
Transcription Factors Mcm1 and Sfp1 May Affect [PSI+] Prion Phenotype by Altering the Expression of the SUP35 Gene
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plasmids
2.2. Strains
2.3. qPCR
2.4. Protein Analysis
2.5. Fluorescence Microscopy
2.6. Bioinformatic Analysis
3. Results
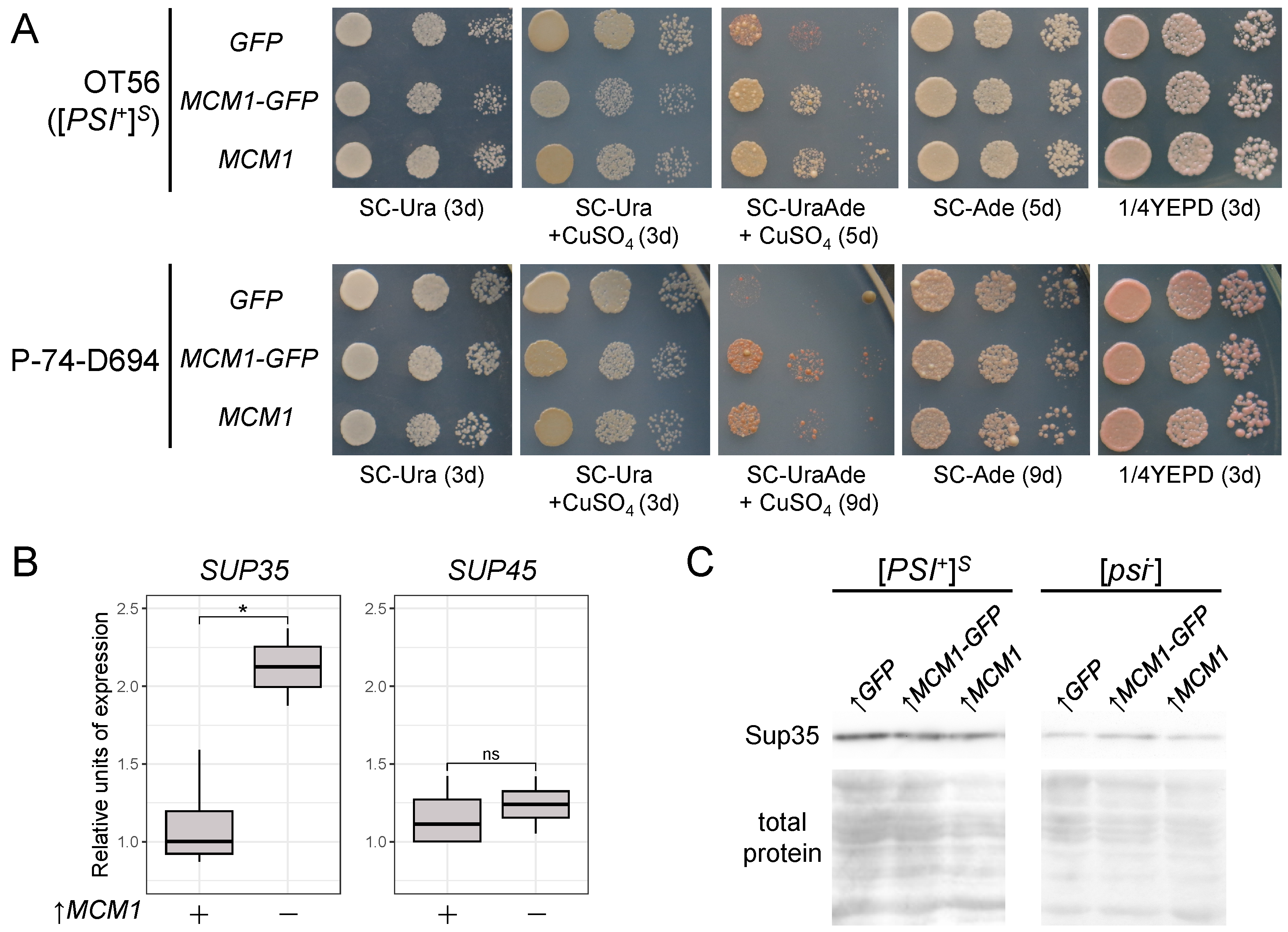
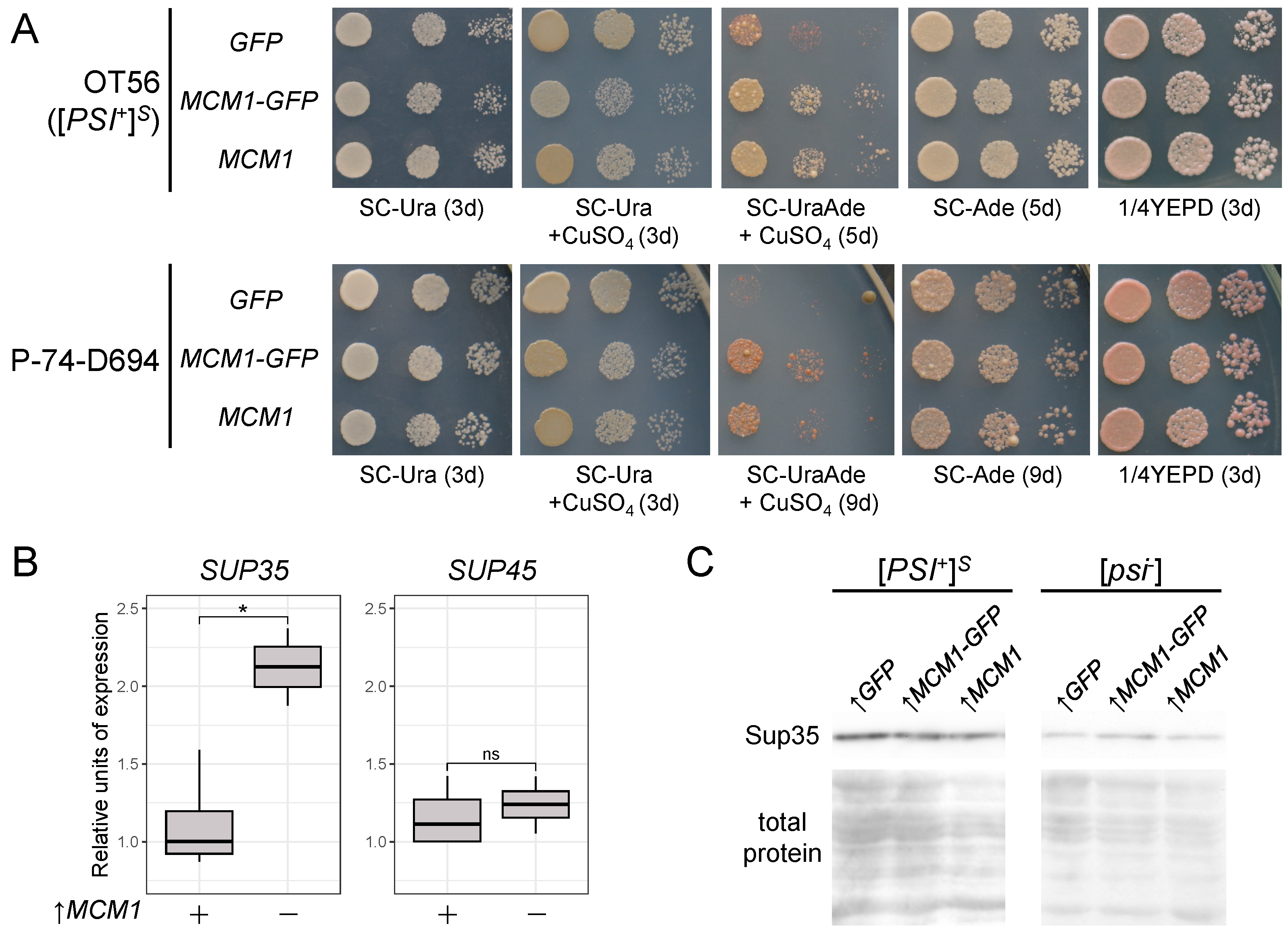
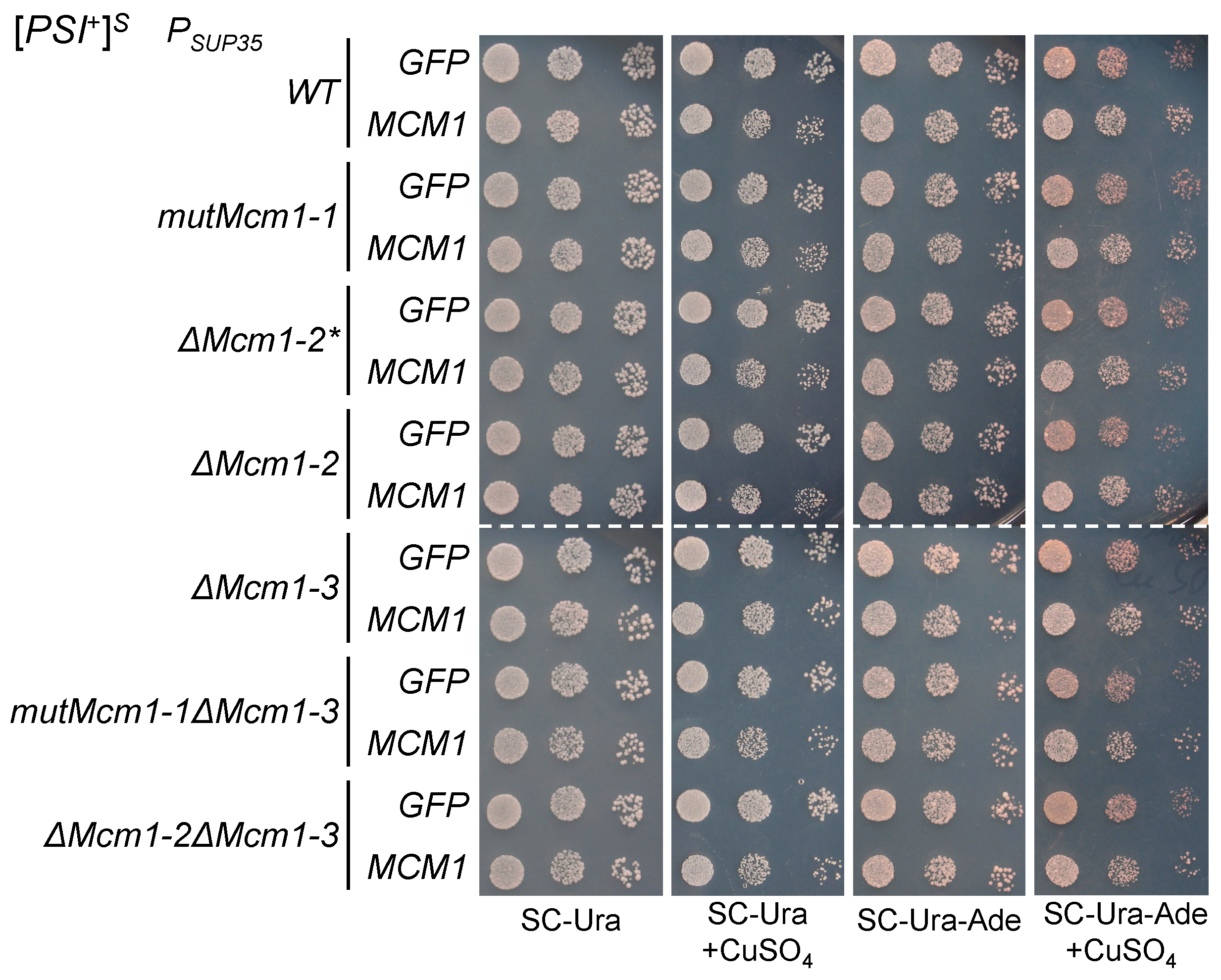
3.1. Transient Overexpression of MCM1 Enhances Nonsense Suppression in [PSI+] Strains
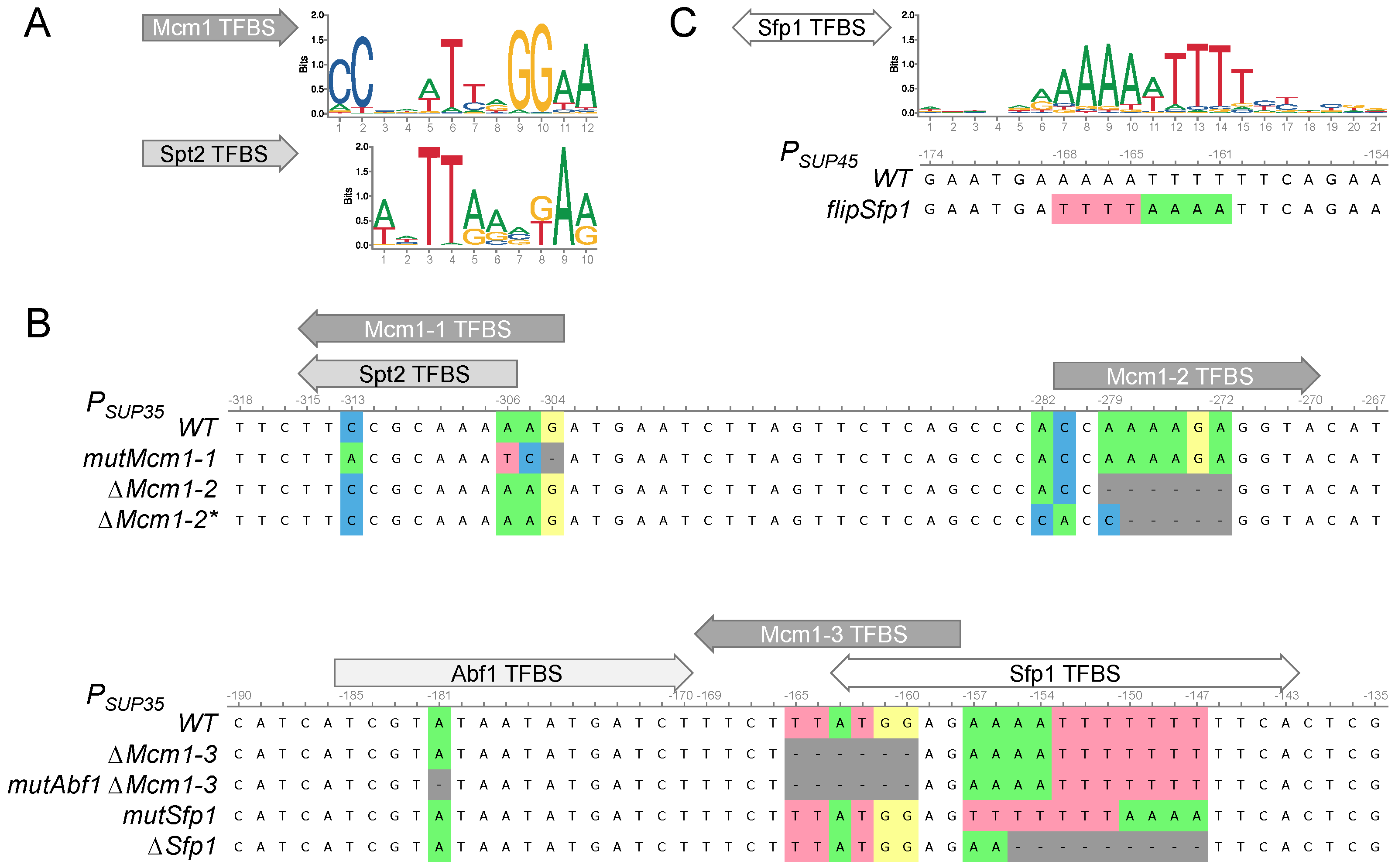
3.2. Search for the Potential Mcm1 and Sfp1 Binding Sites in the SUP35 and SUP45 Promoter Regions
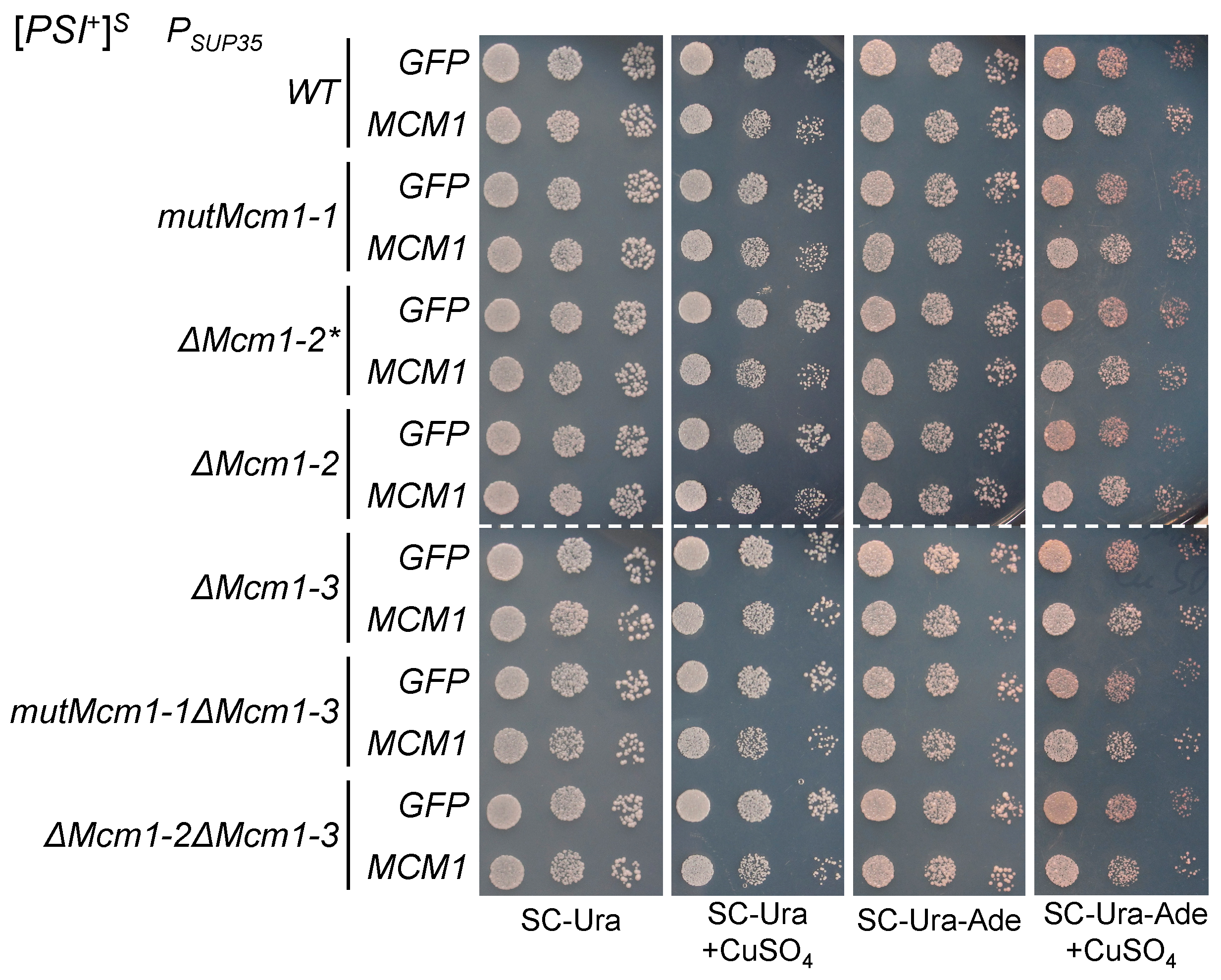
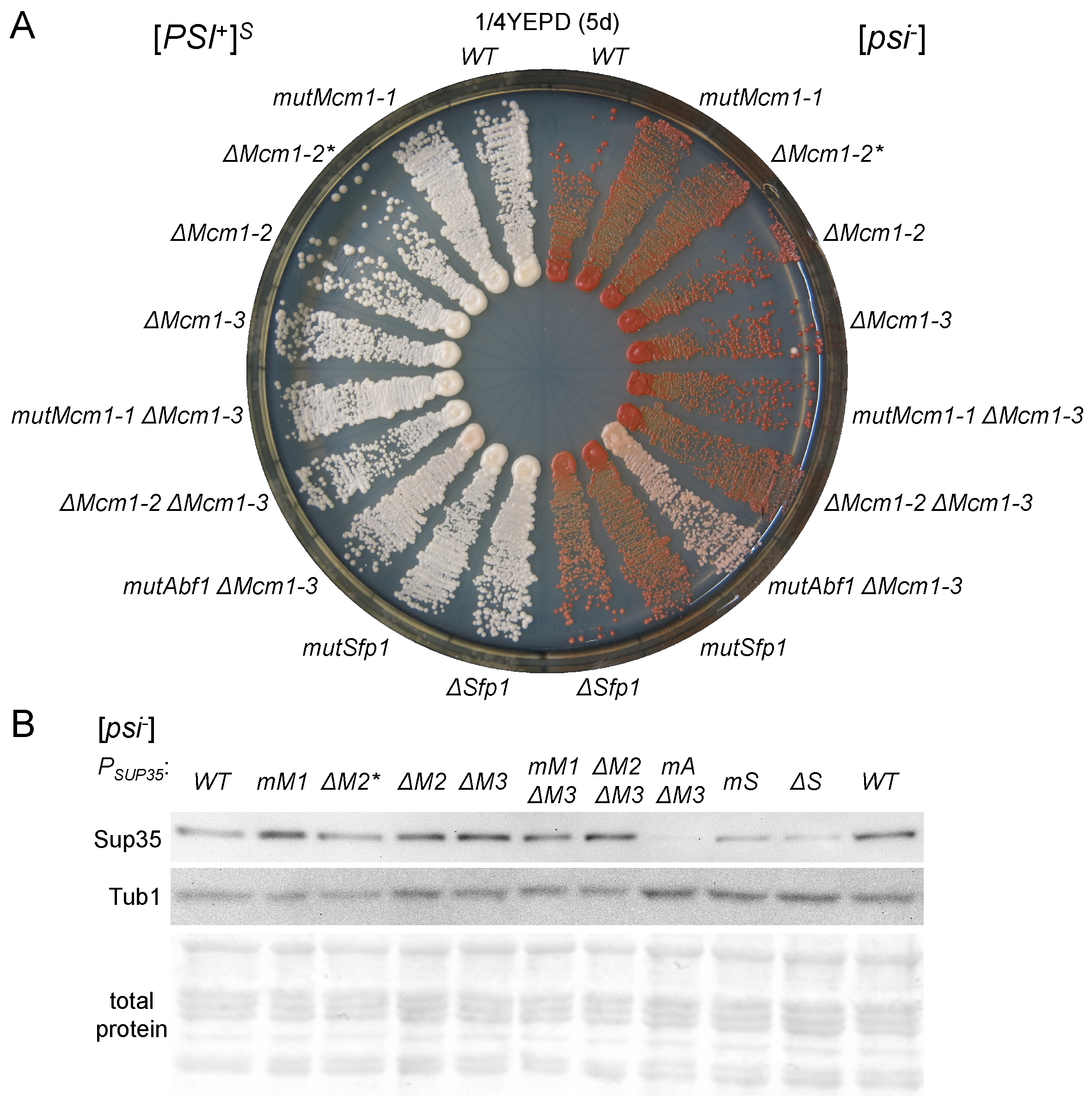
3.3. The Potential Mcm1 and Sfp1 Binding Sites in the SUP35 Promoter Regions Do Not Significantly Influence Nonsense Suppression
3.4. The Effects of Transient MCM1 Overexpression Do Not Depend on the Potential Mcm1 Binding Sites
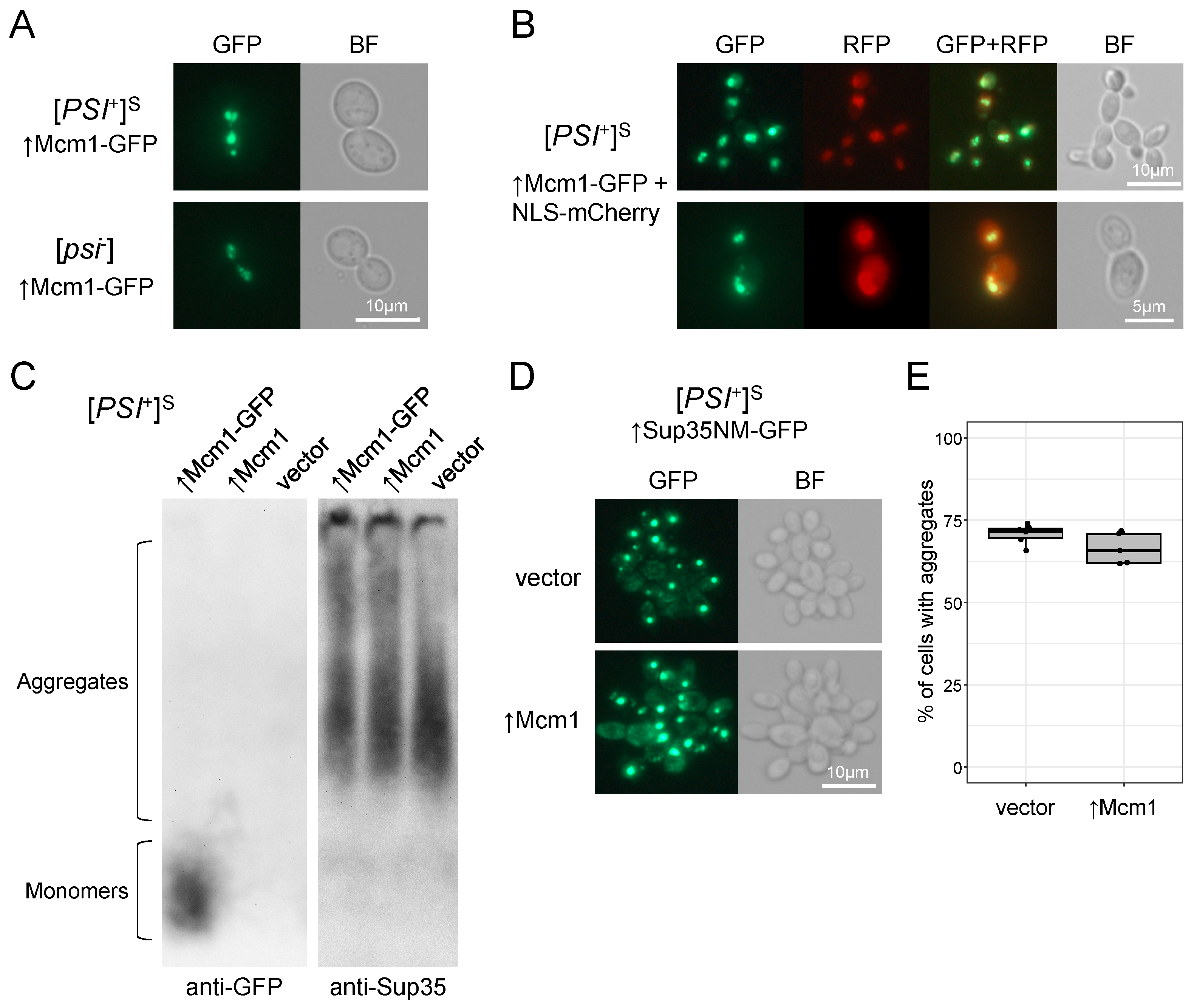
3.5. Mcm1 Does Not Form Detergent-Resistant Aggregates and Does Not Affect Sup35 Aggregation
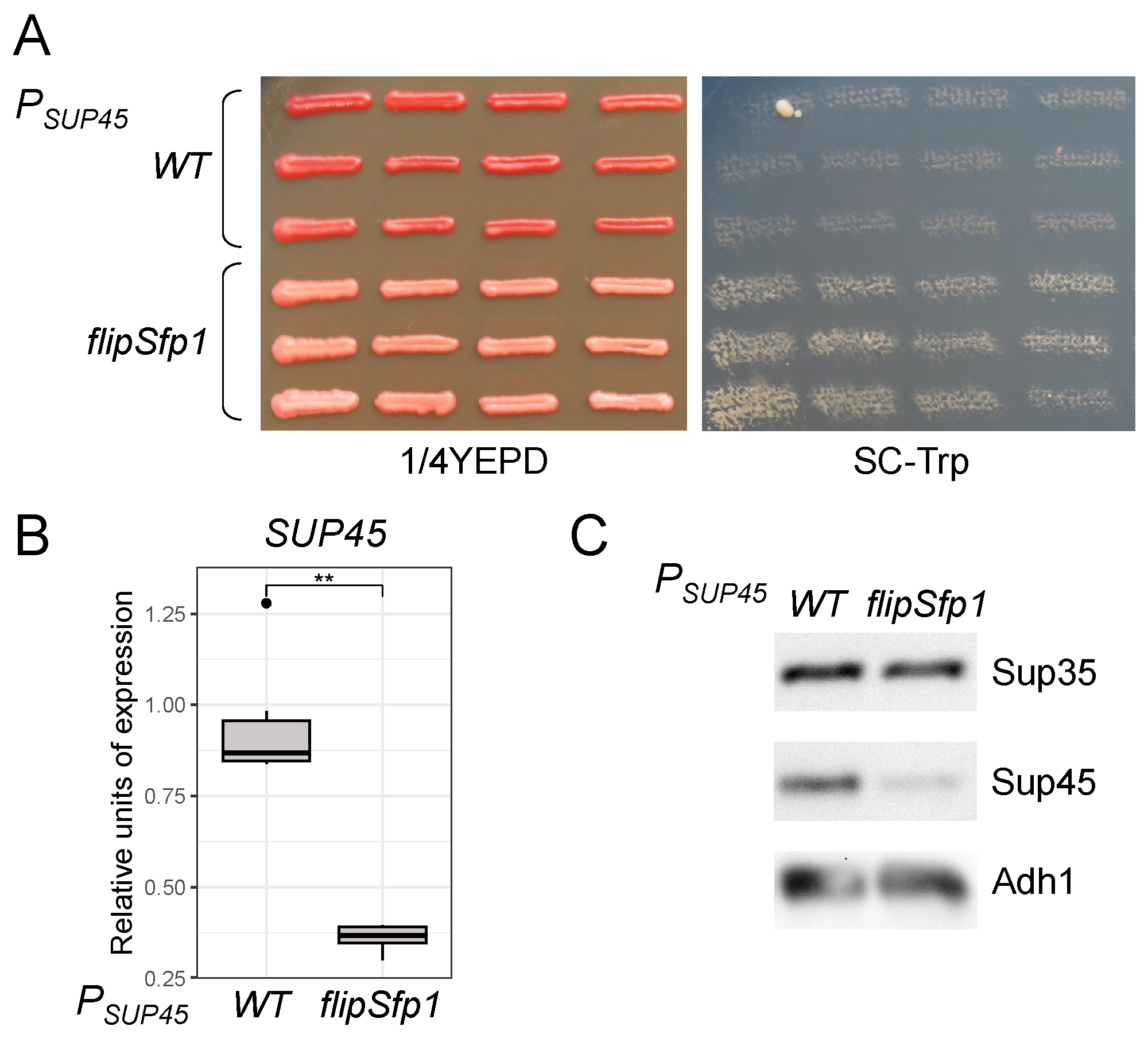
3.6. The Potential Sfp1 Binding Site in the SUP45 Promoter Affects Its Expression and Nonsense Suppression
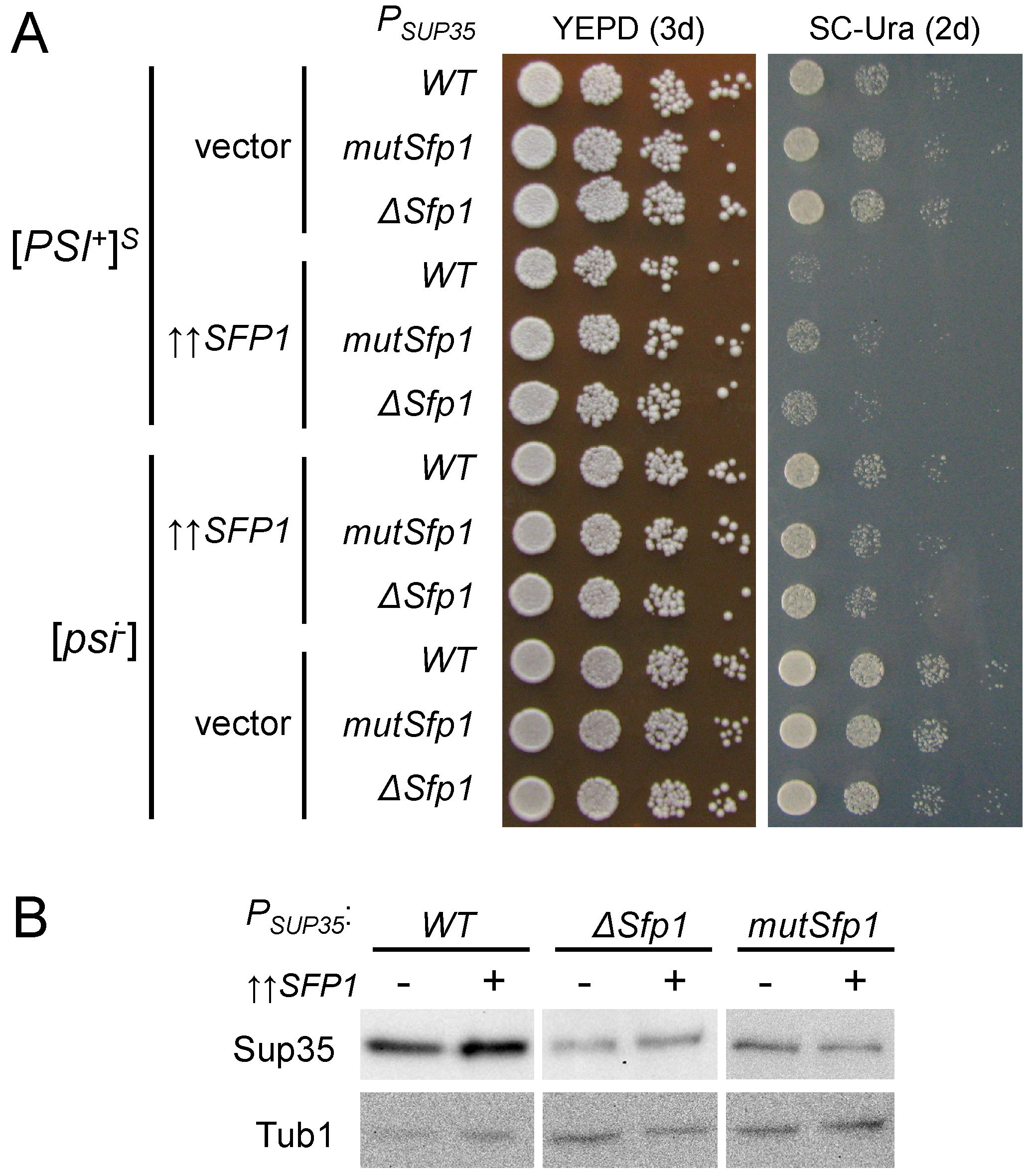
3.7. The Potential Sfp1 Binding Site in the SUP35 Promoter Is Important for [PSI+] Prion Toxicity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| aa | amino acids |
| bp | base pair |
| GFP | green fluorescent protein |
| ChIP | chromatin immunoprecipitation |
| ChEC | chromatin endogenous cleavage |
| 5-FOA | 5-fluoroorotic acid |
| TFBS | transcription factor binding site |
| NLS | nuclear localization signal |
| PQC | protein quality control |
| RP | ribosomal protein |
| RiBi | ribosome biogenesis |
| PCR | polymerase chain reaction |
| qPCR | quantitative polymerase chain reaction |
| SDS-PAGE | sodium dodecyl sulfate–polyacrylamide gel electrophoresis |
| SDD-AGE | semi-denaturating detergent agarose gel electrophoresis |
References
- Liebman, S.W.; Chernoff, Y.O. Prions in yeast. Genetics 2012, 191, 1041–1072. [Google Scholar] [CrossRef] [PubMed]
- Trubitsina, N.; Zemlyanko, O.; Moskalenko, S.; Zhouravleva, G. From past to future: Suppressor mutations in yeast genes encoding translation termination factors. Biol. Commun. 2019, 64, 89–109. [Google Scholar] [CrossRef]
- Stansfield, I.; Akhmaloka, L.E.; Tuite, M.F. Depletion in the levels of the release factor eRF1 causes a reduction in the efficiency of translation termination in yeast. Mol. Microbiol. 1996, 20, 1135–1143. [Google Scholar] [CrossRef]
- Valouev, I.A.; Kushnirov, V.V.; Ter-Avanesyan, M.D. Yeast polypeptide chain release factors eRF1 and eRF3 are involved in cytoskeleton organization and cell cycle regulation. Cell Motil. Cytoskelet. 2002, 52, 161–173. [Google Scholar] [CrossRef] [PubMed]
- Tuite, M.F. Yeast prions and their prion-forming domain. Cell 2000, 191, 289–292. [Google Scholar] [CrossRef] [PubMed]
