
Promising Eco-Friendly Nanoparticles for Managing Bottom Rot Disease in Lettuce (Lactuca sativa var. longifolia)

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Synthesis of Nanomaterials
2.2. Enmiration, Isolation, Identification of Fungal Pathogens Responsible for Bottom Rot
2.3. Pathogenicity Tests
2.4. In Vitro Examination of Antifungal Activity of the Prepared Nanomaterials
2.5. In Vivo Examination of Antifungal Activity of the Prepared Nanomaterials
2.6. Soil Biological Activities
2.6.1. Microbial Populations
2.6.2. Soil Enzymes Activities
2.7. Plant Sampling and Analysis
2.7.1. Estimation of Photosynthetic Pigments
2.7.2. Cell Membrane Stability Index
2.7.3. Anatomical Structure
2.8. Statistical Analysis
3. Results and Discussion
3.1. Characterization of Synthesized Nanomaterials
3.2. Isolation and Frequency Percentages of Causal Agents of Lettuce Bottom Rot
3.3. Pathogenicity Test
3.4. Antifungal Performance of Nano-SiO2, Nano-γFe2O3 and Nano-Copper against R. solani In Vitro Experiment
3.5. Greenhouse Experiment
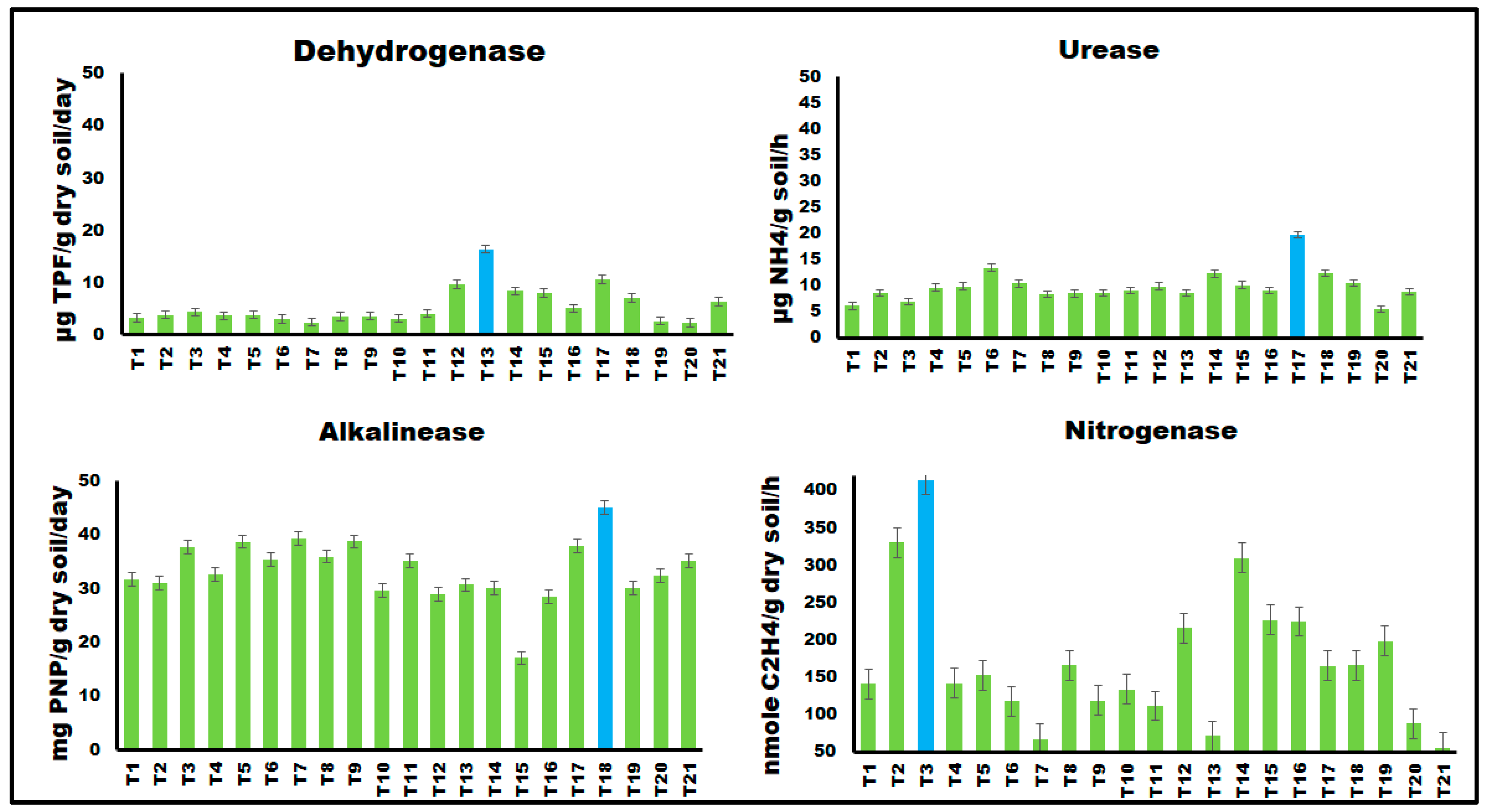
3.6. Effect of Metal Oxide NPs on Soil Biological Activities
3.6.1. Effect on Microbial Community
3.6.2. Effect on Soil Enzymatic Activities
3.7. Effect of Metal Oxide NPs on Endogenous Factors
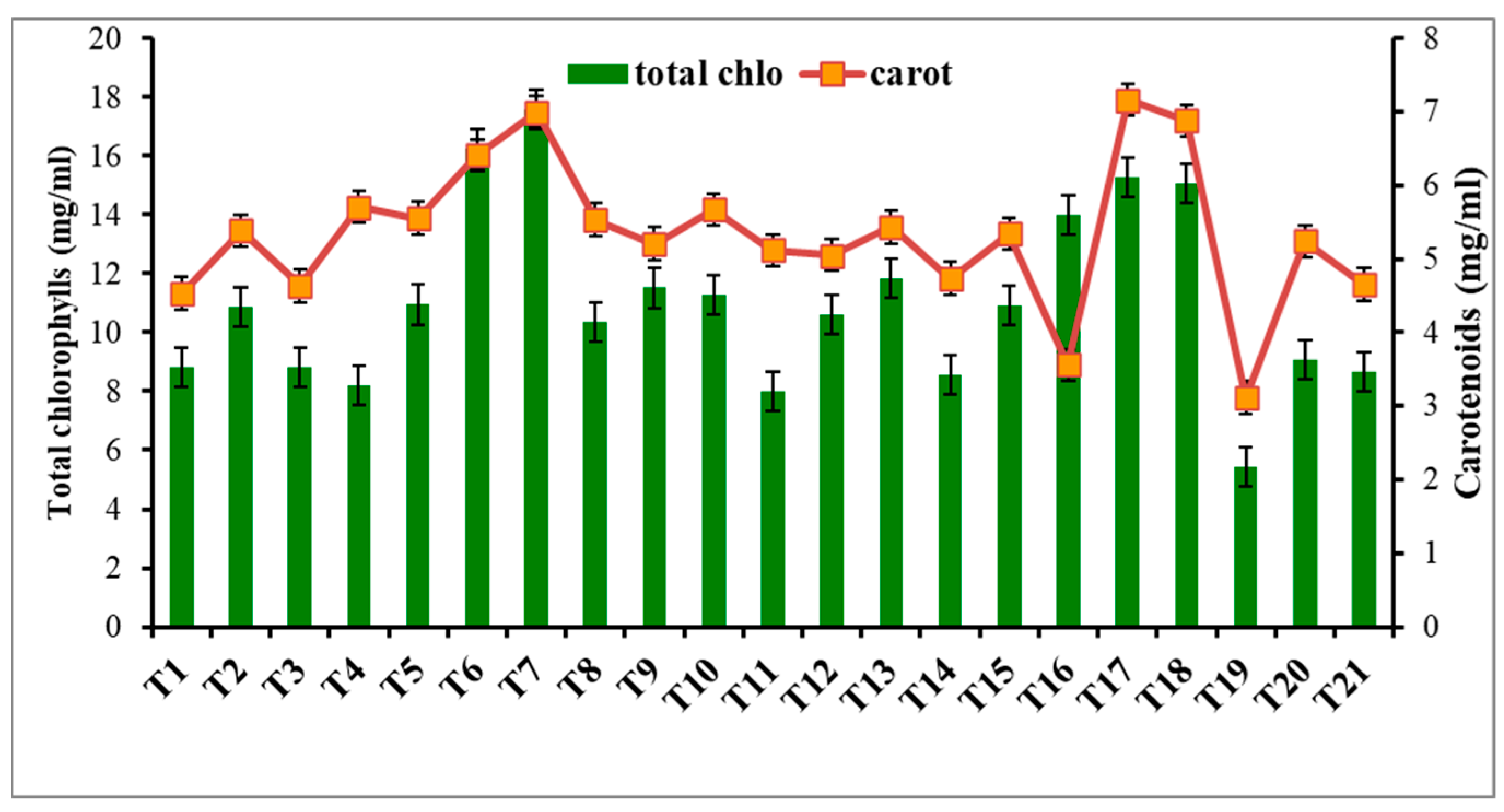
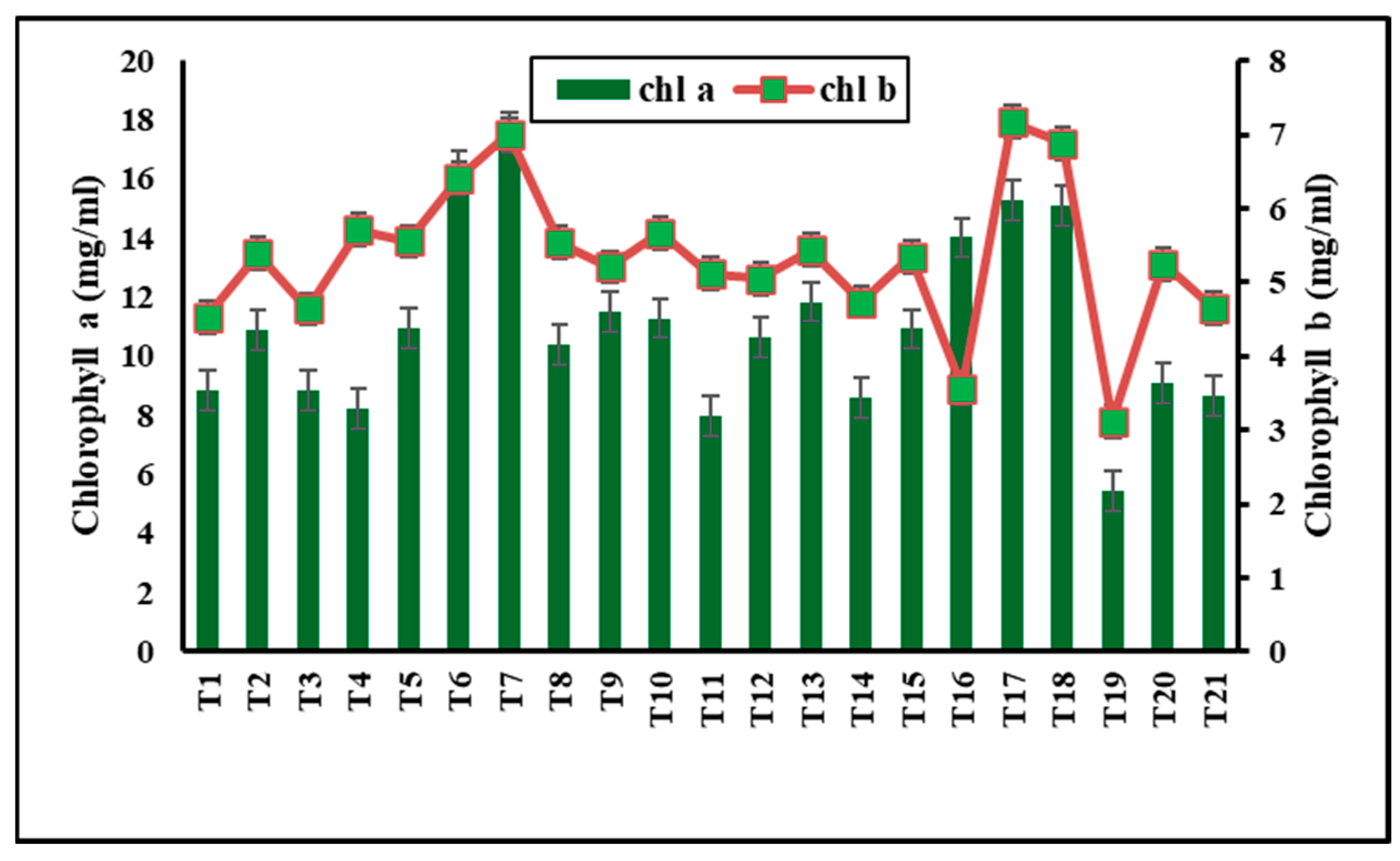
3.7.1. NPs Effect on Photosynthetic Pigments
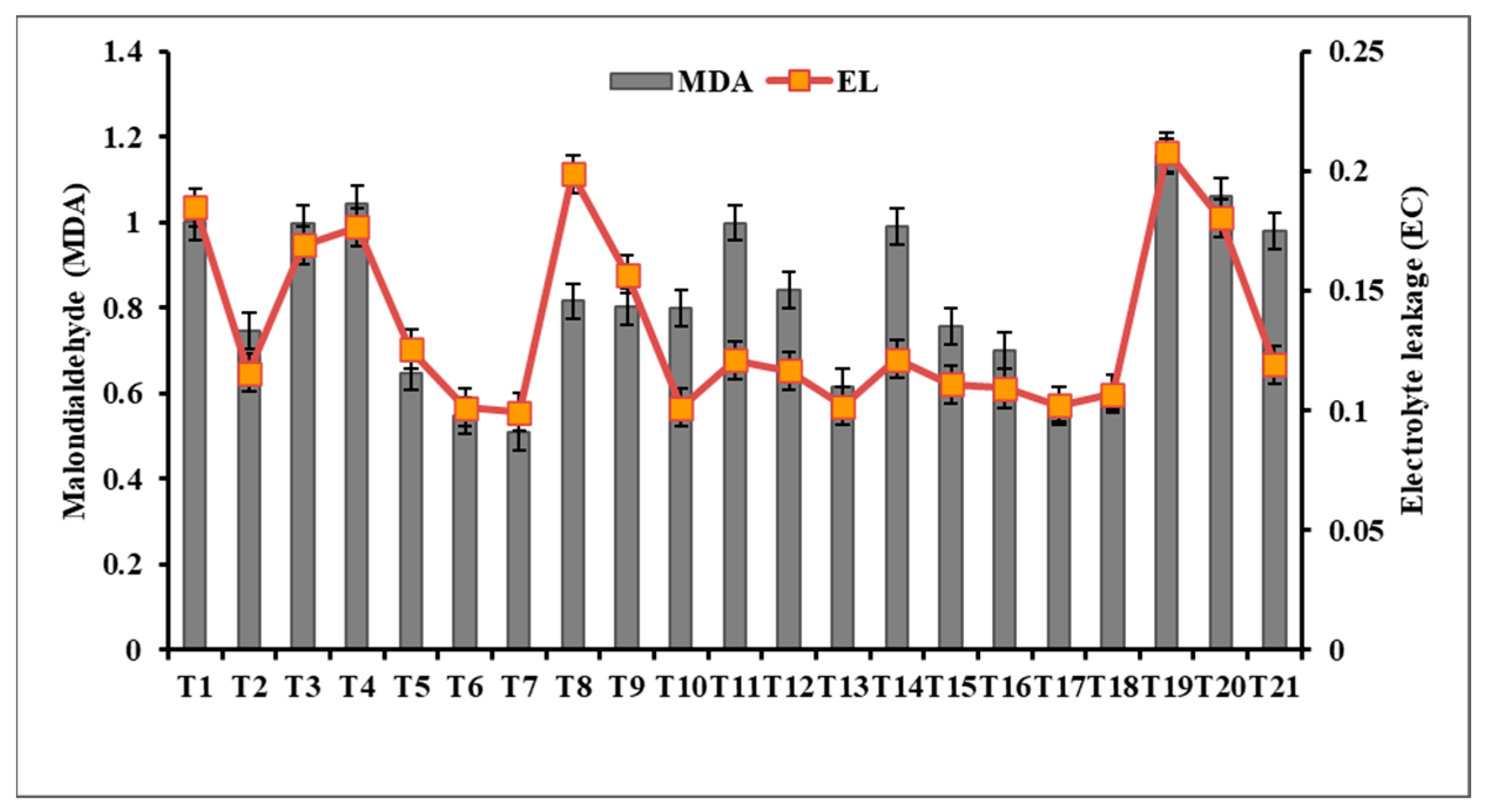
3.7.2. Effect of Metal oxide NPs on Malondialdehyde (MDA) and Electrolyte Leakage (EC)
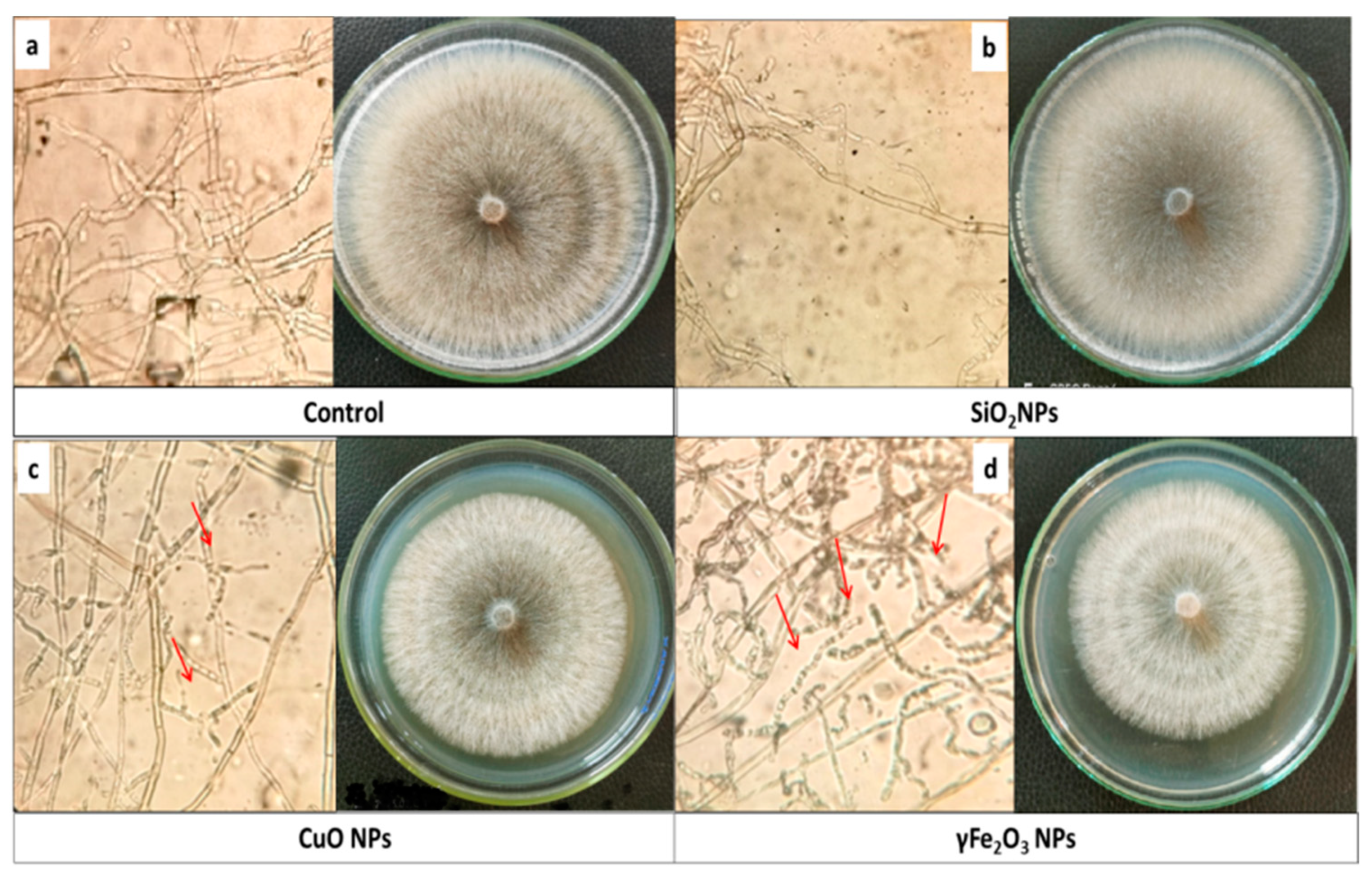
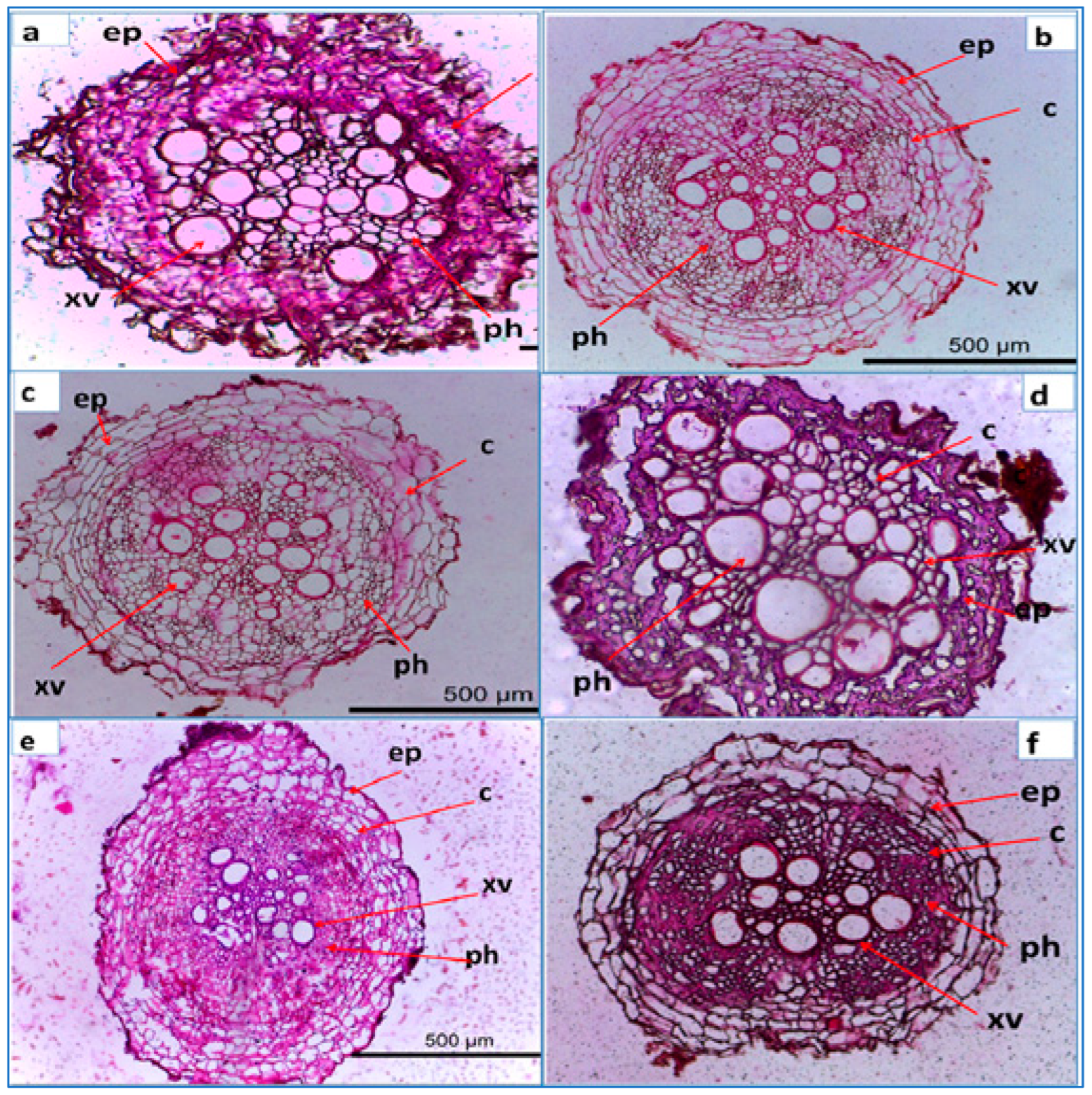
3.8. Anatomical Structure of Lettuce Root Infected with R. solani and Treated with Nano Metal Oxide
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Raza, A.; Razzaq, A.; Mehmood, S.S.; Zou, X.; Zhang, X.; Lv, Y.; Xu, J. Impact of climate change on crops adaptation and strategies to tackle its outcome: A review. Plants 2019, 8, 34. [Google Scholar] [CrossRef] [PubMed]
- Passarelli, M.; Bongiorno, G.; Cucino, V.; Cariola, A. Adopting new technologies during the crisis: An empirical analysis of agricultural sector. Technol. Forecast. Soc. Chang. 2023, 186, 122106. [Google Scholar] [CrossRef]
