
N6-benzyladenine (BAP)-Based Seed Preconditioning Enhances the Shoot Regeneration of Seedling-Derived Explants for Subsequent Indirect Gene Transfer in Soybeans (Glycine max [L.] Merrill.)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Culture Medium
2.2. Preparation of MS Culture Media
2.3. Seed Preconditioning, Germination, and Seedling Establishment
2.4. Explant Preparation and Subculturing for Shoot Induction
2.5. Shoot Elongation, Rooting, and Acclimatization Ex Vitro
2.6. Growth Conditions, Data Collection, and Statistical Analyses
3. Results
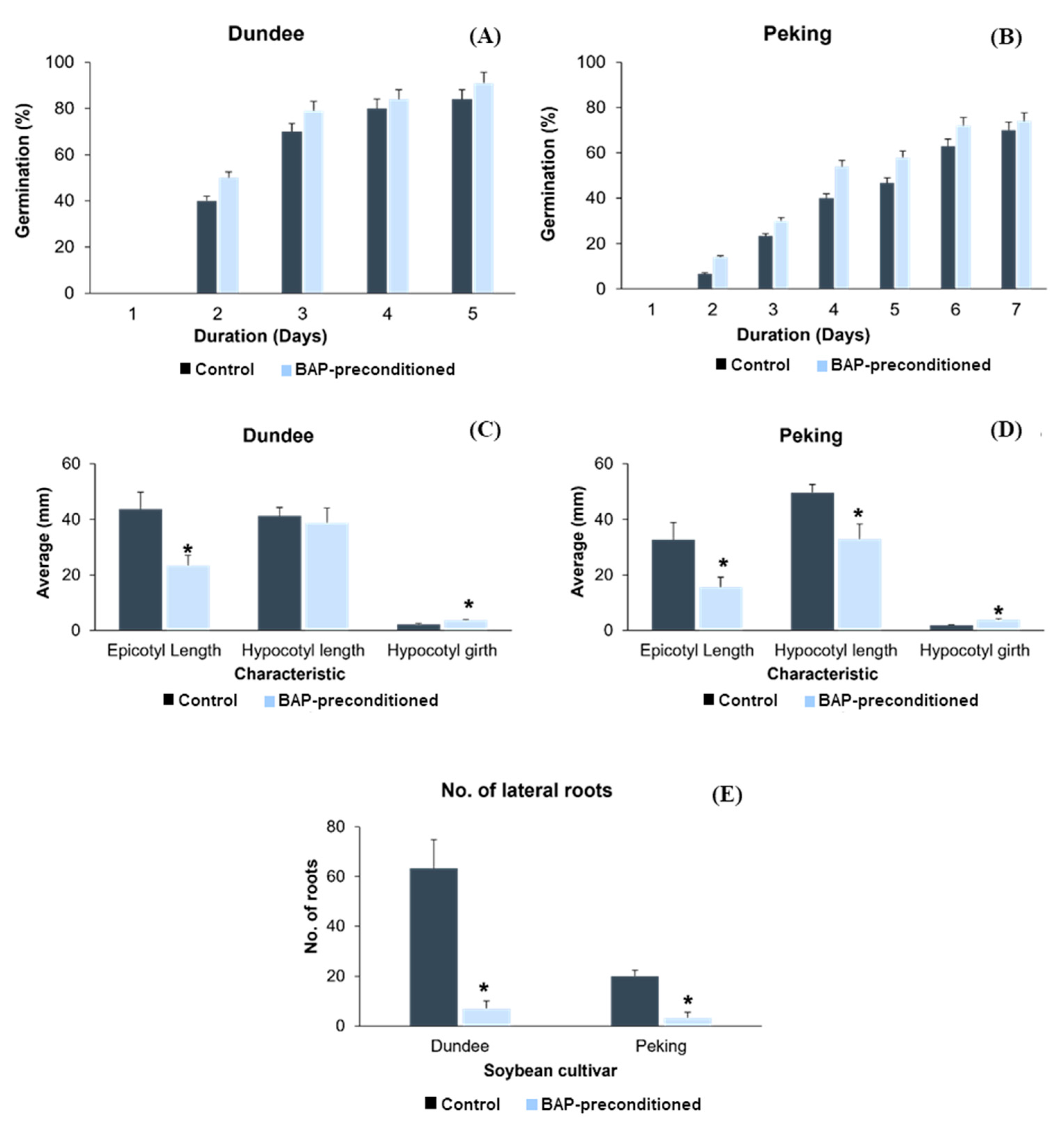
3.1. Effects of BAP Seed Preconditioning on Germination and Seedling Development
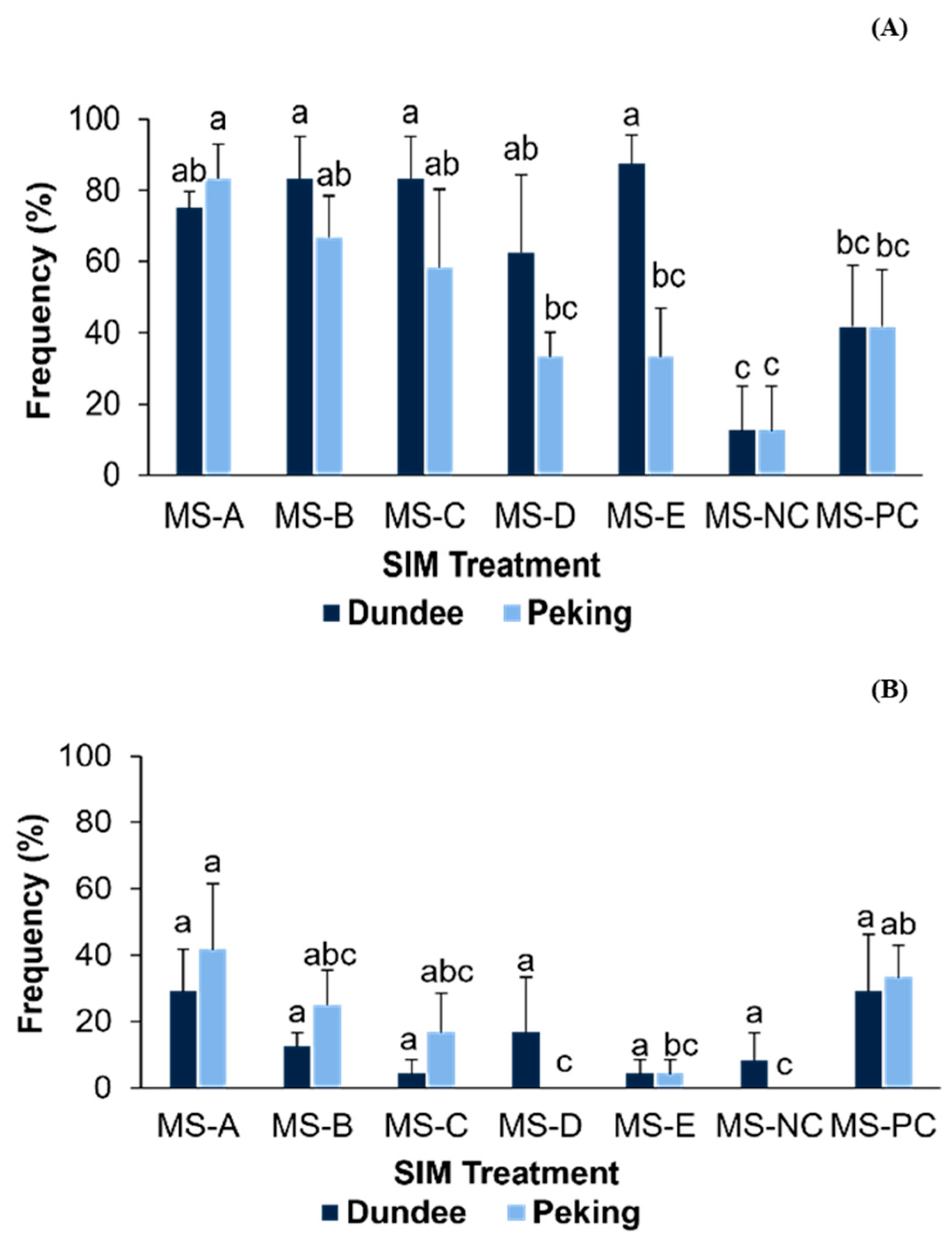
3.2. Effect of Culture Medium and Explants on Shoot Proliferation
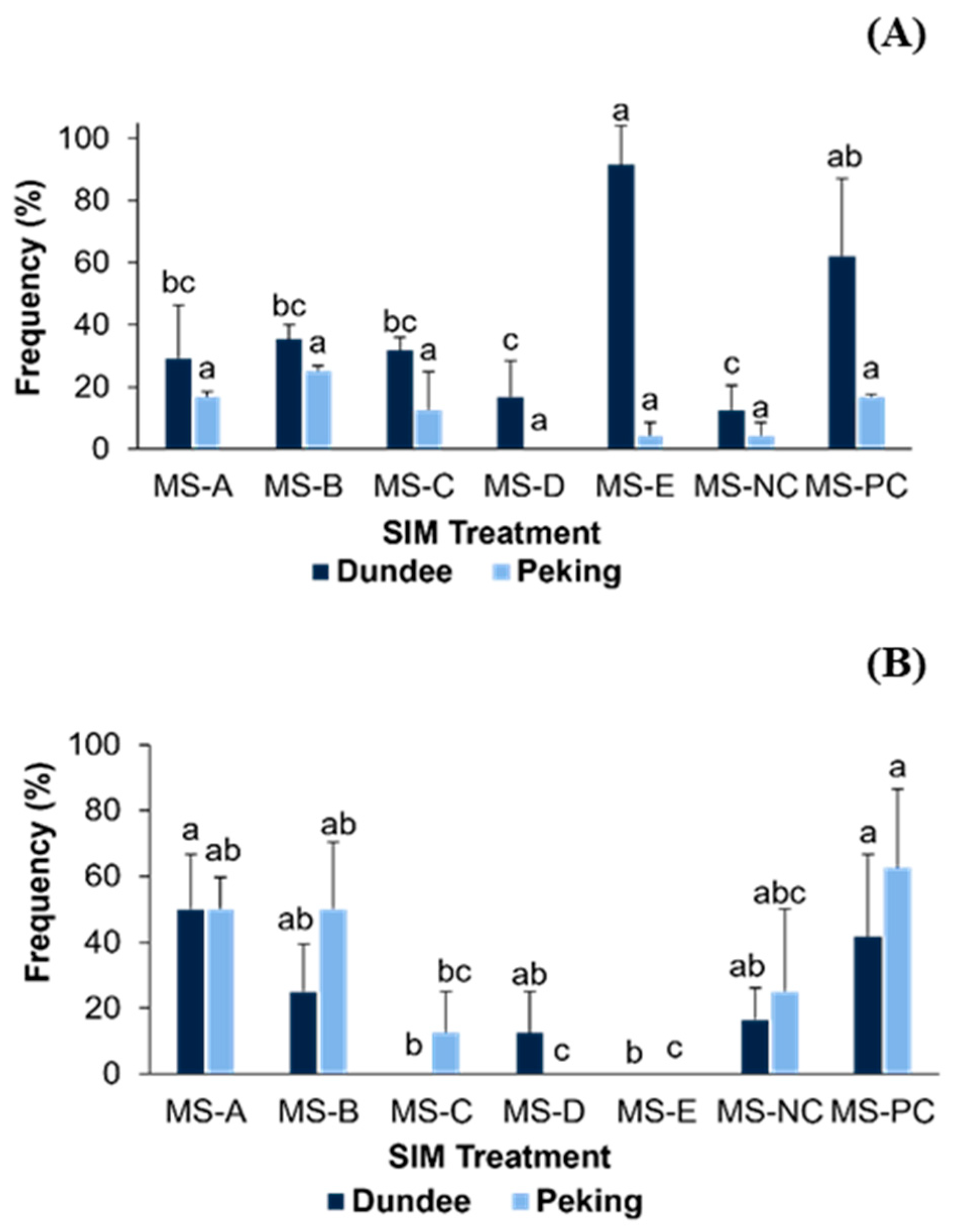
3.3. In Vitro Elongation and Rooting of Induced Shoots
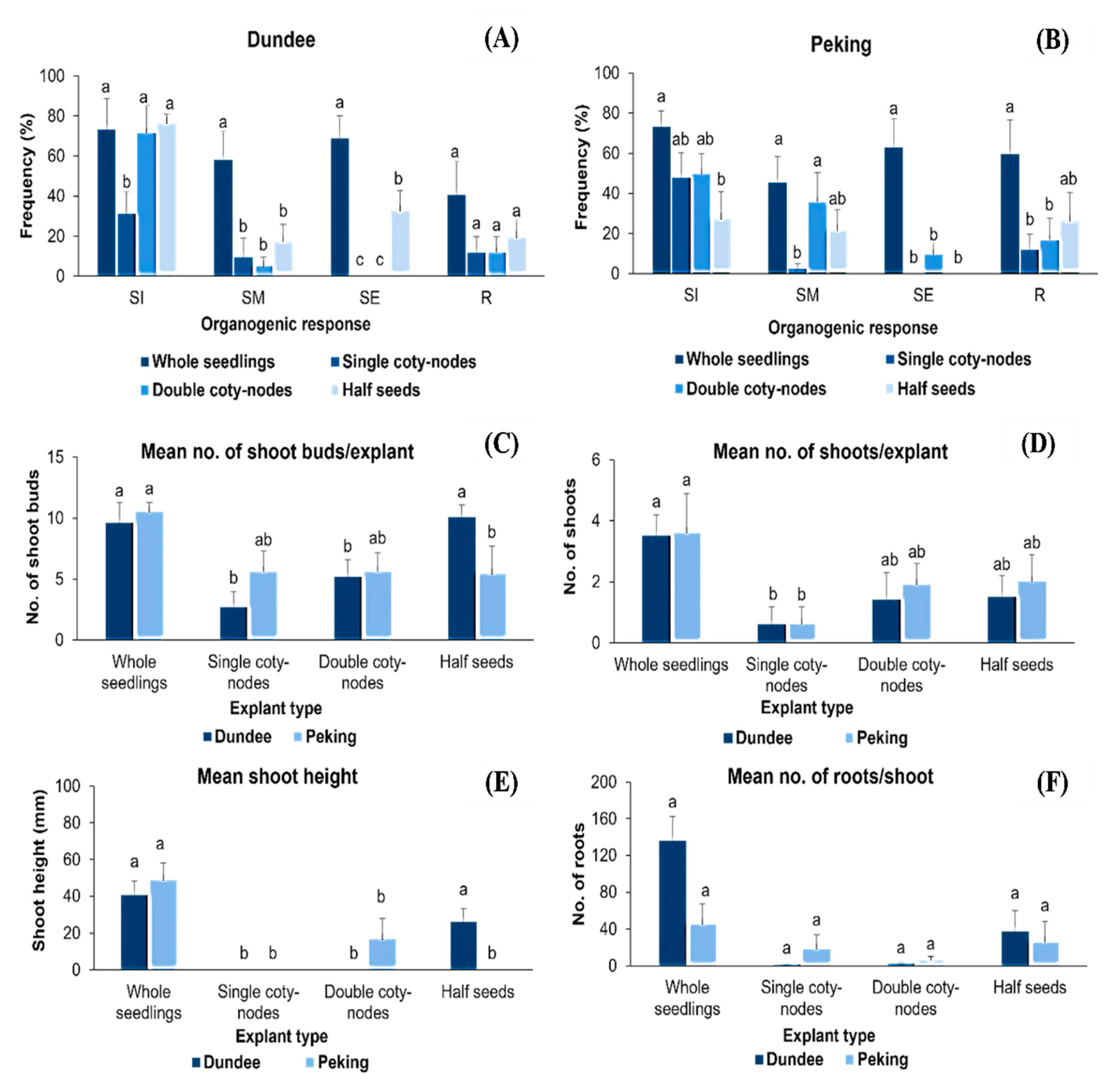
3.4. Overall Analysis of the Role of BAP on Shoot Regeneration in Soybeans
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Raza, G.; Singh, M.B.; Bhalla, P.L. Somatic embryogenesis and plant regeneration from commercial soybean cultivars. Plants 2020, 9, 38. [Google Scholar] [CrossRef]
- Cui, Y.; Zhao, J.; Gao, Y.; Zhao, R.; Zhang, J.; Kong, L. Efficient multi-sites genome editing and plant regeneration via somatic embryogenesis in Picca glauca. Front. Plant Sci. 2021, 12, 751891. [Google Scholar] [CrossRef]