- Kushnirov, V.V.; Dergalev, A.A.; Alieva, M.K.; Alexandrov, A.I. Structural bases of prion variation in yeast. Int. J. Mol. Sci. 2022, 23, 5738. [Google Scholar] [CrossRef] [PubMed]
- Franzmann, T.M.; Jahnel, M.; Pozniakovsky, A.; Mahamid, J.; Holehouse, A.S.; Nüske, E.; Richter, D.; Baumeister, W.; Grill, S.W.; Pappu, R.V.; et al. Phase separation of a yeast prion protein promotes cellular fitness. Science 2018, 359, eaao5654. [Google Scholar] [CrossRef] [PubMed]
- Zhouravleva, G.; Frolova, L.; Le Goff, X.; Le Guellec, R.; Inge-Vechtomov, S.; Kisselev, L.; Philippe, M. Termination of translation in eukaryotes is governed by two interacting polypeptide chain release factors, eRF1 and eRF3. EMBO J. 1995, 14, 4065–4072. [Google Scholar] [CrossRef] [PubMed]
- Stansfield, I.; Jones, K.M.; Kushnirov, V.V.; Dagkesamanskaya, A.R.; Poznyakovski, A.I.; Paushkin, S.V.; Tuite, M.F. The products of the SUP45 (eRF1) and SUP35 genes interact to mediate translation termination in Saccharomyces cerevisiae. EMBO J. 1995, 14, 4365–4373. [Google Scholar] [CrossRef]
- Derkatch, I.L.; Bradley, M.E.; Hong, J.Y.; Liebman, S.W. Prions affect the appearance of other prions: The story of [PIN+]. Cell 2001, 106, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Nizhnikov, A.A.; Magomedova, Z.M.; Rubel, A.A.; Kondrashkina, A.M.; Inge-Vechtomov, S.G.; Galkin, A.P. [NSI+] determinant has a pleiotropic phenotypic manifestation that is modulated by SUP35, SUP45, and VTS1 genes. Curr. Genet. 2012, 58, 35–47. [Google Scholar] [CrossRef] [PubMed]
- Nizhnikov, A.A.; Ryzhova, T.A.; Volkov, K.V.; Zadorsky, S.P.; Sopova, J.V.; Inge-Vechtomov, S.G.; Galkin, A.P. Interaction of prions causes heritable traits in Saccharomyces cerevisiae. PLoS Genet. 2016, 12, e1006504. [Google Scholar] [CrossRef] [PubMed]
- Alberti, S.; Halfmann, R.; King, O.; Kapila, A.; Lindquist, S. A systematic survey identifies prions and illuminates sequence features of prionogenic proteins. Cell 2009, 137, 146–158. [Google Scholar] [CrossRef] [PubMed]
- Matiiv, A.B.; Trubitsina, N.P.; Matveenko, A.G.; Barbitoff, Y.A.; Zhouravleva, G.A.; Bondarev, S.A. Amyloid and amyloid-like aggregates: Diversity and the term crisis. Biochemistry 2020, 85, 1011–1034. [Google Scholar] [CrossRef] [PubMed]
- Zhouravleva, G.A.; Bondarev, S.A.; Trubitsina, N.P. How big is the yeast prion universe? Int. J. Mol. Sci. 2023, 24, 11651. [Google Scholar] [CrossRef] [PubMed]
- Kiktev, D.A.; Chernoff, Y.O.; Archipenko, A.V.; Zhouravleva, G.A. Identification of genes influencing synthetic lethality of genetic and epigenetic alterations in translation termination factors in yeast. Biochem. Biophys. 2011, 438, 117–119. [Google Scholar] [CrossRef] [PubMed]
- Matveenko, A.G.; Zemlyanko, O.M.; Zhouravleva, G.A. Identification of Saccharomyces cerevisiae genes leading to synthetic lethality of prion [PSI+] with SUP45 mutations. Mol. Biol. 2013, 47, 530–537. [Google Scholar] [CrossRef]
