1. Introduction
Rice is a staple food for 2.7 billion people in Asia, where 90% of the world’s rice is grown [
1]. One of the top five rice crop diseases being addressed in the Philippines today is rice blast disease. The disease is caused by the ascomycete fungus
Magnaporthe oryzae (anamorph
Pyricularia oryzae) [
2], which poses a significant threat to food security, damaging as much as 30% of the global rice harvest, with yield losses of 11.9 kg ha
−1 in South and Southeast Asia. In the Philippines, rice yield losses range from 50% to 85% [
3].
Different approaches have been developed to increase productivity and overcome the challenges of rice blast disease. Developing resistant rice crop varieties has had only partial success because of the ability of rice blast fungus to evolve to new races and the dependence of the resistant cultivar on a favorable environment [
1,
4]. Therefore, chemical pesticide and fertilizer application is still widely used for control [
1,
5]. However, extensive use of fungicides may increase the emergence of resistant populations of the pathogen [
6]. The associated risk to human health and the environment is expected to increase rather than decrease in the near future unless peripheral interference provokes alterations toward more sustainable forms [
7]. As a consequence, different biocontrol microbial species have been developed [
6,
8,
9], introducing natural enemies to undergo a population-levelling process whereby the population of one species decreases the number of another species via predation, parasitism, and pathogenicity or competition [
1,
10,
11].
In the Philippines, the Philippine Rice Research Institute (PhilRice), via its ecological resource recovery system project, works on converting biodegradable wastes into fertilizers. The agency developed a low-cost microbial base inoculant to process biodegradable waste such as sawdust, coco dust, carbonized rice hull, and molasses inoculated with a mixture of beneficial microorganisms known as effective microorganisms (EMs) into fertilizers in only two weeks. The use of EMs enhances resistance of crops to stress, reduces soil-borne pathogens and diseases, improves yield and quality, and prolongs a crop’s shelf life [
12]. Among the above-mentioned biodegradable waste applied in inoculants, rice hulls, an agricultural by-product, previously used to control paddy weeds, was reported to contain the allelochemical tricin and its aurone isomer, which function to reduce the incidence of rice seedling rot disease [
13].
The aim of the study was to explore the ability of rice soil bacteria to inhibit the growth of the rice blast fungal pathogen M. oryzae and to develop a sustainable, non-polluting, and cost-effective biofungicide formulation for rice blast disease control. We isolated potential disease control bacteria to combat the rice blast disease pathogen in the development of a biofungicide formulation designed following the EM concept and tested the effect of rice hulls applied with the bacteria strains.
2. Materials and Methods
2.1. Isolation of Rice Blast Fungus (M. oryzae)
The pure culture of M. oryzae was isolated as follows: The infected leaves were collected from College of Agriculture Crop Protection Cluster, University of the Philippines Los Banos (UPLB) (14.167743, 121.243272). After collection, the leaves were washed with sterile distilled water before being dried with a paper towel. The infected area was sliced to 1 inch pieces. A total of 5 leaves pieces were picked and placed on an agar plate. The plates were incubated for 2 days at 25 °C. The isolate was picked using a sterile inoculating loop before being placed in a 1 mL sterile 1 M phosphate buffered saline (PBS) buffer. Then, the serial dilute was suspended to 10−2 to 10−4 and cultured on the plates. The plates were incuabted for another 2 days at 25 °C. Each isolate was cultured in triplicate on either potato dextrose agar (PDA) or rose bengal agar (RBA) plates. The purity of the culture was confirmed at least 4 times with the quadrant streaking technique.
2.2. Isolation of the Potential Biocontrol Bacteria from the Soil Samples
Soil samples were collected from the crop roots in various points of the rice field in UPLB. One gram of soil sample was placed in a 10 mL sterile 1M PBS buffer solution in 3 replicates. The potential biocontrol bacteria were incubated either on trypticase soy agar (TSA) or LB Agar (LBA) for 16–48 h at 25 ± 3 °C.
