Survivability of Probiotic Bacteria in Model Systems of Non-Fermented and Fermented Coconut and Hemp Milks

 ,
, 
 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
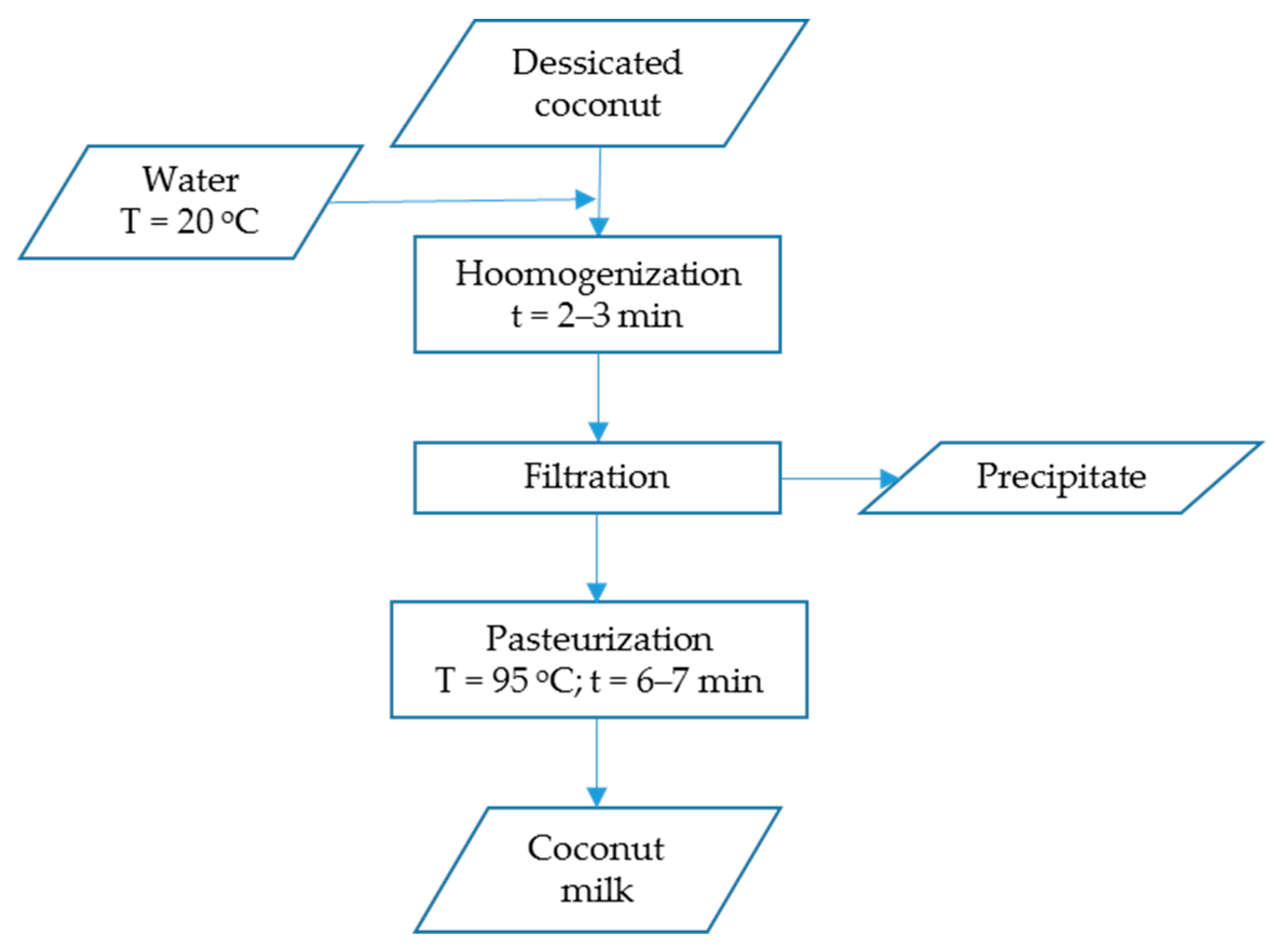
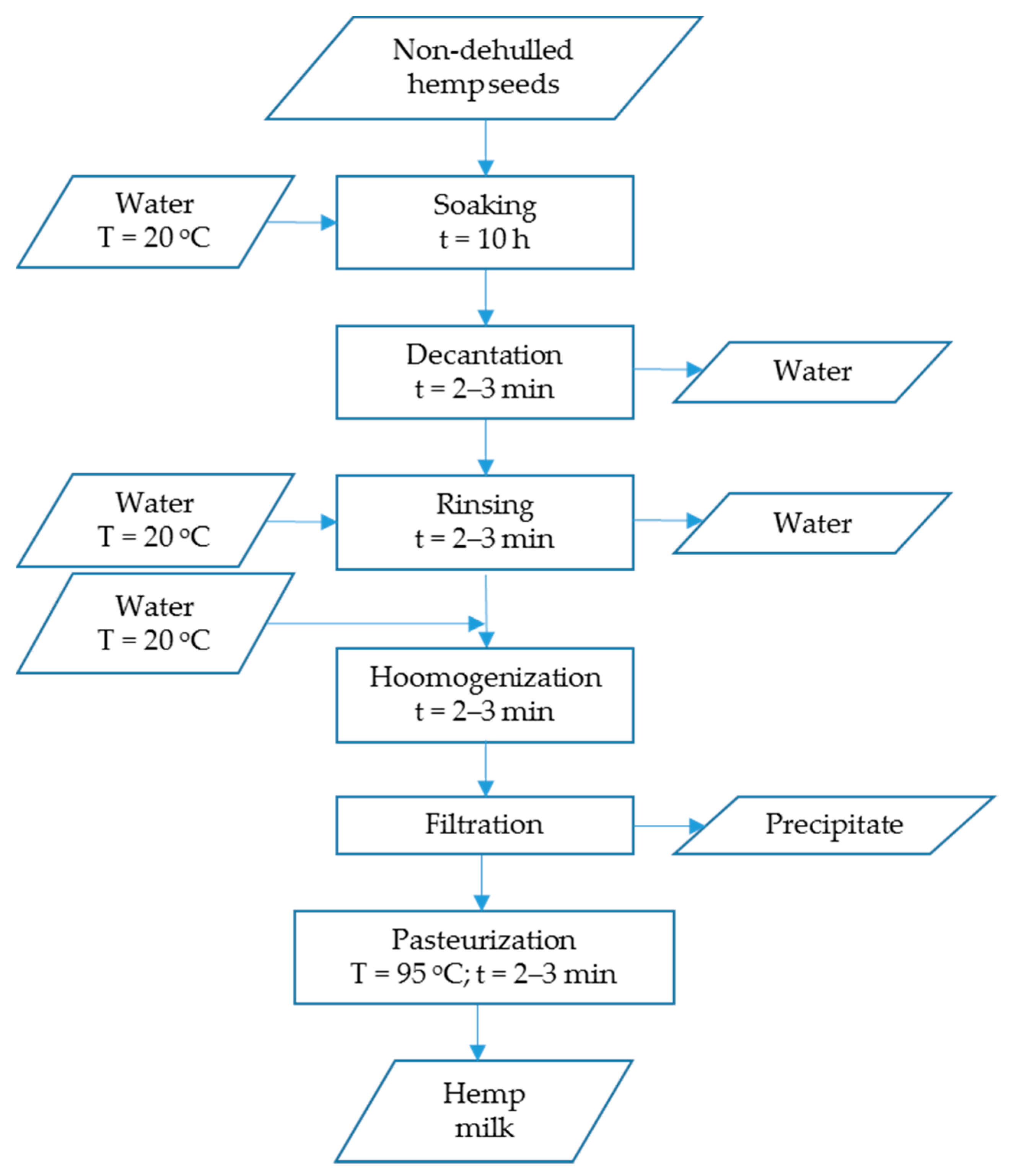
2.1. Plant Material and Beverages Production
2.2. Probiotic Microorganisms
2.3. Chemical Evaluation of Non-Dairy Beverages
2.4. Microbiological Status of Plant Beverages
2.5. Microbiological Analyses of Counts of Viable Bacterial Cells During Storage of Fermented and Non-Fermented Non-Dairy Beverages
2.6. Statistical Analysis
3. Results and Discussion
3.1. Evaluation of the Produced Non-Dairy Beverages
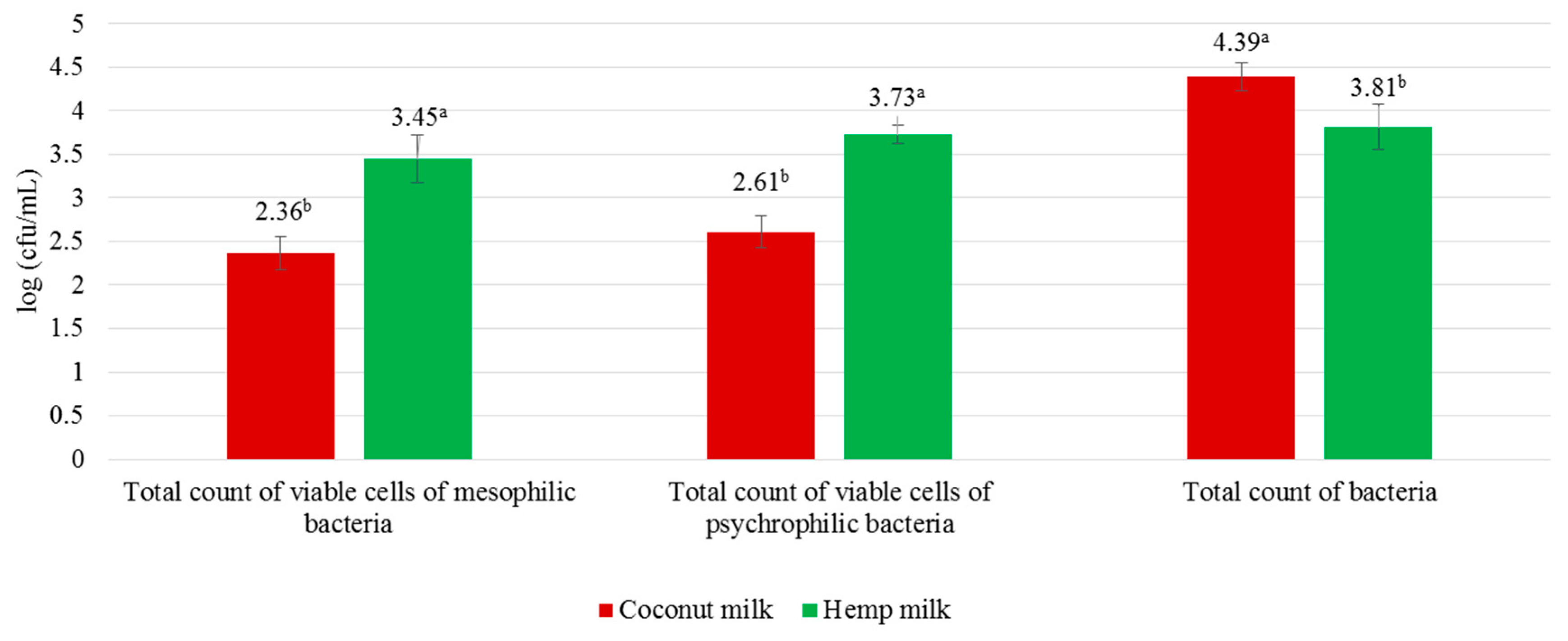
3.2. Microbiological Purity of Non-Dairy Beverages
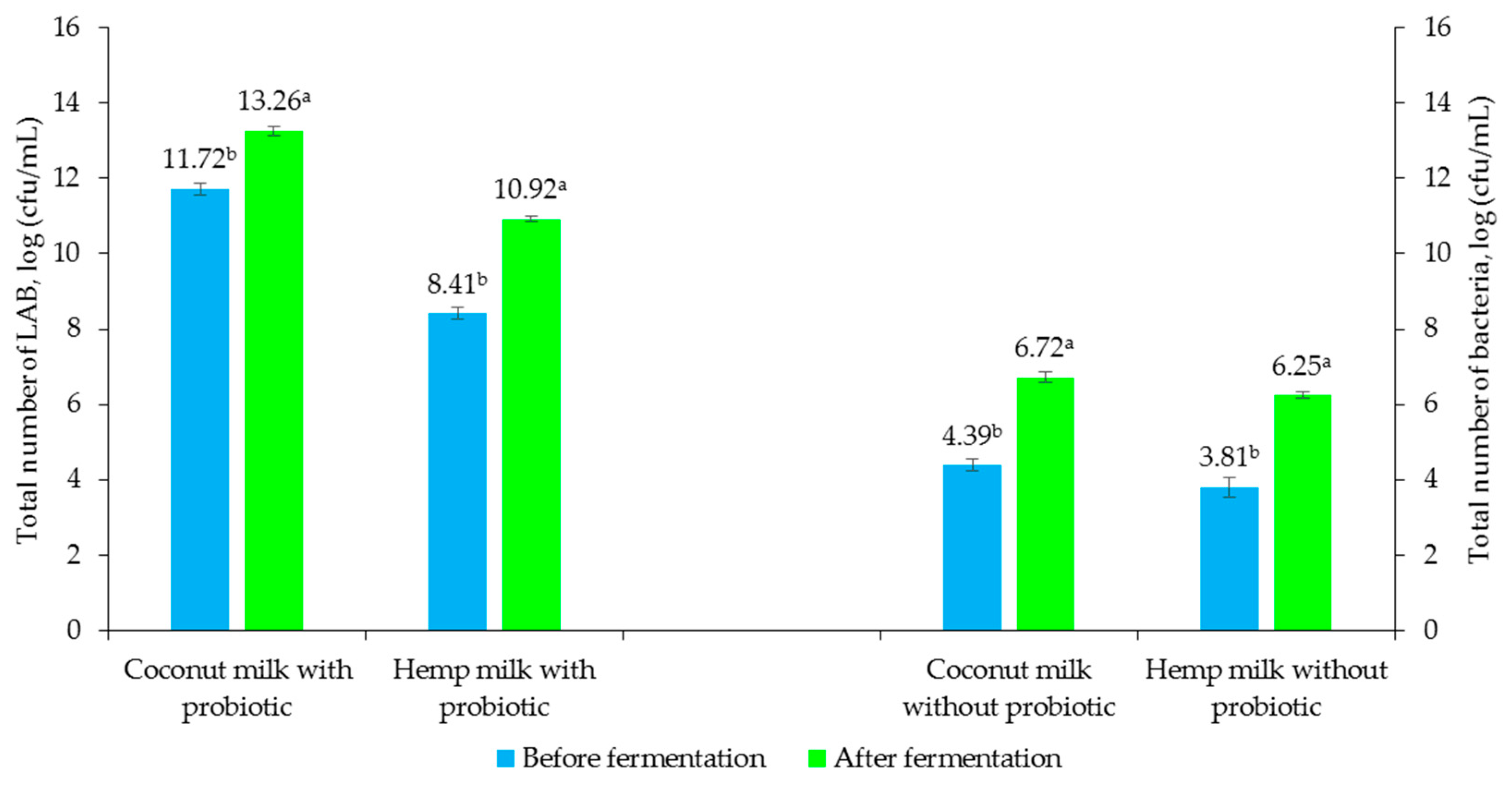
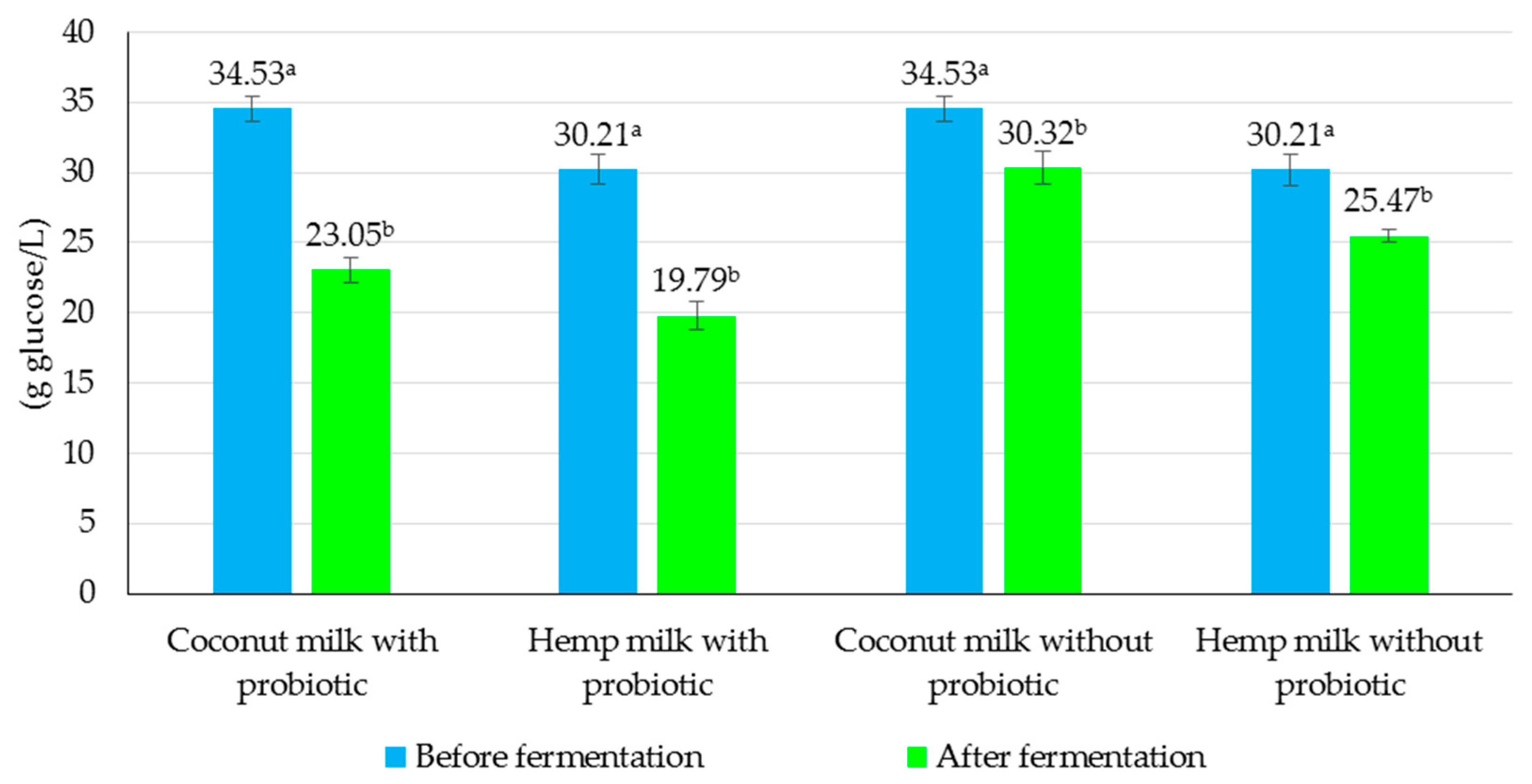
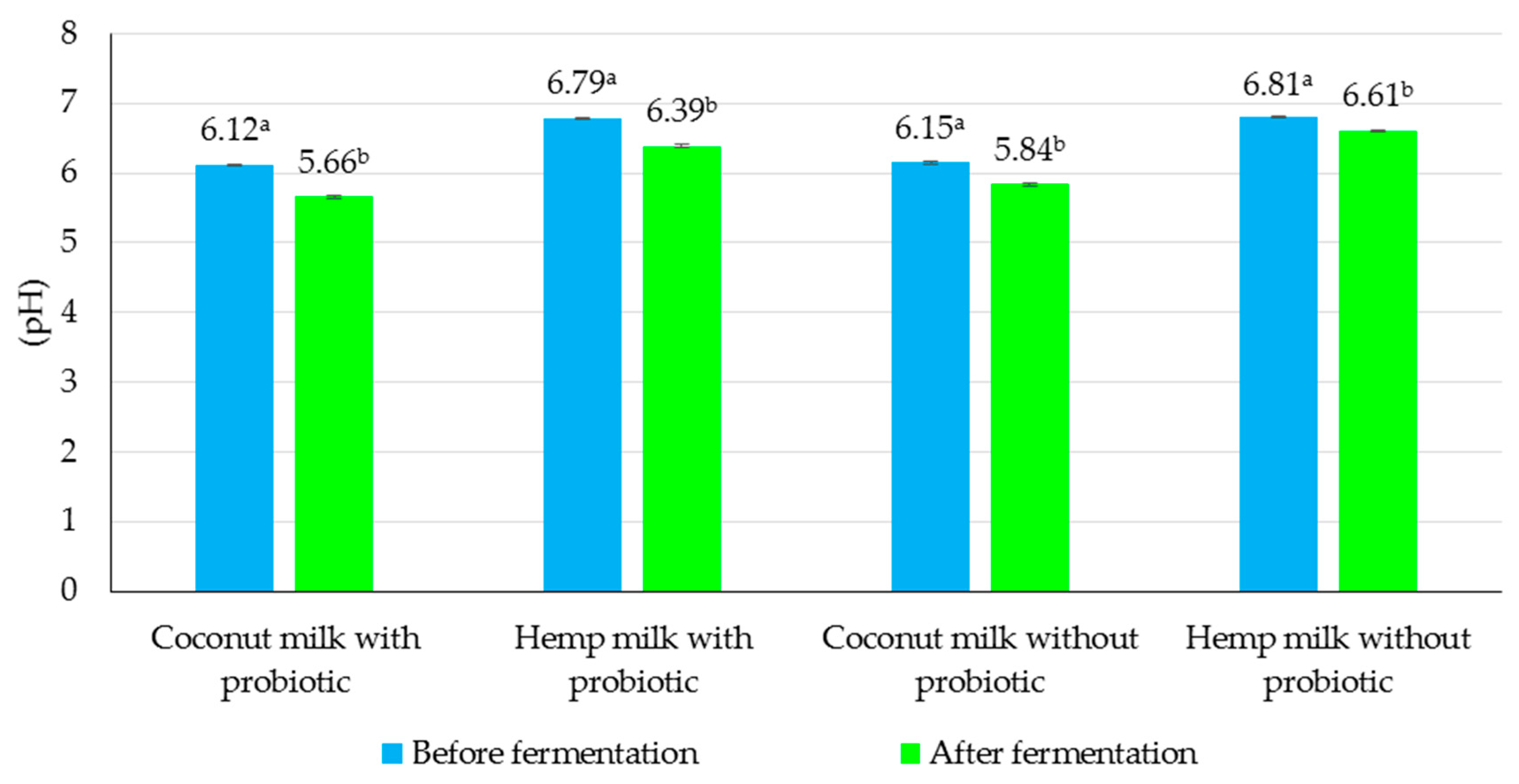
3.3. Evaluation of the Effect of Fermentation Process on Chemical and Microbiological Properties of Non-Dairy Beverages
3.4. Assessment of the Quality of the Produced Non-Dairy Beverages During Cold Storage
3.5. Quantitative Analysis of Viable Bacterial Cells During Storage of Fermented and Non-Fermented Non-Dairy Beverages
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Dam, L.R.; Boafo, Y.A.; Degefa, S.; Gasparatos, A.; Saito, O. Assessing the food security outcomes of industrial crop expansion in smallholder settings: Insights from cotton production in Northern Ghana and sugarcane production in Central Ethiopia. Sustain. Sci. 2017, 677–693. [Google Scholar] [CrossRef]
- Szparaga, A.; Kocira, S. Generalized logistic functions in modelling emergence of Brassica napus L. PLoS ONE 2018, 13, e0201980. [Google Scholar] [CrossRef] [PubMed]
- Alexandratos, N.; Bruinsma, J. World Agriculture: Towards 2015/2030: The 2012 Revision. Available online: http://www.fao.org/3/a-ap106e.pdf (accessed on 24 July 2019).
- FAO. Food security information for action programme. In Food Security Concepts and Frameworks; FAO: Rome, Italy, 2008. [Google Scholar]
- Blay-Palmer, A.; Sonnino, R.; Custot, J. A food politics of the possible? Growing sustainable food systems through networks of knowledge. Agric. Hum. Values 2016, 33, 27–43. [Google Scholar] [CrossRef]
- Szparaga, A.; Stachnik, M.; Czerwińska, E.; Kocira, S.; Dymkowska-Malesa, M.; Jakubowski, M. Multi-objective optimization of osmotic dehydration of plums in sucrose solution and its storage using utopian solution methodology. J. Food Eng. 2019, 245, 104–111. [Google Scholar] [CrossRef]
