
Inheritance of Mitochondria in Pelargonium Section Ciconium (Sweet) Interspecific Crosses

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material, DNA Extraction, and Sequencing
2.2. Mitome Assembly
2.3. Organellar Genotyping Using PCR and KASP Markers
3. Results
3.1. Confirmed F1 Hybrids
3.2. Mitome Assemblies
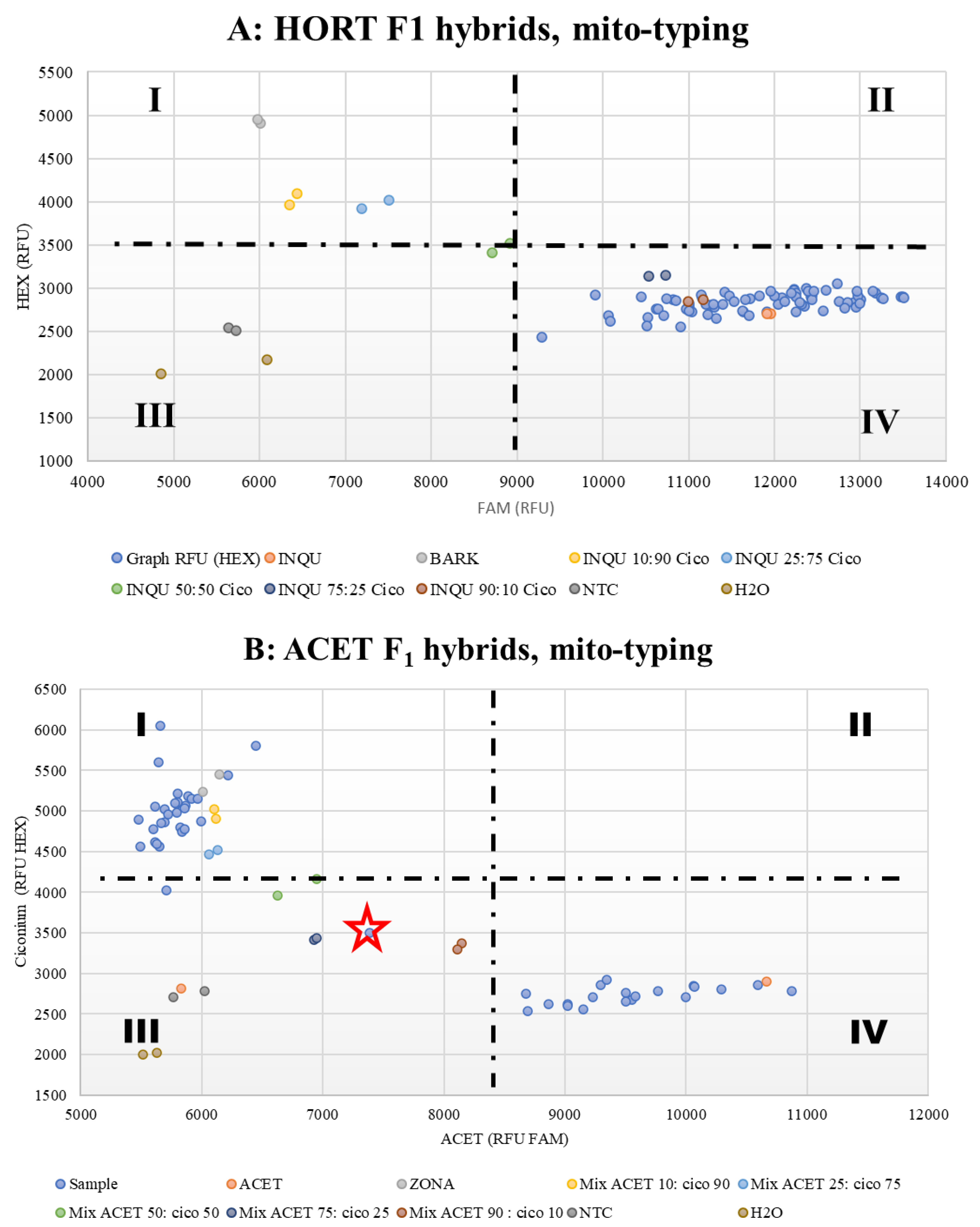
3.3. Mitotyping
4. Discussion
4.1. The Case of P. multibracteatum
4.2. Evolutionary Effects of mCNI
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bateson, W. Heredity and variation in modern lights. In Darwin and Modern Science; Seward, A.C., Ed.; Cambridge University Press: Cambridge, UK, 1909; pp. 85–101. [Google Scholar]
- Dobzhansky, T. Studies on hybrid sterility. II. Localization of sterility factors in Drosophila pseudoobscura hybrids. Genetics 1936, 21, 113–135. [Google Scholar] [CrossRef]
- Müller, H.J. Isolating mechanisms, evolution, and temperature. Biol. Symp. 1942, 6, 71–125. [Google Scholar]
- Schnable, P.S.; Wise, R.P. The molecular basis of cytoplasmic male sterility and fertility restoration. Trends Plant Sci. 1998, 3, 175–180. [Google Scholar] [CrossRef]
- Baur, E. Das Wesen und die Earblichkeitsverhältnisse der ‘varietates albomarginatae hort’ von Pelargonium zonale. Z. Indukt. Abstamm. -Vererbungsl. 1909, 1, 330–351. [Google Scholar] [CrossRef]
- Horn, W. Interspecific crossability and inheritance in Pelargonium. Plant Breed. 1994, 113, 3–17. [Google Scholar] [CrossRef]
- Breman, F.C.; Snijder, R.C.; Korver, J.W.; Pelzer, S.; Sancho-Such, M.; Schranz, M.E.; Bakker, F.T. Interspecific Hybrids Between Pelargonium × hortorum and species from P. section Ciconium reveal biparental plastid inheritance and multi-locus cyto-nuclear incompatibility. Front. Plant Sci. 2020, 11, 614871. [Google Scholar] [CrossRef] [PubMed]
- Tilney-Bassett, R.A.E. The control of plastid inheritance in Pelargonium II. Heredity 1973, 30, 1–13. [Google Scholar] [CrossRef]
- Tilney-Bassett, R.A.E. The control of plastid inheritance in Pelargonium III. Heredity 1974, 33, 353–360. [Google Scholar] [CrossRef]
- Tilney-Bassett, R.A.E. Genetics of variegated plants. In Genetics and Biogenesis of Mitochondria and Chloroplasts; Birky, C.W., Perlman, P.S., Byers, T.J., Eds.; Ohio State University Press: Columbus, OH, USA, 1975; pp. 268–308. [Google Scholar]
- Breman, F.C. Exploring patterns of cytonuclear incompatibility in Pelargonium section Ciconium. Ph.D. Thesis, Wageningen University, Wageningen, The Netherlands, 2021. [Google Scholar] [CrossRef]
