
Extracellular Production of Glutathione by Recombinant Escherichia coli K-12

Abstract
:1. Introduction
2. Materials and Methods
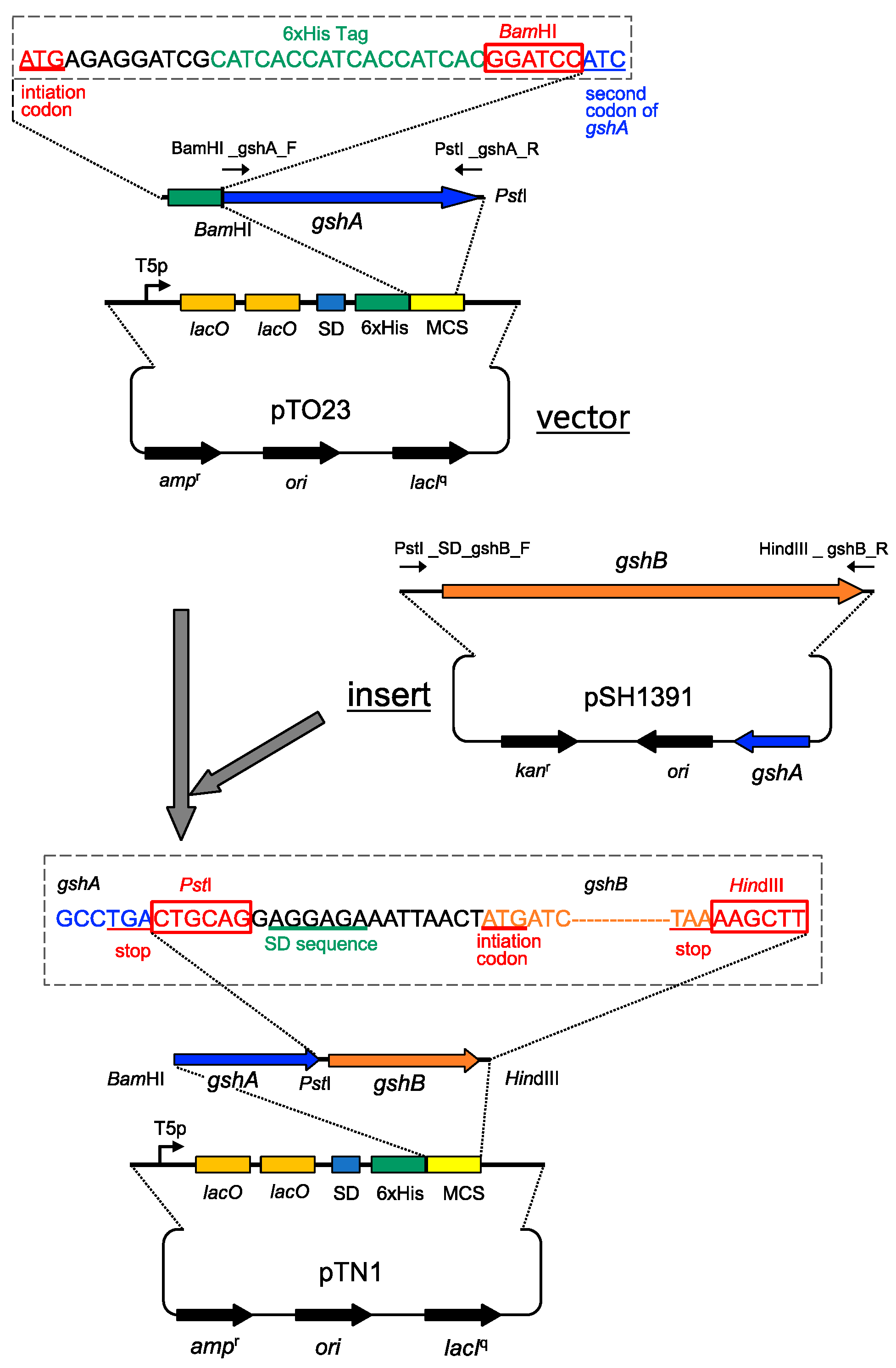
2.1. Strains and Plasmids Used in This Study
2.1.1. Introduction of Mutations into gshA Gene
2.1.2. Cloning of cysE on pQE-80L Plasmid and Introduction of V95R D96P Mutation
2.2. Purification of GshA with Different Combinations of Mutations
2.2.1. Growth Conditions of Strains for Purification of GshA
2.2.2. Purification of GshA by Nickel Sepharose Column
2.2.3. Measurement of GshA Activity
2.3. Glutathione Production
2.3.1. Growth Conditions of Strains for Glutathione Production
2.3.2. Preparation of Supernatants from Extracellular Samples
2.3.3. Measurement of glutathione concentration
- (1)
- Measurement by HPLC equipped with an Amino Na column
- (2)
- Enzymatic method using glutathione reductase
3. Results and Discussion
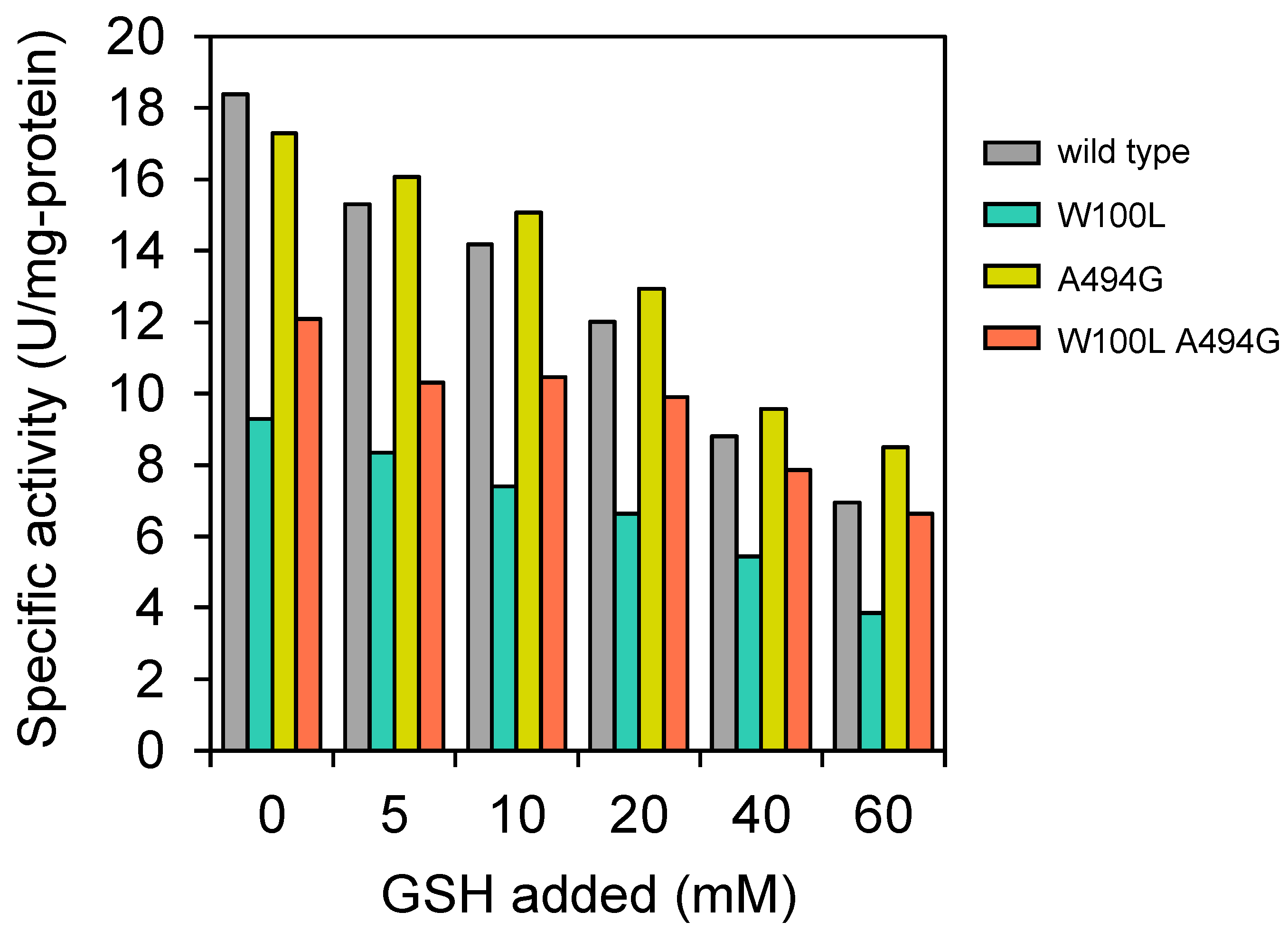
3.1. Evaluation of Desensitized Mutations of GshA
3.2. Effect of The Addition of Sodium Thiosulfate (Na2S2O3) to M9 1% Glucose Medium Containing 1 mM MgSO4

3.3. Effect of Wild-Type and Desensitized CysE on Glutathione Production

3.4. Production of Reduced and Oxidized Glutathione by Addition of Sodium Glutamate and Glycine

4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Janowiak, B.E.; Griffith, O.W. Glutathione synthesis in Streptococcus agalactiae. One protein accounts for γ-glutamylcysteine synthetase and glutathione synthetase activities. J. Biol. Chem. 2005, 280, 11829–11839. [Google Scholar] [CrossRef] [PubMed]
- Vergauwen, B.; De Vos, D.; Van Beeumen, J.J. Characterization of the bifunctional γ-glutamate-cysteine ligase/glutathione synthetase (GshF) of Pasteurella multocida. J. Biol. Chem. 2006, 281, 4380–4394. [Google Scholar] [PubMed]