- Lian, Z.; Ngauyen, C.D.; Liu, L.; Wang, G.; Chen, J.; Wang, S.; Yi, G.; Wilson, S.; Ozias-Akins, P.; Gong, H.; et al. Application of developmental regulators to improve in plants or in vitro transformation in plants. Plant Biotechnol. J. 2022, 20, 1622–1635. [Google Scholar] [CrossRef]
- Raza, G.; Singh, M.B.; Bhalla, P.L. In vitro plant regeneration from commercial cultivars of soybean. BioMed Res. Int. 2017, 2017, 7379693. [Google Scholar] [CrossRef]
- Begum, N.; Zenat, E.A.; Sarkar, M.K.I.; Roy, C.K.; Munshi, J.L.; Jahan, M.A.A. In vitro micro propagation of soybean (Glycine max) BARI-5 variety. Open Microbiol. J. 2019, 13, 177–187. [Google Scholar] [CrossRef]
- Paes de Melo, B.; Lourenco-Tessutti, I.T.; Morgante, C.V.; Santos, N.C.; Pinheiro, L.B.; Barrozo de Jesus Lins, C.; Matar Silva, M.C.; Macedo, L.L.P.; Bastita Fontes, E.P.; Grossi-de-Sa, M.F. Soybean embryogenic axis transformation: Combining biolistic and Agrobacterium-mediated protocols to overcome typical complications of in vitro plant regeneration. Front. Plant Sci. 2020, 11, 1228. [Google Scholar] [CrossRef] [PubMed]
- Jordan, M.C.; Hobbs, S.L.A. Evaluation of a cotyledonary node regeneration system for Agrobacterium-mediated transformation of pea (Pisum sativum L.). Vitr. Cell Dev. Biol. Plant 1993, 29, 77–82. Available online: https://www.jstor.org/stable/4292976 (accessed on 15 June 2023). [CrossRef]