- Tilney-Bassett, R.A.E.; Almouslem, A.B.; Amoate, H.M. Complementary genes control biparental plastid inheritance in Pelargonium. Theor. Appl. Genet. 1992, 85, 317–324. [Google Scholar] [CrossRef]
- Weihe, A.; Apitz, J.; Salinas, A.; Pohlheim, F.; Börner, T. Biparental inheritance of plastidial and mitochondrial DNA and hybrid variegation in Pelargonium. Mol. Genet. Genomics 2009, 282, 587–593. [Google Scholar] [CrossRef]
- Greiner, S.; Sobanski, J.; Bock, R. Why are most organelle genomes transmitted maternally? BioEssays 2015, 37, 80–94. [Google Scholar] [CrossRef] [PubMed]
- Havey, M. Predominant paternal transmission of the mitochondrial genome in cucumber. J. Hered. 1997, 88, 232–235. [Google Scholar] [CrossRef]
- Park, H.-S.; Lee, W.K.; Lee, S.-C.; Lee, H.O.; Joh, H.J.; Park, J.Y.; Kim, S.; Song, K.; Yang, T.-J. Inheritance of chloroplast and mitochondrial genomes in cucumber revealed by four reciprocal F1 hybrid combinations. Sci. Rep. 2021, 11, 2506. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Neale, D.B.; Marshall, K.A.; Sederoff, R.R. Chloroplast and mitochondrial DNA are paternally inherited in Sequoia sempervirens D. Don Endl. Proc. Natl. Acad. Sci. USA 1989, 86, 9347–9349. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Noyer, J.-L.; Horry, J.-P.; Lanaud, C.; Fauré, S.; Carreel, F.; Bakry, F. Maternal inheritance of chloroplast genome and paternal inheritance of mitochondrial genome in bananas (Musa acuminata). Curr. Genet. 1994, 25, 265–269. [Google Scholar] [CrossRef] [PubMed]
- Munasinghe, M.; Ågren, J.A. When and why are mitochondria paternally inherited? Curr. Opin. Genet. Dev. 2023, 80, 102053. [Google Scholar] [CrossRef] [PubMed]
- Khachaturyan, M.; Santer, M.; Reusch, T.B.; Dagan, T. Heteroplasmy is rare in plant mitochondria compared to plastids despite similar mutation rates. bioRxiv 2023. [Google Scholar] [CrossRef] [PubMed]
- Sobanski, J.; Giavalisco, P.; Fischer, A.; Kreiner, J.M.; Walther, D.; Schöttler, M.A.; Pellizzer, T.; Golczyk, H.; Obata, T.; Bock, R.; et al. Chloroplast competition is controlled by lipid biosynthesis in evening primroses. Proc. Natl. Acad. Sci. USA 2019, 116, 5665–5674. [Google Scholar] [CrossRef]
- Kuroiwa, T.; Kawazu, T.; Ushida, H.; Ohta, T.; Kuroiwa, H. Direct evidence of plastid DNA and mitochondrial DNA in sperm cells in relation to biparental inheritance of organelle DNA in Pelargonium zonale by fluorescence/electron microscopy. Eur. J. Cell Biol. 1993, 62, 307–313. [Google Scholar]
- Park, S.; Grewe, F.; Zhu, A.; Ruhlman, T.A.; Sabir, J.; Mower, J.P.; Jansen, R.K. Dynamic evolution of Geranium mitochondrial genomes through multiple horizontal and intracellular gene transfers. New Phytol. 2015, 208, 570–583. [Google Scholar] [CrossRef]