- Saluk-Juszczak, J.; Kołodziejczyk, J.; Babicz, K. Functional food—A role of nutraceuticals in cardiovascular disease prevention. Kosmos 2010, 59, 527–538. [Google Scholar]
- Martirosyan, D.; Singh, J. A new definition of functional food by FFC: What makes a new definition unique? Funct. Food Health Dis. J. 2015, 5, 209–223. [Google Scholar] [CrossRef]
- Dymkowska-Malesa, M.; Szparaga, A.; Czerwińska, E. Evaluation of polychlorinated biphenyls content in chosen vegetablesfrom Warmia and Mazury region. Rocz. Ochr. Srodowiska 2014, 16, 290–299. [Google Scholar]
- Mullin, G.; Delzenne, N.M. Functional foods and dietary supplements in 2017: Food for thought. Curr. Opin. Clin. Nutr. Metab. Care 2017, 20, 453–455. [Google Scholar] [CrossRef]
- Douglas, L.C.; Sanders, M.E. Probiotics and prebiotics in dietetics practice. J. Am. Diet. Assoc. 2008, 108, 510–521. [Google Scholar] [CrossRef]
- Kapka-Skrzypczak, L.; Niedźwiecka, J.; Wojtyła, A.; Kruszewski, M. Probiotics and prebiotics as a bioactive component of functional food. Pediatr. Endocrinol. Diabetes Metab. 2012, 18, 79–83. [Google Scholar]
- Sethi, S.; Tyagi, S.K.; Anurag, R.K. Plant-based milk alternatives an emerging segment of functional beverages: A review. J. Food Sci. Technol. 2016, 53, 3408–3423. [Google Scholar] [CrossRef] [PubMed]
- Kandylis, P.; Pissaridi, K.; Bekatorou, A.; Kanellaki, M.; Koutinas, A. Dairy and non-dairy probiotic beverages. Curr. Opin. Food Sci. 2016, 7, 58–63. [Google Scholar] [CrossRef]
- Hoffman, M.; Kostyra, E. Sensory quality and nutritional value of vegan substitutes of milk. PTPS 2015, 1, 52–57. [Google Scholar]
- Zielińska, D. Lactobacillus Strain Survival Study in Fermeted Soy Beverage. Zywnosc-Nauka Technol. Jakosc 2006, 49, 121–128. [Google Scholar]
- Quasem, J.M.; Mazahreh, A.S.; Abu-Alruz, K. Development of Vegetable Based Milk from Decorticated Sesame (Sesamum Indicum). Am. J. Appl. Sci. 2009, 6, 888–896. [Google Scholar] [CrossRef] [Green Version]
- Yeung, P.S.M.; Sanders, M.E.; Kitts, C.L.; Cano, R.; Tong, P.S. Species specific identification on commercial probiotic strains. J. Dairy Sci. 2002, 8, 1039–1051. [Google Scholar] [CrossRef]
- Hadadji, M.; Bensoltane, A. Growth and lactic acid production by Bifidobacterium longum and Lactobacillus acidophilus in goat’s milk. Afr. J. Biotechnol. 2006, 5, 505–509. [Google Scholar]
