Impacts of Agricultural Management Systems on Biodiversity and Ecosystem Services in Highly Simplified Dryland Landscapes

 ,
,  ,
,
Abstract
:1. Background
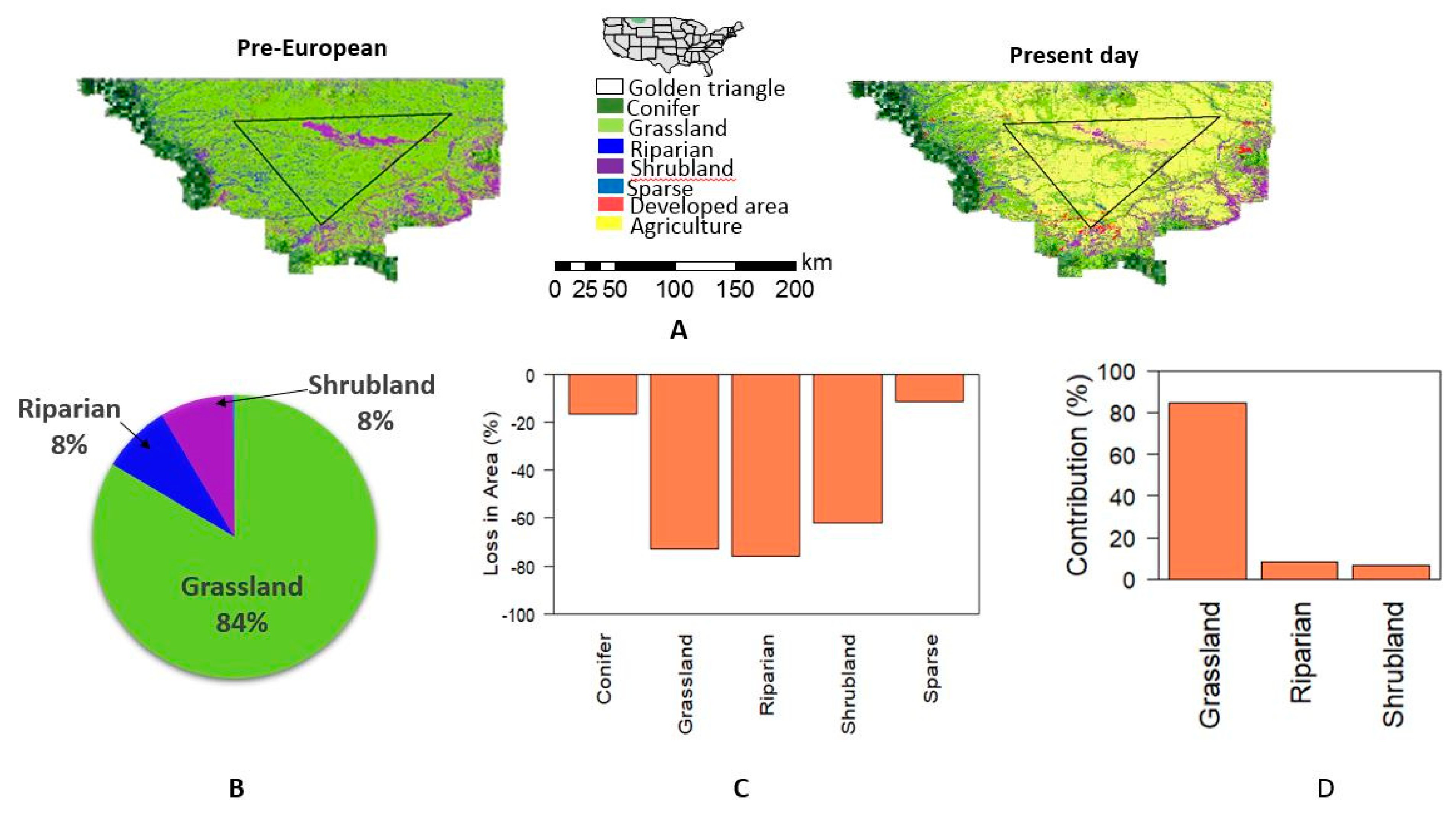
2. A Case Study of Land Use Changes due to Agricultural Expansion in the Drylands of the Northern Great Plains
3. Impacts of Agricultural Management Systems on Biodiversity and Ecosystem Services in Highly Simplified Drylands
3.1. Management Systems and Associated Plant Diversity
3.2. Management Systems, Pest Infestation, Parasitoid Abundance, and Pest Regulation Services
3.3. Management Systems and Bee Colony Success
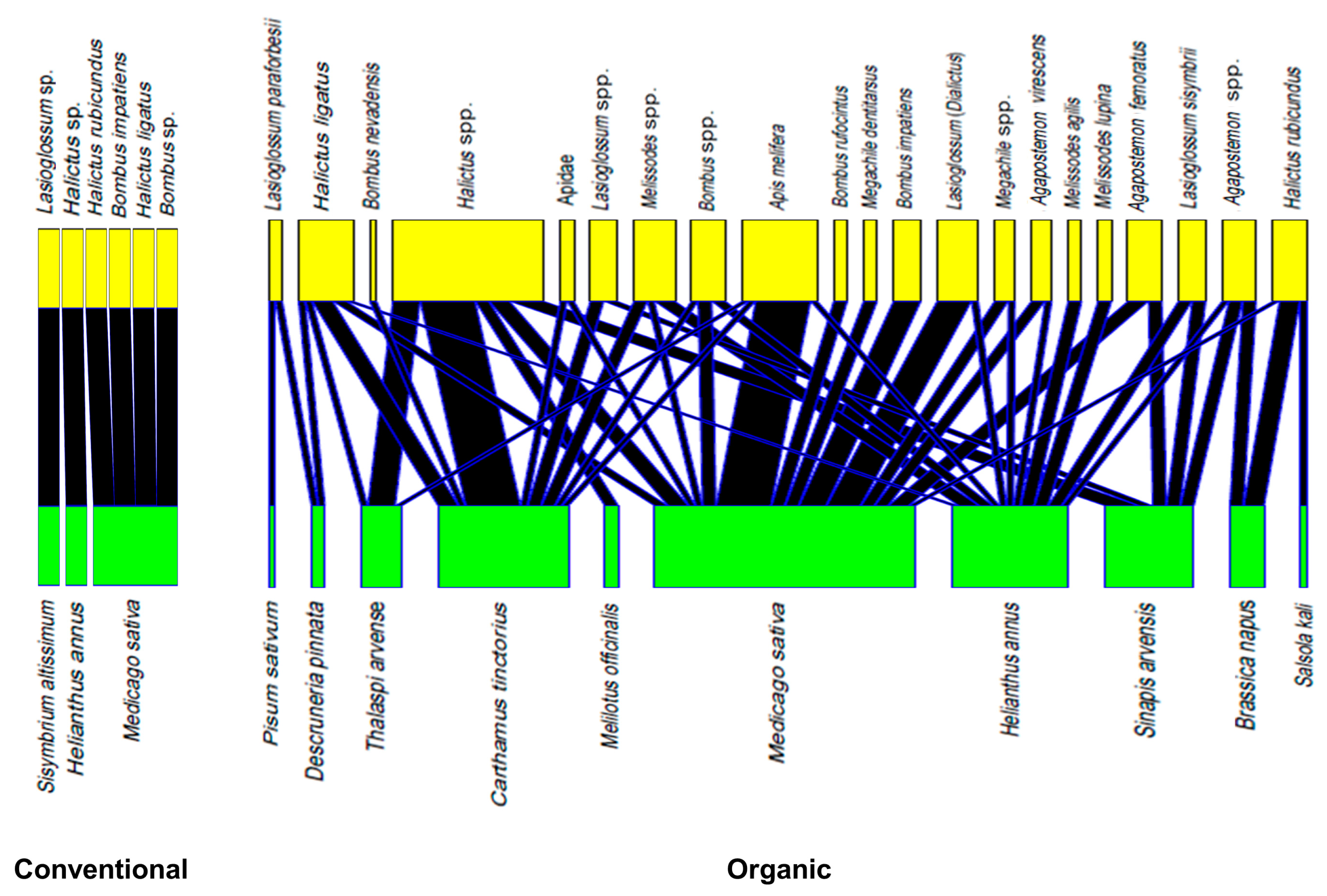
3.4. Management Systems and Bee–Flower Networks
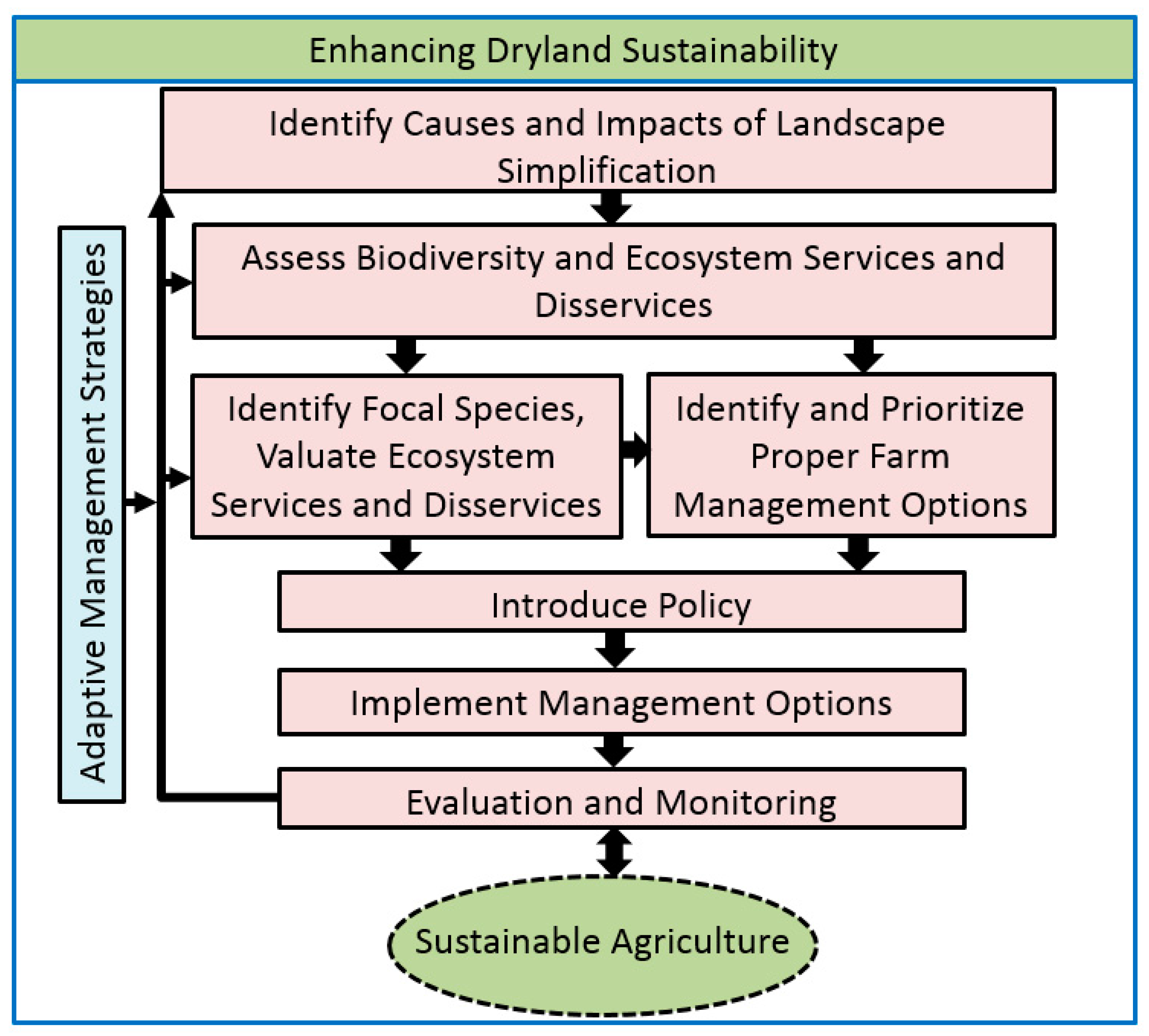
4. Discussion: Enhancing the Sustainability of Dryland Agroecosystems
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- UNCCD. United Nations Convention to Combat Desertification in Those Countries Experiencing Serious Drought and/or Desertification, Particularly in Africa; United Nations Environment Programmes/Convention to Combat Desertification: Geneva, Switzerland, 1994. [Google Scholar]
- Feng, S.; Fu, Q. Expansion of global drylands under a warming climate. Atmos. Chem. Phys. 2013, 13, 10081–10094. [Google Scholar] [CrossRef] [Green Version]
- Yirdaw, E.; Tigabu, M.; Monge, A. Rehabilitation of degraded dryland ecosystems—Review. Silva Fenn. 2017, 51. [Google Scholar] [CrossRef]
- Puigdefábregas, J. Ecological impacts of global change on drylands and their implications for desertification. Land Degrad. Dev. 1998, 9, 393–406. [Google Scholar] [CrossRef]
- Cowie, A.L.; Penman, T.D.; Gorissen, L.; Winslow, M.D.; Lehmann, J.; Tyrrell, T.D.; Twomlow, S.; Wilkes, A.; Lal, R.; Jones, J.W.; et al. Towards sustainable land management in the drylands: Scientific connections in monitoring and assessing dryland degradation, climate change and biodiversity. Land Degrad. Dev. 2011, 22, 248–260. [Google Scholar] [CrossRef]
- Sherwood, S.; Fu, Q. A Drier Future? Climate Change. Science 2014, 343. [Google Scholar] [CrossRef] [PubMed]
- Steffan-Dewenter, I.; Münzenberg, U.; Tscharntke, T. Pollination, seed set and seed predation on a landscape scale. Proc. Biol. Sci. 2001, 268, 1685–1690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaplin-Kramer, R.; O’Rourke, M.E.; Blitzer, E.J.; Kremen, C. A meta-analysis of crop pest and natural enemy response to landscape complexity. Ecol. Lett. 2011, 14, 922–932. [Google Scholar] [CrossRef] [PubMed]
- Darkoh, M.B.K. Regional perspectives on agriculture and biodiversity in the drylands of Africa. J. Arid Environ. 2001, 54, 261–279. [Google Scholar] [CrossRef]