- Schmidt, M.A.; Tucker, D.M.; Cahoon, E.B.; Parrott, W.A. Towards normalization of soybean somatic embryo maturation. Plant Cell Rep. 1998, 24, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassay with tobacco tissue culture. Plant Physiol. 1962, 15, 473–479. [Google Scholar] [CrossRef]
- Sojkova, J.; Zur, I.; Gregorova, Z.; Zimova, M.; Matusikova, I.; Mihalik, D.; Kraic, J.; Moravcikova, J. In vitro regeneration potential of seven commercial soybean cultivars (Glycine max L.) for use in biotechnology. Nova Biotechnol. Chem. 2016, 15, 1–11. [Google Scholar] [CrossRef]
- Marquez-Lopez, R.E.; Quintana-Escobar, A.O.; Loyola-Vargas, V.M. Cytokinins, the Cinderella of plant growth regulators. Phytochem. Rev. 2019, 18, 1387–1408. [Google Scholar] [CrossRef]
- Mangena, P.; Mokwala, P.W. The influence of seed viability on the germination and in vitro multiple shoot regeneration of soybean (Glycine max L.). Agriculture 2019, 9, 35. [Google Scholar] [CrossRef]
- Mangena, P.; Mokwala, P.W.; Nikolova, R.V. In vitro multiple shoot induction in soybean. Int. J. Agric. Biol. 2015, 17, 838–842. [Google Scholar] [CrossRef]
