1. Introduction
Perennial crops can regrow after normal harvest, and have been adopted as part of the global toolkit for climate change mitigation and food security in the long term [
1,
2]. The potential benefits of perennial crops in sustainable farming systems are now drawing the attention of scientific researchers and government officials, not only because of the likely positive ecological effects on landscape and biodiversity conservation, but also the likely significant economic benefits for smallholder farmers [
3,
4,
5,
6]. In particular, the rice-growing lands in Asia are largely threatened due to pressure on soil resources [
1]. Thus, the development of high-yielding perennial rice cultivars would address the environmental limitations of annual rice while helping to feed the rapidly increasing human population.
With a successful wide hybrid being reported between the wild perennial rice
Oryza longistaminata and the annual rice
Oryza sativa [
7], it was originally proposed that perennial rice should be developed in order to stabilize fragile upland farming systems. Greater understanding of the genetic architecture of perenniality [
8,
9] was obtained from the study of viable progeny from the wide hybrid segregating for perenniality [
10,
11]. This led to proposals to introgress additional traits from the wild perennial species into the annual cultivated rice germplasm, and to the perennial rice breeding programs at Yunnan Academy of Agricultural Sciences and Yunnan University.
Rice is one of the most important crops grown worldwide, so the opportunity for the successful development of perennial rice has great potential. Viable progeny from the wide-hybrid segregating for perenniality also acquired nematode resistance and drought tolerance from the wild species, through linkage drag [
10,
11]. For the development of perennial rice to stabilize the fragile soils of rice-based farming systems, perennial rice breeding using derivatives of the original wide-hybrid and research on the genetic control of perenniality in rice have been continued [
1,
8,
9,
10,
11]. These efforts offer the opportunity not only for the commercial use of perennial rice, but also for further understanding of the genetic architecture of perenniality in rice.
A successful perennial rice breeding program has been established in the Yunnan Academy of Agricultural Sciences and Yunnan University, with the high-yielding and broadly-adapted experimental line PR23 recommended for pre-release testing under paddy conditions in southern China and Laos [
12,
13]. Consequently, this paper reports the field evaluation of PR23 in comparison with the main conventional rice cultivars in pre-release testing under paddy conditions in nine ecological regions of Yunnan Province in China, between 2011 and 2017. The objectives were: (1) to compare the survival, regrowth, field performance, and adaptation of perennial rice PR23 with two conventional rice varieties across nine ecological regions of Yunnan; (2) to consider the economic costs and benefits in cultivation of perennial rice relative to annual rice; and (3) to discuss evidence for the commercial release of PR23 as a high-yielding and broadly-adapted perennial rice cultivar for farmers in the Yunnan Province of China.
3. Results
Throughout the duration of the study, long-term mean monthly maximum temperature was generally favourable at all sites, exceeding 30 °C only at Honghe in August and Jing Hong in February–March (
Supplementary Table S1). In contrast, long-term mean monthly minimum temperature was below 15 °C for 3 months in Jing Hong (550 m), 4 months in Honghe (1300 m), 5 months in Wenshang and Dehong (900 and 1260 m), 6 months in Lancang and Mengzhe (1150 and 1255 m), and 7 months in Puer and Yiliang (1305 and 1600 m), respectively (
Supplementary Table S2). Overall, Jinghong at the lowest altitude was warmest with higher evaporative demand, and Yiliang at the highest altitude was coldest with lower evaporative demand (
Supplementary Table S1). Rainfall was higher in 2017 than 2016 at all sites, with December–April generally drier than May–November (
Supplementary Table S2). Soils were generally mildly acidic and sufficient in soil organic matter, total N, and available N, P, and K (
Table 3).
Data were available for G × E analysis from three genotypes at 23 environments (Experiment 1), 12 environments (Experiment 2), and 35 environments (Combined) (
Table 1 and
Supplementary Table S3). Site mean yield ranged from 4.81 to 9.71 t ha
−1, with yields in the first season generally higher than in the second season, and with yields gradually declining in successive crops (
Table 1). Field duration ranged from 100 to 169 days, with longer durations generally at cooler, higher-altitude sites (
Table 1). The three genotypes were quite similar in mean field duration and mean grain yield, averaging 108 days and 6.12 t ha
−1, respectively, but only PR23 was able to successfully regrow in subsequent seasons, averaging 89.8% regrowth over its 23 crops (
Table 2).
Data were analysed separately for Experiments 1 and 2, as well as combined over all environments. In all three analyses, genotype main effects accounted for less than 5% of the total sum of squares, environment main effects accounted for more than 70% of the total sum of squares, while the genotype by environment interactions accounted for 11.4%, 17.3%, and 25.7% of the total sum of squares, for three genotypes by 12, 23, and 35 environments, respectively.