- Hansen, A.J.; Rasker, R.; Maxwell, B.; Rotella, J.J.; Johnson, J.D.; Parmenter, A.W.; Langner, U.; Cohen, W.B.; Lawrence, R.L.; Kraska, M.P.V. Ecological Causes and Consequences of Demographic Change in the New WestAs natural amenities attract people and commerce to the rural west, the resulting land-use changes threaten biodiversity, even in protected areas, and challenge efforts to sustain local communities and ecosystems. Bioscience 2002, 52, 151–162. [Google Scholar] [CrossRef]
- Koohafkan, P.; Stewart, B.A. Water and Cereals in Drylands. Available online: http://www.fao.org/3/i0372e/i0372e.pdf (accessed on 1 June 2019).
- Thomas, R.J.; Akhtar-Schuster, M.; Stringer, L.C.; Marques, M.J.; Escadafal, R.; Abraham, E.; Enne, G. Fertile ground? Options for a science-policy platform for land. Environ. Sci. Policy 2012, 16, 122–135. [Google Scholar] [CrossRef]
- Reynolds, J.F.; Mark, D.; Smith, S.; Lambin, E.F.; Ii, B.L.T.; Mortimore, M.; Batterbury, S.P.J.; Downing, T.E.; Dowlatabadi, H.; Fernández, R.J.; et al. Global Desertification: Building a Science for Dryland Development. Science 2007, 316, 847–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brittain, C.A.; Vighi, M.; Bommarco, R.; Settele, J.; Potts, S.G. Impacts of a pesticide on pollinator species richness at different spatial scales. Basic Appl. Ecol. 2010, 11, 106–115. [Google Scholar] [CrossRef]
- Hole, D.G.; Perkins, A.J.; Wilson, J.D.; Alexander, I.H.; Grice, P.V.; Evans, A.D. Does organic farming benefit biodiversity? Biol. Conserv. 2005, 122, 113–130. [Google Scholar] [CrossRef]
- Tscharntke, T.; Klein, A.M.; Kruess, A.; Steffan-Dewenter, I.; Thies, C. Landscape perspectives on agricultural intensification and biodiversity—Ecosystem service management. Ecol. Lett. 2005, 8, 857–874. [Google Scholar] [CrossRef]
- Kremen, C.; Williams, N.M.; Thorp, R.W. Crop pollination from native bees at risk from agricultural intensification. Proc. Natl. Acad. Sci. USA 2002, 99, 16812–16816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kremen, C.; Williams, N.M.; Bugg, R.L.; Fay, J.P.; Thorp, R.W. The area requirements of an ecosystem service: Crop pollination by native bee communities in California. Ecol. Lett. 2004, 7, 1109–1119. [Google Scholar] [CrossRef]
- Williams, N.M.; Crone, E.E.; Roulston, T.H.; Minckley, R.L.; Packer, L.; Potts, S.G. Ecological and life-history traits predict bee species responses to environmental disturbances. Biol. Conserv. 2010, 143, 2280–2291. [Google Scholar] [CrossRef]
- Isaacs, R.; Kirk, A.K. Pollination services provided to small and large highbush blueberry fields by wild and managed bees. J. Appl. Ecol. 2010, 47, 841–849. [Google Scholar] [CrossRef]
- Isaacs, R.; Tuell, J.; Fiedler, A.; Gardiner, M.; Landis, D. Maximizing arthropod-mediated ecosystem services in agricultural landscapes: The role of native plants. Front. Ecol. Environ. 2009, 7, 196–203. [Google Scholar] [CrossRef]