- Yuliana, N.; Rangga, A. Rakhmiati Manufacture of fermented coco milk-drink containing lactic acid bacteria cultures. Afr. J. Food Sci. 2010, 4, 558–562. [Google Scholar]
- Bartkiene, E.; Zokaityte, E.; Lele, V.; Sakiene, V.; Zavistanaviciute, P.; Klupsaite, D.; Bendoraitiene, J.; Navikaite-Snipaitiene, V.; Ruzauskas, M. Technology and characterisation of whole hemp seed beverages prepared from ultrasonicated and fermented whole seed paste. Int. J. Food Sci. Technol. 2019, 1–14. [Google Scholar] [CrossRef]
- Wang, Q.; Jiang, J.; Xiong, Y.L. High pressure homogenization combined with pH shift treatment: A process to produce physically and oxidatively stable hemp milk. Food Res. Int. 2018, 106, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Scholz-Ahrens, K.E.; Ahrens, F.; Barth, C.A. Nutritional and health attributes of milk and milk imitations. Eur. J. Nutr. 2019, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Kaniewski, R.; Pniewska, I.; Kubacki, A.; Strzelczyk, M.; Chudy, M.; Oleszak, G. Konopie siewne (Cannabis sativa L.)-cenna roślina przydatna i lecznicza. Postępy Fitoter. 2017, 2, 139–144. [Google Scholar] [CrossRef]
- Mojka, K. Probiotics, prebiotics and synbiotics–characteristics and functions. Probl. Hig. Epidemiol. 2014, 95, 541–549. [Google Scholar]
- Zaręba, D.; Ziarno, M.; Obiedziński, M. Viability of yoghurt bacteria and probiotic strains in models of fermented and non-fermented milk. Med. Weter. 2008, 64, 1007–1011. [Google Scholar]
- Jeske, S.; Zannini, E.; Arendt, E.K. Past, present and future: The strength of plant-based dairy substitutes based on gluten-free raw materials. Food Res. Int. 2018, 110, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Akin, Z.; Ozcan, T. Functional properties of fermented milk produced with plant proteins. LWT 2017, 86, 25–30. [Google Scholar] [CrossRef]
- Mårtensson, O.; Öste, R.; Holst, O. Lactic Acid Bacteria in an Oat-based Non-dairy Milk Substitute: Fermentation Characteristics and Exopolysaccharide Formation. LWT-Food Sci. Technol. 2000, 33, 525–530. [Google Scholar] [CrossRef]
- Cruz, N.; Capellas, M.; Jaramillo, D.; Trujillo, A.-J.; Guamis, B.; Ferragut, V. Soymilk treated by ultra high-pressure homogenization: Acid coagulation properties and characteristics of a soy-yogurt product. Food Hydrocoll. 2009, 23, 490–496. [Google Scholar] [CrossRef]
- Chaiwanon, P.; Puwastien, P.; Nitithamyong, A.; Sirichakwal, P.P. Calcium Fortification in Soybean Milk and In Vitro Bioavailability. J. Food Compos. Anal. 2000, 13, 319–327. [Google Scholar] [CrossRef]