- Breman, F.C.; Korver, J.W.; Snijder, R.C.; Bakker, F.T. Plastid-encoded RNA polymerase variation in Pelargonium sect. Ciconium. Hortic. Adv. 2024, 2, 1. [Google Scholar] [CrossRef]
- Bakker, F.T.; Culham, A.; Marais, E.M.; Gibby, M. Nested radiation in Cape Pelargonium. In Plant Species-Level Systematics: New Perspectives on Pattern and Process; Bakker, F.T., Chartrou, L.W., Gravendeel, B., Pielser, P.B., Eds.; A.R.G. Ganter Verlag K.G.: Ruggell, Liechtstein, 2005; pp. 75–100. [Google Scholar]
- Jin, J.-J.; Yu, W.-B.; Yang, J.-B.; Song, Y.; Depamphilis, C.W.; Yi, T.-S.; Li, D.-Z. GetOrganelle: A fast and versatile toolkit for accurate de novo assembly of organelle genomes. Genome Biol. 2020, 21, 241. [Google Scholar] [CrossRef] [PubMed]
- Bakker, F.T.; Breman, F.C.; Merckx, V. DNA sequence evolution in fast evolving mitochondrial DNA nad1 exons in Geraniaceae and Plantaginaceae. Taxon 2006, 55, 887–896. [Google Scholar] [CrossRef]
- Grewe, F.; Zhu, A.; Mower, J.P. Loss of a trans-splicing nad1 intron from Geraniaceae and transfer of the maturase gene matR to the nucleus in Pelargonium. Genome Biol. Evol. 2016, 8, 3193–3201. [Google Scholar] [CrossRef] [PubMed]
- Hiremath, P.J.; Kumar, A.; Penmetsa, R.V.; Farmer, A.; Schlueter, J.A.; Chamarthi, S.K.; Whaley, A.M.; Carrasquilla-Garcia, N.; Gaur, P.M.; Upadhyaya, H.D.; et al. Large-scale development of cost- effective SNP marker assays for diversity assessment and genetic mapping in chickpea and comparative mapping in legumes. Plant Biotechnol. J. 2012, 10, 716–732. [Google Scholar] [CrossRef] [PubMed]
- Semagn, K.; Babu, R.; Hearne, S. Single nucleotide polymorphism genotyping using Kompetitive Allele Specific PCR (KASP): Over view of the technology and its application in crop improvement. Mol. Breed. 2014, 33, 1–14. [Google Scholar] [CrossRef]
- Islam, M.S.; Thyssen, G.N.; Jenkins, J.N.; Fang, D.D. Detection, validation, and application of genotyping-by-sequencing based single nucleotide polymorphisms in upland Cotton. Plant Genome 2014, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Steele, K.A.; Quinton-Tulloch, M.J.; Amgai, R.B.; Dhakal, R.; Khatiwada, S.P.; Vyas, D.; Heine, M.; Witcombe, J.R. Accelerating public sector rice breeding with high-density KASP markers derived from whole genome sequencing of indica rice. Mol. Breed. 2018, 38, 38. [Google Scholar] [CrossRef]
- Bakker, F.T.; Culham, A.; Pankhurst, C.E.; Gibby, M. Mitochondrial and chloroplast DNA-based phylogeny of Pelargonium (Geraniaceae). Am. J. Bot. 2000, 87, 727–734. [Google Scholar] [CrossRef]