- Dainese, M.; Martin, E.A.; Aizen, M.; Albrecht, M.; Bartomeus, I.; Bommarco, R.; Carvalheiro, L.G.; Chaplin-Kramer, R.; Gagic, V.; Garibaldi, L.A.; et al. A global synthesis reveals biodiversity-mediated benefits for crop production. bioRxiv 2019, 554170. [Google Scholar] [CrossRef] [Green Version]
- Hansen, N.C.; Allen, B.L.; Anapalli, S.; Blackshaw, R.E.; Lyon, D.J.; Machado, S. Dryland Agriculture in North America. In Innovations in Dryland Agriculture; Springer International Publishing: Cham, Switzerland, 2016; pp. 415–441. [Google Scholar]
- Adhikari, S.; Burkle, L.A.; O’Neill, K.M.; Weaver, D.K.; Menalled, F.D. Dryland organic farming increases floral resources and bee colony success in highly simplified agricultural landscapes. Agric. Ecosyst. Environ. 2019, 270–271, 9–18. [Google Scholar] [CrossRef]
- Adhikari, S.; Seipel, T.; Menalled, F.D.; Weaver, D.K. Farming system and wheat cultivar affect infestation of, and parasitism on, Cephus cinctus in the Northern Great Plains. Pest Manag. Sci. 2018, 74, 2480–2487. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, S.; Menalled, F. Impacts of Dryland Farm Management Systems on Weeds and Ground Beetles (Carabidae) in the Northern Great Plains. Sustainability 2018, 10, 2146. [Google Scholar] [CrossRef]
- Huang, J.; Yu, H.; Guan, X.; Wang, G.; Guo, R. Accelerated dryland expansion under climate change. Nat. Clim. Chang. 2016, 6, 166–171. [Google Scholar] [CrossRef]
- Millennium Ecosystem Assessment. Ecosystems and Human Well-Being: Wetlands and Water; World Resources Institute: Washington, DC, USA, 2005. [Google Scholar]
- Brester, G.W.; Grant, B.; Boland, M.A. Marketing Organic Pasta from Big Sandy to Rome: It’s a Long Kamut®. Rev. Agric. Econ. 2009, 31, 359–369. [Google Scholar] [CrossRef]
- Johnston, A.M.; Tanaka, D.L.; Miller, P.R.; Brandt, S.A.; Nielsen, D.C.; Lafond, G.P.; Riveland, N.R. Oilseed Crops for Semiarid Cropping Systems in the Northern Great Plains. Agron. J. 2002, 94, 231–240. [Google Scholar] [CrossRef] [Green Version]
- North Dakota Wheat Commission. North Dakota Wheat Commission: Building Bigger Better Markets. Available online: http://www.ndwheat.com/default.asp (accessed on 29 October 2018).
- Montana Agricultural Statistics. USDA—National Agricultural Statistics Service and Montana Department of Agriculture; Montana Agricultural Statistics, USDA: Washington, DC, USA, 2016.
- Lakkakula, P.; Olson, F.; Ripplinger, D. Pea and Lentil Market Analysis; North Dakota State University: Fargo, ND, USA, 2017. [Google Scholar]
- Barrett, S.; Havlina, D.; Jones, J.; Hann, W.; Frame, C.; Hamilton, D.; Schon, K.; Demeo, T.; Hutter, L.; Menakis, J. Interagency Fire Regime Condition Class Guidebook. Version 3.0. Available online: https://www.landfire.gov/frcc/frcc_guidebooks.php (accessed on 29 October 2018).