- Shang, Y.; Hasan, M.K.; Ahammed, G.J.; Li, M.; Yin, H.; Zhou, J. Applications of Nanotechnology in Plant Growth and Crop Protection: A Review. Molecules 2019, 24, 2558. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Khan, A.U.; Hasan, M.A.; Yadav, K.K.; Pinto, M.M.C.; Malik, N.; Yadav, V.K.; Khan, A.H.; Islam, S.; Sharma, G.K. Agro-nanotechnology as an emerging field: A novel sustainable approach for improving plant growth by reducing biotic stress. Appl. Sci. 2021, 11, 2282. [Google Scholar] [CrossRef]
- Grillo, R.; Abhilash, P.C.; Fraceto, L.F. Nanotechnology applied to bio-encapsulation of pesticides. J. Nanosci. Nanotechnol. 2016, 16, 1231–1234. [Google Scholar] [CrossRef] [PubMed]
- Kah, M.; Tufenkji, N.; White, J.C. Nano-enabled strategies to enhance crop nutrition and protection. Nat. Nanotechnol. 2019, 14, 532–540. [Google Scholar] [CrossRef] [PubMed]
- Massalimov, I.; Ram, P.; Amr, I.M.I.; Ahmed, I.S.A. Modern Prospects of Nanotechnology in Plant Pathology; Springer: Berlin/Heidelberg, Germany, 2017; pp. 1–372. [Google Scholar] [CrossRef]
- Wolf, P.F.J.; Lenz, R.; Baron, K.; Verreet, J.A. Quaternary integrated pest management concept for powdery mildew in sugar beet. I. Analysis of epidemic determinants to predict disease onset. J. Plant Dis. Prot. 2006, 113, 61–67. [Google Scholar] [CrossRef]
- Elshahawy, I.E.; El-Mohamedy, R.S. Biological control of Pythium damping-off and root-rot diseases of tomato using Trichoderma isolates employed alone or in combination. J. Plant Pathol. 2019, 101, 597–608. [Google Scholar] [CrossRef]
- Wallon, T.; Sauvageau, A.; Van der Heyden, H. Detection and quantification of rhizoctonia solani and rhizoctonia solani ag1-ib causing the bottom rot of lettuce in tissues and soils by multiplex qpcr. Plants 2021, 10, 57. [Google Scholar] [CrossRef]
- Martins, S.A.; Schurt, D.A.; Seabra, S.S.; Martins, S.J.; Ramalho, M.A.P.; Moreira, F.M.d.S.; da Silva, J.C.P.; da Silva, J.A.G.; de Medeiros, F.H.V. Common bean (Phaseolus vulgaris L.) growth promotion and biocontrol by rhizobacteria under Rhizoctonia solani suppressive and conducive soils. Appl. Soil Ecol. 2018, 127, 129–135. [Google Scholar] [CrossRef]
- Mondal, P.; Kumar, R.; Gogoi, R. Bioorganic Chemistry fungicidal evaluation against Sclerotium rolfsii, Rhizoctonia bataticola and Rhizoctonia solani. Bioorg. Chem. 2017, 70, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Raghupati, K.R.; Koodali, R.T.; Manna, A.C. Size-dependent bacterial growth inhibition and mechanism of antibacterial activity of zinc oxide nanoparticles. Langmuir 2011, 27, 4020–4028. [Google Scholar] [CrossRef]