- Matveenko, A.G.; Belousov, M.V.; Bondarev, S.A.; Moskalenko, S.E.; Zhouravleva, G.A. Identification of new genes that affect [PSI+] prion toxicity in Saccharomyces cerevisiae yeast. Mol. Biol. 2016, 50, 710–718. [Google Scholar] [CrossRef]
- Sikorski, R.S.; Hieter, P. A system of shuttle vectors and yeast host strains designed for efficient manipulation of DNA in Saccharomyces cerevisiae. Genetics 1989, 122, 19–27. [Google Scholar] [CrossRef]
- Christianson, T.W.; Sikorski, R.S.; Dante, M.; Shero, J.H.; Hieter, P. Multifunctional yeast high-copy-number shuttle vectors. Gene 1992, 110, 119–122. [Google Scholar] [CrossRef] [PubMed]
- Rogoza, T.; Goginashvili, A.; Rodionova, S.; Ivanov, M.; Viktorovskaya, O.; Rubel, A.; Volkov, K.; Mironova, L. Non-Mendelian determinant [ISP+] in yeast is a nuclear-residing prion form of the global transcriptional regulator Sfp1. Proc. Natl. Acad. Sci. USA 2010, 107, 10573–10577. [Google Scholar] [CrossRef] [PubMed]
- Nizhnikov, A.A.; Kondrashkina, A.M.; Antonets, K.S.; Galkin, A.P. Overexpression of genes encoding asparagine-glutamine-rich transcriptional factors causes nonsense suppression in Saccharomyces cerevisiae. Russ. J. Genet. Appl. Res. 2014, 4, 122–130. [Google Scholar] [CrossRef]
- Serio, T.R.; Cashikar, A.G.; Moslehi, J.J.; Kowal, A.S.; Lindquist, S.L. Yeast prion [psi+] and its determinant, Sup35p. Methods Enzymol. 1999, 309, 649–673. [Google Scholar] [PubMed]
- Malinovska, L.; Kroschwald, S.; Munder, M.C.; Richter, D.; Alberti, S. Molecular chaperones and stress-inducible protein-sorting factors coordinate the spatiotemporal distribution of protein aggregates. Mol. Biol. Cell 2012, 23, 3041–3056. [Google Scholar] [CrossRef] [PubMed]
- Volkov, K.V.; Aksenova, A.Y.; Soom, M.J.; Osipov, K.V.; Svitin, A.V.; Kurischko, C.; Shkundina, I.S.; Ter-Avanesyan, M.D.; Inge-Vechtomov, S.G.; Mironova, L.N. Novel non-Mendelian determinant involved in the control of translation accuracy in Saccharomyces cerevisiae. Genetics 2002, 160, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Moskalenko, S.E.; Chabelskaya, S.V.; Inge-Vechtomov, S.G.; Philippe, M.; Zhouravleva, G.A. Viable nonsense mutants for the essential gene SUP45 of Saccharomyces cerevisiae. BMC Mol. Biol. 2003, 4, 2. [Google Scholar] [CrossRef]
- Danilov, L.G.; Matveenko, A.G.; Ryzhkova, V.E.; Belousov, M.V.; Poleshchuk, O.I.; Likholetova, D.V.; Sokolov, P.A.; Kasyanenko, N.A.; Kajava, A.V.; Zhouravleva, G.A.; et al. Design of a new [PSI+]-no-more mutation in SUP35 with strong inhibitory effect on the [PSI+] prion propagation. Front. Mol. Neurosci. 2019, 12, 274. [Google Scholar] [CrossRef] [PubMed]
- Chernoff, Y.O.; Lindquist, S.L.; Ono, B.; Inge-Vechtomov, S.G.; Liebman, S.W. Role of the chaperone protein Hsp104 in propagation of the yeast prion-like factor [psi+]. Science 1995, 268, 880–884. [Google Scholar] [CrossRef] [PubMed]
- Derkatch, I.L.; Bradley, M.E.; Zhou, P.; Chernoff, Y.O.; Liebman, S.W. Genetic and environmental factors affecting the de novo appearance of the [PSI+] prion in Saccharomyces cerevisiae. Genetics 1997, 147, 507–519. [Google Scholar] [CrossRef] [PubMed]