- Pollnac, F.W.; Rew, L.J.; Maxwell, B.D.; Menalled, F.D. Spatial patterns, species richness and cover in weed communities of organic and conventional no-tillage spring wheat systems. Weed Res. 2008, 48, 398–407. [Google Scholar] [CrossRef]
- Danner, N.; Molitor, A.M.; Schiele, S.; Härtel, S.; Steffan-Dewenter, I. Season and landscape composition affect pollen foraging distances and habitat use of honey bees. Ecol. Appl. 2016. [Google Scholar] [CrossRef]
- Landis, D.A.; Menalled, F.D.; Costamagna, A.C.; Wilkinson, T.K. Manipulating plant resources to enhance beneficial arthropods in agricultural landscapes. Weed Sci. 2005, 53, 902–908. [Google Scholar] [CrossRef]
- Pywell, R.F.; Warman, E.A.; Carvell, C.; Sparks, T.H.; Dicks, L.V.; Bennett, D.; Wright, A.; Critchley, C.N.R.; Sherwood, A. Providing foraging resources for bumblebees in intensively farmed landscapes. Biol. Conserv. 2005, 121, 479–494. [Google Scholar] [CrossRef]
- Nicholls, C.I.; Altieri, M.A. Plant biodiversity enhances bees and other insect pollinators in agroecosystems. A review. Agron. Sustain. Dev. 2013, 33, 257–274. [Google Scholar] [CrossRef]
- Losey, J.E.; Vaughn, M. The economic value of ecological services provided by insects. Bioscience 2006, 56, 311–323. [Google Scholar] [CrossRef]
- Barbosa, P. Agroecosystems and conservation biological control. In Conservation Biological Control; Academic Press: San Diego, CA, USA, 1998; pp. 39–54. [Google Scholar]
- Landis, D.A. Designing agricultural landscapes for biodiversity-based ecosystem services. Basic Appl. Ecol. 2017, 18, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Power, A.G. Ecosystem services and agriculture: Tradeoffs and synergies. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2010, 365, 2959–2971. [Google Scholar] [CrossRef] [PubMed]
- Lesieur, V.; Martin, J.-F.; Weaver, D.K.; Hoelmer, K.A.; Smith, D.R.; Morrill, W.L.; Kadiri, N.; Peairs, F.B.; Cockrell, D.M.; Randolph, T.L.; et al. Phylogeography of the Wheat Stem Sawfly, Cephus cinctus Norton (Hymenoptera: Cephidae): Implications for Pest Management. PLoS ONE 2016, 11, e0168370. [Google Scholar] [CrossRef] [PubMed]
- Beres, B.L.; Dosdall, L.M.; Weaver, D.K.; Cárcamo, H.A.; Spaner, D.M. Biology and integrated management of wheat stem sawfly and the need for continuing research. Can. Entomol. 2011, 143, 105–125. [Google Scholar] [CrossRef]
- Weaver, D.K.; Nansen, C.; Runyon, J.B.; Sing, S.E.; Morrill, W.L. Spatial distributions of Cephus cinctus Norton (Hymenoptera: Cephidae) and its braconid parasitoids in Montana wheat fields. Biol. Control 2005, 34, 1–11. [Google Scholar] [CrossRef]