- Jones, N.; Ray, B.; Ranjit, K.T.; Manna, A.C. Antibacterial activity of ZnO nanoparticles suspensions on a broad spectrum of microorganisms. FEMS Microbiol. Lett. 2008, 279, 71–76. [Google Scholar] [CrossRef]
- Azam, A.; Ahmed, A.S.; Oves, M.; Khan, M.S.; Habib, S.S.; Memic, A. Antimicrobial activity of metal oxide nanoparticles against Gram-positive and Gram-negative bacteria: A comparative study. Int. J. Nanomed. 2012, 7, 6003–6009. [Google Scholar] [CrossRef]
- Subhapriya, S.; Gomathipriya, P. Green synthesis of titanium dioxide (TiO2) nanoparticles by Trigonella foenum-graecum extract and its antimicrobial properties. Microb. Pathog. 2018, 116, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Tortella, G.; Rubilar, O.; Pieretti, J.C.; Fincheira, P.; de Melo Santana, B.; Fernández-Baldo, M.A.; Benavides-Mendoza, A.; Seabra, A.B. Nanoparticles as a Promising Strategy to Mitigate Biotic Stress in Agriculture. Antibiotics 2023, 12, 338. [Google Scholar] [CrossRef]
- Day, J.M.; Döbereiner, J. Physiological aspects of N2-fixation by a Spirillum from Digitaria roots. Soil Biol. Biochem. 1976, 8, 45–50. [Google Scholar] [CrossRef]
- Xiong, L.; Tong, Z.H.; Chen, J.J.; Li, L.L.; Yu, H.Q. Morphology-dependent antimicrobial activity of Cu/Cu𝑥O nanoparticles. Ecotoxicology 2015, 24, 2067–2072. [Google Scholar] [CrossRef]
- Khatami, M.; Varma, R.; Heydari, M.; Peydayesh, M.; Sedighi, A.; Askari, H.A.; Rohani, M.; Baniasadi, M.; Arkia, S.; Seyedi, F.; et al. Copper oxide nanoparticles greener aynthesis using tea and its antifungal efficiency on Fusarium solani. Geomicrobiol. J. 2019, 36, 777–781. [Google Scholar] [CrossRef]
- Kamel, S.M.; Elgobashy, S.F.; Omara, R.I.; Derbalah, A.S.; Abdelfatah, M.; El-Shaer, A.; Al-Askar, A.A.; Abdelkhalek, A.; Abd-Elsalam, K.A.; Essa, T.; et al. Antifungal Activity of Copper Oxide Nanoparticles against Root Rot Disease in Cucumber. J. Fungi 2022, 8, 911. [Google Scholar] [CrossRef]
- Elmer, W.H.; De La Torre-Roche, R.; Pagano, L.; Majumdar, S.; Zuverza-Mena, N.; Dimpka, C.; Gardea-Torresdey, J.; White, W. Effect of metalloid and metallic oxide nanoparticles on Fusarium wilt of watermelon. Plant. Dis. 2018, 102, 1394–1401. [Google Scholar] [CrossRef]