- Paz, M.M.; Martinez, J.C.; Kalvig, A.B.; Fonger, T.M.; Wang, K. Improved cotyledonary node method using an alternative explant derived from mature seed for efficient Agrobacterium-mediated soybean transformation. Plant Cell Rep. 2006, 25, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Wijewardana, C.; Alsajri, F.A.; Reddy, K.R. Soybean seed germination response to in vitro osmotic stress. Seed Technol. 2018, 39, 143–154. Available online: https://www.jstor.org/stable/45135884 (accessed on 7 July 2023).
- Walne, C.H.; Alsajri, F.A.; Gajanayake, B.; Lokhande, S.; Seepaul, R.; Wijewardana, C.; Reddy, K.R. In vitro seed germination response of corn, cotton, and soybean to temperature. J. Miss. Acad. Sci. 2020, 65, 374–384. [Google Scholar]
- Phat, P.; Rehman, S.U.; Jung, H.-I.; Ju, H.-J. Optimisation of soybean (Glycine max L.) regeneration for Korean cultivars. Pak. J. Bot. 2015, 47, 2379–2385. [Google Scholar]
- Ngomuo, M.; Mneney, E.; Ndakidemi, P. The effects of auxins and cytokinin on growth and development of (Musa sp.) var “Yangambi” explants in tissue culture. Am. J. Plant Sci. 2013, 4, 2174–2180. [Google Scholar] [CrossRef]
- Bhattacharya, J.; Renukdas, N.N.; Khuspe, S.S.; Rawal, S.K. Multiple shoot regeneration from immature embryo explants of papaya. Biol. Plant 2003, 47, 327–331. Available online: https://bp.ueb.cas.cz/pdfs/bpl/2003/03/04.pdf (accessed on 15 June 2023). [CrossRef]
