Hi-sAFe: A 3D Agroforestry Model for Integrating Dynamic Tree–Crop Interactions

 , ,
, ,  , , , and
, , , and 
Abstract
:1. Introduction
- systems across pedoclimatic environments and management regimes;
- above- and belowground interactions for light, water, and nutrients;
- the multiple possible yields, including food, fiber, and fuel; and
- ecosystem services such as the capture of excess nutrients, soil erosion, and carbon sequestration [17].
- three-dimensional tree–crop interactions for light, water, and nitrogen;
- plastic aboveground and belowground tree architecture responsiveness to resource availability;
- the full lifetime of the system, from tree planting to harvest, on a daily time-step; and
- the principal agroforestry design and management strategies, such as branch pruning, tree thinning, root pruning, and the incorporation of an uncropped area around each tree or strip along the tree row.
2. Model Specification
2.1. Hi-sAFe Scene
2.2. STICS Crop Model in Hi-sAFe
2.3. Water and Nitrogen Cycles
2.4. sAFe-Tree Model
2.4.1. Tree Architecture
2.4.2. Phenology
2.4.3. Light Interception
2.4.4. Carbon Assimilation
2.4.5. Carbon Allocation
2.4.6. Growth and Allometry
2.4.7. Water Demand and Rain Interception
2.4.8. Nitrogen Demand and Allocation
2.4.9. Tree Management Interventions
2.5. Water Competition and Uptake
2.6. Nitrogen Competition and Uptake
- A weighted mean root diameter was used to account for differences in root diameter among plant species;
- To account for differences in current demand that may make the zero-sink assumption an overestimation for some plants, current demand per unit root length at the plant level was used as an additional weighing factor;
- To prevent plant uptake in a mixture from being more than it would be in a monoculture, a series of constraints were added to ensure that including roots of a nondemanding plant does not increase uptake by others.
3. Discussion
3.1. Model Approach
3.2. Model Applications
3.3. Model Limitations
3.4. Model Implementation and Distribution
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gold, M.A.; Hanover, J.W. Agroforestry systems for the temperate zone. Agrofor. Syst. 1987, 5, 109–121. [Google Scholar] [CrossRef]
- Wilson, M.H.; Lovell, S.T. Agroforestry—The next step in sustainable and resilient agriculture. Sustainability 2016, 8, 574. [Google Scholar] [CrossRef]
- Mosquera-Losada, M.R.; Moreno, G.; Pardini, A.; McAdam, J.H.; Papanastasis, V.; Burgess, P.J.; Lamersdorf, N.; Castro, M.; Liagre, F.; Rigueiro-Rodríguez, A. Past, present and future of agroforestry systems in Europe. In Agroforestry—The Future of Global Land Use; Nair, P., Garrity, D., Eds.; Advances in Agroforestry; Springer: Dordrecht, The Netherlands, 2012; Volume 9, pp. 285–312. [Google Scholar]
- Eichhorn, M.P.; Paris, P.; Herzog, F.; Incoll, L.D.; Liagre, F.; Mantzanas, K.; Mayus, M.; Moreno, G.; Papanastasis, V.P.; Pilbeam, D.J.; et al. Silvoarable systems in Europe—Past, present and future prospects. Agrofor. Syst. 2006, 67, 29–50. [Google Scholar] [CrossRef]
- Wolz, K.J.; DeLucia, E.H. Alley cropping: Global patterns of species composition and function. Agric. Ecosyst. Environ. 2018, 252, 61–68. [Google Scholar] [CrossRef]
- Graves, A.R.; Burgess, P.J.; Palma, J.H.N.; Herzog, F.; Moreno, G.; Bertomeu, M.; Dupraz, C.; Liagre, F.; Keesman, K.; van der Werf, W.; et al. Development and application of bio-economic modelling to compare silvoarable, arable, and forestry systems in three European countries. Ecol. Eng. 2007, 29, 434–449. [Google Scholar] [CrossRef]
- Osman, A.N.; Bayala, J. Performance of cowpea (Vigna unguiculata) and pearl millet (Pennisetum glaucum) intercropped under Parkia biglobosa in an agroforestry system in Burkina Faso. Afr. J. Agric. Res. 2011, 6, 882–891. [Google Scholar]
- Dubey, S.K.; Sharma, N.; Sharma, J.P.; Sharma, A.; Kishore, N. Assessing citrus (lemon) based intercropping in the irrigated areas of northern plains of Haryana. Indian J. Hortic. 2016, 73, 441–444. [Google Scholar] [CrossRef]
- Dyack, B.J.; Rollins, K.; Gordon, A.M. A model to calculate ex ante the threshold value of interaction effects necessary for proposed intercropping projects to be feasible to the landowner and desirable to society. Agrofor. Syst. 1999, 44, 197–214. [Google Scholar] [CrossRef]