2.3. Inhibition Test
The antifungal activity of the isolated soil microorganisms was measured by the modified agar disc method [
14,
15]. The plates were incubated at 25 ± 3 °C for 3–5 days. Three replications were made for each strain.
The shortest and the longest diameter of the cleared zones were measured and averaged. The results are expressed in diameter (mm). Antifungal activity around the soil microorganism was evaluated by the following ratings [
14]: (1) no inhibition: Mycelial growth not different from control (−); (2) weak inhibition: Partial inhibition of mycelia growth, measured as 5 to 9 mm (+); (3) moderate inhibition: Almost complete inhibition of mycelial growth, measured as a diameter of 10 to 19 mm (++); and (4) strong inhibition: Complete inhibition, in which most mycelia did not grow, measured as a diameter of >20 mm (+++). A blank agar disc was used as the control.
2.4. Identification of Potential Biofungicide Bacteria
2.4.1. DNA Extraction
Pure culture bacteria were grown in a liquid LB medium at 37 °C for one day. A total of 1 mL of bacterial inoculant was added into a microcentrifuge tube. The tube was centrifuged for 1 min at 12,000 rpm and the supernatant was discarded. The procedure was conducted twice to increase the yield. The pellet was mixed with 1 mL of sodium chloride-tris-EDTA (STE) buffer. The mixture was centrifuged for 1 min at 12,000 rpm and the supernatant was discarded. The STE buffer (200 µL) was added with a sodium dodecyl sulfate (SDS) buffer (20 µL, 10%) into a microcentrifuge tube and then vortexed. The microcentrifuge tube was then placed in a water bath at 65 °C for 20–30 min. After the water bath, 4 µL of Protein ASEK (10 mg/mL Proteinase K) was added to the solution, and was again suspended in a water bath overnight at 56 °C. After the second water bath, a 400 µL STE buffer was added into the tube and mixed gently. The solution was washed using phenol and chloroform: 10 drops of phenol and 10 drops of chloroform were added to the solution. It was mixed vigorously and then centrifuged for 5 min at 12,000 rpm. The phenol and chloroform wash was repeated until a clear solution was obtained. An equal volume of chloroform was added to the clear solution and then centrifuged at 12,000 rpm for 5 min. The supernatant was transferred to a new microcentrifuge tube. Three molar NaOAc (sodium acetate) (pH 5.2, 1/10 volume of the supernatant) and 95% ethanol were added into the tube to increase the volume up to 1 mL before being vortexed. The solution was allowed to sit in the freezer for 10 min. The solution was then centrifuged for 15 min at 12,000 rpm and the alcohol was gently discarded. The microcentrifuge tube was centrifuged again and the rest of the alcohol was discarded. The tube was dried in an oven for 5 min at 60 °C to evaporate any remaining alcohol. Distilled water (20 µL) was added to the dry DNA pellets. The solution was then given a quick spin in the centrifuge and was allowed to sit for 5 min to ensure the DNA had dissolved in the water.
2.4.2. Polymerase Chain Reaction (PCR)
The 16S rDNA fragments were amplified using universal primers for bacteria: Reverse primer 1492r: 5′ TAC GGT TAC CTT GTT ACGACT T 3′ and forward primer 27f: 5′ GAG TTT GAT CAT GGC TCA G 3′ [
16]. PCR reagents were purchased from Ginomics BioScience and Technology and the 50-µL PCR solution was prepared as follows: 34.5 µL ddH
2O, 5 µL 10xPCR Buffer, 8 µL dNTP, 0.5 µL forward primer, 0.5 µL reverse primer, 1 µL Temp DNA (already diluted by the ratio of 1 µL DNA extraction product + 99 µL ddH
2O), and 0.5 µL enzyme (rTaq, DNA Polymerase). PCR (BIO-RAD T-100 Thermal Cycler) was conducted following the protocol: Temp 1: 94.0 °C/5.00 min; 40 cycles of Temp 2: 94.0 °C/30 s, 55.0 °C/30 s, 72.0 °C/30 s, and a final extension Temp 3: 72.0 °C/7 min, 4.0 °C/∞).