- Damanik, R.I.; Manurung, B.H.; Bayu, E.S. Effects of hypoxia condition in embryogenic callus growth of soybean cell culture. IOP Conf. Ser. Earth Environ. Sci. 2018, 122, 1–6. [Google Scholar] [CrossRef]
- Silue, S.; Diarrassouba, N.; Fofana, I.J.; Muhovski, Y.; Toussaint, A.; Mergeai, G.; Jacquemin, J.-M.; Beudoin, J.-P. Description of Phaseolus vulgaris L. aborting embryos from ethyl methanesulfonate (EMS) mutagenised plants. Biotechnol. Agron. Soc. Environ. 2013, 17, 564–571. [Google Scholar]
- Poniewozik, M.; Szot, P.; Parzymies, M. Tissue culture multiplication of Paphiopedilum insigne depending on the medium type, growth regulators and natural supplements. Acta Sci. Pol. Hortorum Cultus 2021, 20, 125–134. [Google Scholar] [CrossRef]
- Sevik, H.; Guney, K. Effects of IAA, IBA, NAA, and GA3 on rooting and morphological features of Melissa officinalis L. stem cuttings. Sci. World J. 2013, 30, 909507. [Google Scholar] [CrossRef]
- Yan, H.; Lu, L.; Alzate, A.; Ceballos, H.; Hershey, C.; Chen, S. Fruit, seed, and embryo development of different cassava (Manihot esculeta Crantz) genotypes and embryo rescue. Afr. J. Biotechnol. 2014, 131, 1524–1528. [Google Scholar] [CrossRef]
- Polisetty, R.; Paul, V.; Deveshwar, J.J.; Khetarpal, S.; Suresh, K.; Chandra, R. Multiple shoot induction by benzyladenine and complete plant regeneration from seed explants of chickpea (Cicer arietinum L.). Plant Cell Rep. 1997, 16, 565–571. [Google Scholar] [CrossRef]