- El-Batal, A.I.; El-Sayyad, G.S.; Al-Shammari, B.M.; Abdelaziz, A.M.; Nofel, M.M.; Gobara, M.; Elkhatib, W.F.; Eid, N.A.; Salem, M.S.; Attia, M.S. Protective role of iron oxide nanocomposites on disease index, and biochemical resistance indicators against Fusarium oxysporum induced-cucumber wilt disease: In vitro, and in vivo studies. Microb. Pathog. 2023, 180, 106131. [Google Scholar] [CrossRef] [PubMed]
- Filho, J.A.W.; Duarte, H.S.S.; Rodrigues, F.A. Effect of foliar application of potassium silicate and fungicide on the severity of leaf rust and yellow leaf spot in wheat. Rev. Ceres 2013, 60, 726–730. [Google Scholar]
- Rodrigues, F.A.; Dallagnol, L.J.; Duarte, H.S.S.; Datnoff, L.E. Silicon control of foliar diseases in monocots and dicots. In Silicon and Plant Diseases; Springer International Publishing: Cham, Switzerland, 2015. [Google Scholar]
- Abdelrhim, A.S.; Mazrou, Y.S.A.; Nehela, Y.; Atallah, O.O.; El-Ashmony, R.M.; Dawood, M.F.A. Silicon Dioxide Nanoparticles Induce Innate Immune Responses and Activate Antioxidant Machinery in Wheat against Rhizoctonia solani. Plants 2021, 10, 2758. [Google Scholar] [CrossRef] [PubMed]
- Ainsworth, G.C.; James, P.W. Ainsworth and Bisby’s Dictionary of Fungi, 6th ed.; Commonwealth Mycological Institute: Kew, UK, 1971. [Google Scholar]
- Alexopoulos, C.J.; Mims, C.W.; Blackwell, M. Introductory Mycology, 4th ed.; John Wiley and Sons: New York, NY, USA, 1996. [Google Scholar]
- Liu, S.; Wei, L.; Hao, L.; Fang, N.; Chang, M.W.; Xu, R.; Yang, Y.; Chen, Y. Sharper and faster “Nano darts” kill more bacteria: A study of antibacterial activity of individually dispersed pristine single-walled carbon nanotube. ACS Nano 2009, 3, 3891–3902. [Google Scholar] [CrossRef]
- Abd-El-Khair, H.; El-Nagdi, W.M.A. Field application of bio-control agents for controlling fungal root rot and root-knot nematode in potato. Arch. Phytopathol. Plant Prot. 2014, 47, 1218–1230. [Google Scholar] [CrossRef]
- Fratemale, D.; Giamperi, L.; Ricci, D. Chemical Composition and antifungal activity of essential oil obtained from in vitro plants of Thymus mastichina L. J. Essent. Oil Res. 2003, 15, 278–281. [Google Scholar] [CrossRef]