- Yang, J.; Li, W.; Wang, D.; Wu, H.; Li, Z.; Ye, Q. Characterization of bifunctional L-glutathione synthetases from Actinobacillus pleuropneumoniae and Actinobacillus succinogenes for efficient glutathione biosynthesis. Appl. Microbiol. Biotechnol. 2016, 100, 6279–6289. [Google Scholar]
- Vairetti, M.; Pasqua, L.G.D.; Cagna, M.; Richelmi, P.; Ferrigo, A.; Berardo, C. Changes in glutathione content in liver diseases: An update. Antioxidants 2021, 10, 364. [Google Scholar] [PubMed]
- Ueda, Y.; Yonemitsu, M.; Tsubuku, T.; Sakaguchi, M.; Miyajima, R. Flavor characteristics of glutathione in raw and cooked foodstuffs. Biosci. Biotechnol. Biochem. 1997, 61, 1977–1980. [Google Scholar] [CrossRef]
- Ueda, Y.; Sakaguchi, M.; Hirayama, K.; Miyajima, R.; Kimizuka, A. Characteristic flavor constituents in water extract of garlic. Agric. Biol. Chem. 1990, 54, 163–169. [Google Scholar]
- Ogawa, K.; Henmi, K. Plant growth regulators and their use. JP 5452022 2014. [Google Scholar]
- Gul, N.; Ahmad, P.; Wani, T.A.; Tyagi, A.; Aslam, S. Glutathione improves low temperature stress tolerance in pusa sheetal cultivar of Solanum lycopersicum. Sci. Rep. 2022, 12, 12548. [Google Scholar]
- Schmacht, M.; Lorenz, E.; Senz, M. Microbial production of glutathione. World J. Microbiol. Biotechnol. 2017, 33, 106. [Google Scholar]
- Suzuki, H.; Kumagai, H.; Tochikura, T. Isolation, genetic mapping, and characterization of Escherichia coli K-12 mutants lacking γ-glutamyltranspeptidase. J. Bacteriol. 1987, 169, 3926–3931. [Google Scholar]
- Suzuki, H.; Koyanagi, T.; Izuka, S.; Onishi, A.; Kumagai, H. The yliA, -B, -C, and -D genes of Escherichia coli K-12 encode a novel glutathione importer with an ATP-binding cassette. J. Bacteriol. 2005, 187, 5861–5867. [Google Scholar]
- Suzuki, H.; Hashimoto, W.; Kumagai, H. Glutathione metabolism in Escherichia coli. J. Mol. Catal. B 1999, 6, 175–184. [Google Scholar]