- Newnam, G.P.; Wegrzyn, R.D.; Lindquist, S.L.; Chernoff, Y.O. Antagonistic interactions between yeast chaperones Hsp104 and Hsp70 in prion curing. Mol. Cell. Biol. 1999, 19, 1325–1333. [Google Scholar] [CrossRef]
- Drozdova, P.B.; Tarasov, O.V.; Matveenko, A.G.; Radchenko, E.A.; Sopova, J.V.; Polev, D.E.; Inge-Vechtomov, S.G.; Dobrynin, P.V. Genome sequencing and comparative analysis of Saccharomyces cerevisiae strains of the Peterhof. PLoS ONE 2016, 11, e0154722. [Google Scholar] [CrossRef] [PubMed]
- Matveenko, A.G.; Drozdova, P.B.; Belousov, M.V.; Moskalenko, S.E.; Bondarev, S.A.; Barbitoff, Y.A.; Nizhnikov, A.A.; Zhouravleva, G.A. SFP1-mediated prion-dependent lethality is caused by increased Sup35 aggregation and alleviated by Sis1. Genes Cells 2016, 21, 1290–1308. [Google Scholar] [CrossRef] [PubMed]
- Chernoff, Y.O.; Galkin, A.P.; Lewitin, E.; Chernova, T.A.; Newnam, G.P.; Belenkiy, S.M. Evolutionary conservation of prion-forming abilities of the yeast Sup35 protein. Mol. Microbiol. 2000, 35, 865–876. [Google Scholar] [CrossRef] [PubMed]
- Matveenko, A.G.; Drozdova, P.B.; Moskalenko, S.E.; Tarasov, O.V.; Zhouravleva, G.A. Whole genome sequencing data and analyses of the underlying SUP35 transcriptional regulation for a Saccharomyces cerevisiae nonsense suppressor mutant. Data Brief 2019, 23, 103694. [Google Scholar] [CrossRef] [PubMed]
- Barbitoff, Y.A.; Matveenko, A.G.; Matiiv, A.B.; Maksiutenko, E.M.; Moskalenko, S.E.; Drozdova, P.B.; Polev, D.E.; Beliavskaia, A.Y.; Danilov, L.G.; Predeus, A.V.; et al. Chromosome-level genome assembly and structural variant analysis of two laboratory yeast strains from the Peterhof Genetic Collection lineage. G3 2021, 11, jkab029. [Google Scholar] [CrossRef]
- Matveenko, A.G.; Ryzhkova, V.E.; Zaytseva, N.A.; Danilov, L.G.; Mikhailichenko, A.S.; Barbitoff, Y.A.; Zhouravleva, G.A. Processing of fluorescent proteins may prevent detection of prion particles in [PSI+] cells. Biology 2022, 11, 1688. [Google Scholar] [CrossRef] [PubMed]
- Maksiutenko, E.M.; Barbitoff, Y.A.; Matveenko, A.G.; Moskalenko, S.E.; Zhouravleva, G.A. Gene amplification as a mechanism of yeast adaptation to nonsense mutations in release factor genes. Genes 2021, 12, 2019. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Fritsch, E.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]
- Kaiser, C.; Michaelis, S.; Mitchell, A. Methods in Yeast Genetics; Number 316; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1994; p. 234. [Google Scholar]
- Eaglestone, S.S.; Ruddock, L.W.; Cox, B.S.; Tuite, M.F. Guanidine hydrochloride blocks a critical step in the propagation of the prion-like determinant [PSI+] of Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2000, 97, 240–244. [Google Scholar] [CrossRef] [PubMed]