2.4.3. 16.S rDNA Gene Sequence Analysis
16S rDNA gene sequence analysis was conducted by Genomics (New Taipei City, Taiwan). The DNA sequence data for species were analyzed with a BLASTn algorithm (
https://blast.ncbi.nlm.nih.gov/Blast.cgi).
2.5. Biofungicide Formulation
2.5.1. Rice Hull–Rice Grain Biofungicide Formulation
The biofungicide bacteria were cultured in LB and incubated in the dark for 5 days at 25 ± 3 °C. The rice hull and rice grain mixture was prepared in a 500-mL conical flask (300 g rice hull, 100 g rice grain, and 200 mL distilled water) and sterilized in an autoclave for 30 min at 121 °C [
17]. An amount of 80 mL of LB medium inoculated with biocontrol agent bacteria was aseptically transferred to the mixture and incubated at 25 ± 3 °C for 2 weeks.
2.5.2. Liquid Biofungicide Formulation
An amount of 5 mL 1 M glucose solution was added to 120-mL sterilized LB and was inoculated with the biocontrol agent bacteria in the dark for 5 days at 25 ± 3 °C.
2.6. Crop Test
The biofungicide formulas were tested for their ability to suppress rice blast disease under a controlled environment. The rice cultivar used in this study was UPLRi-5 (upland rice), a rice blast resistant crop variety developed by UPLB. The method used was adapted and modified from previous studies [
1,
4,
17]. The rice seeds were soaked in distilled water for 24 h, then placed on a tray with a wet paper towel, and incubated for 24 h. The germinated seeds were submerged in the formula at 2% concentration for 30 min while the control seeds were soaked in distilled water. After treatments, seeds were sown on a tray with sterile paddy soil for 12 days before transplanting.
The crop test was performed in plastic pots (15 cm diameter) filled with the mixture of the sterile paddy soil (taken from <20 cm depth paddy field) and rice hull–rice grain biofungicide formulation (ratio 3:1). The liquid biofungicide formula was applied by spraying 5 mL of liquid biofungicide to the plants. Four treatments (in triplicates) were conducted as follows: (A) Bacillus subtilis 5 biofungicide mixture + B. subtilis 5 liquid biofungicide formula; (B) Bacillus cereus 3S5 biofungicide mixture + B. cereus 3S5 liquid biofungicide formula; and (C) Pseudomonas fluorecens 10S2 biofungicide mixture + P. fluorecens 10S2 liquid biofungicide formula. At 19 days after transplanting, 1 g of rice hull–rice grain biofungicide formula was placed between the stem and the basal leaf sheath of each tiller and in the roots, whereas 20 mL of liquid biofungicide formula was sprayed on the whole plant.
The infection of the pathogen was conducted 1 day after the application of biofungicide by spraying the plant with 20 mL of a pathogen spore suspension (2-week-old PDA culture of disease pathogen spores were harvested by using 7 mL of sterile distilled water containing 0.5% gelatin, filtered through 0.2-μm nylon mesh that was transferred immediately to a flask on ice to prevent spore germination). After inoculation, the plant was covered with black plastic for 12 h to stimulate infection.
Two days after the infection, the biofungicide treatments were conducted again as described previously (1 g of rice hull–rice grain biofungicide formula and 20 mL of liquid biofungicide formula).