- Keesman, K.J.; van der Werf, W.; Van Keulen, H. Production ecology of agroforestry systems: A minimal mechanistic model and analytical derivation of the land equivalent ratio. Math. Biosci. 2007, 209, 608–623. [Google Scholar] [CrossRef]
- Artru, S.; Garré, S.; Dupraz, C.; Hiel, M.-P.; Blitz-Frayret, C.; Lassois, L. Impact of spatio-temporal shade dynamics on wheat growth and yield, perspectives for temperate agroforestry. Eur. J. Agron. 2017, 82, 60–70. [Google Scholar] [CrossRef]
- Lovell, S.T.; Dupraz, C.; Gold, M.; Jose, S.; Revord, R.; Stanek, E.; Wolz, K.J. Temperate agroforestry research: Considering multifunctional woody polycultures and the design of long-term field trials. Agrofor. Syst. 2017, 263, 1–19. [Google Scholar] [CrossRef]
- Thevathasan, N.V.; Gordon, A.M. Ecology of tree intercropping systems in the north temperate region: Experiences from southern Ontario, Canada. Agrofor. Syst. 2004, 61, 257–268. [Google Scholar]
- Dupraz, C.; Auclair, D.; Barthélémy, D.; Caraglio, Y.; Sabatier, S.; Bariteau, M.; Kreiter, S.; Tixier Garcin, M.-S.; Maillet, J. Programme Intégré de Recherches en Agroforesterie à Restinclières (PIRAT); INRA: Montpellier, France, 2000; pp. 1–34. [Google Scholar]
- Udawatta, R.P.; Krstansky, J.J.; Henderson, G.S.; Garrett, H.E. Agroforestry practices, runoff, and nutrient loss: A paired watershed comparison. J. Environ. Qual. 2002, 31, 1214–1225. [Google Scholar] [CrossRef] [PubMed]
- Malézieux, E.; Crozat, Y.; Dupraz, C.; Laurans, M. Mixing plant species in cropping systems: Concepts, tools and models: A review. Agron. Sustain. Dev. 2009, 29, 43–62. [Google Scholar] [CrossRef]
- Luedeling, E.; Smethurst, P.J.; Baudron, F.; Bayala, J.; Huth, N.I.; van Noordwijk, M.; Ong, C.K.; Mulia, R.; Lusiana, B.; Muthuri, C.; et al. Field-scale modeling of tree–crop interactions: Challenges and development needs. Agric. Syst. 2016, 142, 51–69. [Google Scholar] [CrossRef]
- De Angelis, D.L.; Mooij, W.M. In praise of mechanistically rich models. In Models in Ecosystem Science; Canham, C.D., Cole, J., Lauenroth, W., Eds.; Princeton University Press: Princeton, NJ, USA, 2003; pp. 61–82. [Google Scholar]
- Oreske, N. The role of quantitative models in science. In Models in Ecosystem Science; Canham, C.D., Cole, J., Lauenroth, W., Eds.; Princeton University Press: Princeton, NJ, USA, 2003; pp. 13–31. [Google Scholar]
- Aumann, C.A. A methodology for developing simulation models of complex systems. Ecol. Model. 2007, 202, 385–396. [Google Scholar] [CrossRef]
- Boote, K.; Jones, J.; Hoogenboom, G. Simulation of crop growth CROPGRO model. In Agricultural System Modeling and Simulation; Peart, R., Cury, R., Eds.; CRC Press: New York, NY, USA, 1998; pp. 651–693. [Google Scholar]
- Brisson, N.; Mary, B.; Ripoche, D.; Jeuffroy, M.-H.; Ruget, F.; Nicoullaud, B.; Gate, P.; Devienne-Barret, F.; Antonioletti, R.; Durr, C.; et al. STICS: A generic model for the simulation of crops and their water and nitrogen balances. I. Theory and parameterization applied to wheat and corn. Agronomie 1998, 18, 311–346. [Google Scholar] [CrossRef]
- Zamora, D.S.; Jose, S.; Jones, J.W.; Cropper, W.P. Modeling cotton production response to shading in a pecan alleycropping system using CROPGRO. Agrofor. Syst. 2009, 76, 423–435. [Google Scholar] [CrossRef]
- Dufour, L.; Metay, A.; Talbot, G.; Dupraz, C. Assessing light competition for cereal production in temperate agroforestry systems using experimentation and crop modelling. J. Agron. Crop Sci. 2013, 199, 217–227. [Google Scholar] [CrossRef]
- Williams, J.R.; Jones, C.A.; Kiniry, J.R.; Spanel, D.A. The EPIC Crop Growth Model. Trans. ASAE 1989, 32, 497–511. [Google Scholar] [CrossRef]
- Easterling, W. Modelling the effect of shelterbelts on maize productivity under climate change: An application of the EPIC model. Agric. Ecosyst. Environ. 1997, 61, 163–176. [Google Scholar] [CrossRef]
- Qi, X.; Mize, C.W.; Batchelor, W.D.; Takle, E.S.; Litvina, I.V. SBELTS: A model of soybean production under tree shelter. Agrofor. Syst. 2001, 52, 53–61. [Google Scholar] [CrossRef]