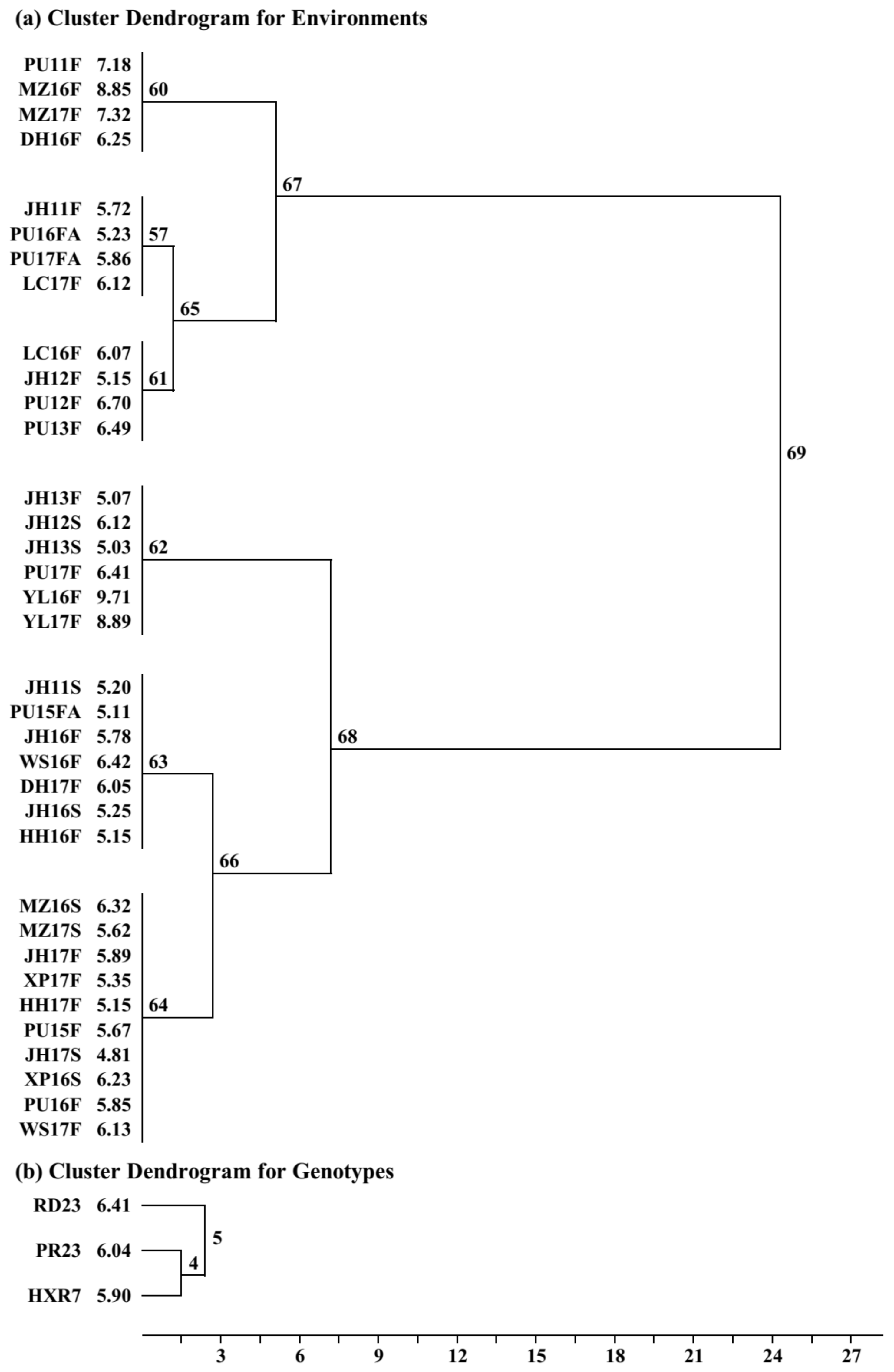
Membership of environment groupings from the three analyses is shown in
Table 4, with groupings aligned by environment membership. Remarkably, groupings from the separate analyses are retained in the combined analysis, with two groups in combined (E58 and E56) being composed solely of groups from Experiment 1 (E38 and E36), one group in combined (E34) being composed solely of one group from Experiment 2 (E11), and the remainder combining groups from both Experiments 1 and 2. There were only 3 of 35 environments which were exceptions, as underlines in
Table 4. Consequently, combined analysis was chosen for interpretation of the data.
Cluster analysis on environment-standardized residuals was used to identify six environment groups for the three genotypes, which preserved 95.8% of the G × E sum of squares. The cluster dendrogram for environments (
Figure 1a) initially separated a set of 12 environments (Fusion 67) from the other 23 environments (Fusion 68). Among the 12-environment set (Fusion 67), a group of four environments separated first (Environment group E60), then the remainder split into two groups of four environments (E61 and E57). Likewise, among the 23-environment set (Fusion 68), a group of six environments separated first (E62), then the remainder split into groups of seven and ten (E63 and E64), respectively. Membership of these groups is shown in
Table 4, with E62 comprising E34 and E59, and E64 comprising E58 and E56. Although RD23 separated from PR23 and HRX7 in the cluster dendrogram for genotypes (
Figure 1b), all three genotypes were retained for interpretation. Consequently, cluster analysis reduced the matrix from 3 genotypes × 35 individual environments (=105) to 3 genotypes × 6 environment groups (=18), whilst retaining the repeatable G × E variation (95.8%) for interpretation.