2.7. Disease Assessment
At 30 days after pathogen infection, the disease was assessed from each leaf with the following formula developed by IRRI (1996):
DI: disease intensity
ni: number of leaves with i score
N: number of total leaves observed
V: the highest scale of disease severity
v: scales of disease severity (0–9)
Scale description:
0 = no lesions
1 = small brown, specks of pinhead size
2 = larger brown specks
3 = small, roundish to slightly elongated, necrotic gray spots about 1 to 2 mm in diameter
4 = typical blast lesions, elliptical, 1 to 2 cm long, usually confined to the area of the two main veins infecting <2% of the total leaf area
5 = typical blast lesions infecting <10% of the leaf area
6 = typical blast lesions infecting 10%–25% of the leaf area
7 = typical blast lesions infecting 26%–50% of the leaf area
8 = typical blast lesions infecting 51%–75% of the leaf area
9 = all leaves dead
3. Results
3.1. Selection of Bacterial Strains
A total of 300 rhizobacteria strains were isolated and cultured from the paddy soil before the inhibition test. Only 6% of the tested soil microorganisms had a positive result on the inhibition test.
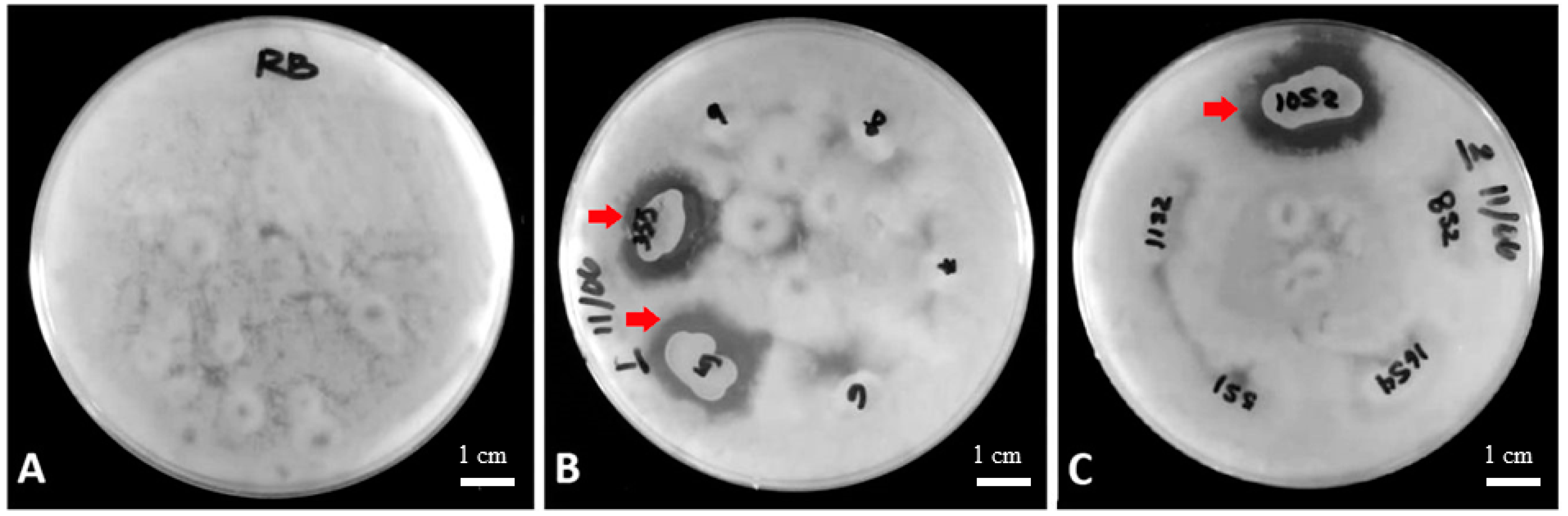
Table 1 lists the most effective bacterial strains among the inhibition tests. The strains were isolated from the different sampling sites of the rice rhizosphere, including those from healthy crops, rice blast-infected crops, and bacterial blight-infected crops. The inhibition zones of the potential biofungicides ranged from 14.5 to 24.3 mm in diameter.
Figure 1B,C show the inhibition zones from the most effective bacterial strains ID 5, 3S5, and 10S2.