- Palmer, J.D.; Adams, K.L.; Cho, Y.; Parkinson, C.L.; Qiu, Y.L.; Song, K. Dynamic evolution of plant mitochondrial genomes: Mobile genes and introns and highly variable mutation rates. Proc. Natl. Acad. Sci. USA 2000, 97, 6960–6966. [Google Scholar] [CrossRef]
- Choi, K.; Weng, M.-L.; Ruhlman, T.A.; Jansen, R.K. Extensive variation in nucleotide substitution rate and gene/intron loss in mitochondrial genomes of Pelargonium. Mol. Phylogenet. Evol. 2021, 155, 106986. [Google Scholar] [CrossRef]
- Mower, J.P.; Touzet, P.; Gummow, J.S.; Delph, L.F.; Palmer, J.D. Extensive variation in synonymous substitution rates in mitochondrial genes of seed plants. BMC Evol. Biol. 2007, 7, 135. [Google Scholar] [CrossRef] [PubMed]
- van de Kerke, S.J.; Shrestha, B.; Ruhlman, T.A.; Weng, M.-L.; Jansen, R.K.; Jones, C.S.; Schlichting, C.D.; Hosseini, S.; Mohammadin, S.; Schranz, M.E.; et al. Plastome based phylogenetics and younger crown node age in Pelargonium. Mol. Phylogenet. Evol. 2019, 137, 33–43. [Google Scholar] [CrossRef]
- Guo, F.L.; Hu, S.Y. Cytological evidence of biparental inheritance of plastids and mitochondria in Pelargonium. Protoplasma 1995, 186, 201–207. [Google Scholar] [CrossRef]
- Kirk, J.T.O.; Tilney-Bassett, R.A.E. The Plastids; Freeman and, Co.: London, UK, 1967. [Google Scholar]
- Chung, K.P.; Gonzalez-Duran, E.; Ruf, S.; Endries, P.; Bock, R. Control of plastid inheritance by environmental and genetic factors. Nat. Plants 2023, 9, 68–80. [Google Scholar] [CrossRef]
- Sakamoto, W.; Takami, T. Maternal plastid inheritance: Two abating factors identified. Trends Genet. 2023, 39, 342–343. [Google Scholar] [CrossRef]
- Havey, M.J. Organellar genomes of the cucurbits. In Genetics and Genomics of Cucurbitaceae; Grumet, R., Katzir, N., Garcia-Mas, J., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; pp. 241–252. [Google Scholar]
- Apitz, J.; Weihe, A.; Pohlheim, F.; Börner, T. Biparental inheritance of organelles in Pelargonium: Evidence for intergenomic recombination of mitochondrial DNA. Planta 2013, 237, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Kuroiwa, H.; Kuroiwa, T. Giant mitochondria in the mature egg cell of Pelargonium zonale. Protoplasma 1992, 168, 184–188. [Google Scholar] [CrossRef]
- Maréchal, A.; Brisson, N. Recombination and the maintenance of plant organelle genome stability. New Phytol. 2010, 186, 299–317. [Google Scholar] [CrossRef]
- Barnard-Kubow, K.B.; McCoy, M.A.; Galloway, L.F. Biparental chloroplast inheritance leads to rescue from cytonuclear incompatibility. New Phytol. 2017, 213, 1466–1476. [Google Scholar] [CrossRef]
