Oceania, American. | SARS-CoV-2 susceptibility | 158 and 374 typed samples | [70] | |
| Hong Kong Chinese | Associated with COVID-19 | 190 COVID-19 cases | p = 0.045 | [64] | |
| HLA-B*56:04 | Hong Kong Chinese | Associated with COVID-19 | 190 COVID-19 cases | p = 0.029 | [64] |
| HLA-B*58 | Iranian | Protection | 143 controls and 142 COVID-19-infected individuals | p = 0.0376 | [43] |
| 74 countries | Risk of death | p = 0.0089 (insignificant after adjustment). | [54] | ||
| HLA-B*58:01 | 209 populations | High risk for COVID-19 susceptibility, severity, and mortality in population-based studies. | 0.0062 | [57] | |
| Italian | Positively associated with COVID-19 | 99 patients | p = 0.01317 | [72] | |
| HLA-C | |||||
| HLA-C*01 | Prevalent in mild COVID-19 infection compared to severe COVID-19 | 228 controls, 235 COVID-19 patients | p = 0.004 | [37] | |
| Italian | Permissive to SARS-CoV-2. More prevalent in the deceased than in survivors. | p = 0.09 | [36,58] | ||
| HLA-C*03 | Italian | Positively associated with the incidence of SARS-CoV-2 infection | 370,000 individuals and an additional 120,926 individuals | p > 0.0001 | [58] |
| Saudi | Increased COVID-19 severity | 136 COVID-19 patients | p = 0.047 | [69] | |
| HLA-C*04:01 | Germany, Spain, Switzerland, and the United States | Severe COVID-19 | 435 | p = 0.0074 | [73] |
| Europeans | Severe COVID-19 | 619 controls and 182 infected individuals. | [74] | ||
| Russian | Associated with COVID-19 susceptibility. | 12,139 | [75] | ||
| Sardinian | Susceptibility to SARS-CoV-2 infection | 619 controls, 182 SARS-CoV-2 patients | p = 0.001 | [66] | |
| Increased risk of COVID-19 | p = 0.005 | [76] | |||
| Albany, NY, USA | Severe COVID-19 | 100 hospitalized COVID-19 infections and 26 controls | p = 0.0087 | [47] | |
| Increased risk of hospitalization measured by days with ventilation | p = 0.0023 | ||||
| United Arab Emirates (15 nationalities) | COVID-19 severity | 92 COVID-19-infected individuals | p = 0.0077 | [65] | |
| HLA-C*05 | Spain | Severe COVID-19 | 9373 COVID-19-infected individuals and 5943 controls | p = 0.045 | [48] |
| 74 countries | Risk of COVID-19 death | p = 0.000027 | [54] | ||
| HLA-C*05:01 | Brazilian | COVID-19 protection | [49] | ||
| HLA-C*06:02 | Manchester and Leeds | Worse COVID-19 disease outcome | 80 infected out of 308 | [39] | |
| HLA-C*7 | Egyptian | Associated with protection from death | 69 COVID-19 patients | p = 0.001 | [60] |
| HLA-C*07:01 | Sardinian | Negatively correlates with SARS-CoV-2 susceptibility and mortality | 619 controls, 182 SARS-CoV-2 patients | p = 0.0406 | [66] |
| Brazilian | COVID-19 protection | [49,77] | |||
| German | Decreased risk of symptomatic COVID-19 | 6919 COVID-19-infected individuals | p = 0.001 | [40] | |
| HLA-C*07:29 | Chinese | Higher expression in COVID-19 patients than controls. | 82 | p = 0.001 | [59] |
| HLA-C*08:02 | Sardinian | Increased susceptibility SARS-CoV-1 | 619 controls, 182 SARS-CoV-2 patients | HLA-A*30:02, HLA-B*14:02, and HLA-C*08:02 haplotypes (p = 0.0008) | [66] |
| Brazilian | COVID-19 protection | [49] | |||
| Spanish Mediterranean Caucasian | Mild COVID-19 | p = 0.0014 | [48] | ||
| Spanish | Reduced risk of COVID-19 | 9373 COVID-19 positive cases and 5943 controls | p = 0.024 | [48] | |
| HLA-C*12 | Manchester and Leeds | Prevalent in the control population | 80 COVID-19-infected, 308 wait-listed renal transplants (control), and 10,000 deceased donors (control) | p = 0.0286 | [39] |
| Egyptian | Protection from death | 69 COVID-19 patients | p = 0.008 | [60] | |
| HLA-C*12:02 | Han | High response against COVID-19 | 5 | [51] | |
| HLA-C*12:03 | Spanish Mediterranean Caucasian | Mild COVID-19 compared to critical COVID-19 | 72 individuals, 24 COVID-19-infected individuals, and 48 hospitalized | p = 0.0001 | [78] |
| HLA-C*14:02 | Chinese | Severe COVID-19 | 332 | p = 0.003 | [45] |
| HLA-C*15:02 | Brazilian | COVID-19 protection | [49] | ||
| HLA-C*16 | Egyptian | COVID-19 severity | 69 COVID-19 patients | [60] | |
| Spain | Increased COVID-19 infection | 3886 | p = 0.02 | [36] | |
| HLA-C*16:01 | Spanish Mediterranean Caucasian population | Associated more with mild COVID-19 when compared to critical than severe | 72 individuals, 24 COVID-19-infected individuals, and 48 hospitalized | p = 0.0014 | [78] |
| HLA-C*17 | Egyptian | COVID-19 severity | 69 COVID-19 patients | [60] | |
| HLA-C*17:01 | Brazilian | Associated with COVID-19 protection | [49,77] | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Naidoo, L.; Arumugam, T.; Ramsuran, V. Narrative Review Explaining the Role of HLA-A, -B, and -C Molecules in COVID-19 Disease in and around Africa. Infect. Dis. Rep. 2024, 16, 380-406. https://doi.org/10.3390/idr16020029
Naidoo L, Arumugam T, Ramsuran V. Narrative Review Explaining the Role of HLA-A, -B, and -C Molecules in COVID-19 Disease in and around Africa. Infectious Disease Reports. 2024; 16(2):380-406. https://doi.org/10.3390/idr16020029
Chicago/Turabian StyleNaidoo, Lisa, Thilona Arumugam, and Veron Ramsuran. 2024. "Narrative Review Explaining the Role of HLA-A, -B, and -C Molecules in COVID-19 Disease in and around Africa" Infectious Disease Reports 16, no. 2: 380-406. https://doi.org/10.3390/idr16020029
APA StyleNaidoo, L., Arumugam, T., & Ramsuran, V. (2024). Narrative Review Explaining the Role of HLA-A, -B, and -C Molecules in COVID-19 Disease in and around Africa. Infectious Disease Reports, 16(2), 380-406. https://doi.org/10.3390/idr16020029