3.2. Identification of Potential Biofungicides Strains
To identify the possible species of potential biofungicides bacteria, we analyzed the 16S rDNA sequences for bacterial strains ID 5, 3S5, and 10S2, then performed a BLAST search against the NCBI database (
Table 2). From the results, we obtained the possible species of the potential biofungicides:
Bacillus subtilis,
B. cereus, and
Pseudomonas fluorescens (ID 5, 3S5, and 10S2, respectively).
3.3. Crop Test of Biofungicide Formulations
The biofungicide formulas were tested on the rice cultivar UPLRi-5 under controlled conditions.
Figure 2 shows the severity of rice blast symptoms by different treatments in the pot test. Rice blast disease was effectively inhibited by the biofungicide formulas A, B, and C.
Rice blast disease control was 52% with the control rice crop but 80% to 85% with treatments, showing 31% to 33% higher resistance to the pathogen.
The disease intensity of the control rice crop was 42.9%, considerably higher than with treatments, which ranged from 14.9% to 19.3% (
Table 3). The highest disease intensity was with treatment C (
P. fluorescens).
4. Discussion
In sustainable agriculture, a biocontrol solution is needed for rice blast disease in order to make significant progress in environmental concerns and rice cultivation. In this study, we developed novel biofungicide formulas by using indigenous antagonistic bacteria to help improve rice blast disease control. The bacteria sequence can be manipulated by genetic modification to adapt the various paddy environments but requires further assessment and comprehensive understanding of pathosystems before modification. Yet one biological control can be an effective measure via competition in various living habitats, and the application of indigenous antagonist bacteria without modification is acceptable.
The antifungal activity of the microorganisms we found (
B. subtilis 5,
B. cereus 3S5, and
P. fluorecens 10S2) demonstrated their ability to inhibit the growth of the rice pathogen (
M. oryzae) and were strong candidates for biocontrol agents for rice blast disease compared with previous studies, which were 19.27% and 53.32% inhibition rates [
18]. In contrast to chemical fungicides, biocontrol bacteria produce a mixture of antifungal compounds that can fluctuate in content based on environmental cues [
8]. The application of biofungicide strains could lead to a better environmentally-friendly strategy for reducing rice blast disease than applying chemical fungicides because of the lower possibility of introducing fungicide-resistant pathogen strains. Furthermore, as living organisms, these biofungicide microbes may continue to evolve with their rhizospheric neighbor plants. For example, if the host plant modulates the root exudate composition to attract more plant-resistance triggering microbes, the pathogen inhibition activity in the soil also improves [
8]. Therefore, the direct antifungal activity of the biofungicide strains found in this study could have ecologically relevant implications in preventing blast infection.
Antibiotic compounds produced by antagonist bacteria contribute to the suppression of plant pathogens [
19]. Previous studies have reported that enzymes increase in activity after the use of biocontrol agents such as peroxidase [
20], an enzyme that acts as a catalyst for the final stage of lignin biosynthesis [
21]. It is a pathogenesis-related protein (PR-protein) that is toxic to the pathogen and increases the resistance of the cell wall against degrading enzymes produced by the pathogen [
19,
22]. Peroxidase is found in healthy plant tissue with its activity markedly increased when the plant is infected with the pathogen [
17].
B. subtilis (reported in this study),
B. polymyxa, and
P. fluorescens (reported in this study) were reported to increase plant health by increasing peroxidase activity in the plant [
4,
23].
The precise mechanism by which rice rhizospheric microbes induce physiological effects on the host (rice crop) is not known, but the microorganisms we identified were able to trigger a defense mechanism and reduce disease symptoms during pathogen infection. Plant roots encounter diverse microbial populations in soil and generate a unique ecological niche for microbes by secreting resources into the rhizosphere. These rhizospheric resources are limited and some microbes have evolved antimicrobial traits to reduce competition from other microbes and bolster the health of their plant host [
8]. However, we still require a comprehensive understanding of the contribution of individual microbial strains to plant growth and protection. Since biofungicide has been recognized as a possible approach to crop protection, further studies are needed to classify probable biofungicide agents from the diverse pool of soil microorganisms and to understand the mechanisms by which they affect plant health, pathogen resistance, and productivity.