- Peterson, R.K.D.; Buteler, M.; Weaver, D.K.; Macedo, T.B.; Sun, Z.; Perez, O.G.; Pallipparambil, G.R. Parasitism and the demography of wheat stem sawfly larvae, Cephus cinctus. Biol. Control 2011, 56, 831–839. [Google Scholar] [CrossRef]
- Bekkerman, A.; Weaver, D.K. Modeling Joint Dependence of Managed Ecosystems Pests: The Case of the Wheat Stem Sawfly. J. Agric. Resour. Econ. 2018, 43, 172–194. [Google Scholar]
- Morrill, W.L.; Kushnak, G.D.; Gabor, J.W. Parasitism of the Wheat Stem Sawfly (Hymenoptera: Cephidae) in Montana. Biol. Control 1998, 12, 159–163. [Google Scholar] [CrossRef]
- Runyon, J.B.; Hurley, R.L.; Morrill, W.L.; Weaver, D.K. Distinguishing adults of Bracon cephi and Bracon lissogaster (Hymenoptera: Braconidae), parasitoids of the wheat stem sawfly (Hymenoptera: Cephidae). Can. Entomol. 2001, 133, 215–217. [Google Scholar] [CrossRef]
- Runyon, J.B.; Morrill, W.L.; Weaver, D.K.; Miller, P.R. Parasitism of the Wheat Stem Sawfly (Hymenoptera: Cephidae) by Bracon cephi and B. lissogaster (Hymenoptera: Braconidae) in Wheat Fields Bordering Tilled and Untilled Fallow in Montana. J. Econ. Entomol. 2002, 95, 1130–1134. [Google Scholar] [CrossRef] [PubMed]
- Weaver, D.K.; Sing, S.E.; Runyon, J.B.; Morrill, W.L.; Weaver, D. Potential Impact of Cultural Practices on Wheat Stem Sawfly (Hymenoptera: Cephidae) and Associated Parasitoids. J. Agric. Urban Entomol. 2004, 21, 271–287. [Google Scholar]
- Buteler, M.; Weaver, D.K.; Miller, P.R. Wheat stem sawfly-infested plants benefit from parasitism of the herbivorous larvae. Agric. For. Entomol. 2008, 10, 347–354. [Google Scholar] [CrossRef]
- Rand, T.A.; Waters, D.K.; Blodgett, S.L.; Knodel, J.J.; Harris, M.O. Increased area of a highly suitable host crop increases herbivore pressure in intensified agricultural landscapes. Agric. Ecosyst. Environ. 2014, 186, 135–143. [Google Scholar] [CrossRef] [Green Version]
- Tscharntke, T.; Karp, D.S.; Chaplin-Kramer, R.; Batáry, P.; DeClerck, F.; Gratton, C.; Hunt, L.; Ives, A.; Jonsson, M.; Larsen, A.; et al. When natural habitat fails to enhance biological pest control—Five hypotheses. Biol. Conserv. 2016, 204, 449–458. [Google Scholar] [CrossRef]
- Karp, D.S.; Chaplin-Kramer, R.; Meehan, T.D.; Martin, E.A.; DeClerck, F.; Grab, H.; Gratton, C.; Hunt, L.; Larsen, A.E.; Martínez-Salinas, A.; et al. Crop pests and predators exhibit inconsistent responses to surrounding landscape composition. Proc. Natl. Acad. Sci. USA 2018, 115, E7863–E7870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Settele, J.; Settle, W.H. Conservation biological control: Improving the science base. Proc. Natl. Acad. Sci. USA 2018, 115, 8241–8243. [Google Scholar] [CrossRef] [Green Version]
- NASS. 2016 Wheat Varieties Grown in Montana; NASS: Washington, DC, USA, 2016.