- Mayus, M.; Van Keulen, H.; Stroosnijder, L. A model of tree–crop competition for windbreak systems in the Sahel: Description and evaluation. Agrofor. Syst. 1999, 43, 183–201. [Google Scholar] [CrossRef]
- Van der Werf, W.; Keesman, K.; Burgess, P.; Graves, A.; Pilbeam, D.; Incoll, L.D.; Metselaar, K.; Mayus, M.; Stappers, R.; van Keulen, H.; et al. Yield-SAFE: A parameter-sparse, process-based dynamic model for predicting resource capture, growth, and production in agroforestry systems. Ecol. Eng. 2007, 29, 419–433. [Google Scholar] [CrossRef]
- Graves, A.R.; Burgess, P.J.; Palma, J.; Keesman, K.J.; van der Werf, W.; Dupraz, C.; Van Keulen, H.; Herzog, F.; Mayus, M. Implementation and calibration of the parameter-sparse Yield-SAFE model to predict production and land equivalent ratio in mixed tree and crop systems under two contrasting production situations in Europe. Ecol. Model. 2010, 221, 1744–1756. [Google Scholar] [CrossRef]
- Huth, N.I.; Carberry, P.S.; Poulton, P.L.; Brennan, L.E.; Keating, B.A. A framework for simulating agroforestry options for the low rainfall areas of Australia using APSIM. Eur. J. Agron. 2002, 18, 171–185. [Google Scholar] [CrossRef]
- Mobbs, D.C.; Cannell, M.G.R.; Crout, N.M.J.; Lawson, G.J.; Friend, A.D.; Arah, J. Complementarity of light and water use in tropical agroforests I. Theoretical model outline, performance and sensitivity. For. Ecol. Manag. 1998, 102, 259–274. [Google Scholar] [CrossRef]
- Friend, A.D.; Stevens, A.K.; Knox, R.G.; Cannell, M.G.R. A process-based, terrestrial biosphere model of ecosystem dynamics (Hybrid v3.0). Ecol. Model. 1997, 95, 249–287. [Google Scholar] [CrossRef]
- Stephens, W.A.H.T.M. Modelling the benefits of soil water conservation using the PARCH model—A case study from a semi-arid region of Kenya. J. Arid Environ. 1999, 41, 335–344. [Google Scholar] [CrossRef]
- Cannell, M.G.R.; Mobbs, D.C.; Lawson, G.J. Complementarity of light and water use in tropical agroforests II. Modelled theoretical tree production and potential crop yield in arid to humid climates. For. Ecol. Manag. 1998, 102, 275–282. [Google Scholar] [CrossRef]
- Van Noordwijk, M.; Lusiana, B. WaNuLCAS, a model of water, nutrient and light capture in agroforestry systems. Agrofor. Syst. 1999, 43, 217–242. [Google Scholar] [CrossRef]
- Walker, A.P.; Mutuo, P.K.; van Noordwijk, M.; Albrecht, A.; Cadisch, G. Modelling of planted legume fallows in Western Kenya using WaNuLCAS. (I) Model calibration and validation. Agrofor. Syst. 2007, 70, 197–209. [Google Scholar] [CrossRef]
- Martin, F.S.; van Noordwijk, M. Trade-offs analysis for possible timber-based agroforestry scenarios using native trees in the Philippines. Agrofor. Syst. 2009, 76, 555–567. [Google Scholar] [CrossRef]
- Pansak, W.; Hilger, T.; Lusiana, B.; Kongkaew, T.; Marohn, C.; Cadisch, G. Assessing soil conservation strategies for upland cropping in Northeast Thailand with the WaNuLCAS model. Agrofor. Syst. 2010, 79, 123–144. [Google Scholar] [CrossRef]
- Cahyo, A.N.; Babel, M.S.; Datta, A.; Prasad, K.C.; Clemente, R. Evaluation of land and water management options to enhance productivity of rubber plantation using WaNuLCas model. Agrivita J. Agr. Sci. 2016, 38, 93–103. [Google Scholar] [CrossRef]
- Dupraz, C.; Burgess, P.; Gavaland, A.; Graves, A.; Herzog, F.; Incoll, L.D.; Jackson, N.; Keesman, K.; Lawson, G.; Lecomte, I.; et al. Synthesis of the Silvoarable Agroforestry For Europe (SAFE) Project; INRA-UMR System: Montpellier, France, 2005; pp. 1–254. [Google Scholar]
- Talbot, G. L’intégration Spatiale et Temporelle du Partage des Ressources dans un Système Agroforestier Noyers-Céréales: Une Clef pour en Comprendre la Productivité? Ecosystems; Université Montpellier II—Sciences et Techniques du Languedoc: Montpellier, France, 2011; pp. 1–298. [Google Scholar]
- Cannell, M.G.R.; van Noordwijk, M.; Ong, C.K. The central agroforestry hypothesis: The trees must acquire resources that the crop would not otherwise acquire. Agrofor. Syst. 1996, 34, 27–31. [Google Scholar] [CrossRef]
- Jose, S.; Gillespie, A.R.; Pallardy, S.G. Interspecific interactions in temperate agroforestry. Agrofor. Syst. 2004, 61–62, 237–255. [Google Scholar]