5. Conclusions
In this study, we isolated three rhizobacteria B. subtilis 5, B. cereus 3S5, and P. fluorecens 10S2 which can significantly inhibit the growth of rice blast fungus in plate tests. We developed three novel biofungicide formulations inoculated with these bacteria species, exhibiting a significantly rice blast disease intensity inhibition with the mean disease control rate being approximately 31% higher than in the control, making them promising novel treatments for rice blast disease.
Author Contributions
Conceptualization, T.-Y.C.; Data curation, W.-C.C. and A.L.D.; Formal analysis, W.-C.C. and A.L.D.; Funding acquisition, C.-S.L.; Investigation, A.L.D.; Methodology, W.-C.C., T.-Y.C., A.L.D. and C.-S.L.; Project administration, T.-Y.C. and C.-S.L.; Resources, C.-S.L.; Supervision, W.-C.C. and C.-S.L.; Writing – original draft, W.-C.C., A.L.D. and C.-S.L.; Writing – review & editing, W.-C.C., T.-Y.C. and C.-S.L.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gnanamanickam, S.S.; Mew, T.W. Biological control of blast disease of rice (Oryza sativa L.) with antagonistic bacteria and its mediation by a Pseudomonas antibiotic. Ann. Phytopathol. Soc. Jpn. 1992, 58, 380–385. [Google Scholar] [CrossRef]
- Kuyek, D. Blast, Biotech, and Big Business: Implications of Corporate Strategies in Rice Research in Asia. Agricultural R. & D. 2000. Available online: https://www.grain.org/article/entries/36-blast-biotech-and-big-business-implications-of-corporate-strategies-on-rice-research-in-asia (accessed on 1 March 2019).
- Leung, H. Disease- and Pest-Resistant Rice. 2018. Available online: http://irri.org/our-work/research/better-rice-varieties/disease-and-pest-resistant-rice (accessed on 1 March 2019).
- Suprapta, D.N.; Quintao, V.; Khalimi, K. Effectiveness of rhizobacteria to reduce rice blast disease intensity. J. Biol. Agric. Healthc. 2014, 4, 35–41. [Google Scholar]
- Singh, A.; Parmar, N.; Kuhad, R.C.; Ward, O.P. Bioaugmentation, biostimulation, and biocontrol in soil biology. In Bioaugmentation, Biostimulation and Biocontrol; Singh, A., Parmar, N., Kuhad, R.C., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 1–23. [Google Scholar]
- Khalil, M.S.; Moubasher, H.; Hasan, F.F. Biological control of rice blast disease by Streptomyces flavotricini. Res. J. Pharm. Biol. Chem. Sci. 2014, 5, 1453–1461. [Google Scholar]
- Snelder, D.J.; Masipiqueña, M.D.; de Snoo, G.R. Risk assessment of pesticide usage by smallholder farmers in the Cagayan Valley (Philippines). Crop Prot. 2008, 27, 747–762. [Google Scholar] [CrossRef]
- Spence, C.; Alff, E.; Johnson, C.; Ramos, C.; Donofrio, N.; Sundaresan, V.; Bais, H. Natural rice rhizospheric microbes suppress rice blast infections. BMC Plant Biol. 2014, 14, 130. [Google Scholar] [CrossRef] [PubMed]
- Law, J.W.; Ser, H.L.; Khan, T.M.; Chuah, L.H.; Pusparajah, P.; Chan, K.G.; Goh, B.H.; Lee, L.H. The potential of Streptomyces as biocontrol agents against the rice blast fungus, Magnaporthe oryzae (Pyricularia oryzae). Front Microbiol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Resource Manual on Integrated Production and Pest Management (IPPM) in Rice; World Education (INGO) Philippines, Inc.: Laguna, Philippines, 2005.