Grain yields, growth durations and regrowth percentages are shown for all three genotypes across each of six environmental groups in
Table 5,
Table 6 and
Table 7, respectively. On average, grain yields were highest (7.40 t ha
−1) in E60 (PU11F, MZ16F, MZ17F, DH16F) and lowest (5.66 t ha
−1) in E64 (ten sites), E63 (seven sites), and E57 (four sites) (
Table 5). RD23 yielded 6.41 t ha
−1 on average, was highest yielding in E62, E64, and E63, and was lowest yielding in E61 and E60. HXR7 yielded 5.90 t ha
−1 on average, was highest yielding in E61 and E57, but lowest in E64 and E63. In contrast, PR23 was more stable in grain yield, averaging 6.04 t ha
−1, and was generally intermediate in yield, except in E62 and E60, where it ranked third and first, respectively. On average, growth duration was longest at E60, where yields were highest (
Table 6). HXR7 was longer in growth duration (151–171 days), RD23 was intermediate (145–161 days), and PR23 was shortest in growth duration (137–147 days), except in E60, where PR23 took 163 days and was highest yielding (8.49 t ha
−1). PR23 successfully regrew in all six environment groups, whereas RD23 and HXR7 did not (
Table 7).
Cost–benefit analysis of perennial and annual rice across eight locations in Yunnan in Experiment 1 was compared by location (
Table 8) and by growth habit and season (
Table 9). Although locations did not differ significantly in cost–benefit per unit of investment, cost–benefit per unit of labour, or profit per unit of investment, locations did differ significantly in profit per day (
Table 8), with Yiliang (226 Yuan/d) being the most profitable, and Puer the least profitable (118.5 Yuan/d). In contrast, all four parameters were statistically significant for the interaction between growth habit and season (
Table 9). In each case, the ratoon crop of PPR23 was more profitable than the re-sown crop of HXR7 in the second season. As a consequence, the second crop was more profitable than the first, and PR23 was more profitable than HXR7 on average. Thus, allowing the perennial rice PR23 to ratoon resulted in greater profit, especially per unit of labour, due to substantial savings in the amount and cost of labour.
Different traits of perennial rice PR23 and annual rice RD23 and HXR7 are presented in
Table 10. The grain yield of PR23 (7.05 t ha
−1) was significantly higher than RD23 (5.69 t ha
−1) and HXR7 (5.89 t ha
−1) in Experiment 1 in both seasons. The major causes for this result were the lower number of panicles and the higher 1000-grain weight of PR23 in comparison with RD23 and HXR7. However, the number of spikelets per panicle of PR23 was slightly fewer than those of RD23 and HXR7. The growth duration of PR23 (135 d) was shorter than RD23 (152 d) and HXR7 (152 d) in the second season, though they were similar in the first season (157 d). Panicle length of PR23 (20.0 cm) was less than RD23 (25.4 cm) and HXR7 (22.9 cm) in both seasons. There was no significant difference in plant height. The duration of flowering in PR23 was longer than RD23 and HXR7 in both seasons, which may be conducive to adequate pollination and fertilization in PR23. Grain quality of PR23 was similar to RD23, although it ranked behind HXR7, which farmers prefer. Nevertheless, the cooking quality of PR23 was comparable with HXR7, and higher than RD23. In addition, PR23 was preferred by millers due to its high rice yield and high milling percentage (73%). Overall, farmers preferred PR23 due to the savings of labour and decreased intensity of labour, as a result of not having to till and replant each season. Thus, PR23 had obvious economic benefits, resulting in its growing popularity with farmers.
Finally, perennial rice PR23 was successfully grown and mechanically harvested from four large demonstration areas of 1.0 to 13.0 ha on-farm in southern Yunnan (
Table 11). Total grain yield of PR23 per year exceeded 13 t/ha in each demonstration area, with Mengzhe yielding 17.4 t ha
−1 in 2006 from seasons 1 and 2, and 15.1 t ha
−1 in 2017 from seasons 3 and 4. Ratoon percentage of PR23 ranged from 90.0–98.2%, including 93.3% and 90.0% in seasons 3 and 4 at Mengzhe, indicating its strong perenniality across a range of irrigated environments. Grain yield of PR23 was higher in the first season than in the second season, due to the shorter growth duration, fewer panicles, and fewer spikelets per panicle in the second season. These results confirmed that PR23 produced high yield, excellent regrowth, and adaptability when grown at commercial scale on-farm. Consequently, there has been an upsurge in demand for PR23 among local subsistence farmers and large commercial growers, indicating a bright future for perennial rice production and application across wider areas. This evidence is consistent with the need to release PR23 to farmers in Yunnan, as the first-ever perennial rice grain crop.



{kind=link}