- Barnard-Kubow, K.B.; So, N.; Galloway, L.F. Cytonuclear incompatibility contributes to the early stages of speciation. Evolution 2016, 70, 2752–2766. [Google Scholar] [CrossRef] [PubMed]
- Greiner, S.; Bock, R. Tuning a ménage à trois: Co-evolution and co-adaptation of nuclear and organellar genomes in plants. BioEssays 2013, 35, 354–365. [Google Scholar] [CrossRef] [PubMed]
- Röschenbleck, J.; Albers, F.; Müller, K.; Weinl, S.; Kudla, J. Phylogenetics, character evolution and a subgeneric revision of the genus Pelargonium (Geraniaceae). Phytotaxa 2014, 159, 31–76. [Google Scholar] [CrossRef]
- Bakker, F.T.; Culham, A.; Daugherty, L.C.; Gibby, M. A trnL-F based phylogeny for species of (Geraniaceae) with small chromosomes. Plant Syst. Evol. 1999, 216, 309–324. [Google Scholar] [CrossRef]
- Dodsworth, S.; Jang, T.S.; Struebig, M.; Chase, M.W.; Weiss-Schneeweiss, H.; Leitch, A.R. Genome-wide repeat dynamics reflect phylogenetic distance in closely related allotetraploid Nicotiana (Solanaceae). Plant Syst. Evol. 2017, 303, 1013–1020. [Google Scholar] [CrossRef]
- Breman, F.C.; Chen, G.; Snijder, R.C.; Schranz, M.E.; Bakker, F.T. Repeatome-Based Phylogenetics in Pelargonium Section Ciconium (Sweet) Harvey. Genome Biol. Evol. 2021, 13, evab269. [Google Scholar] [CrossRef]
- Knuth, R. Geraniaceae.—In Engler, A.; (Ed.): Das Pflanzenreich 4, 1–9.—Leipzig (1912), Germany Engelmann. Available online: https://www.biodiversitylibrary.org/bibliography/71251 (accessed on 23 April 2024).
- Van der Walt, J.J.A.; Albers, F.; Gibby, M. Delimitation of Pelargonium sect. Glaucophyllum (Geraniaceae). Plant Syst. Evol. 1990, 171, 15–26. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Species | Herbarium Voucher Accession | Institute 1 | Acronym Used in Text |
|---|---|---|---|
| P. acetosum | 1243 | STEU | ACET |
| P. acraeum | 1975 | STEU | ACRA |
| P. alchemilloides | 1885 | STEU | ALCH2x |
| P. alchemilloides | 1882 | STEU | ALCH4x |
| P. articulatum | 1972055 | WAG | ARTI |
| P. barklyi | 1972061 | WAG | BARK |
| P. frutetorum | 0754 | STEU | FRUT |
| P. inquinans | 0682 | STEU | INQU |
| P. multibracteatum | 2902 | STEU | MULT |
| P. peltatum | 1890 | STEU | PELT |
| P. quinquelobatum | 1972049 | WAG | QUIN |
| P. ranuncolophyllum | A3651 | MSUN * | RANU |
| P. tongaense | 3074 | STEU | TONG |
| P. zonale | 1896 | STEU | ZONA |
| P. elongatum | 0854 | STEU | ELON |
| P. aridum | 1847 | STEU | ARID |
| P. insularis | 19990489 | RBGE | INSU |
| P. yemenensesp. nov | 1972037 | WAG | YEME |
| P. omanensesp. nov | 2184 | RBGE | OMAN |
| P. somalense | V-067490 | V | SOMA |
| F1 Types | # Plants/Cross | # Marker Pairs/Cross | Fertility Phenotype | (M), (P), (B), (H), (R), (WT) |