- Kalia, A.; Mudhar, R.K. Biological control of pests. In Bioaugmentation, Biostimulation and Biocontrol; Singh, A., Parmar, N., Kuhad, R.C., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 223–240. [Google Scholar]
- Corales, R.G.; Tadeo, B.D.; Cordero, J.D. Rice-based Microbial Inoculant. In Rice Technology Bulletin; Department of Agriculture, Philippine Rice Research Institute (PhilRice): Science City of Muñoz, Philippines, 2003; Volume 81, No. 48; ISSN 0117-9799. [Google Scholar]
- Kong, C.H.; Xu, X.H.; Zhang, M.; Zhang, S.Z. Allelochemical tricin in rice hull and its aurone isomer against rice seedling rot disease. Pest. Manag. Sci. 2010, 66, 1018–1024. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimi Zarandi, M.; Shahidi Bonjar, G.H.; Padasht Dehkaei, F.; Ayatollahi Moosavi, S.A.; Rashid Farokhi, P.; Aghighi, S. Biological control of rice blast (Magnaporthe oryzae) by use of Streptomyces sindeneusis isolate 263 in greenhouse. Am. J. Appl. Sci. 2009, 6, 194–199. [Google Scholar] [CrossRef]
- Naureen, Z.; Price, A.H.; Hafeez, F.Y.; Roberts, M.R. Identification of rice blast disease-suppressing bacterial strains from the rhizosphere of rice grown in Pakistan. Crop Prot. 2009, 28, 1052–1060. [Google Scholar] [CrossRef] [Green Version]
- Miller, C.S.; Handley, K.M.; Wrighton, K.C.; Frischkorn, K.R.; Thomas, B.C.; Banfield, J.F. Short-read assembly of full-length 16S amplicons reveals bacterial diversity in subsurface sediments. PLoS ONE 2013, 8, e56018. [Google Scholar] [CrossRef] [PubMed]
- Nagarajkumara, M.; Jayarajb, J.; Muthukrishnanb, S.; Bhaskarana, R.; Velazhahan, R. Detoxification of oxalic acid by Pseudomonas fluorescens strain PfMDU2: Implications for the biological control of rice sheath blight caused by Rhizoctonia solani. Microbiol. Res. 2005, 160, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Suryadi, Y.; Susilowati, D.; Riana, E.; Mubarik, N. Management of rice blast disease (Pyricularia oryzae) using formulated bacterial consortium. Emir. J. Food Agric 2013, 25, 349–357. [Google Scholar] [CrossRef]
- Whipps, J.M. Microbial interactions and biocontrol in the rhizosphere. J. Expt. Bot. 2001, 52 (Suppl. 1), 487–511. [Google Scholar] [CrossRef]
- Chen, C.; Belanger, R.R.; Benhamou, N.; Paulitz, T.C. Defense enzymes induced in cucumber roots by treatment with plant growth-promoting rhizobacteria (PGPR) and Pythium aphanidermantum. Physiol. Mol. Plant. Pathol. 2000, 56, 13–23. [Google Scholar] [CrossRef]
- Vance, C.P.; Kirk, T.K.; Sherwod, R.T. Lignification as a mechanism of disease resistance. Annu. Rev. Phytopathol. 1980, 18, 259–288. [Google Scholar] [CrossRef]
- Huang, J.S. Degradation of Cell Walls by Plant Pathogens. In Plant Pathogenesis and Resistance; Kluwer Academic Publishers: Dordrecht, The Netherlands; Boston, MA, USA; London, UK, 2001; pp. 51–130. [Google Scholar]
- Sha, Y.; Wang, Q.; Li, Y. Suppression of Magnaporthe oryzae and interaction between Bacillus subtilis and rice plants in the control of rice blast. Springerplus 2016, 5, 1238. [Google Scholar] [CrossRef] [PubMed]
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).







{kind=link}
{kind=link}