- Wang, Q.; Xiong, D.; Zhao, P.; Yu, X.; Tu, B.; Wang, G. Effect of applying an arsenic-resistant and plant growth-promoting rhizobacterium to enhance soil arsenic phytoremediation by Populus deltoides LH05-17. J. Appl. Microbiol. 2011, 111, 1065–1074. [Google Scholar] [CrossRef]
- Black, C.A. Methods of Soil Analysis. Part 2. Chemical and Microbiological Properties, 2nd ed.; American Society of Agronomy: Madison, WI, USA, 1982. [Google Scholar]
- Atlas, R.M. Handbook of Microbiological Media for the Examination of Food; CRC Press: Boca Raton, FL, USA, 1995. [Google Scholar]
- Difco. Difco & BBL Manual: Manual of Microbiological Culture Media, 2nd ed.; Citeseer: College Park, MD, USA, 2009. [Google Scholar] [CrossRef]
- Döbereiner, J.; Day, J.M.; Dart, P.J. Nitrogenase activity in the rhizosphere of sugar cane and some other tropical grasses. Plant Soil 1972, 37, 191–196. [Google Scholar] [CrossRef]
- Abd-el-Malek, Y.; Ishac, Y.Z. Evaluation of Methods Used in Counting Azotobacters. J. Appl. Bacteriol. 1968, 31, 267–275. [Google Scholar] [CrossRef]
- Pikovskaya, R.I. Mobilization of phosphorus in soil connection with the vital activity of some microbial species. Microbiologiya 1984, 17, 362–370. [Google Scholar]
- Zahra, M.K. Studies of Silicate Bacteria. Master’s Thesis, Faculty of Agriculture Cairo University, Giza, Egypt, 1969; p. 111. [Google Scholar]
- Casida, L.E., Jr. Microbial Metabolic Activity in Soil as Measured by Dehydrogenase Determinationst. Appl. Environ. Microbiol. 1977, 34, 630–636. [Google Scholar] [CrossRef]
- Tabatabai, M.A.; Bremner, J.M. Assay of urease activity in soils. Soil Biol. Biochem. 1972, 4, 479–487. [Google Scholar] [CrossRef]
- Senwo, Z.N.; Ranatunga, T.D.; Tazisong, I.A.; Taylor, R.W.; He, Z. Phosphatase activity of Ultisols and relationship to soil fertility indices. J. Food Agric. Environ. 2007, 5, 262–266. [Google Scholar]
- Young, A.J. The photoprotective role of carotenoids in higher plants. Physiol. Plant. 1991, 83, 702–708. [Google Scholar] [CrossRef]
- Zedan, A.; Omar, S. Nano selenium: Reduction of severe hazards of atrazine and promotion of changes in growth and geneexpression patterns on Vicia faba seedlings. Afr. J. Biotechnol. 2019, 18, 502–510. [Google Scholar]
- Blokhina, O.; Virolainen, E.; Fagerstedt, K.V. Antioxidants, oxidative damage and oxygen deprivation stress: A review. Ann. Bot. 2003, 91, 179–194. [Google Scholar] [CrossRef]