|---|---|---|---|---|
| ACET_X_FRUT | 2 | 1 | P | M |
| ACET_X_INQU | 1 | 2 | P | M |
| ACET_X_ZONA | 12 | 1 | -- | M (11) P (1) |
| ALCH(4X)_X_BARK | 1 | 1 | -- | M |
| ALCH(4X)_X_YEME | 2 | 1 | P | P |
| BARK_X_FRUT | 3 | 1 | MS | M |
| BARK_X_INQU | 1 | 2 | -- | P |
| BARK_X_MULT | 3 | 2 | MS | M |
| BARK_X_QUIN | 2 | 1 | -- | M(2) P(1) |
| FRUT_X_ACET | 1 | 1 | P | M |
| FRUT_X_BARK | 3 | 1 | MS | M |
| HORT(4X)_X_ARTI(4X) | 8 | 1 | MS | M |
| (HORT_X_ZONA)_X_ARID | 5 | 1 | MS | M |
| HORT_X_ACET | 3 | 2 | P | M |
| HORT_X_ACRA | 1 | 1 | P | M |
| HORT_X_ALCH | 1 | 1 | MS | M |
| HORT_X_ARID | 6 | 2 | MS | M |
| HORT_X_BARK | 2 | 1 | -- | M |
| HORT_X_FRUT | 1 | 1 | F | M |
| HORT_X_MULT | 1 | 1 | MS | M |
| HORT_X_QUIN | 15 | 1 | MS | M(8) P(7) |
| HORT_X_TONG | 8 | 1 | P | M |
| HORT_X_TONG(4X) | 1 | 1 | P | M |
| HORT_X_ZONA | 26 | 1 | P | M |
| TONG_X_ACET | 7 | 1–2 | P | B(1)/P |
| YEME_X_ALCH(4X) | 1 | 1 | P | P |
| P. inquinans | 1 | 1 | F | WT |
| P. peltatum | 1 | 1 | F | WT |
| P. salmoneum | 1 | 2 | F | WT |
| P. x hortorum_4x | 1 | 1 | P | WT * |
| P. quinquelobatum | 1 | 3 | F | WT |
| P. yemenense | 1 | 3 | F | WT |
| P. barklyi | 1 | 3 | F | |
| P. aridum | 1 | 3 | F | WT |
| P. quinquelobatum | 1 | 3 | F | WT |
| P. alchemilloides | 1 | 3 | F | WT |
| P. tongaense | 1 | 1 | F | WT |
| P. articulatum | 1 | 1 | F | WT |
| P. multibracteatum | 1 | 2 | F | WT/H |
| mult_x_acet | 8 | 2-3 | -- | B/H * |
| mult_x_alch | 14 | 2 | P | P/H * |
| mult_x_arid | 3 | 1 | MS | P/H * |
| mult_x_bark | 5 | 2 | MS | B/H * |
| mult_x_inqu | 3 | 1 | -- | P/H * |
| mult_x_pelt | 3 | 1 | MS | P/H * |
| mult_x_quin | 6 | 2 | P | P/H * |
| mult_x_ranu | 9 | 1–2 | P | P/H * |
| mult_x_tong | 2 | 1 | MS | P/H * |
| mult_x_zona | 2 | 2 | MS | P/H * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Breman, F.C.; Korver, J.; Snijder, R.; Schranz, M.E.; Bakker, F.T. Inheritance of Mitochondria in Pelargonium Section Ciconium (Sweet) Interspecific Crosses. Int. J. Plant Biol. 2024, 15, 586-598. https://doi.org/10.3390/ijpb15030044
Breman FC, Korver J, Snijder R, Schranz ME, Bakker FT. Inheritance of Mitochondria in Pelargonium Section Ciconium (Sweet) Interspecific Crosses. International Journal of Plant Biology. 2024; 15(3):586-598. https://doi.org/10.3390/ijpb15030044
Chicago/Turabian StyleBreman, Floris C., Joost Korver, Ronald Snijder, M. Eric Schranz, and Freek T. Bakker. 2024. "Inheritance of Mitochondria in Pelargonium Section Ciconium (Sweet) Interspecific Crosses" International Journal of Plant Biology 15, no. 3: 586-598. https://doi.org/10.3390/ijpb15030044