- Brisson, N.; Ruget, F.O.; Gate, P.; Lorgeou, J.; Nicoullaud, B.; Tayot, X.; Plenet, D.; Jeuffroy, M.-H.L.N.; Bouthier, A.; Ripoche, D.; et al. STICS: A generic model for simulating crops and their water and nitrogen balances. II. Model validation for wheat and maize. Agronomie 2002, 22, 69–92. [Google Scholar] [CrossRef]
- De Coligny, F.; Ancelin, P.; Cornu, G.; Courbaud, B.; Dreyfus, P.; Goreaud, F.; Gourlet-Fleury, S.; Meredieu, C.; Saint-André, L. CAPSIS: Computer-Aided Projection for Strategies in Silviculture: Advantages of a Shared Forest-Modelling Platform; CABI: Sesimbra, Portugal, 2002; pp. 1–4. [Google Scholar]
- Mulia, R.; Dupraz, C.; van Noordwijk, M. Reconciling root plasticity and architectural ground rules in tree root growth models with voxel automata. Plant Soil 2010, 337, 77–92. [Google Scholar] [CrossRef]
- Courbaud, B.; de Coligny, F.; Cordonnier, T. Simulating radiation distribution in a heterogeneous Norway spruce forest on a slope. Agric. For. Meteorol. 2003, 116, 1–18. [Google Scholar] [CrossRef]
- Dauzat, J. Radiative transfer simulation on computer models of Elaeis guineensis. Oléagineux 1994, 49, 8–90. [Google Scholar]
- Brunner, A. A light model for spatially explicit forest stand models. For. Ecol. Manag. 1998, 107, 19–46. [Google Scholar] [CrossRef]
- Canham, C.D.; Coates, K.D.; Bartemucci, P.; Quaglia, S. Measurement and modeling of spatially explicit variation in light transmission through interior cedar-hemlock forests of British Columbia. Can. J. For. Res. 1999, 29, 1775–1783. [Google Scholar] [CrossRef]
- Moon, P.; Spencer, D. Illumination from a non-uniform sky. Illum. Eng. 1942, 37, 707–726. [Google Scholar]
- Martens, S.N.; Breshears, D.D.; Meyer, C.W. Spatial distributions of understory light along the grassland/forest continuum: Effects of cover, height, and spatial pattern of tree canopies. Ecol. Model. 2000, 126, 79–93. [Google Scholar] [CrossRef]
- Monteith, J.L. Climate and the efficiency of crop production in Britain. Philos. Trans. R. Soc. Lond. B 1977, 281, 277–294. [Google Scholar] [CrossRef]
- Lacointe, A.; Kajji, A.; Daudet, F.-A.; Archer, P.; Frossard, J.-S. Seasonal Variation of Photosynthetic Carbon Flow Rate into Young Walnut and its Partitioning among the Plant Organs and Functions. J. Plant Physiol. 1995, 146, 222–230. [Google Scholar] [CrossRef]
- Poorter, H.; Nagel, O. The role of biomass allocation in the growth response of plants to different levels of light, CO2, nutrients and water: A quantitative review. Aust. J. Plant Physiol. 2000, 27, 595–607. [Google Scholar]
- De Willigen, P.; Nielsen, N.E.; Claassen, N.; Castrignanò, A.M. Modeling water and nutrient uptake. In Root Methods, a Handbook; Smit, A.L., Bengough, A.G., Engels, C., van Noordwijk, M., Pellerin, S., van de Geijn, S.C., Eds.; Springer: Berlin/Heidelberg, Germany, 2000; pp. 509–544. [Google Scholar]
- Van Noordwijk, M.; de Willigen, P.; Lusiana, B.; Mulia, R.; Ozier-Lafontaine, H.; Radersma, S.; Dupraz, C. The Hi-sAFe Process-Based Algorithm for Water Uptake by Roots of Trees and Crops Rooted in the Same Volume of Soil; INRA: Montpellier, France, 2018; Available online: https://www1.montpellier.inra.fr/wp-inra/hi-safe/en/ (accessed on 1 November 2018).
- De Willigen, P.; van Noordwijk, M. Uptake potential of non-regularly distributed roots. J. Plant Nutr. 1987, 10, 1273–1280. [Google Scholar] [CrossRef]
- De Willigen, P.; van Noordwijk, M. Diffusion and mass flow to a root with constant nutrient demand or behaving as a zero-sink. Soil Sci. 1994, 157, 162–170. [Google Scholar] [CrossRef]
- Schachtman, D.P.; Goodger, J.Q.D. Chemical root to shoot signaling under drought. Trends Plant Sci. 2008, 13, 281–287. [Google Scholar] [CrossRef]
- Tardieu, F. Too many partners in root-shoot signals. Does hydraulics qualify as the only signal that feeds back over time for reliable stomatal control? New Phytol. 2016, 212, 802–804. [Google Scholar] [CrossRef]
- Lusiana, B.; van Noordwijk, M.; Dupraz, C. The Hi-sAFe Algorithm for Sharing Nutrient Uptake between Trees and Crops Rooted in the Same Volume of Soil; INRA: Montpellier, France, 2018; Available online: https://www1.montpellier.inra.fr/wp-inra/hi-safe/en/ (accessed on 1 November 2018).