- Kevan, P.G. Pollinators as bioindicators of the state of the environment: Species, activity and diversity. Agric. Ecosyst. Environ. 1999, 74, 373–393. [Google Scholar] [CrossRef]
- Moisset, B.B.; Buchmann, S. Bee Basics: An Introduction to Our Native Bees; Lulu.com: Morrisville, NC, USA, 2011. [Google Scholar]
- Goulson, D.; Nicholls, E.; Botias, C.; Rotheray, E.L. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 2015, 347, 1255957. [Google Scholar] [CrossRef]
- Goulson, D.; Lye, G.C.; Darvill, B. Decline and Conservation of Bumble Bees. Annu. Rev. Entomol. 2008, 53, 191–208. [Google Scholar] [CrossRef] [PubMed]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Kremen, C.; Miles, A. Ecosystem Services in Biologically Diversified versus Conventional Farming Systems: Benefits, Externalitites, and Trade-Offs. Ecol. Soc. 2012, 17, 1–23. [Google Scholar] [CrossRef]
- Gallai, N.; Salles, J.M.; Settele, J.; Vaissière, B.E. Economic valuation of the vulnerability of world agriculture confronted with pollinator decline. Ecol. Econ. 2009, 68, 810–821. [Google Scholar] [CrossRef]
- Adhikari, S. Impacts of Dryland Farming Systems on Biodiversity, Plant-Insect Interactions, and Ecosystem Services; Montana State University: Bozeman, MT, USA, 2018. [Google Scholar]
- Adhikari, S.; Burkle, L.A.; O’Neill, K.M.; Weaver, D.K.; Delphia, C.M.; Menalled, F.D. Dryland Organic Farming Partially Offsets Negative Effects of Highly Simplified Agricultural Landscapes on Forbs, Bees, and Bee–Flower Networks. Environ. Entomol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Kehinde, T.; Samways, M.J. Insect-flower interactions: Network structure in organic versus conventional vineyards. Anim. Conserv. 2014, 17, 401–409. [Google Scholar] [CrossRef]
- Power, E.F.; Stout, J.C. Organic dairy farming: Impacts on insect-flower interaction networks and pollination. J. Appl. Ecol. 2011, 48, 561–569. [Google Scholar] [CrossRef]
- Tucker, E.M.; Rehan, S.M. Wild Bee Community Assemblages Across Agricultural Landscapes. J. Agric. Urban Entomol. 2017, 33, 77–104. [Google Scholar] [CrossRef]
- Spiesman, B.J.; Bennett, A.; Isaacs, R.; Gratton, C. Bumble bee colony growth and reproduction depend on local flower dominance and natural habitat area in the surrounding landscape. Biol. Conserv. 2017, 206, 217–223. [Google Scholar] [CrossRef] [Green Version]
- Landis, D.A.; Wratten, S.D.; Gurr, G.M. Habitat Management to Conserve Natural Enemies of Arthropod Pests in Agriculture. Annu. Rev. Entomol. 2000, 45, 175–201. [Google Scholar] [CrossRef]
- Rusch, A.; Chaplin-Kramer, R.; Gardiner, M.M.; Hawro, V.; Holland, J.; Landis, D.; Thies, C.; Tscharntke, T.; Weisser, W.W.; Winqvist, C.; et al. Agricultural landscape simplification reduces natural pest control: A quantitative synthesis. Agric. Ecosyst. Environ. 2016, 221, 198–204. [Google Scholar] [CrossRef] [Green Version]
- Letourneau, D.K.; Bothwell Allen, S.G.; Stireman, J.O. Perennial habitat fragments, parasitoid diversity and parasitism in ephemeral crops. J. Appl. Ecol. 2012, 49, 1405–1416. [Google Scholar] [CrossRef]
- Chaplin-Kramer, R.; de Valpine, P.; Mills, N.J.; Kremen, C. Detecting pest control services across spatial and temporal scales. Agric. Ecosyst. Environ. 2013, 181, 206–212. [Google Scholar] [CrossRef] [Green Version]
- Philip Robertson, G.; Gross, K.L.; Hamilton, S.K.; Landis, D.A.; Schmidt, T.M.; Snapp, S.S.; Swinton, S.M. Farming for Ecosystem Services: An Ecological Approach to Production Agriculture. Bioscience 2014, 64, 404–415. [Google Scholar] [CrossRef] [PubMed]
- Mallinger, R.E.; Gibbs, J.; Gratton, C. Diverse landscapes have a higher abundance and species richness of spring wild bees by providing complementary floral resources over bees’ foraging periods. Landsc. Ecol. 2016, 31, 1523–1535. [Google Scholar] [CrossRef]
- Walther-Hellwig, K.; Frankl, R. Foraging habitats and foraging distances of bumblebees, Bombus spp. (Hym., Apidae), in an agricultural landscape. J. Appl. Entomol. 2000, 124, 299–306. [Google Scholar] [CrossRef]
- Morandin, L.A.; Winston, M.L.; Abbott, V.A.; Franklin, M.T. Can pastureland increase wild bee abundance in agriculturally intense areas? Basic Appl. Ecol. 2007, 8, 117–124. [Google Scholar] [CrossRef]
- Morandin, L.A.; Kremen, C. Hedgerow restoration promotes pollinator populations and exports native bees to adjacent fields. Ecol. Appl. 2013, 23, 829–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morandin, L.A.; Winston, M.L. Pollinators provide economic incentive to preserve natural land in agroecosystems. Agric. Ecosyst. Environ. 2006, 116, 289–292. [Google Scholar] [CrossRef]
- Peterson, C.A.; Eviner, V.T.; Gaudin, A.C.M. Ways forward for resilience research in agroecosystems. Agric. Syst. 2018, 162, 19–27. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Ricketts, T.H.; Kremen, C.; Carney, K. Ecosystem services and dis-services to agriculture. Ecol. Econ. 2007, 64, 253–260. [Google Scholar] [CrossRef] [Green Version]
- Duru, M.; Therond, O.; Martin, G.; Martin-Clouaire, R.; Magne, M.-A.; Justes, E.; Journet, E.-P.; Aubertot, J.-N.; Savary, S.; Bergez, J.-E.; et al. How to implement biodiversity-based agriculture to enhance ecosystem services: A review. Agron. Sustain. Dev. 2015, 35, 1259–1281. [Google Scholar] [CrossRef]
- Sandifer, P.A.; Sutton-Grier, A.E.; Ward, B.P. Exploring connections among nature, biodiversity, ecosystem services, and human health and well-being: Opportunities to enhance health and biodiversity conservation. Ecosyst. Serv. 2015, 12, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Birgé, H.E.; Allen, C.R.; Garmestani, A.S.; Pope, K.L. Adaptive management for ecosystem services. J. Environ. Manag. 2016, 183, 343–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, B.B. Resilience in Agriculture through Crop Diversification: Adaptive Management for Environmental Change. Bioscience 2011, 61, 183–193. [Google Scholar] [CrossRef] [Green Version]
- Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services (IPBES); IPBES Secretariat, U.C.G. IPBES Global Assessment Report. Available online: https://www.ipbes.net/news/ipbes-global-assessment-preview (accessed on 2 June 2019).