- Soto, N.; Delgado, C.; Hernandez, Y.; Rosabal, Y.; Ferreira, A.; Pujol, M.; Aragao, F.J.L.; Enriquez, G.A. Efficient particle bombardment-mediated transformation of Cuban soybean (INCASoy-36) using glyphosate as a selective. Plant Cell Tissue Organ. Cult. 2016, 128, 187–196. [Google Scholar] [CrossRef]
- Sindhu, M.; Kumar, A.; Yadav, H.; Chaudhary, D.; Jaiwal, R.; Jaiwal, P.K. Current advances and future directions in genetic enhancement of a climate resilient food legume crop, cowpea (Vigna unguiculata L. Walp.). Plant Cell Tissue Organ. Cult. 2019, 139, 429–453. [Google Scholar] [CrossRef]
- Khabbazi, S.D.; Barpete, S.; Sancak, C.; Ozcan, S. Preconditioning effect of BAP on in vitro multiplication of mature embryo of lentil (Lens culinaris Medik.). Fresenius Env. Environ. Bull. 2017, 26, 919–925. [Google Scholar]
- Pratap, A.; Prajapati, U.; Singh, C.M.; Gupta, S.; Rathore, M.; Malviya, N.; Tomar, R.; Gupta, A.K.; Tripathi, S.; Singh, N.P. Potential, constraints, and applications of in vitro methods in improving grain legumes. Plant Breed. 2018, 137, 235–249. [Google Scholar] [CrossRef]