- Wolz, K.J.; Lovell, S.T.; Branham, B.E.; Eddy, W.C.; Keeley, K.; Revord, R.S.; Wander, M.M.; Yang, W.H.; DeLucia, E.H. Frontiers in alley cropping: Transformative solutions for temperate agriculture. Glob. Chang. Biol. 2018, 24, 883–894. [Google Scholar] [CrossRef]
- Andrianarisoa, K.S.; Dufour, L.; Bienaimé, S.; Zeller, B.; Dupraz, C. The introduction of hybrid walnut trees (Juglans nigra × regia cv. NG23) into cropland reduces soil mineral N content in autumn in southern France. Agrofor. Syst. 2016, 90, 193–205. [Google Scholar] [CrossRef]
- Buttoud, G. Advancing Agroforestry on the Policy Agenda; FAO: Rome, Italy, 2013; pp. 1–37. [Google Scholar]
- Schoeneberger, M.; Bentrup, G.; de Gooijer, H.; Soolanayakanahally, R.; Sauer, T.; Brandle, J.; Zhou, X.; Current, D. Branching out: Agroforestry as a climate change mitigation and adaptation tool for agriculture. J. Soil Water Conserv. 2012, 67, 128A–136A. [Google Scholar] [CrossRef]
- Anderson, S.H.; Udawatta, R.P.; Seobi, T.; Garrett, H.E. Soil water content and infiltration in agroforestry buffer strips. Agrofor. Syst. 2009, 75, 5–16. [Google Scholar] [CrossRef]
- Böhm, C.; Kanzler, M.; Freese, D. Wind speed reductions as influenced by woody hedgerows grown for biomass in short rotation alley cropping systems in Germany. Agrofor. Syst. 2014, 88, 579–591. [Google Scholar] [CrossRef]
- Nasielski, J.; Furze, J.R.; Tan, J.; Bargaz, A.; Thevathasan, N.V.; Isaac, M.E. Agroforestry promotes soybean yield stability and N2-fixation under water stress. Agron. Sustain. Dev. 2015, 35, 1541–1549. [Google Scholar] [CrossRef]
- Van der Putten, W.H.; Bardgett, R.D.; Bever, J.D.; Bezemer, T.M.; Casper, B.B.; Fukami, T.; Kardol, P.; Klironomos, J.N.; Kulmatiski, A.; Schweitzer, J.A.; et al. Plant-soil feedbacks: The past, the present and future challenges. J. Ecol. 2013, 101, 265–276. [Google Scholar] [CrossRef]
- Tilman, D. Diversity and productivity in a long-term grassland experiment. Science 2001, 294, 843–845. [Google Scholar] [CrossRef] [PubMed]
- Vandermeer, J. The Ecology of Intercropping; Cambridge University Press: Cambridge, UK, 1989. [Google Scholar]
- Li, L.; Tilman, D.; Lambers, H.; Zhang, F.S. Plant diversity and overyielding: Insights from belowground facilitation of intercropping in agriculture. New Phytol. 2014, 203, 63–69. [Google Scholar] [CrossRef]
- Brooker, R.W.; Maestre, F.T.; Callaway, R.M.; Lortie, C.L.; Cavieres, L.A.; Kunstler, G.; Liancourt, P.; Tielbörger, K.; Travis, J.M.J.; Anthelme, F.; et al. Facilitation in plant communities: The past, the present, and the future. J. Ecol. 2008, 96, 18–34. [Google Scholar] [CrossRef]
- Perry, D.; Margolis, H.; Choquette, C.; Molina, R. Ectomycorrhizal mediation of competition between coniferous tree species. New Phytol. 1989, 112, 501–511. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, H.Y.H.; Reich, P.B. Forest productivity increases with evenness, species richness and trait variation: A global meta-analysis. J. Ecol. 2012, 100, 742–749. [Google Scholar] [CrossRef]
- Collet, C.; Ningre, F.; Barbeito, I.; Arnaud, A.; Piboule, A. Response of tree growth and species coexistence to density and species evenness in a young forest plantation with two competing species. Ann. Bot. 2014, 113, 711–719. [Google Scholar] [CrossRef]
- Sapijanskas, J.; Paquette, A.; Potvin, C.; Kunert, N. Tropical tree diversity enhances light capture through crown plasticity and spatial and temporal niche differences. Ecology 2014, 95, 2479–2492. [Google Scholar] [CrossRef]
- Dupraz, C. Adequate design of control treatments in long term agroforestry experiments with multiple objectives. Agrofor. Syst. 1999, 43, 35–48. [Google Scholar] [CrossRef]