- Gietz, R.D.; Woods, R.A. Transformation of yeast by lithium acetate/single-stranded carrier DNA/polyethylene glycol method. Methods Enzymol. 2002, 350, 87–96. [Google Scholar] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(−ΔΔ C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Lei, J.; Yang, H.; Xu, K.; Wang, R.; Zhang, Z. An improved method for whole protein extraction from yeast Saccharomyces cerevisiae. Yeast 2011, 28, 795–798. [Google Scholar] [CrossRef] [PubMed]
- Kushnirov, V.V.; Alexandrov, I.M.; Mitkevich, O.V.; Shkundina, I.S.; Ter-Avanesyan, M.D. Purification and analysis of prion and amyloid aggregates. Methods 2006, 39, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Halfmann, R.; Lindquist, S. Screening for amyloid aggregation by semi-denaturing detergent-agarose gel electrophoresis. J. Vis. Exp. JoVE 2008, 17, 838. [Google Scholar]
- Chabelskaya, S.; Kiktev, D.; Inge-Vechtomov, S.; Philippe, M.; Zhouravleva, G. Nonsense mutations in the essential gene SUP35 of Saccharomyces cerevisiae are non-lethal. Mol. Genet. Genom. MGG 2004, 272, 297–307. [Google Scholar] [CrossRef] [PubMed]
- Kiktev, D.; Moskalenko, S.; Murina, O.; Baudin-Baillieu, A.; Rousset, J.P.; Zhouravleva, G. The paradox of viable sup45 STOP mutations: A necessary equilibrium between translational readthrough, activity and stability of the protein. Mol. Genet. Genom. MGG 2009, 282, 83–96. [Google Scholar] [CrossRef]
- Kwon, A.T.; Arenillas, D.J.; Hunt, R.W.; Wasserman, W.W. oPOSSUM-3: Advanced analysis of regulatory motif over-representation across genes or ChIP-Seq datasets. G3 2012, 2, 987–1002. [Google Scholar] [CrossRef] [PubMed]
- Castro-Mondragon, J.A.; Riudavets-Puig, R.; Rauluseviciute, I.; Lemma, R.B.; Turchi, L.; Blanc-Mathieu, R.; Lucas, J.; Boddie, P.; Khan, A.; Pérez, N.M.; et al. JASPAR 2022: The 9th release of the open-access database of transcription factor binding profiles. Nucleic Acids Res. 2022, 50, D165–D173. [Google Scholar] [CrossRef] [PubMed]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Venters, B.J.; Wachi, S.; Mavrich, T.N.; Andersen, B.E.; Jena, P.; Sinnamon, A.J.; Jain, P.; Rolleri, N.S.; Jiang, C.; Hemeryck-Walsh, C.; et al. A comprehensive genomic binding map of gene and chromatin regulatory proteins in Saccharomyces. Mol. Cell 2011, 41, 480–492. [Google Scholar] [CrossRef] [PubMed]
- Michelitsch, M.D.; Weissman, J.S. A census of glutamine/asparagine-rich regions: Implications for their conserved function and the prediction of novel prions. Proc. Natl. Acad. Sci. USA 2000, 97, 11910–11915. [Google Scholar] [CrossRef] [PubMed]
- Bondarev, S.A.; Antonets, K.S.; Kajava, A.V.; Nizhnikov, A.A.; Zhouravleva, G.A. Protein Co-Aggregation Related to Amyloids: Methods of Investigation, Diversity, and Classification. Int. J. Mol. Sci. 2018, 19, 2292. [Google Scholar] [CrossRef] [PubMed]
- Chakrabortee, S.; Byers, J.S.; Jones, S.; Garcia, D.M.; Bhullar, B.; Chang, A.; She, R.; Lee, L.; Fremin, B.; Lindquist, S.; et al. Intrinsically Disordered Proteins Drive Emergence and Inheritance of Biological Traits. Cell 2016, 167, 369–381. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.; Richmond, T.J. Crystal structure of the yeast MATalpha2/MCM1/DNA ternary complex. Nature 1998, 391, 660–666. [Google Scholar] [CrossRef] [PubMed]