- Patel, H.; Pandiella, S.; Wang, R.; Webb, C. Influence of malt, wheat, and barley extracts on the bile tolerance of selected strains of lactobacilli. Food Microbiol. 2004, 21, 83–89. [Google Scholar] [CrossRef]
- Vera-Pingitore, E.; Jimenez, M.E.; DallAgnol, A.; Belfiore, C.; Fontana, C.; Fontana, P.; Von Wright, A.; Vignolo, G.; Plumed-Ferrer, C. Screening and characterization of potential probiotic and starter bacteria for plant fermentations. LWT 2016, 71, 288–294. [Google Scholar] [CrossRef]
- Navarro, R.R.; Faronilo, K.M.L.; Eom, S.H.; Jeon, K.H. Isolation, characterization, and identification of probiotic lactic acid bacterium from sabeng, a Philippine fermented drink. J. ISSAAS 2018, 24, 127–136. [Google Scholar]
- Blasco, M. Milk Power. Mleko roślinne 80 przepisów; Wydawnictwo Burda: Warszawa, Poland, 2014. [Google Scholar]
- Szakuła, M. Konopie w kuchni; Smiling Spoon: Warszawa, Poland, 2015. [Google Scholar]
- Zielińska, D.; Kołożyn-Krajewska, D.; Goryl, A. Survival models of potentially probiotic Lactobacillus casei KN291 bacteria in a fermented soy beverage. Zywnosc-Nauka Technol. Jakosc 2008, 5, 126–134. [Google Scholar]
- Zielińska, D.; Uzarowicz, U. Development of ripening and storage conditions of probiotic soy beverage. Zywnosc-Nauka Technol. Jakosc 2007, 5, 186–193. [Google Scholar]
- Zielińska, D. Selecting Suitable Bacterial Strains of Lactobacillus and Identifying Soya Drink Fermentation Conditions. Zywnosc-Nauka Technol. Jakosc 2005, 2, 289–297. [Google Scholar]
- AOAC Official Method. 18 Fat Content of Raw and Pasteurized Whole Milk Gerber Method by Weight (Method I) First Action; AOAC International: Rockville, MD, USA, 2000. [Google Scholar]
- AOAC. Official Methods of Analysis of AOAC International, 17th ed.; Horwitz, W., Ed.; AOAC International: Gaithersburg, MD, USA, 2000. [Google Scholar]
- AOAC. Official Method of Analysis of the Association of Official Analytical Chemists; AOAC International: Arlington, VA, USA, 1997. [Google Scholar]
- Miller, G.L. Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Czerwińska, E.; Piotrowski, W. Potential Sources of Milk Contamination Influencing its Quality for Consumption. Rocz. Ochr. Srodowiska 2011, 13, 635–652. [Google Scholar]
- Gustaw, W.; Kozioł, J.; Waśko, A.; Skrzypczak, K.; Michalak-Majewska, M.; Nastaj, M. Physicochemical Properties and Survival of Lactobacillus casei in Fermented Milk Beverages Produced With Addition of Selected Milk Protein Preparations. Zywnosc-Nauka Technol. Jakosc 2015, 6, 129–139. [Google Scholar] [CrossRef]