- Santos, L.O.; Silva, P.G.P.; Lemos, W.J.F., Jr.; de Oliveira, V.S.; Anschau, A. Glutathione production by Saccharomyces cerevisiae: Current state and perspectives. Appl. Microbiol. Biotechnol. 2022, 106, 1879–1894. [Google Scholar] [PubMed]
- Fujio, T. Method for producing substances by utilizing the ATP recycling activity of stationary bacterial cells. In Great Development of Microorganisms; Imanaka, T., Ed.; NTS: Tokyo, Japan, 2002; pp. 1135–1140. (In Japanese) [Google Scholar]
- Kumagai, H.; Suzuki, H.; Shimizu, M.; Tochikura, T. Utilization of the γ-glutamyltranspeptidase reaction for glutathione synthesis. J. Biotechnol. 1989, 9, 129–138. [Google Scholar] [CrossRef]
- Watanabe, K.; Yamano, Y.; Murata, K.; Kimura, A. The nucleotide sequence of the gene for γ-glutamylcysteine synthetase of Escherichia coli. Nucleic Acids Res. 1986, 14, 4393–4400. [Google Scholar] [CrossRef] [PubMed]
- Kai, Y.; Kashiwagi, T.; Ishikawa, K.; Ziyatdinov, M.K.; Redkina, E.I.; Kiriukhin, M.Y.; Gusyatiner, M.M.; Kobayashi, S.; Takagi, H.; Suzuki, E. Engineering of Escherichia coli L-serine O-acetyltransferase on the basis of crystal structure: Desensitization to feedback inhibition by L-cysteine. Prot. Eng. Des. Sel. 2006, 19, 163–167. [Google Scholar]
- Miller, J.H. Experiments in Molecular Genetics; Cold Spring Harbor Laboratory: Cold Spring Harbor, NY, USA, 1972. [Google Scholar]
- Suzuki, H.; Kumagai, H.; Tochikura, T. γ-Glutamyltranspeptidase from Escherichia coli K-12: Purification and properties. J. Bacteriol. 1986, 168, 1325–1331. [Google Scholar]
- Kurihara, S.; Oda, S.; Tsuboi, Y.; Kim, H.G.; Oshida, M.; Kumagai, H.; Suzuki, H. γ-Glutamylputrescine synthetase in the putrescine utilization pathway of Escherichia coli K-12. J. Biol. Chem. 2008, 283, 19981–19990. [Google Scholar]
- Liu, H.; Fang, G.; Wu, H.; Li, Z.; Ye, Q. L-Cysteine production in Escherichia coli based on rational metabolic engineering and modular strategy. Biotechnol. J. 2018, 13, e1700695. [Google Scholar]