- Tye, B.K.; Chang, V.K. Dual functional regulators coordinate DNA replication and gene expression in proliferating cells. Front. Biosci. 2004, 9, 2548–2555. [Google Scholar] [CrossRef] [PubMed]
- Wittenberg, C.; Reed, S.I. Cell cycle-dependent transcription in yeast: Promoters, transcription factors, and transcriptomes. Oncogene 2005, 24, 2746–2755. [Google Scholar] [CrossRef] [PubMed]
- Dastidar, R.G.; Hooda, J.; Shah, A.; Cao, T.M.; Henke, R.M.; Zhang, L. The nuclear localization of SWI/SNF proteins is subjected to oxygen regulation. Cell Biosci. 2012, 2, 30. [Google Scholar] [CrossRef] [PubMed]
- Dolan, J.W.; Fields, S. Cell-type-specific transcription in yeast. Biochim. Biophys. Acta Gene Struct. Expr. 1991, 1088, 155–169. [Google Scholar] [CrossRef] [PubMed]
- Darieva, Z.; Clancy, A.; Bulmer, R.; Williams, E.; Pic-Taylor, A.; Morgan, B.A.; Sharrocks, A.D. A competitive transcription factor binding mechanism determines the timing of late cell cycle-dependent gene expression. Mol. Cell 2010, 38, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Sorrells, T.R.; Johnson, A.N.; Howard, C.J.; Britton, C.S.; Fowler, K.R.; Feigerle, J.T.; Weil, P.A.; Johnson, A.D. Intrinsic cooperativity potentiates parallel cis-regulatory evolution. eLife 2018, 7, e37563. [Google Scholar] [CrossRef]
- Rossi, M.J.; Lai, W.K.M.; Pugh, B.F. Genome-wide determinants of sequence-specific DNA binding of general regulatory factors. Genome Res. 2018, 28, 497–508. [Google Scholar] [CrossRef] [PubMed]
- Primig, M.; Winkler, H.; Ammerer, G. The DNA binding and oligomerization domain of MCM1 is sufficient for its interaction with other regulatory proteins. EMBO J. 1991, 10, 4209–4218. [Google Scholar] [CrossRef]
- Bruhn, L.; Hwang-Shum, J.J.; Sprague, G.F., Jr. The N-terminal 96 residues of MCM1, a regulator of cell type-specific genes in Saccharomyces cerevisiae, are sufficient for DNA binding, transcription activation, and interaction with alpha 1. Mol. Cell. Biol. 1992, 12, 3563–3572. [Google Scholar] [PubMed]
- Chang, H.Y.; Lin, J.Y.; Lee, H.C.; Wang, H.L.; King, C.Y. Strain-specific sequences required for yeast [PSI+] prion propagation. Proc. Natl. Acad. Sci. USA 2008, 105, 13345–13350. [Google Scholar] [CrossRef]
- Krobitsch, S.; Lindquist, S. Aggregation of huntingtin in yeast varies with the length of the polyglutamine expansion and the expression of chaperone proteins. Proc. Natl. Acad. Sci. USA 2000, 97, 1589–1594. [Google Scholar] [CrossRef] [PubMed]
- Muchowski, P.J.; Schaffar, G.; Sittler, A.; Wanker, E.E.; Hayer-Hartl, M.K.; Hartl, F.U. Hsp70 and hsp40 chaperones can inhibit self-assembly of polyglutamine proteins into amyloid-like fibrils. Proc. Natl. Acad. Sci. USA 2000, 97, 7841–7846. [Google Scholar] [CrossRef] [PubMed]