- De Man, J.C.; Rogosa, M.; Sharpe, M.E. A MEDIUM FOR THE CULTIVATION OF LACTOBACILLI. J. Appl. Bacteriol. 1960, 23, 130–135. [Google Scholar] [CrossRef]
- Champagnem, C.P.; Rastall, R.A. Some technological challenges in the addition of probiotic bacteria to foods. In Prebiotics and Probiotics Science and Technology, 2nd ed.; Charalampopoulos, D., Rastall, R.A., Eds.; Springer: London, UK, 2009; pp. 763–806. [Google Scholar]
- Yamaguishi, C.T.; Spier, M.R.; Lindner, J.D.D.; Soccol, V.T.; Soccol, C.R. Current Market Trends and Future Directions. In Probiotics; Springer: Berlin, Germany, 2011; Volume 21, pp. 299–319. [Google Scholar] [CrossRef]
- Szydłowska, A.; Kołożyn-Krajewska, D. Applying potencially probiotic bacterial strains to pumpkin pulp fermentation. Zywnosc-Nauka Technol. Jakosc 2010, 73, 109–119. [Google Scholar] [CrossRef]
- Sobczyk, M.; Ziarno, M. Characteristics of Microbial Quality and Nutrient Content (Protein, Fat, Mineral Compounds) Some Petals Muesli. ZPPNR 2013, 575, 119–129. [Google Scholar]
- Jessa, J.; Hozyasz, K.K. Health value of coconut products. Pediatr. Pol. 2015, 90, 415–423. [Google Scholar] [CrossRef]
- Aizpurua-Olaizola, O.; Omar, J.; Navarro, P.; Olivares, M.; Etxebarria, N.; Usobiaga, A. Identification and quantification of cannabinoids in Cannabis sativa L. plants by high performance liquid chromatography-mass spectrometry. Anal. Bioanal. Chem. 2014, 406, 7549–7560. [Google Scholar] [CrossRef] [PubMed]
- Belewu, M.A.; Belewu, K.Y. Comparative Physico-Chemical Evulation of Tiger-nut, Soybean and Coconut Milk Sources. Int. J. Agric. Biol. 2007, 9, 785–787. [Google Scholar]
- Yeo, S.-K.; Liong, M.-T.; Yeo, S.; Liong, M. Effect of prebiotics on viability and growth characteristics of probiotics in soymilk. J. Sci. Food Agric. 2010, 90, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Jurkowski, M.; Błaszczyk, M. Physiology and Biochemistry of Lactic Acid Bacteria. Kosmos 2012, 61, 493–504. [Google Scholar]
- Nogala-Kałucka, M. Analiza żywności. Wybrane metody oznaczeń jakościowych i ilościowych składników żywności. Wyd; Uniwersytetu Przyrodniczego w Poznaniu: Poznań, Poland, 2016. [Google Scholar]
- Dłużewska, E.; Nizler, M.; Maszewska, M. Technological Aspects of Obtaining Soy Beverages. Acta Sci. Technol. Aliment 2004, 3, 93–102. [Google Scholar]