- Global Drylands: A UN System-Wide Response; United Nations Environment Management Groups: Geneva, Switzerland, 2011.
- Farooq, M.; Siddique, K.H.M. Innovations in Dryland Agriculture. In Innovations in Dryland Agriculture; Springer International Publishing: Cham, Switzerland, 2016; pp. 215–236. [Google Scholar]
- Lu, N.; Wang, M.; Ning, B.; Yu, D. Research advances in ecosystem services in drylands under global environmental changes. Curr. Opin. Environ. Sustain. 2018, 33, 92–98. [Google Scholar] [CrossRef]
- Schild, J.E.M.; Vermaat, J.E.; de Groot, R.S.; Quatrini, S.; van Bodegom, P.M. A global meta-analysis on the monetary valuation of dryland ecosystem services: The role of socio-economic, environmental and methodological indicators. Ecosyst. Serv. 2018, 32, 78–89. [Google Scholar] [CrossRef] [Green Version]
- Schild, J.E.M.; Vermaat, J.E.; van Bodegom, P.M. Differential effects of valuation method and ecosystem type on the monetary valuation of dryland ecosystem services: A quantitative analysis. J. Arid Environ. 2018, 159, 11–21. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, P.G.; Maxwell, B.D.; Rew, L.J.; Ellis, C.; Bekkerman, A. Vulnerability of dryland agricultural regimes to economic and climatic change. Ecol. Soc. 2018, 23, art34. [Google Scholar] [CrossRef]
- Adhikari, A.; Hansen, A.J. Land use change and habitat fragmentation of wildland ecosystems of the North Central United States. Landsc. Urban Plan. 2018, 177, 196–216. [Google Scholar] [CrossRef]
- DeFries, R.S.; Foley, J.A.; Asner, G.P. Land-use choices: Balancing human needs and ecosystem function. Front. Ecol. Environ. 2004, 2, 249–257. [Google Scholar] [CrossRef]
- Altieri, M.A. The ecological role of biodiversity in agroecosystems. Agric. Ecosyst. Environ. 1999, 74, 19–31. [Google Scholar] [CrossRef] [Green Version]
- Benton, T.G.; Vickery, J.A.; Wilson, J.D. Farmland biodiversity: Is habitat heterogeneity the key? Trends Ecol. Evol. 2003, 18, 182–188. [Google Scholar] [CrossRef]
- Gaba, S.; Lescourret, F.; Boudsocq, S.; Enjalbert, J.; Hinsinger, P.; Journet, E.-P.; Navas, M.-L.; Wery, J.; Louarn, G.; Malézieux, E.; et al. Multiple cropping systems as drivers for providing multiple ecosystem services: From concepts to design. Agron. Sustain. Dev. 2015, 35, 607–623. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Plants | Family |
|---|---|
| Graminoids | |
| Koeleria macrantha (Ledeb.) J.A. Schultes | Poaceae |
| Hesperostipa comata (Trin. & Rupr.) Barkworth | Poaceae |
| Pascopyrum smithii (Rydb.) A. Love | Poaceae |
| Pseudoroegneria spicata (Pursh) A. Love | Poaceae |
| Muhlenbergia cuspidata (Torr. ex Hook.) Rydb. | Poaceae |
| Calamovilfa longifolia (Hook.) Scribn | Poaceae |
| Poa sandbergii Vasey | Poaceae |
| Schizachyrium scoparium (Michx.) Nash | Poaceae |
| Bouteloua gracilis (Willd. ex Kunth.) Lag. ex Griffiths | Poaceae |
| Nassella viridula (Trin.) Barkworth | Poaceae |
| Carex filifolia Nutt. | Cyperaceae |
| * Bromus tectorum L. | Poaceae |
| * Bromus japonicus Thunb. | Poaceae |
| * Agropyron cristatum (L.) Gaertn. | Poaceae |
| * Poa pratensis L. | Poaceae |
| Forbs | |
| Pediomelum argophyllum (Pursh) J. Grimes | Fabaceae |
| Phlox hoodii Richardson | Polemoniaceae |
| Antennaria neglecta Greene | Asteraceae |
| Heterotheca villosa (Pursh) Shinners | Asteraceae |
| Cirsium undulatum (Nutt.) Spreng. | Asteraceae |
| Penstemon grandiflorus Nutt. | Scrophulariaceae |
| Plantago elongata Pursh | Plantaginaceae |
| Lygodesmia juncea (Pursh) D. Don ex Hook. | Asteraceae |
| Comandra umbellata (L.) Nutt. | Santalaceae |
| Thermopsis rhombifolia (Nutt. ex Pursh) Nutt. ex Richardson | Fabaceae |
| Gaillardia aristata Pursh | Asteraceae |
| Achillea millefolium L. var. occidentalis DC. | Asteraceae |
| Potentilla glandulosa Lindl. | Rosaceae |
| Allium textile A. Nelson & J.F. Macbr. | Liliaceae |
| Linum lewisii Pursh | Linaceae |
| Tetraneuris acaulis (Pursh) Greene | Asteraceae |
| Sphaeralcea coccinea (Nutt.) Rydb. | Malvaceae |
| Vicia americana Muhl. ex Willd. | Fabaceae |
| Liatris punctata Hook. | Asteraceae |
| Nothocalais cuspidata (Pursh) Greene | Asteraceae |
| Symphyotrichum ericoides (L.) G.L. Nesom var. pansum (S.F. Blake) G.L. Nesom | Asteraceae |
| Machaeranthera pinnatifida (Hook.) Shinners | Asteraceae |
| Ratibida columnifera (Nutt.) Wooton & Stand | Asteraceae |