- Lott, J.E.; Ong, C.K.; Black, C.R. Understorey microclimate and crop performance in a Grevillea robusta-based agroforestry system in semi-arid Kenya. Agric. For. Meteorol. 2009, 149, 1140–1151. [Google Scholar] [CrossRef]
- Gea-Izquierdo, G.; Montero, G.; Cañellas, I. Changes in limiting resources determine spatio-temporal variability in tree–grass interactions. Agrofor. Syst. 2009, 76, 375–387. [Google Scholar] [CrossRef]
- Mead, R.; Willey, R.W. The concept of a “land equivalent ratio” and advantages in yields from intercropping. Exp. Agric. 1980, 16, 217–228. [Google Scholar] [CrossRef]
- Grimm, V. Mathematical models and understanding in ecology. Ecol. Model. 1994, 75–76, 641–651. [Google Scholar] [CrossRef]
- Van Nes, E.H.; Scheffer, M. A strategy to improve the contribution of complex simulation models to ecological theory. Ecol. Model. 2005, 185, 153–164. [Google Scholar] [CrossRef]
- Rykiel, E.J. Testing ecological models: The meaning of validation. Ecol. Model. 1996, 90, 229–244. [Google Scholar] [CrossRef]
- Carpenter, S.R. The need for fast-and-frugal models. In Models in Ecosystem Science; Canham, C.D., Cole, J., Lauenroth, W., Eds.; Princeton University Press: Princeton, NJ, USA, 2003; pp. 455–460. [Google Scholar]
- Brauer, D.; Ares, A.; Reid, W.; Thomas, A.; Slusher, J.P. Nut-yield Variations and Yield–diameter Relationships in Open-canopy Black Walnut Trees in Southern USA. Agrofor. Syst. 2006, 67, 63–72. [Google Scholar] [CrossRef]
- Villalobos, F.J.; Testi, L.; Hidalgo, J.; Pastor, M.; Orgaz, F. Modelling potential growth and yield of olive (Olea europaea L.) canopies. Eur. J. Agron. 2006, 24, 296–303. [Google Scholar] [CrossRef]
- Petriccione, M.; Ciarmiello, L.F.; Boccacci, P.; De Luca, A.; Piccirillo, P. Evaluation of “Tonda di Giffoni” hazelnut (Corylus avellana L.) clones. Sci. Hortic. 2010, 124, 153–158. [Google Scholar] [CrossRef]
- Rosenstock, T.S.; Rosa, U.A.; Plant, R.E.; Brown, P.H. A reevaluation of alternate bearing in pistachio. Sci. Hortic. 2010, 124, 149–152. [Google Scholar] [CrossRef]
- Farquhar, G.; Caemmerer, S. A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta 1980, 149, 78–90. [Google Scholar] [CrossRef] [PubMed]
- Warren, J.M.; Norby, R.J.; Wullschleger, S.D. Elevated CO2 enhances leaf senescence during extreme drought in a temperate forest. Tree Physiol. 2011, 31, 117–130. [Google Scholar] [CrossRef]
- Augspurger, C.K. Reconstructing patterns of temperature, phenology, and frost damage over 124 years: Spring damage risk is increasing. Ecology 2013, 94, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Augspurger, C.K.; Bartlett, E.A. Differences in leaf phenology between juvenile and adult trees in a temperate deciduous forest. Tree Physiol. 2003, 23, 517–525. [Google Scholar] [CrossRef]
- Morin, X.; Lechowicz, M.J.; Augspurger, C.; O’Keefe, J.; Viner, D.; Chuine, I. Leaf phenology in 22 North American tree species during the 21st century. Glob. Chang. Biol. 2009, 15, 961–975. [Google Scholar] [CrossRef]
- Kramer, K.; Leinonen, I.; Loustau, D. The importance of phenology for the evaluation of impact of climate change on growth of boreal, temperate and Mediterranean forests ecosystems: An overview. Int. J. Biometeorol. 2000, 44, 67–75. [Google Scholar] [CrossRef]
- Germon, A.; Cardinael, R.; Prieto, I.; Mao, Z.; Kim, J.; Stokes, A.; Dupraz, C.; Laclau, J.-P.; Jourdan, C. Unexpected phenology and lifespan of shallow and deep fine roots of walnut trees grown in a silvoarable Mediterranean agroforestry system. Plant Soil 2015, 401, 409–426. [Google Scholar] [CrossRef]
- Valle, B.; Simonneau, T.; Sourd, F.; Pechier, P.; Hamard, P.; Frisson, T.; Ryckewaert, M.; Christophe, A. Increasing the total productivity of a land by combining mobile photovoltaic panels and food crops. Appl. Energy 2017, 206, 1495–1507. [Google Scholar] [CrossRef]