- Serpionov, G.V.; Alexandrov, A.I.; Antonenko, Y.N.; Ter-Avanesyan, M.D. A protein polymerization cascade mediates toxicity of non-pathological human huntingtin in yeast. Sci. Rep. 2015, 5, 18407. [Google Scholar] [CrossRef] [PubMed]
- Hughes, R.E.; Lo, R.S.; Davis, C.; Strand, A.D.; Neal, C.L.; Olson, J.M.; Fields, S. Altered transcription in yeast expressing expanded polyglutamine. Proc. Natl. Acad. Sci. USA 2001, 98, 13201–13206. [Google Scholar] [CrossRef] [PubMed]
- Cao, F.; Levine, J.J.; Li, S.H.; Li, X.J. Nuclear aggregation of huntingtin is not prevented by deletion of chaperone Hsp104. Biochim. Biophys. Acta Mol. Basis Dis. 2001, 1537, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Kukushkin, Y.; Gupta, R.; Chen, T.; Konagai, A.; Hipp, M.S.; Hayer-Hartl, M.; Hartl, F.U. PolyQ proteins interfere with nuclear degradation of cytosolic proteins by sequestering the Sis1p chaperone. Cell 2013, 154, 134–145. [Google Scholar] [CrossRef]
- Miller, S.B.; Ho, C.T.; Winkler, J.; Khokhrina, M.; Neuner, A.; Mohamed, M.Y.; Guilbride, D.L.; Richter, K.; Lisby, M.; Schiebel, E.; et al. Compartment-specific aggregases direct distinct nuclear and cytoplasmic aggregate deposition. EMBO J. 2015, 34, 778–797. [Google Scholar] [CrossRef] [PubMed]
- Blumberg, H.; Silver, P. A split zinc-finger protein is required for normal yeast growth. Gene 1991, 107, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Norris, D. The SFP1 gene product of Saccharomyces cerevisiae regulates G2/M transitions during the mitotic cell cycle and DNA-damage response. Genetics 1998, 150, 1419–1428. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, P.; Nishikawa, J.L.; Breitkreutz, B.J.; Tyers, M. Systematic identification of pathways that couple cell growth and division in yeast. Science 2002, 297, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Fingerman, I.; Nagaraj, V.; Norris, D.; Vershon, A.K. Sfp1 plays a key role in yeast ribosome biogenesis. Eukaryot. Cell 2003, 2, 1061–1068. [Google Scholar] [CrossRef] [PubMed]
- Marion, R.M.; Regev, A.; Segal, E.; Barash, Y.; Koller, D.; Friedman, N.; O’Shea, E.K. Sfp1 is a stress- and nutrient-sensitive regulator of ribosomal protein gene expression. Proc. Natl. Acad. Sci. USA 2004, 101, 14315–14322. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, P.; Rupes, I.; Sharom, J.R.; Schneper, L.; Broach, J.R.; Tyers, M. A dynamic transcriptional network communicates growth potential to ribosome synthesis and critical cell size. Genes Dev. 2004, 18, 2491–2505. [Google Scholar] [CrossRef] [PubMed]
- Cipollina, C.; van den Brink, J.; Daran-Lapujade, P.; Pronk, J.T.; Porro, D.; de Winde, J.H. Saccharomyces cerevisiae SFP1: At the crossroads of central metabolism and ribosome biogenesis. Microbiology 2008, 154, 1686–1699. [Google Scholar] [CrossRef] [PubMed]
- Drozdova, P.; Rogoza, T.; Radchenko, E.; Lipaeva, P.; Mironova, L. Transcriptional response to the [ISP+] prion of Saccharomyces cerevisiae differs from that induced by the deletion of its structural gene, SFP1. FEMS Yeast Res. 2014, 14, 1160–1170. [Google Scholar] [CrossRef] [PubMed]