- Rozporządzenie Komisji (WE) 2073/2005 i 2019/229. Available online: https://eur-lex.europa.eu/legal-content/PL/TXT/PDF/?uri=CELEX:32019R0229&from=ES (accessed on 26 August 2019).
- Lee, S.-Y.; Dougherty, R.H.; Kang, D.-H. Inhibitory Effects of High Pressure and Heat on Alicyclobacillus acidoterrestris Spores in Apple Juice. Appl. Environ. Microbiol. 2002, 68, 4158–4161. [Google Scholar] [CrossRef] [Green Version]
- Ouwehand, A.C.; Salminen, S.J. The Health Effects of Cultured Milk Products with Viable and Non-viable Bacteria. Int. Dairy J. 1998, 8, 749–758. [Google Scholar] [CrossRef]
- Shah, N.P. Functional foods from probiotics and prebiotics. Food Technol. 2001, 55, 46–53. [Google Scholar]
- Gupta, S.; Abu-Ghannam, N. Probiotic Fermentation of Plant Based Products: Possibilities and Opportunities. Crit. Rev. Food Sci. Nutr. 2012, 52, 183–199. [Google Scholar] [CrossRef] [PubMed]
- Olszewska, M.; Łaniewska-Trokenheim, Ł. Study of the nonculturable state of lactic acid bacteria cells under adverse growth conditions. Med. Weter. 2011, 67, 105–109. [Google Scholar]
- Warmińska-Radyko, I.; Olszewska, M.; Mikś-Krajnik, M. Effect of temperature and sodium chloride on the growth and metabolism of Lactococcus strains in long-term incubation of milk. Milchwissenschaft 2010, 65, 32–35. [Google Scholar]
- Joux, F.; LeBaron, P. Use of fluorescent probes to assess physiological functions of bacteria at single-cell level. Microbes Infect. 2000, 2, 1523–1535. [Google Scholar] [CrossRef]
- Lahtinen, S.J.; Gueimonde, M.; Ouwehand, A.C.; Reinikainen, J.P.; Salminen, S.J. Probiotic Bacteria May Become Dormant during Storage. Appl. Environ. Microbiol. 2005, 71, 1662–1663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaręba, D. Fatty Acid Profile of Soya Milk Fermented by Various Bacteria Strains of Lactic Acid Fermentation. Zywnosc-Nauka Technol. Jakosc 2009, 67, 59–71. [Google Scholar]
- Czaczyk, K.; Marciniak, A.; Białas, W.; Mueller, A.; Myszka, K. The Effect of Environmental Factors Influencing Lipopeptide Biosurfactants Biosynthesis by Bacillus Spp. Zywnosc-Nauka Technol. Jakosc 2007, 50, 140–149. [Google Scholar]
- Huy, D.N.A.; Haom, P.A.; Hungm, P.V. Screening and identification of Bacillus sp. Isolated from traditional Vietnamese soybean-fermented products for high fibrinolytic enzyme production. Int. Food Res. J. 2016, 23, 326–331. [Google Scholar]
- Food and Environmental Hygiene Department FEHD. Available online: https://www.cfs.gov.hk/english/whatsnew/whatsnew_act/files/MBGL_RTE%20food_e.pdf (accessed on 2 September 2019).