- Lennox, E.S. Transduction of linked genetic characters of the host by bacteriophage P1. Virology 1955, 1, 190–206. [Google Scholar]
- Suzuki, H.; Izuka, S.; Minami, H.; Miyakawa, N.; Ishihara, S.; Kumagai, H. Use of bacterial γ-glutamyltranspeptidase for enzymatic synthesis of γ-D-glutamyl compounds. Appl. Environ. Microbiol. 2003, 69, 6399–6404. [Google Scholar] [PubMed]
- Fahey, R.C.; Brody, S.; Mikolajczyk, S.D. Changes in the glutathione thiol-disulfide status of Neurospora crassa conidia during germination and aging. J. Bacteriol. 1975, 121, 144–151. [Google Scholar]
- Sakamoto, A.; Terui, Y.; Yoshida, T.; Yamamoto, T.; Suzuki, H.; Yamamoto, K.; Ishihama, A.; Igarashi, K.; Kashiwagi, K. Three members of polyamine modulon under oxidative stress conditions: Two transcription factors (SoxR and EmrR) and a glutathione synthetic enzyme (GshA). PLoS ONE 2015, 10, e0124883. [Google Scholar]
- Richman, P.G.; Meister, A. Regulation of γ-glutamyl-cysteine synthetase by nonallosteric feedback inhibition by glutathione. J. Biol. Chem. 1975, 250, 1422–1426. [Google Scholar] [PubMed]
- Hibi, T.; Nii, H.; Nakatsu, T.; Kimura, A.; Kato, H.; Hiratake, J.; Oda, J. Crystal structure of γ-glutamylcysteine synthetase: Insights into the mechanism of catalysis by a key enzyme for glutathione homeostasis. Proc. Natl. Acad. Sci. USA 2004, 101, 15052–15057. [Google Scholar]
- Gushima, H.; Miya, T.; Murata, K.; Kimura, A. Construction of glutathione-producing strains of Escherichia coli B by recombinant DNA techniques. J. Appl. Biochem. 1983, 5, 43–52. [Google Scholar]
- Watanabe, K.; Yamano, Y.; Murata, K.; Kimura, A. Glutathione production by Escherichia coli cells with hybrid plasmid containing tandemly polymerized genes for glutathione synthetase. Appl. Microbiol. Biotechnol. 1986, 24, 375–378. [Google Scholar] [CrossRef]