| Opuntia polyacantha Haw. | Cactaceae |
| Packera plattensis (Nutt.) W.A. Weber & Á. Löve | Asteraceae |
| Lycopodium L., spp. | Lycopodiaceae |
| Bryopsida (Limpr.) Rothm., spp. | |
| * Medicago sativa L. | Fabaceae |
| * Melilotus officinalis (L.) Lam. | Fabaceae |
| * Sinapis arvensis L. ssp. arvensis | Brassicaceae |
| * Tragopogon pratensis L. | Asteraceae |
| ** Taraxacum officinale F.H. Wigg | Asteraceae |
| ** Urtica dioica L. | Urticaceae |
| Shrubs | |
| Symphoricarpos occidentalis Hook. | Caprifoliaceae |
| Artemisia ludoviciana Nutt. | Asteraceae |
| Artemisia dracunculus L. | Asteraceae |
| Artemisia longifolia Nutt. | Asteraceae |
| Artemisia frigida Willd. | Asteraceae |
| Krascheninnikovia lanata (Pursh) A. Meeuse & Smi | Chenopodiaceae |
| Artemisia cana Pursh | Asteraceae |
| Rosa arkansana Porter | Rosaceae |
| Rosa acicularis Lindl. | Rosaceae |
| Gutierrezia sarothrae (Pursh) Britton & Rusby | Asteraceae |
| Weed Taxa | Percent Composition (Overall) | |
|---|---|---|
| Conventional | Organic | |
| Amaranthus blitoides S. Watson | <0.01 | <0.01 |
| Amaranthus retroflexus L. | <0.01 | 0.47 |
| Arabidopsis thaliana (L.) Heynh. | - | <0.01 |
| Artemisia frigida Willd. | - | <0.01 |
| Avena fatua L. * | 4.71 | 4.70 |
| Bassia scoparia (L.) A.J. Scott | 0.07 | 0.14 |
| Brassicaceae sp. | - | 0.04 |
| Bromus arvensis L. * | 0.08 | <0.01 |
| Bromus tectorum L. * | 8.40 | 0.20 |
| Buglossoides arvensis (L.) I.M. Johnst. | - | <0.01 |
| Carthamus tinctorius L. | - | 0.69 |
| Chenopodium album L. | - | 7.43 |
| Chenopodium murale L. | - | <0.01 |
| Cirsium arvense (L.) Scop. | 2.55 | <0.01 |
| Descurainia pinnata (Walter) Britton | <0.01 | 0.03 |
| Descurainia sophia (L.) Webb ex Prantl | - | 0.01 |
| Fabaceae sp. | - | <0.01 |
| Fagopyrum esculentum Moench | - | 0.03 |
| Helianthus annuus L. | <0.01 | 2.72 |
| Hordeum jubatum L.* | 5.03 | 0.04 |
| Lactuca serriola L. | 0.12 | 0.43 |
| Lens culinaris Medik. | - | 0.15 |
| Leptochloa fusca (L.) Kunth * | <0.01 | - |
| Medicago lupulina L. | - | 0.01 |
| Medicago sativa L. | 0.89 | 3.50 |
| Melilotus officinalis (L.) Lam. | - | 0.07 |
| Monolepis nuttalliana (Schult.) Greene | - | <0.01 |
| Pisum sativum L. | - | 1.08 |
| Poa annua L.* | <0.01 | - |
| Polygonum aviculare L. | 0.01 | 0.14 |
| Polygonum convolvulus L. | 0.13 | 6.70 |
| Pseudoroegneria spicata (Pursh) * | 0.81 | - |
| Salsola kali L. | 2.72 | 36.15 |
| Salvia reflexa Hornem. | 0.10 | 0.01 |
| Setaria viridis (L.) P. Beauv. * | 0.13 | 0.05 |
| Silene latifolia Poir. | - | <0.01 |
| Sinapis arvensis L. | - | 2.23 |
| Sisymbrium altissimum L. | - | 0.04 |
| Solanum triflorum Nutt. | - | <0.01 |
| Taraxacum officinale F.H. Wigg. | 0.34 | 0.04 |
| Thlaspi arvense L. | - | 1.31 |
| Tragopogon dubius Scop. | <0.01 | - |
| Trifolium sp. | - | <0.01 |
| Triticum aestivum L. * | - | 3.50 |
| Unknown (dicot) sp. | <0.01 | 0.81 |
| Unknown (grass) sp. * | <0.01 | 0.03 |
| Vaccaria hispanica (Mill.) Rauschert | - | 0.63 |
| Vicia americana Muhl. ex Willd. | <0.01 | 0.43 |
| Farming Systems | Infested Stems | Parasitoids | Cut Stems | Total Stems Observed |
|---|---|---|---|---|
| Conventional | 684 | 37 | 359 | 1835 |
| Organic | 195 | 54 | 48 | 969 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adhikari, S.; Adhikari, A.; Weaver, D.K.; Bekkerman, A.; Menalled, F.D. Impacts of Agricultural Management Systems on Biodiversity and Ecosystem Services in Highly Simplified Dryland Landscapes. Sustainability 2019, 11, 3223. https://doi.org/10.3390/su11113223
Adhikari S, Adhikari A, Weaver DK, Bekkerman A, Menalled FD. Impacts of Agricultural Management Systems on Biodiversity and Ecosystem Services in Highly Simplified Dryland Landscapes. Sustainability. 2019; 11(11):3223. https://doi.org/10.3390/su11113223
Chicago/Turabian StyleAdhikari, Subodh, Arjun Adhikari, David K. Weaver, Anton Bekkerman, and Fabian D. Menalled. 2019. "Impacts of Agricultural Management Systems on Biodiversity and Ecosystem Services in Highly Simplified Dryland Landscapes" Sustainability 11, no. 11: 3223. https://doi.org/10.3390/su11113223
APA StyleAdhikari, S., Adhikari, A., Weaver, D. K., Bekkerman, A., & Menalled, F. D. (2019). Impacts of Agricultural Management Systems on Biodiversity and Ecosystem Services in Highly Simplified Dryland Landscapes. Sustainability, 11(11), 3223. https://doi.org/10.3390/su11113223