- Shuttleworth, W.J.; Wallace, J.S. Evaporation from sparse crops-an energy combination theory. Q. J. R. Meteorol. Soc. 1985, 111, 839–855. [Google Scholar] [CrossRef]
- sBlack, C.K.; Masters, M.D.; LeBauer, D.S.; Anderson-Teixeira, K.J.; DeLucia, E.H. Root volume distribution of maturing perennial grasses revealed by correcting for minirhizotron surface effects. Plant Soil 2017, 419, 391–404. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
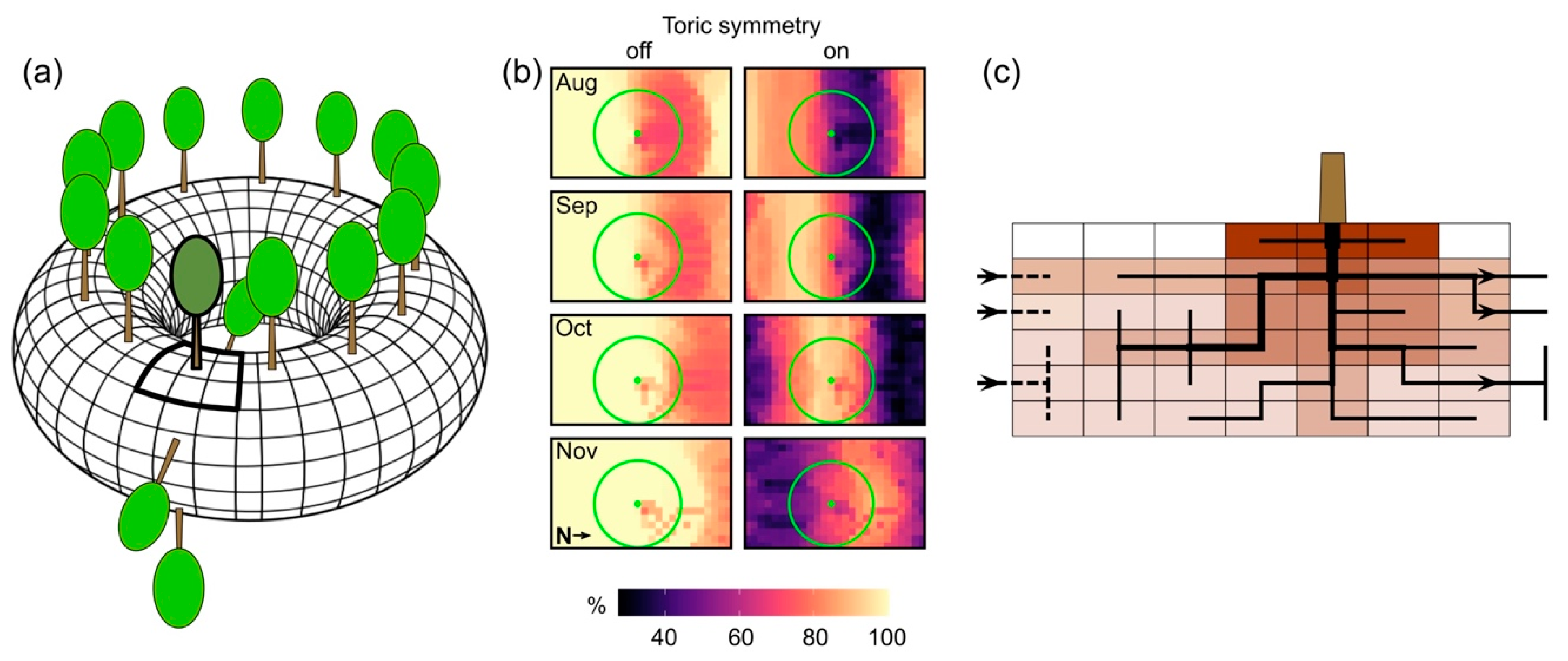{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter Definition | Units | Allowed Values |
|---|---|---|
| Latitude | degrees | (−90, 90] |
| Cell width | m | (0, -) |
| Orientation of north | degrees clockwise relative to +y axis | [0, 360) |
| Plot length (y) | m | [cell width, -) |
| Plot width (x) | m | [cell width, -) |
| Magnitude of the slope of the soil surface | degrees downward from horizontal | [0, 90) |
| Aspect of the slope of the soil surface | degrees clockwise from north | [0, 360) |
| Maximum thickness of a voxel | m | [0.01, -) |
| Distance from the tree to the alley crop edge | m | [0, plot width) |
| PBC in the +x direction | - | on/off |
| PBC in the -x direction | - | on/off |
| PBC in the +y direction | - | on/off |
| PBC in the -y direction | - | on/off |
| Parameter | Definition | Units | Allowed Values |
|---|---|---|---|
| Phenology | |||
| D0 | Date to start accumulation of temperature for budburst | Julian day | [1, 365] |
| ST | Threshold of accumulated degree-days to trigger budburst | °C-days | [0, -) |
| DLE | Duration of leaf expansion | days | [1, 365] |
| DV | Bud burst to leaf fall duration | days | [1, 365] |
| TLF | Threshold for frost mortality of leaves | °C | (-, -) |
| DLF | Duration of leaf fall | days | [1, 365] |
| Light interception | |||
| μ | Correction parameter to account for leaf clumping. A clumping coefficient below 1, equal to 1, or above 1 indicates a clumped, random, or regular distribution of leaves inside the crown, respectively. | - | [0, -) |
| σPAR | Leaf light absorption coefficient for photosynthetically active radiation | - | [0, 1] |
| σNIR | Leaf light absorption coefficient for near infrared radiation | - | [0, 1] |
| WAD | Wood area density for light interception by tree branches | m2 m−3 | [0, -) |
| Carbon assimilation | |||
| LUEmax | Maximum potential light-use efficiency | g C MJ−1 | (0, -) |
| ωLUE | Sensitivity of LUE to water stress | - | [0, -) |
| ηLUE | Sensitivity of LUE to N stress | - | [0, -) |
| τmax | Leaf age at which maximum LUE occurs | days | [1, 365] |
| τleaves | Leaf senescence time constant | days−2 | (0, -) |
| Carbon allocation | |||
| aNSC | Threshold imbalance above which remobilization of CNSC is triggered | - | (0, 1] |
| bNSC | Limits maximum daily ΔCNSC as a fraction of CNSC | - | (0, 1] |
| cNSC | Limits maximum daily ΔCNSC as a fraction of Cleaves | - | (0, 1] |
| αNSC* | Target CNSC as a fraction of the tree woody C pool | - | (0, 1] |
| LFR*0 | Initial LFR* at tree planting | kg kg−1 | (0, 1) |
| ΔLFR*max | Maximum daily change in LFR* | kg kg−1 | [0, -) |
| δLFR | Target daily upward drift in LFR* | kg kg−1 | [0, -) |
| εLFR | Sensitivity of LFR* to N satisfaction when there is no stress | day−1 | [0, -) |
| ωLFR | Sensitivity of LFR* to water stress | - | [0, -) |
| ηLFR | Sensitivity of LFR* to N stress | - | [0, -) |
| LFR*min | Minimum allowed LFR* | kg kg−1 | (0, 1) |
| LFR*max | Maximum allowed LFR* | kg kg−1 | (0, 1) |
| Growth and senescence | |||
| SLM | Leaf dry mass per unit leaf area | kg m−2 | (0, -) |
| θleaves | Leaf C content | g C g−1 dry biomass | (0, 1) |