- Li, W.; Li, Z.; Yang, J.; Ye, Q. Production of glutathione using a bifunctional enzyme encoded by gshF from Streptococcus thermophilus expressed in Escherichia coli. J. Biotechnol. 2011, 154, 261–268. [Google Scholar]
- Wang, D.; Wang, C.; Wu, H.; Li, Z.; Ye, Q. Glutathione production by recombinant Escherichia coli expressing bifunctional glutathione synthetase. J. Ind. Microbiol. Biotechnol. 2016, 43, 45–53. [Google Scholar]
- Zhang, J.; Quan, C.; Wang, C.; Wu, H.; Li, Z.; Ye, Q. Systematic manipulation of glutathione metabolism in Escherichia coli for improved glutathione production. Microb. Cell Fact. 2016, 15, 38. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Source or Reference | |
|---|---|---|
| DH5α | F- Φ80(ΔlacZΔM15) Δ(lacZYA-argF)U169 deoR recA1 endA1 hsdR17(rK−, mK+) phoA supE44 λ- thi-1 gyrA96 relA1 | Toyobo |
| SH1391 | pSH1391/DH5α | [11] |
| SI97 | rph-1 Δggt-2 htp+-Tn10 ΔgshA ΔgsiAB | [11] |
| TN4 | pTN1/SI97 | |
| TN9 | pTN5/SI97 | |
| TN10 | pTN6/SI97 | |
| TN13 | pTN16/SI97 | |
| TN31 | pTN29/SI97 | |
| TN37 | pTN36/SI97 | |
| TN40 | rph-1 Δggt-2 htp+-Tn10 ΔgshA ΔgsiAB ΔcysE::kanr-FRT | |
| TN48 | pTN36/TN40 | |
| Plasmid | Genotype | |
| pQE-80L | ColE1 ampr lacIq | Qiagen |
| pSH1391 | ColEI replicon rop+ amps::gshA+ gshB+ kanr tets | [11] |
| pSH1564 | ColEI replicon rop+ amps::gshAL1M W100L gshB+ kanr tets | Laboratory stock |
| pSH1778 | ColE1 replicon ampr T5p-cysE+ | |
| pTN35 | ColE1 replicon ampr T5p-cysEV95R D96P | |
| pTO23 | ColE1 replicon ampr lacIq T5p-ATG-His6-gshA | |
| pTN1 | ColE1 replicon ampr lacIq T5p-ATG-His6-gshA-gshB | |
| pTN5 | ColE1 replicon ampr lacIq T5p-ATG-His6-gshAW100L-gshB | |
| pTN6 | ColE1 replicon ampr lacIq T5p-ATG-His6-gshAA494G-gshB | |
| pTN16 | ColE1 replicon ampr lacIq T5p-ATG-His6-gshAW100L A494G-gshB | |
| pTN29 | ColE1 replicon ampr lacIq T5p-ATG-His6-gshA-gshB-cysE | |
| pTN36 | ColE1 replicon ampr lacIq T5p-ATG-His6-gshA-gshB-cysEV95R D96P | |