- Wee, Y.J.; Yun, J.S.; Kim, D.; Ryu, H.W. Batch and repeated batch production of L(+)-lactic acid by Enterococcus faecalis RKY1 using wood hydrolyzate and corn steep liquor. J. Ind. Microbiol. Biotechnol. 2006, 33, 431–435. [Google Scholar] [CrossRef]
- Libudzisz, Z.; Kowal, K.; Żakowska, Z. Mikrobiologia Techniczna; Wydawnictwo Naukowe PWN: Warszawa, Polnd, 2007. [Google Scholar]
- Gajewska, J.; Błaszczyk, M.K. Probiotyczne bakterie fermentacji mlekowej (LAB). Postępy Mikrobiol. 2012, 51, 55–65. [Google Scholar]
- Makarova, K.; Slesarev, A.; Wolf, Y.; Sorokin, A.; Mirkin, B.; Koonin, E.; Pavlov, A.; Pavlova, N.; Karamychev, V.; Polouchine, N.; et al. Comparative genomics of the lactic acid bacteria. Proc. Natl. Acad. Sci. 2006, 103, 15611–15616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, S.; Cox, S.; Abu-Ghannam, N. Process optimization for the development of a functional beverage based on lactic acid fermentation of oats. Biochem. Eng. J. 2010, 52, 199–204. [Google Scholar] [CrossRef]
- Paseephol, T.; Sherkat, F. Probiotic stability of yoghurts containing Jerusalem artichoke inulins during refrigerated storage. J. Funct. Foods 2009, 1, 311–318. [Google Scholar] [CrossRef]
- Colakoglu, H.; Gursoy, O. Effect of lactic adjunct cultures on conjugated linoleic acid (CLA) concentration of yoghurt drink. J. Food Agric. Environ. 2011, 9, 60–64. [Google Scholar]
- Guo, Z.; Wang, J.; Yan, L.; Chen, W.; Liu, X.-M.; Zhang, H.-P. In vitro comparison of probiotic properties of Lactobacillus casei Zhang, a potential new probiotic, with selected probiotic strains. LWT 2009, 42, 1640–1646. [Google Scholar] [CrossRef]
- Fenderya, S.; Akbulut, N.; Akalın, A.S. Viability and activity of bifidobacteria in yoghurt containing fructooligosaccharide during refrigerated storage. Int. J. Food Sci. Technol. 2004, 39, 613–621. [Google Scholar] [CrossRef]
- Bonczar, G. The effects of certain factors on the properties of yoghurt made from ewe’s milk. Food Chem. 2002, 79, 85–91. [Google Scholar] [CrossRef]
- Bernat, N.; Cháfer, M.; Chiralt, A.; González-Martínez, C. Vegetable milks and their fermented derivative products. Int. J. Food Stud. 2014, 3, 93–124. [Google Scholar] [CrossRef]
- Bernat, N.; Cháfer, M.; González-Martínez, C.; Rodríguez-García, J.; Chiralt, A. Optimisation of oat milk formulation to obtain fermented derivatives by using probiotic Lactobacillus reuteri microorganisms. Food Sci. USA Technol. Int. 2015, 21, 145–157. [Google Scholar] [CrossRef]
- Blandino, A.; Al-Aseeri, M.; Pandiella, S.; Cantero, D.; Webb, C. Cereal-based fermented foods and beverages. Food Res. Int. 2003, 36, 527–543. [Google Scholar] [CrossRef]
- Gomes, A.M.; Malcata, F.; Gomes, A.M.; Malcata, F. Bifidobacterium spp. and Lactobacillus acidophilus: Biological, biochemical, technological and therapeutical properties relevant for use as probiotics. Trends Food Sci. Technol. 1999, 10, 139–157. [Google Scholar] [CrossRef]
- Rivera-Espinoza, Y.; Gallardo-Navarro, Y. Non-dairy probiotic products. Food Microbiol. 2010, 27, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Skrzyplonek, K.; Jasińska, M. Quality of Fermented Probiotic Beverages Made From Frozen Acid Whey and Milk During Refrigerated Storage. Zywnosc-Nauka Technol. Jakosc 2016, 104, 32–44. [Google Scholar] [CrossRef]
- At-Otaibi, M.M. Evaluation of some probiotic fermented milk products from Al-Ahsa markets, Saudi Arabia. J. Food Technol. 2009, 4, 1–8. [Google Scholar] [CrossRef]
- Bakirci, I.; Kavaz, A. An investigation of some properties of banana yogurts made with commercial ABT-2 starter culture during storage. Int. J. Dairy Technol. 2008, 61, 270–276. [Google Scholar] [CrossRef]
- Pescuma, M.; Hébert, E.M.; Mozzi, F.; De Valdez, G.F. Functional fermented whey-based beverage using lactic acid bacteria. Int. J. Food Microbiol. 2010, 141, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Olson, D.; Aryana, K. An excessively high Lactobacillus acidophilus inoculation level in yogurt lowers product quality during storage. LWT 2008, 41, 911–918. [Google Scholar] [CrossRef]
- Mousavi, Z.E.; Mousavi, S.M.; Razavi, S.H.; Emam-Djomeh, Z.; Kiani, H. Fermentation of pomegranate juice by probiotic lactic acid bacteria. World J. Microbiol. Biotechnol. 2011, 27, 123–128. [Google Scholar] [CrossRef]
- Pereira, A.L.F.; Maciel, T.C.; Rodrigues, S. Probiotic beverage from cashew apple juice fermented with Lactobacillus casei. Food Res. Int. 2011, 44, 1276–1283. [Google Scholar] [CrossRef]
- Salas, M.L.; Mounier, J.; Valence, F.; Coton, M.; Thierry, A.; Coton, E. Antifungal Microbial Agents for Food Biopreservation—A Review. Microorganisms 2017, 5, 37. [Google Scholar] [CrossRef]
- Wang, Y.-C.; Yu, R.-C.; Chou, C.-C. Growth and survival of bifidobacteria and lactic acid bacteria during the fermentation and storage of cultured soymilk drinks. Food Microbiol. 2002, 19, 501–508. [Google Scholar] [CrossRef]
- Frank, J.F.; Marth, E.M. Fermentations. In Fundamentals of Dairy Chemistry; Wong, N.P., Jenness, R., Keeney, M., Marth, E.H., Eds.; Springer: Boston, MA, USA, 1988; pp. 655–738. [Google Scholar] [CrossRef]
- Medina, L.M.; Jordano, R. Survival of constitutive microflora in commercially fermented milk containing Bifidobacteria during refrigerated storage. J. Food Prot. 1994, 56, 731–733. [Google Scholar] [CrossRef] [PubMed]