- Radchenko, E.; Rogoza, T.; Khokhrina, M.; Drozdova, P.; Mironova, L. SUP35 expression is enhanced in yeast containing [ISP+], a prion form of the transcriptional regulator Sfp1. Prion 2011, 5, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Drozdova, P.B.; Radchenko, E.A.; Rogoza, T.M.; Khokhrina, M.A.; Mironova, L.N. The SFP1 controls translation termination in Saccharomyces cerevisiae via regulation of Sup35p (eRF3) level. Mol. Biol. 2013, 47, 242–247. [Google Scholar] [CrossRef]
- Hughes, J.D.; Estep, P.W.; Tavazoie, S.; Church, G.M. Computational identification of cis-regulatory elements associated with groups of functionally related genes in Saccharomyces cerevisiae. J. Mol. Biol. 2000, 296, 1205–1214. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Byers, K.J.; McCord, R.P.; Shi, Z.; Berger, M.F.; Newburger, D.E.; Saulrieta, K.; Smith, Z.; Shah, M.V.; Radhakrishnan, M.; et al. High-resolution DNA-binding specificity analysis of yeast transcription factors. Genome Res. 2009, 19, 556–566. [Google Scholar] [CrossRef] [PubMed]
- Albert, B.; Tomassetti, S.; Gloor, Y.; Dilg, D.; Mattarocci, S.; Kubik, S.; Shore, D. Sfp1 regulates transcriptional networks driving cell growth and division through multiple promoter-binding modes. Genes Dev. 2019, 33, 288–293. [Google Scholar] [CrossRef] [PubMed]
- Dagkessamanskaya, A.; Ter-Avanesyan, M.; Mager, W.H. Transcriptional regulation of SUP35 and SUP45 in Saccharomyces cerevisiae. Yeast 1997, 13, 1265–1274. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Transcription Factor | Gene | TFBS Start (Rel. to ORF Start) | TFBS End (Rel. to ORF Start) | %Score |
|---|---|---|---|---|
| Mcm1 | SUP35 | −315 | −304 | 81.3% |
| −281 | −270 | 82.7% | ||
| −169 | −158 | 80.4% | ||
| −40 | −29 | 75.6% | ||
| SUP45 | −43 | −32 | 83.6% | |
| −67 | −56 | 81.5% | ||
| −165 | −154 | 76.4% | ||
| −388 | −377 | 75.2% | ||
| −435 | −424 | 76% | ||
| −475 | −464 | 76.6% | ||
| Sfp1 | SUP35 | −426 | −406 | 75.1% |
| −328 | −308 | 81.9% | ||
| −163 | −143 | 93.3% | ||
| SUP45 | −20 | 1 | 77.3% | |
| −174 | −154 | 92.6% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matveenko, A.G.; Mikhailichenko, A.S.; Drozdova, P.B.; Zhouravleva, G.A. Transcription Factors Mcm1 and Sfp1 May Affect [PSI+] Prion Phenotype by Altering the Expression of the SUP35 Gene. Microbiol. Res. 2024, 15, 508-524. https://doi.org/10.3390/microbiolres15020034
Matveenko AG, Mikhailichenko AS, Drozdova PB, Zhouravleva GA. Transcription Factors Mcm1 and Sfp1 May Affect [PSI+] Prion Phenotype by Altering the Expression of the SUP35 Gene. Microbiology Research. 2024; 15(2):508-524. https://doi.org/10.3390/microbiolres15020034
Chicago/Turabian StyleMatveenko, Andrew G., Anastasiia S. Mikhailichenko, Polina B. Drozdova, and Galina A. Zhouravleva. 2024. "Transcription Factors Mcm1 and Sfp1 May Affect [PSI+] Prion Phenotype by Altering the Expression of the SUP35 Gene" Microbiology Research 15, no. 2: 508-524. https://doi.org/10.3390/microbiolres15020034