| pYU4 | ColEI replicon rop+ amps::gshAL1M A494G gshB+ kanr tets | Laboratory stock |
| Primer | Nucleotide sequence | |
| BamHI_gshA_F | CCCggatccATCCCGGACGTATCACAGGCG | |
| PstI_gshA_R | CCCctgcagTCAGGCGTGTTTTTCCAGCC | |
| PstI_gshB_F | CCCctgcagGAGGAGAAATTAACTATGATCAAGCTCGGCATCGTGATG | |
| HindIII_gshB_R | CCCaagcttTTACTGCTGCTGTAAACGTG | |
| EcoRI_SD_cysE_N | CCCgaattcATTAAAGAGGAGAAATTAACTATGTCGTGTGAAGAACTGGAAATTG | |
| HindIII_cysE_C | CCCaagcttTTAGATCCCATCCCCATACTC | |
| HindIII_SD_cysE_F | CCCaagcttGAGGAGAAATTAACTATGTCG | |
| V95R D96P | GTGCGTACCCGCGACCCGGCACGCCCGAAATACTCAACCCCGTTGTTATACC | |
| V95R D96P comp | GGTATAACAACGGGGTTGAGTATTTCGGGCGTGCCGGGTCGCGGGTACGCAC |
| Ingredients | Volume Added |
|---|---|
| 10× M9 buffer | 6.0 mL |
| 100 mM CaCl2 | 0.06 mL |
| 10% (weight/volume) glucose | 3.0 mL |
| 100 mM Na2S2O3 | 3.0 mL |
| 100 mM Na2SO4 | 3.0 mL |
| Lennox broth [22] | 6.0 mL |
| 1 M MgSO4 | 0.6 mL |
| Distilled water | 38.34 mL |
| Total | 60 mL |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suzuki, H.; Nishida, K.; Nakamura, T. Extracellular Production of Glutathione by Recombinant Escherichia coli K-12. Microbiol. Res. 2023, 14, 1199-1209. https://doi.org/10.3390/microbiolres14030080
Suzuki H, Nishida K, Nakamura T. Extracellular Production of Glutathione by Recombinant Escherichia coli K-12. Microbiology Research. 2023; 14(3):1199-1209. https://doi.org/10.3390/microbiolres14030080
Chicago/Turabian StyleSuzuki, Hideyuki, Kazuki Nishida, and Tatsuya Nakamura. 2023. "Extracellular Production of Glutathione by Recombinant Escherichia coli K-12" Microbiology Research 14, no. 3: 1199-1209. https://doi.org/10.3390/microbiolres14030080
APA StyleSuzuki, H., Nishida, K., & Nakamura, T. (2023). Extracellular Production of Glutathione by Recombinant Escherichia coli K-12. Microbiology Research, 14(3), 1199-1209. https://doi.org/10.3390/microbiolres14030080