| θwood | C content of all compartments except leaves | g C g−1 dry biomass | (0, 1) |
| ρwood | Wood density of branches, stems, stump, and coarse roots | kg m−3 | (0, -) |
| γ | Ratio of coarse root cross-sectional area to fine root length | m2 m−1 | (0, -) |
| SRL | Fine root length per unit dry mass | m g−1 of dry matter | (0, -) |
| τfr | Mean lifespan of fine roots not in anoxic voxel | days | (0, -) |
| τfr,anoxia | Mean lifespan of fine roots in anoxic voxel | days | (0, -) |
| τcr | Number of days of anoxia to kill coarse roots | days | (0, -) |
| Root growth algorithm [47] | |||
| α | Threshold for root colonization | m of root m−3 voxel | (0, -) |
| β | The proportion of C allocated to fine roots that is allocated to colonization | - | (0, 1) |
| λ | Fraction of root colonization to horizontal voxels | - | [0, 1] |
| η | Fraction of vertical root colonization to lower voxel | - | [0, 1] |
| ϕ | Fine root proliferation weighting factor for water uptake efficiency | - | [0, -) |
| υ | Fine root proliferation weighting factor for N uptake efficiency | - | [0, -) |
| ρ | Fine root proliferation weighting factor for voxel–stem base topological distance | - | [0, -) |
| Water demand and rain interception | |||
| Γ | Scaling coefficient for transpiration | - | (0, -) |
| ω | Wettability of leaves | mm lai−1 | [0, -) |
| SFmax | Maximum fraction of intercepted rain routed to stem flow | - | [0, 1] |
| cSF | Scaling coefficient for stem flow | - | [0, -) |
| Nitrogen demand | |||
| ηbranches | Functional optimum N/C ratio in branches | kg N kg C−1 | (0, -) |
| ηcr | Functional optimum N/C ratio in coarse roots | kg N kg C−1 | (0, -) |
| ηfr | Functional optimum N/C ratio in fine roots | kg N kg C−1 | (0, -) |
| ηleaves | Functional optimum N/C ratio in foliage | kg N kg C−1 | (0, -) |
| ηstem | Functional optimum N/C ratio in stem | kg N kg C−1 | (0, -) |
| ηstump | Functional optimum N/C ratio in stump | kg N kg C−1 | (0, -) |
| αN | Coefficient applied to optimum N content to define target N content | - | [1, -) |
| βN | Coefficient defining “luxury” N content | ||
| κleaves | Fraction of N content recovered from leaves during senescence | - | [0, 1] |
| κfr | Fraction of N content recovered from fine roots during senescence | - | [0, 1] |
| (A1) | |
| (A2) | |
| (A3) | |
| (A4) | |
| (A5) | |
| (A6) | |
| (A7) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dupraz, C.; Wolz, K.J.; Lecomte, I.; Talbot, G.; Vincent, G.; Mulia, R.; Bussière, F.; Ozier-Lafontaine, H.; Andrianarisoa, S.; Jackson, N.; et al. Hi-sAFe: A 3D Agroforestry Model for Integrating Dynamic Tree–Crop Interactions. Sustainability 2019, 11, 2293. https://doi.org/10.3390/su11082293
Dupraz C, Wolz KJ, Lecomte I, Talbot G, Vincent G, Mulia R, Bussière F, Ozier-Lafontaine H, Andrianarisoa S, Jackson N, et al. Hi-sAFe: A 3D Agroforestry Model for Integrating Dynamic Tree–Crop Interactions. Sustainability. 2019; 11(8):2293. https://doi.org/10.3390/su11082293
Chicago/Turabian StyleDupraz, Christian, Kevin J. Wolz, Isabelle Lecomte, Grégoire Talbot, Grégoire Vincent, Rachmat Mulia, François Bussière, Harry Ozier-Lafontaine, Sitraka Andrianarisoa, Nick Jackson, and et al. 2019. "Hi-sAFe: A 3D Agroforestry Model for Integrating Dynamic Tree–Crop Interactions" Sustainability 11, no. 8: 2293. https://doi.org/10.3390/su11082293
APA StyleDupraz, C., Wolz, K. J., Lecomte, I., Talbot, G., Vincent, G., Mulia, R., Bussière, F., Ozier-Lafontaine, H., Andrianarisoa, S., Jackson, N., Lawson, G., Dones, N., Sinoquet, H., Lusiana, B., Harja, D., Domenicano, S., Reyes, F., Gosme, M., & Van Noordwijk, M. (2019). Hi-sAFe: A 3D Agroforestry Model for Integrating Dynamic Tree–Crop Interactions. Sustainability, 11(8), 2293. https://doi.org/10.3390/su11082293