- Kosikowska, M.; Jakubczyk, E. Metody oznaczania bakterii probiotycznych w produktach mlecznych. Przeg. Mlecz. 2007, 57, 12–17. [Google Scholar]
- Pereira, A.L.F.; Almeida, F.D.L.; de Jesus, A.L.T.; da Costa, J.M.C.; Rodrigues, S. Storage Stability and Acceptance of Probiotic Beverage from Cashew Apple Juice. Food Bioprocess Technol. 2013, 6, 3155–3165. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Product | Protein | Lipids | Sugars |
|---|---|---|---|
| (%) | |||
| Desiccated coconut | 5.6 | 63.2 | 5.9 |
| Non-dehulled seeds of hemp | 25.2 | 36.1 | 5.4 |
| Non-Dairy Beverage | Protein | Lipids | Reducing Sugars | Active Acidity |
|---|---|---|---|---|
| (% ± SD) | (g glucose/L ± SD) | (pH ± SD) | ||
| Coconut milk | 3.23 ± 0.28 | 21.08 ± 0.41 | 34.53 ± 0.39 | 6.15 ± 0.15 |
| Hemp milk | 6.96 ± 0.19 | 18.02 ± 0.54 | 30.21 ± 0.33 | 6.81 ± 0.11 |
| Treatment | Storage Time | |||
|---|---|---|---|---|
| Day 1 | Day 7 | Day 14 | Day 21 | |
| Fermented | ||||
| CP | 5.61 ± 0.02 aF | 5.11 ± 0.02 bG | 4.93 ± 0.02 cB | 4.81 ± 0.02 dB |
| HP | 6.47 ± 0.01 aC | 6.08 ± 0.02 bA | 5.91 ± 0.02 cA | 5.78 ± 0.02 dA |
| C | 5.84 ± 0.01 aE | 4.85 ± 0.01 bH | 4.08 ± 0.01 cG | 3.58 ± 0.02 dG |
| H | 6.62 ± 0.02 aB | 5.49 ± 0.02 bD | 4.39 ± 0.02 cF | 3.41 ± 0.01 dH |
| Non-Fermented | ||||
| CP | 6.15 ± 0.02 aD | 5.31 ± 0.02 bE | 4.88 ± 0.02 cC | 4.21 ± 0.01 dC |
| HP | 6.81 ± 0.02 aA | 5.87 ± 0.02 bB | 4.65 ± 0.01 cD | 3.95 ± 0.02 dE |
| C | 6.15 ± 0.01 aD | 5.25 ± 0.02 bF | 4.53 ± 0.02 cE | 4.01 ± 0.02 dD |
| H | 6.81 ± 0.02 aA | 5.71 ± 0.01 bC | 4.35 ± 0.02 cF | 3.70 ± 0.02 dF |
| Treatment | Storage Time | |||
|---|---|---|---|---|
| Day 1 | Day 7 | Day 14 | Day 21 | |
| Fermented | ||||
| CP | 3.02 ± 0.21 aB | 2.95 ± 0.23 aB | 2.84 ± 0.15 aB | 2.67 ± 0.26 aB |
| HP | 6.68 ± 0.26 aA | 6.41 ± 0.28 aA | 6.33 ± 0.21 aA | 6.19 ± 0.24 aA |
| C | 3.02 ± 0.19 aB | 2.96 ± 0.17 aB | 2.87 ± 0.18 aB | 2.71 ± 0.18 aB |
| H | 6.68 ± 0.29 aA | 6.45 ± 0.25 aA | 6.34 ± 0.23 aA | 6.22 ± 0.23 aA |
| Non-Fermented | ||||
| CP | 3.23 ± 0.22 aB | 3.17 ± 0.19 aB | 3.10 ± 0.21 aB | 3.01 ± 0.24 aB |
| HP | 6.96 ± 0.27 aA | 6.91 ± 0.25 aA | 6.82 ± 0.23 aA | 6.68 ± 0.26 aA |
| C | 3.23 ± 0.18 aB | 3.15 ± 0.20 aB | 3.09 ± 0.20 aB | 2.98 ± 0.18 aB |
| H | 6.96 ± 0.27 aA | 6.89 ± 0.22 aA | 6.78 ± 0.21 aA | 6.63 ± 0.25 aA |
| Treatment | Storage Time | |||
|---|---|---|---|---|
| Day 1 | Day 7 | Day 14 | Day 21 | |
| Fermented | ||||
| CP | 21.06 ± 0.06 aA | 20.98 ± 0.06 abA | 20.87 ± 0.06 bcA | 20.75 ± 0.06 cA |
| HP | 18.01 ± 0.06 aB | 17.95 ± 0.06 abB | 17.81 ± 0.06 bcB | 17.66 ± 0.06 cB |
| C | 21.06 ± 0.07 aA | 20.96 ± 0.07 aA | 20.89 ± 0.07 abA | 20.73 ± 0.06 bA |
| H | 18.01 ± 0.06 aB | 17.89 ± 0.05 abB | 17.78 ± 0.06 bB | 17.62 ± 0.06 cB |
| Non-Fermented | ||||
| CP | 21.08 ± 0.02 aA | 21.01 ± 0.06 aA | 20.94 ± 0.06 aA | 20.84 ± 0.06 bA |
| HP | 18.02 ± 0.02 aB | 17.97 ± 0.06 aB | 17.92 ± 0.06 aB | 17.69 ± 0.06 bB |
| C | 21.08 ± 0.02 aA | 21.02 ± 0.06 aA | 20.93 ± 0.06 abA | 20.79 ± 0.06 bA |
| H | 18.02 ± 0.02 aB | 17.95 ± 0.06 aB | 17.89 ± 0.06 aB | 17.66 ± 0.06 bB |
| Treatment | Storage Time | |||
|---|---|---|---|---|
| Day 1 | Day 7 | Day 14 | Day 21 | |
| Fermented | ||||
| CP | 13.26 ± 0.14 aA | 11.46 ± 0.18 bA | 11.26 ± 0.17 bA | 9.41 ± 0.24 cA |
| HP | 10.92 ± 0.10 aC | 10.31 ± 0.25 aB | 8.28 ± 0.33 bB | 7.35 ± 0.26 cB |
| Non-Fermented | ||||
| CP | 11.72 ± 0.04 aB | 6.81 ± 0.13 bC | 5.42 ± 0.15 cC | 3.12 ± 0.13 dC |
| HP | 8.41 ± 0.18 aD | 6.35 ± 0.16 bC | 4.53 ± 0.08 cD | 3.54 ± 0.20 dC |
| Treatment | Storage Time | |||
|---|---|---|---|---|
| Day 1 | Day 7 | Day 14 | Day 21 | |
| Fermented | ||||
| C | 6.72 ± 0.17 aA | 6.23 ± 0.30 aA | 5.54 ± 0.24 bA | 3.82 ± 0.18 cB |
| H | 6.25 ± 0.13 aB | 5.69 ± 0.29 aA | 4.37 ± 0.20 bB | 3.93 ± 0.27 bB |
| Non-Fermented | ||||
| C | 4.39 ± 0.17 bC | 4.41 ± 0.15 bB | 4.79 ± 0.19 bB | 5.67 ± 0.18 aA |
| H | 3.81 ± 0.14 bD | 3.86 ± 0.08 bB | 5.76 ± 0.18 aA | 5.92 ± 0.16 aA |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szparaga, A.; Tabor, S.; Kocira, S.; Czerwińska, E.; Kuboń, M.; Płóciennik, B.; Findura, P. Survivability of Probiotic Bacteria in Model Systems of Non-Fermented and Fermented Coconut and Hemp Milks. Sustainability 2019, 11, 6093. https://doi.org/10.3390/su11216093
Szparaga A, Tabor S, Kocira S, Czerwińska E, Kuboń M, Płóciennik B, Findura P. Survivability of Probiotic Bacteria in Model Systems of Non-Fermented and Fermented Coconut and Hemp Milks. Sustainability. 2019; 11(21):6093. https://doi.org/10.3390/su11216093
Chicago/Turabian StyleSzparaga, Agnieszka, Sylwester Tabor, Sławomir Kocira, Ewa Czerwińska, Maciej Kuboń, Bartosz Płóciennik, and Pavol Findura. 2019. "Survivability of Probiotic Bacteria in Model Systems of Non-Fermented and Fermented Coconut and Hemp Milks" Sustainability 11, no. 21: 6093. https://doi.org/10.3390/su11216093
APA StyleSzparaga, A., Tabor, S., Kocira, S., Czerwińska, E., Kuboń, M., Płóciennik, B., & Findura, P. (2019). Survivability of Probiotic Bacteria in Model Systems of Non-Fermented and Fermented Coconut and Hemp Milks. Sustainability, 11(21), 6093. https://doi.org/10.3390/su11216093