- Saeed, M.; Dodd, P.B.; Sohail, L. Anatomical studies of stems, roots and leaves of.pdf. J. Hortic. For. 2010, 2, 87–94. [Google Scholar]
- Hoshmand, A. Design of Experiments for Agriculture and the Natural Sciences, 2nd ed.; Chapman and Hall: New York, NY, USA, 2006. [Google Scholar]
- Premaratne, W.A.P.J.; Priyadarshana, W.M.G.I.; Gunawardena, S.H.P.; De Alwis, A.A.P. Synthesis of nanosilica from paddy husk ash and their surface functionalization. J. Sci. Univ. Kelaniya Sri Lanka 2013, 8, 33–48. [Google Scholar] [CrossRef]
- Jeon, C.S.; Baek, K.; Park, J.K.; Oh, Y.K.; Lee, S.D. Adsorption characteristics of As (V) on iron-coated zeolite. J. Hazard. Mater. 2009, 163, 804–808. [Google Scholar] [CrossRef]
- Chen, J.P.; Lim, L.L. Key Factors in Chemical Reduction by Hydrazine for Recovery of Precious Metals. Chemosphere 2002, 49, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Rashid, A.; Younas, R.; Chong, R. A chemical reduction approach to the synthesis of copper nanoparticles. Int. Nano Lett. 2016, 6, 21–26. [Google Scholar] [CrossRef]
- Raghunath, A.; Perumal, E. Metal oxide nanoparticles as antimicrobial agents: A promise for the future. Int. J. Antimicrob. Agents 2017, 49, 137–152. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; He, Y.; Irwin, P.L.; Jin, T.; Shi, X. Antibacterial activity and mechanism of action of zinc oxide nanoparticles against Campylobacter jejuni. Appl. Environ. Microbiol. 2011, 77, 2325–2331. [Google Scholar] [CrossRef] [PubMed]
- Oussou-Azo, A.F.; Nakama, T.; Nakamura, M.; Futagami, T.; Vestergaard, M.C.M. Antifungal potential of nanostructured crystalline copper and its oxide forms. Nanomaterials 2020, 10, 1003. [Google Scholar] [CrossRef] [PubMed]
- Nashwa, A.H.F.; Mervat, A.H. Reduction of Urea Transformation in Soil Using Aqueous Extracted Leaves of Neem (Azadirachta indica) and Olive (Olea europaea L). Indian J. Environ. Prot. IJEP 2020, 40, 12–22. Available online: https://www.e-ijep.co.in/january-2020/ (accessed on 15 January 2023).
- Nashwa, A.H.F.; Massoud, O.N.; Ebtsam, M.M.; Khalil, H.M. Biological Evaluation of Soil Cultivated with Egyptian Clover (Trifolium alexndrinum L.) through Long Term Trial at Bahtim Region, Egypt. Sciences 2015, 5, 515–525. Available online: https://www.curresweb.com/mejas/mejas/2015/515-525.pdf (accessed on 12 May 2023).