- Zia, M.; Rizvi, Z.F.; Rehman, R.; Chaudhary, M.F. Micropropagation of two Pakistani soybean (Glycine max L.) cultivars from cotyledonary nodes. Span. J. Agric. Res. 2010, 8, 448–453. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sehaole, E.M.K.; Mangena, P. N6-benzyladenine (BAP)-Based Seed Preconditioning Enhances the Shoot Regeneration of Seedling-Derived Explants for Subsequent Indirect Gene Transfer in Soybeans (Glycine max [L.] Merrill.). Int. J. Plant Biol. 2024, 15, 254-266. https://doi.org/10.3390/ijpb15020022
Sehaole EMK, Mangena P. N6-benzyladenine (BAP)-Based Seed Preconditioning Enhances the Shoot Regeneration of Seedling-Derived Explants for Subsequent Indirect Gene Transfer in Soybeans (Glycine max [L.] Merrill.). International Journal of Plant Biology. 2024; 15(2):254-266. https://doi.org/10.3390/ijpb15020022
Chicago/Turabian StyleSehaole, Esmerald Michel Khomotso, and Phetole Mangena. 2024. "N6-benzyladenine (BAP)-Based Seed Preconditioning Enhances the Shoot Regeneration of Seedling-Derived Explants for Subsequent Indirect Gene Transfer in Soybeans (Glycine max [L.] Merrill.)" International Journal of Plant Biology 15, no. 2: 254-266. https://doi.org/10.3390/ijpb15020022
APA StyleSehaole, E. M. K., & Mangena, P. (2024). N6-benzyladenine (BAP)-Based Seed Preconditioning Enhances the Shoot Regeneration of Seedling-Derived Explants for Subsequent Indirect Gene Transfer in Soybeans (Glycine max [L.] Merrill.). International Journal of Plant Biology, 15(2), 254-266. https://doi.org/10.3390/ijpb15020022