- Rangaraj, S.; Gopalu, K.; Rathinam, Y.; Periasamy, P.; Venkatachalam, R.; Narayanasamy, K. Effect of silica nanoparticles on microbial biomass and silica availability in maize rhizosphere. Biotechnol. Appl. Biochem. 2014, 61, 668–675. [Google Scholar] [CrossRef]
- Díaz-Raviña, M.; de Anta, R.C.; Bååth, E. Tolerance (PICT) of the Bacterial Communities to Copper in Vineyards Soils from Spain. J. Environ. Qual. 2007, 36, 1760–1764. [Google Scholar] [CrossRef]
- Bakshi, M.; Kumar, A. Copper-based nanoparticles in the soil-plant environment: Assessing their applications, interactions, fate and toxicity. Chemosphere 2021, 281, 130940. [Google Scholar] [CrossRef]
- He, S.; Feng, Y.; Ren, H.; Zhang, Y.; Gu, N.; Lin, X. The impact of iron oxide magnetic nanoparticles on the soil bacterial community. J. Soils Sediments 2011, 11, 1408–1417. [Google Scholar] [CrossRef]
- Xu, J.; Chen, Y.; Luo, J.; Xu, J.; Zhou, G.; Yu, Y.; Xue, L.; Yang, L.; He, S. Fe3O4 nanoparticles affect paddy soil microbial-driven carbon and nitrogen processes: Roles of surface coating and soil types. Environ. Sci. Nano 2022, 9, 2440–2452. [Google Scholar] [CrossRef]
- Ju-Nam, Y.; Lead, J.R. Manufactured nanoparticles: An overview of their chemistry, interactions and potential environmental implications. Sci. Total Environ. 2008, 400, 396–414. [Google Scholar] [CrossRef] [PubMed]
- Nowack, B. The behavior and effects of nanoparticles in the environment. Environ. Pollut. 2009, 157, 1063–1064. [Google Scholar] [CrossRef]
- Khalifa, M.; Fetyan, N.A.H.; Magid, M.S.A.; El-Sherry, N.I. Effectiveness of potassium silicate in suppression white rot disease and enhancement physiological resistance of onion plants, and its role on the soil microbial community. Middle East J. Agric. Res. 2017, 6, 376–394. [Google Scholar]
- Stone, M.M.; DeForest, J.L.; Plante, A.F. Changes in extracellular enzyme activity and microbial community structure with soil depth at the Luquillo Critical Zone Observatory. Soil Biol. Biochem. 2014, 75, 237–247. [Google Scholar] [CrossRef]
- Bandick, A.K.; Dick, R.P. Field management effects on soil enzyme activities. Soil Biol. Biochem. 1999, 31, 1471–1479. [Google Scholar] [CrossRef]
- Outten, F.W.; Theil, E.C. Iron-based redox switches in biology. Antioxid. Redox Signal. 2009, 11, 1029–1046. [Google Scholar] [CrossRef]
- Asadishad, B.; Chahal, S.; Akbari, A.; Cianciarelli, V.; Azodi, M.; Ghoshal, S.; Tufenkji, N. Amendment of Agricultural Soil with Metal Nanoparticles: Effects on Soil Enzyme Activity and Microbial Community Composition. Environ. Sci. Technol. 2018, 52, 1908–1918. [Google Scholar] [CrossRef]
- Jośko, I.; Oleszczuk, P.; Futa, B. The effect of inorganic nanoparticles (ZnO, Cr2O3, CuO and Ni) and their bulk counterparts on enzyme activities in different soils. Geoderma 2014, 232–234, 528–537. [Google Scholar] [CrossRef]
- Kukreti, B.; Sharma, A.; Chaudhary, P.; Agri, U.; Maithani, D. Influence of nanosilicon dioxide along with bioinoculants on Zea mays and its rhizospheric soil. 3 Biotech 2020, 10, 345. [Google Scholar] [CrossRef] [PubMed]
- Naushad, M. Surfactant assisted nano-composite cation exchanger: Development, characterization and applications for the removal of toxic Pb2+ from aqueous medium. Chem. Eng. J. 2014, 235, 100–108. [Google Scholar] [CrossRef]
- Zhao, W.; Liu, L.; Shen, Q.; Yang, J.; Han, X.; Tian, F.; Wu, J. Effects of water stress on photosynthesis, yield, and water use efficiency in winter wheat. Water 2020, 12, 2127. [Google Scholar] [CrossRef]
- Briat, J.F.; Duc, C.; Ravet, K.; Gaymard, F. Ferritins and iron storage in plants. Biochim. Biophys. Acta Gen. Subj. 2010, 1800, 806–814. [Google Scholar] [CrossRef]
- Aguirre, G.; Pilon, M. Copper delivery to chloroplast proteins and its regulation. Front. Plant Sci. 2016, 6, 1250. [Google Scholar] [CrossRef]
- Jurkow, R.; Sękara, A.; Pokluda, R.; Smoleń, S.; Kalisz, A. Biochemical response of oakleaf lettuce seedlings to different concentrations of some metal(oid) oxide nanoparticles. Agronomy 2020, 10, 997. [Google Scholar] [CrossRef]
- Badawy, A.A.; Abdelfattah, N.A.H.; Salem, S.S.; Awad, M.F.; Fouda, A. Efficacy assessment of biosynthesized copper oxide nanoparticles (Cuo-nps) on stored grain insects and their impacts on morphological and physiological traits of wheat (Triticum aestivum L.) plant. Biology 2021, 10, 233. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Description | Treatments | Description |
|---|---|---|---|
| T1 | 50 mg L−1 SiO2 soil drench | T12 | 200 mg L−1 SiO2 roots dipping + soil drench |
| T2 | 100 mg L−1 SiO2 soil drench | T13 | 50 mg L−1 CuO roots dipping + soil drench |
| T3 | 200 mg L−1 SiO2 soil drench | T14 | 100 mg L−1 CuO roots dipping + soil drench |
| T4 | 50 mg L−1 CuO soil drench | T15 | 200 mg L−1 CuO roots dipping + soil drench |
| T5 | 100 mg L−1 CuO soil drench | T16 | 50 mg L−1 Fe2O3 roots dipping + soil drench |
| T6 | 200 mg L−1 CuO soil drench | T17 | 100 mg L−1Fe2O3 roots dipping + soil drench |
| T7 | 50 mg L−1 Fe2O3 soil drench | T18 | 200 mg L−1 Fe2O3 roots dipping + soil drench |
| T8 | 100 mg L−1 Fe2O3 soil drench | T19 | Control (plants inoculated with fungal pathogen) |
| T9 | 200 mg L−1 Fe2O3 soil drench | T20 | Control (plants inoculated with fungal pathogen) + Rizolex® 2.5 g L−1 |
| T10 | 50 mg L−1 SiO2 roots dipping + soil drench | T21 | Control (healthy plants) |
| T11 | 100 mg L−1 SiO2 roots dipping + soil drench |
| El Mansoryh | Nahia | Total | ||||
|---|---|---|---|---|---|---|
| Isolates | No. | Frequency % | No. | Frequency % | No. | Frequency % |
| Rhizoctonia solani | 12 | 70.53% | 9 | 60.00 | 21 | 63.64% |
| Fusarium solani | 3 | 17.91% | 4 | 26.67 | 7 | 21.21% |
| Fusarium oxysporum | 2 | 11.56% | 2 | 13.33 | 5 | 15.15% |
| Total | 17 | 100% | 15 | 100 | 33 | 100% |
| LSD at 0.05% | 20.06 | 12.89 | ||||
| Isolates | Disease Incidence % | Disease Severity % |
|---|---|---|
| Rhizoctonia solani | 83.33 | 64.77 |
| Fusarium solani | 33.33 | 3.24 |
| Fusarium oxysporum | 16.67 | 1.48 |
| Control | 0.00 | 0.00 |
| LSD at 0.05% | 21.25 | 2.74 |
| Treatments | Concentration ppm | Linear Growth | Reduction % |
|---|---|---|---|
| SiO2 | 50 | 9 a | 0.0 |
| 100 | 9 a | 0.0 | |
| 200 | 9 a | 0.0 | |
| CuO | 50 | 8.26 ab | 8.22 |
| 100 | 8.26 ab | 8.22 | |
| 200 | 7.93 bc | 11.89 | |
| Fe2O3 | 50 | 7.46 bc | 17.11 |
| 100 | 7.36 c | 18.22 | |
| 200 | 6.9 c | 23.33 | |
| Rizolex | 2.5 g/L | 4.5 d | 50 |
| Control | 0 | 9.0 a | 0 |
| Treatments | Disease Incidence % | Disease Severity % | Fresh Weight (g) | Dry Weight (g) |
|---|---|---|---|---|
| T1 | 50.0 cd | 36.50 e | 33.7 | 10.3 |
| T2 | 50.0 cd | 36.18 e | 24.5 | 8.6 |
| T3 | 50.0 cd | 34.11 f | 58.4 | 14.8 |
| T4 | 83.3 b | 82.14 a | 34.4 | 7.9 |
| T5 | 83.3 b | 80.12 b | 40.0 | 12.3 |
| T6 | 33.3 d | 27.34 g | 61.4 | 13.9 |
| T7 | 50.0 cd | 55.32 d | 31.4 | 9.6 |
| T8 | 66.7 bc | 25.43 h | 50.3 | 12.1 |
| T9 | 33.3 d | 24.84 h | 62.0 | 18.8 |
| T10 | 33.3 b | 36.15 b | 38.5 | 10.2 |
| T11 | 16.7 b | 16.54 c | 32.7 | 11.0 |
| T12 | 16.7 b | 3.25 g | 50.6 | 16.2 |
| T13 | 33.3 b | 15.50 d | 36.8 | 8.3 |
| T14 | 33.3 b | 13.42 ef | 38.8 | 7.3 |
| T15 | 0.00 c | 0.00 h | 38.0 | 8.7 |
| T16 | 16.7 b | 12.32 f | 44.6 | 10.4 |
| T17 | 16.7 b | 14.23 de | 38.9 | 9.7 |
| T18 | 0.0 c | 0.00 h | 58.7 | 16.9 |
| T19 | 100.0 a | 78.43 c | 37.5 | 9.7 |
| T20 | 33.3 d | 18.32 i | 21.2 | 9.4 |
| T21 | 0.00 e | 0.00 j | 41.9 | 6.9 |
| LSD | 19.19 | 1.03 | 5.371 | 2.645 |
| Treatments | T.B.C | T.F | T.Actin | PSB | SSB | Azotobacter sp. | Azospirillum sp. |
|---|---|---|---|---|---|---|---|
| CFU × 106 | CFU × 104 | CFU × 103 | CFU × 104 | CFU × 104 | CFU × 104 | CFU × 104 | |
| T1 | 9.47 | 10.33 | 5.67 | 10.33 | 0.84 | 1.05 | 0.37 |
| T2 | 11.33 | 11.00 | 7.00 | 8.37 | 0.64 | 0.87 | 0.34 |
| T3 | 12.87 | 11.67 | 7.33 | 10.27 | 0.84 | 1.13 | 0.33 |
| T4 | 11.67 | 7.00 | 6.67 | 9.40 | 0.79 | 0.84 | 0.35 |
| T5 | 14.33 | 4.00 | 6.00 | 9.14 | 0.90 | 0.86 | 0.31 |
| T6 | 9.00 | 5.33 | 7.00 | 5.75 | 0.63 | 0.73 | 0.17 |
| T7 | 20.33 | 14.33 | 10.33 | 8.70 | 0.85 | 0.76 | 0.30 |
| T8 | 24.67 | 10.00 | 9.00 | 8.27 | 0.82 | 0.91 | 0.36 |
| T9 | 17.33 | 8.00 | 13.33 | 11.20 | 0.87 | 0.71 | 0.21 |
| T10 | 13.27 | 6.67 | 4.33 | 9.77 | 0.82 | 0.74 | 0.34 |
| T11 | 12.17 | 10.00 | 5.67 | 7.77 | 0.71 | 0.90 | 0.06 |
| T12 | 18.33 | 8.67 | 5.67 | 10.93 | 0.58 | 0.83 | 0.12 |
| T13 | 19.64 | 6.67 | 3.33 | 8.07 | 0.87 | 0.87 | 0.42 |
| T14 | 20.20 | 8.33 | 3.33 | 8.64 | 0.96 | 0.96 | 0.14 |
| T15 | 11.40 | 8.67 | 2.67 | 4.73 | 0.47 | 0.70 | 0.08 |
| T16 | 10.27 | 5.00 | 5.67 | 8.97 | 0.86 | 0.88 | 0.27 |
| T17 | 9.87 | 5.00 | 3.67 | 7.33 | 0.87 | 0.69 | 0.24 |
| T18 | 8.43 | 3.33 | 2.00 | 5.13 | 0.73 | 0.90 | 0.30 |
| T19 | 13.87 | 14.67 | 3.00 | 5.13 | 0.58 | 0.81 | 0.14 |
| T20 | 12.13 | 5.67 | 3.67 | 5.03 | 0.91 | 0.95 | 0.12 |
| T21 | 10.33 | 10.67 | 6.00 | 8.83 | 0.99 | 0.71 | 0.44 |
| LSD | 2.324 | 2.612 | 1.729 | 2.463 | 0.236 | 0.142 | 0.111 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fetyan, N.A.H.; Essa, T.A.; Salem, T.M.; Taha, A.A.; Elgobashy, S.F.; Tharwat, N.A.; Elsakhawy, T. Promising Eco-Friendly Nanoparticles for Managing Bottom Rot Disease in Lettuce (Lactuca sativa var. longifolia). Microbiol. Res. 2024, 15, 196-212. https://doi.org/10.3390/microbiolres15010014
Fetyan NAH, Essa TA, Salem TM, Taha AA, Elgobashy SF, Tharwat NA, Elsakhawy T. Promising Eco-Friendly Nanoparticles for Managing Bottom Rot Disease in Lettuce (Lactuca sativa var. longifolia). Microbiology Research. 2024; 15(1):196-212. https://doi.org/10.3390/microbiolres15010014
Chicago/Turabian StyleFetyan, Nashwa A. H., Tarek A. Essa, Tamer M. Salem, Ahmed Aboueloyoun Taha, Samah Fawzy Elgobashy, Nagwa A. Tharwat, and Tamer Elsakhawy. 2024. "Promising Eco-Friendly Nanoparticles for Managing Bottom Rot Disease in Lettuce (Lactuca sativa var. longifolia)" Microbiology Research 15, no. 1: 196-212. https://doi.org/10.3390/microbiolres15010014
APA StyleFetyan, N. A. H., Essa, T. A., Salem, T. M., Taha, A. A., Elgobashy, S. F., Tharwat, N. A., & Elsakhawy, T. (2024). Promising Eco-Friendly Nanoparticles for Managing Bottom Rot Disease in Lettuce (Lactuca sativa var. longifolia). Microbiology Research, 15(1), 196-212. https://doi.org/10.3390/microbiolres15010014